
Serovars, Virulence and Antimicrobial Resistance Genes of Non-Typhoidal Salmonella Strains from Dairy Systems in Mexico

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Prevalence of Salmonella in the Different Samples
2.2. Antimicrobial Susceptibility Testing
2.3. Serological Identification of Salmonella enterica O Serogroups
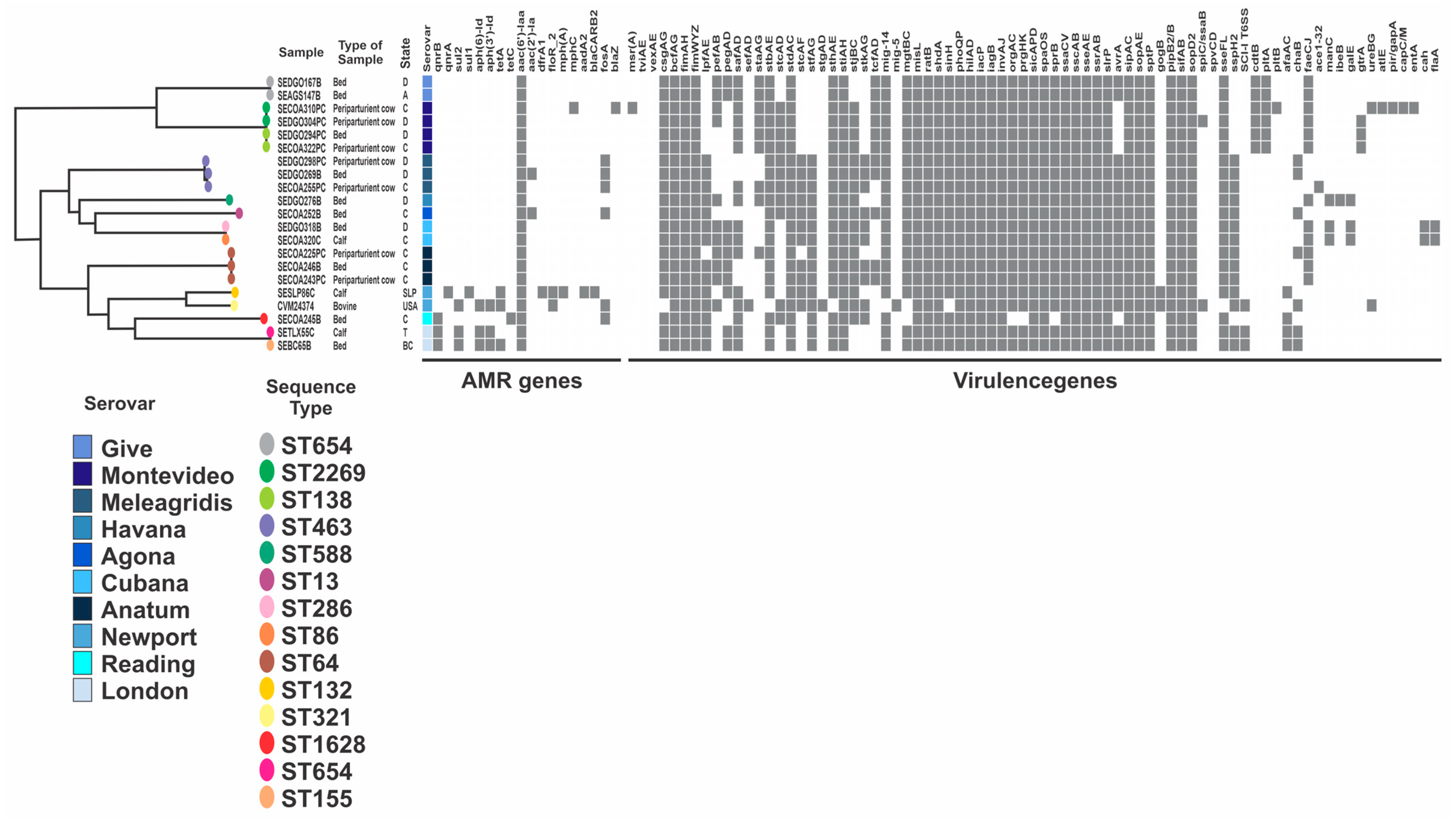
2.4. Genomic Characteristics
2.4.1. In Silico Serovar Identification
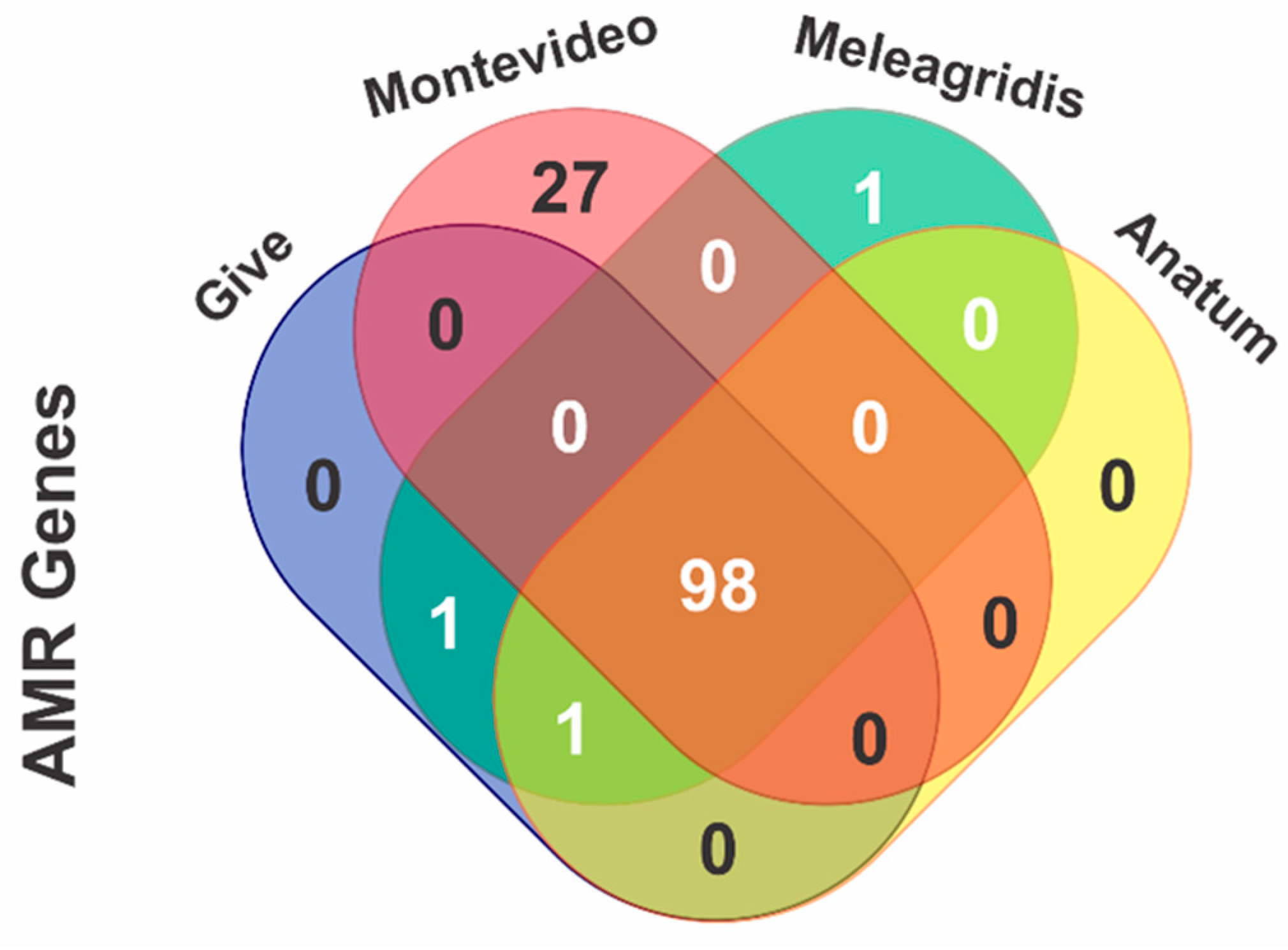
2.4.2. In Silico Detection of Antimicrobial Resistance Genes
2.4.3. Plasmid Profiling
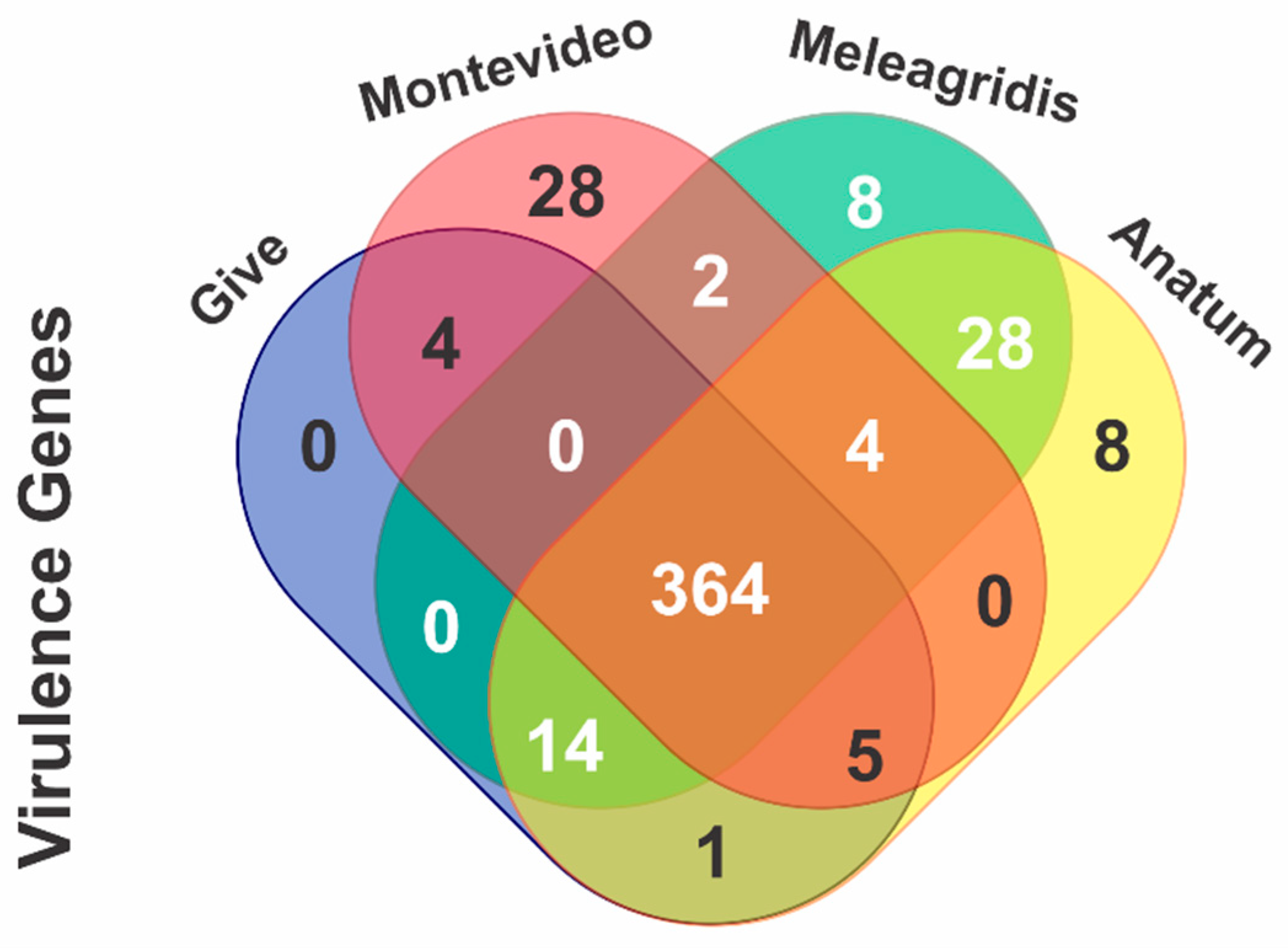
2.4.4. In Silico Identification of Virulence Genes
2.4.5. In Silico Identification of Salmonella Pathogenicity Island
2.5. Phylogenetic Diversity
3. Discussion
4. Materials and Methods
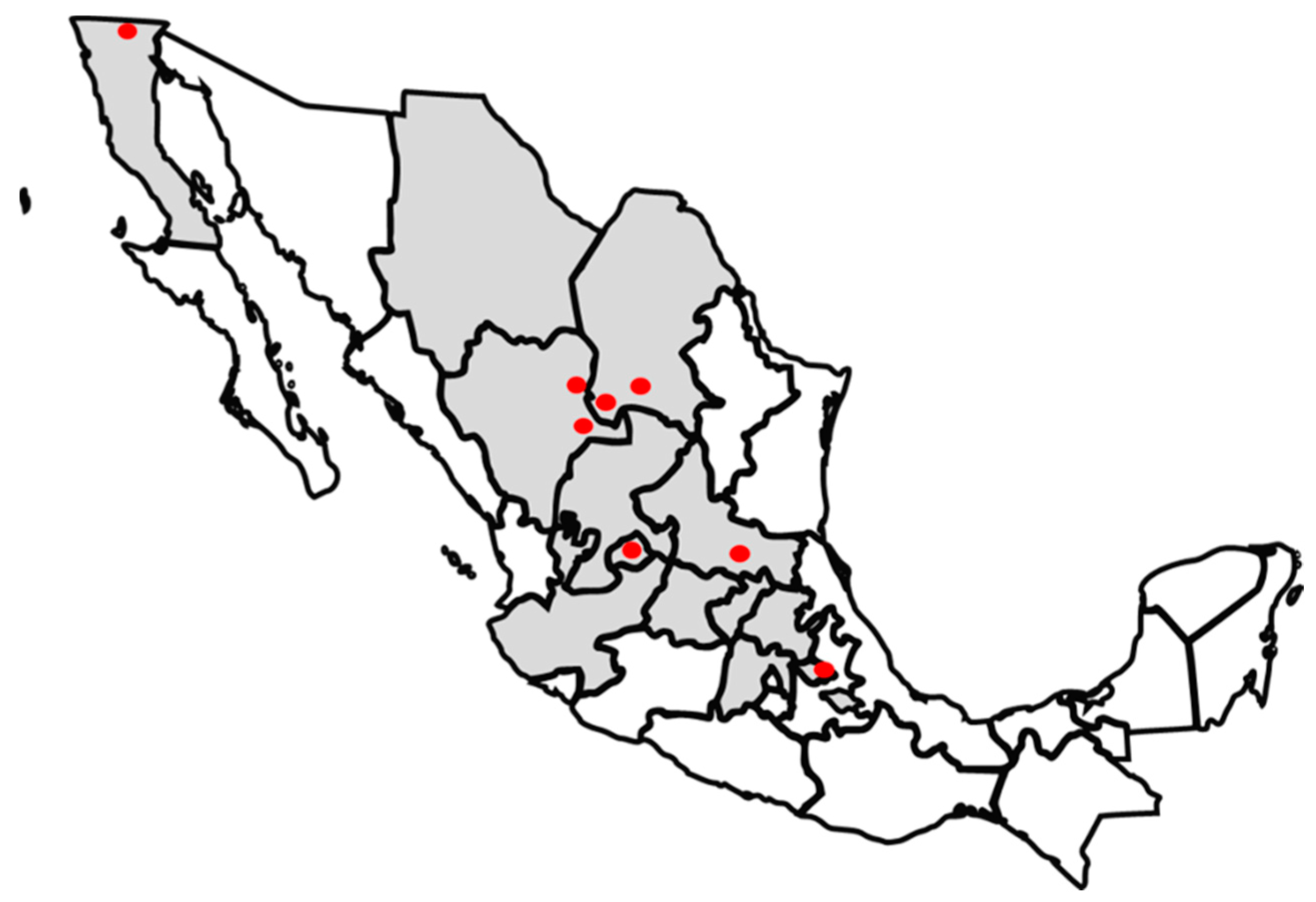
4.1. Sample Collection
4.2. Salmonella Isolation from Bovine Fecal Samples and Environment
4.3. DNA Extraction
4.4. Detection of ipaB Gene for Salmonella Genus
4.5. Antimicrobial Susceptibility Testing
4.6. Serological Identification of Salmonella enterica O Serogroups
4.7. Genome Sequencing and Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grimont, P.; Weill, F. Antigenic Formulae of the Salmonella serovars, 9th ed.; WHO Collaborating Centre for Reference and Research on Salmonella; Institut Pateur: Paris, France, 2007. [Google Scholar]
- Thompson, C.P.; Doak, A.; Amirani, N.; Schroeder, E.; Wright, J.; Kariyawasam, S.; Lamendella, R.; Shariat, N. High-resolution identification of multiple Salmonella serovars in a single sample by using CRISPRSeroSeq. Appl. Environ. Microbiol. 2018, 84, e01859-18. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Juniora, C.A. Worldwide epidemiology of Salmonella serovars in animal-based foods: A meta-analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed]
- Cao, T.; Liu, P.; Li, Y.; Cui, M.; Zhang, C.; Wang, Y.; Shen, J.; Ke, Y.; Wang, Y.; Shen, J.; et al. Prevalence of Salmonella and Antimicrobial Resistance in Isolates from Food Animals—Six PLADs, China, 2019. China CDC Wkly. 2021, 3, 514–517. [Google Scholar] [CrossRef]
- Cummings, K.J.; Warnick, L.D.; Elton, M.; Grö, Y.T.; Mcdonough, P.L.; Siler, J.D. The Effect of Clinical Outbreaks of Salmonellosis on the Prevalence of Fecal Salmonella Shedding Among Dairy Cattle in New York. Foodborne Pathog. Dis. 2010, 7, 815–823. [Google Scholar] [CrossRef]
- Waldner, L.L.; MacKenzie, K.D.; Köster, W.; White, A.P. From exit to entry: Long-term survival and transmission of Salmonella. Pathogens 2012, 1, 128–155. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services; CDC: Atlanta, GA, USA, 2019.
- World Health Organization. Antimicrobial Resistance Global Report on Surveillance; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Koutsoumanis, K.; Allende, A.; Álvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Herman, L.; Peixe, L. Role played by the environment in the emergence and spread of antimicrobial resistance (AMR) through the food chain. EFSA J. 2021, 19, e06651. [Google Scholar] [CrossRef]
- Borges, K.A.; Furian, T.Q.; De Souza, S.; Tondo, E.C.; Streck, A.F.; Salle, C.T.; De Souza-Moraes, H.L.; Do Nascimento, V.P. Spread of a Major Clone of Salmonella enterica Serotype Enteritidis in Poultry and in salmonellosis Outbreaks in Southern Brazil. J. Food Prot. 2017, 80, 158–163. [Google Scholar] [CrossRef]
- Li, S.; He, Y.; Mann, D.A.; Deng, X. Global spread of Salmonella Enteritidis via centralized sourcing and international trade of poultry breeding stocks. Nat. Commun. 2021, 12, 5109. [Google Scholar] [CrossRef]
- Punchihewage-Don, A.J.; Hawkins, J.; Adnan, A.M.; Hashem, F.; Parveen, S. The outbreaks and prevalence of antimicrobial resistant Salmonella in poultry in the United States: An overview. Heliyon 2022, 8, e11571. [Google Scholar] [CrossRef]
- Sadiq, S.; Ahmad, M.; Chaudhry, M.; Akbar, H.; Muhammad Hassan, M.; Shehzad, F.; Hassan, S.; Khan, M.U.Z. Molecular epidemiology of zoonotic Salmonella enteritidis isolated from poultry and human sources by multi locus sequence typing. Pak. Vet. J. 2021, 41, 264–268. [Google Scholar]
- Castro-Vargas, R.E.; Herrera-Sánchez, M.P.; Rodríguez-Hernández, R.; Rondón-Barragán, I.S. Antibiotic resistance in Salmonella spp. isolated from poultry: A global overview. Vet. World 2020, 13, 2070–2084. [Google Scholar] [CrossRef]
- Mthembu, T.P.; Zishiri, O.T.; el Zowalaty, M.E. Genomic characterization of antimicrobial resistance in food chain and livestock-associated Salmonella species. Animals 2021, 11, 872. [Google Scholar] [CrossRef] [PubMed]
- Osterhaus, A.; Valangendonck, C.; Barbeschi, M.; Bruschke, C.; Christensen, R.; Dazak, P.; de Groot, F.; Doherty, P.; Drury, P.; Wagenaar, J. Make science evolve into a One Health approach to improve health and security: A white paper. One Health Outlook 2020, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Delgado-González, R.A.; González-Álvarez, V.; Rodríguez-Martínez, R.; Véliz-Deras, F.G. Prevalence of Escherichia coli and Salmonella spp. in Diarrheic Holstein Calves from La Comarca Lagunera, Mexico. AGROFAZ 2016, 16, 57–64. [Google Scholar]
- Ballesteros-Nova, N.E.; Sánchez, S.; Steffani, J.; Sierra, L.; Chen, Z.; Ruíz-López, F.; Bell, R.; Reed, E.; Balkey, M.; Dudley, E. Genomic Epidemiology of Salmonella enterica Circulating in Surface Waters Used in Agriculture and Aquaculture in Central Mexico. Appl. Environ. Microbiol. 2022, 88, e0214921. [Google Scholar] [CrossRef] [PubMed]
- Schlundt, J.; Toyofuku, H.; Jansen, J.; Herbst, S.A. Emerging food-borne zoonoses. OIE Rev. Sci. Tech. 2004, 23, 513–533. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, C.J.; Cardwell, D.M.; Moeller, R.B.; Gray, G.C. Humans and cattle: A review of bovine zoonoses. Vector-Borne Zoonotic Dis. 2014, 14, 1–19. [Google Scholar] [CrossRef]
- Cummings, K.J.; Warnick, L.D.; Alexander, K.A.; Cripps, C.J.; Gröhn, Y.T.; McDonough, P.L.; Nydam, D.V.; Reed, K.E. The incidence of salmonellosis among dairy herds in the northeastern United States. J. Dairy. Sci. 2009, 92, 3766–3774. [Google Scholar] [CrossRef] [PubMed]
- de la Garza-García, J.; Lozano, M.C.; Wacher-Rodarte, M.; Ocaña, A.; Hernández-Castro, R.; Xicohtencatl-Cortes, J.; Suárez, E. Frecuencia de contaminación y de serotipos de Salmonella enterica y Escherichia coli en una operación integrada de matanza y deshuese de bovinos. Rev. Mex. Cienc. Pecu. 2021, 11, 971–990. [Google Scholar] [CrossRef]
- Gutema, F.D.; Agga, G.E.; Abdi, R.D.; de Zutter, L.; Duchateau, L.; Gabriël, S. Prevalence and serotype diversity of Salmonella in apparently healthy cattle: Systematic review and meta-analysis of published studies, 2000-2017. Front. Vet. Sci. 2019, 6, 102. [Google Scholar] [CrossRef]
- U.S Department of Agriculture. Salmonella, Listeria, and Campylobacter on U.S Dairy Operations, 1996–2007; USDA-APHIS, VS, CEAH: Fort Collins, CO, USA, 2007.
- Crucello, A.; Furtado, M.M.; Chaves, M.D.R.; Sant’Ana, A.S. Transcriptome sequencing reveals genes and adaptation pathways in Salmonella typhimurium inoculated in four low water activity foods. Food Microbiol. 2019, 82, 426–435. [Google Scholar] [CrossRef]
- Doyle, M.P.; Buchanan, R.L. Food Microbiology: Fundamentals and Frontiers, 4th ed.; ASM Press: Washington, DC, USA, 2012; pp. 229–230. [Google Scholar]
- Spector, M.P.; Kenyon, W.J. Resistance and survival strategies of Salmonella enterica to environmental stresses. Food Res. Int. 2012, 45, 455–481. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Desvaux, M.; Hébraud, M.; Møretrø, T.; Langsrud, S.; Doulgeraki, A.; Nychas, G.; Kacániová, M.; Simões, M. Intra- and inter-species interactions within biofilms of important foodborne bacterial pathogens. Front. Microbiol. 2015, 6, 841. [Google Scholar] [CrossRef]
- Gutiérrez-Cogco, L.; Montiel-Vázquez, E.; Aguilera-Pérez, P.; González-Andrade, M.C. Serotipos de Salmonella identificados en los servicios de salud de México. Salud Publica Mex. 2000, 42, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Habing, G.G.; Manning, S.; Bolin, C.; Cui, Y.; Rudrik, J.; Dietrich, S.; Kaneen, J. Within-farm changes in dairy farm-associated Salmonella subtypes and comparison to human clinical isolates in Michigan, 2000–2001 and 2009. Appl Environ. Microbiol 2015, 81, 5724–5735. [Google Scholar] [CrossRef] [PubMed]
- Banerji, S.; Simon, S.; Tille, A.; Fruth, A.; Flieger, A. Genome-based Salmonella serotyping as the new gold standard: A report of the Establishing Next Generation Sequencing Ability for Genomic Analysis in Europe (ENGAGE) consortium identified four sero-typing tools, specifically Metric-Oriented Sequence Typer (MOST). Sci. Rep. 2020, 10, 4333. [Google Scholar] [CrossRef]
- Diep, B.; Barretto, C.; Portmann, A.; Fournier, A.; Karczmarek, A.; Voets, G.; Li, S.; Deng, X.; Klijn, A. Salmonella Serotyping; Comparison of the Traditional Method to a Microarray-Based Method and an in silico Platform Using Whole Genome Sequencing Data. Front. Microbiol. 2019, 10, 2554. [Google Scholar] [CrossRef] [PubMed]
- Siddique, A.; Ullah, N.; Ali, A.; Patel, A.; Moore, T.; Kenney, S.; Ganda, E.; Rahman, A. Draft genome sequences of 25 Salmonella enterica serovar Agona strains isolated from poultry and associated food products harbouring multiple antibiotic resistance genes. J. Glob. Antimicrob. Resist. 2022, 29, 131–135. [Google Scholar] [CrossRef]
- Gopal, M.; Elumali, S.; Arumugam, S.; Durairajpandian, V.; Kannan, M.; Selvam, E.; Seetharaman, S. GyrA ser83 and ParC trp106 Mutations in Salmonella enterica Serovar Typhi Isolated from Typhoid Fever Patients in Tertiary Care Hospital. J. Clin. Diagn. Res. 2016, 10, DC14–DC18. [Google Scholar] [CrossRef] [PubMed]
- Davidson, K.E.; Byrne, B.A.; Pires, A.F.A.; Magdesian, K.G.; Pereira, R. Antimicrobial resistance trends in fecal Salmonella isolates from northern California dairy cattle admitted to a veterinary teaching hospital, 2002–2016. PLoS ONE 2018, 13, e0199928. [Google Scholar] [CrossRef]
- Chuanchuen, R.; Ajariyakhajorn, K.; Koowatananukul, C.; Wannaprasat, W.; Khemtong, S.; Samngamnim, S. Antimicrobial Resistance and Virulence Genes in Salmonella enterica Isolates from Dairy Cows. Foodborne Pathog. Dis. 2010, 7, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Addis, Z.; Kebede, N.; Worku, Z.; Gezahegn, H.; Yirsaw, A.; Kassa, T. Prevalence and antimicrobial resistance of Salmonella isolated from lactating cows and in contact humans in dairy farms of Addis Ababa: A cross sectional study. BMC Infect. Dis. 2011, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, R.; Barh, D.; Weimer, B.; Viana, M.; Profeta, R.; Sousa, T.; Aburjaile, F.; Quino, W.; Souza, R.; Mestanza, O.; et al. WGS-Based Lineage and Antimicrobial Resistance Pattern of Salmonella typhimurium Isolated during 2000–2017 in Peru. Antibiotics 2022, 11, 1170. [Google Scholar] [CrossRef] [PubMed]
- Calia, C.; Oliva, M.; Ferrara, M.; Minervini, C.; Scrascia, M.; Monno, R.; Mulè, G.; Cumbo, C.; Marzella, A.; Pazzani, C. Identification and Characterisation of pST1023 A Mosaic, Multidrug-Resistant and Mobilisable IncR Plasmid. Microorganisms 2022, 10, 1592. [Google Scholar] [CrossRef]
- Jibril, A.H.; Okeke, I.N.; Dalsgaard, A.; Menéndez, V.G.; Olsen, J.E. Genomic analysis of antimicrobial resistance and resistance plasmids in Salmonella serovars from poultry in Nigeria. Antibiotics 2021, 10, 99. [Google Scholar] [CrossRef]
- Wang, J.; Xue, K.; Yi, P.; Zhu, X.; Peng, Q.; Wang, Z.; Peng, Y.; Chen, Y.; Robertson, I.D.; Li, X.; et al. An abattoir-based study on the prevalence of salmonella fecal carriage and esbl related antimicrobial resistance from culled adult dairy cows in Wuhan, China. Pathogens 2020, 9, 853. [Google Scholar] [CrossRef]
- Thahir, S.S.A.; Rajendiran, S.; Shaharudin, R.; Veloo, Y. Multidrug-Resistant Salmonella Species and Their Mobile Genetic Elements from Poultry Farm Environments in Malaysia. Antibiotics 2023, 12, 1330. [Google Scholar] [CrossRef]
- Ramos-Morales, F. Impact of Salmonella enterica Type III Secretion System Effectors on the Eukaryotic Host Cell. Cell Biol. 2012, 2012, 787934. [Google Scholar] [CrossRef]
- Spanò, S.; Ugalde, J.E.; Galán, J.E. Delivery of a Salmonella typhi Exotoxin from a Host Intracellular Compartment. Cell Host Microbe 2008, 3, 30–38. [Google Scholar] [CrossRef]
- Mthembu, T.P.; Zishiri, O.T.; el Zowalaty, M.E. Detection and Molecular Identification of Salmonella Virulence Genes in Livestock Production Systems in South Africa. Pathogens 2019, 8, 124. [Google Scholar] [CrossRef]
- Juhas, M.; van der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.V.; Harhay, D.; Bono, J.; Smith, T.; Fields, P.; Dinsmore, B.; Santovenia, M.; Wang, R.; Bosilevac, J.; Harhay, J. Comparative genomics of Salmonella enterica serovar Montevideo reveals lineage-specific gene differences that may influence ecological niche association. Microb. Genom. 2018, 4, e000202. [Google Scholar] [CrossRef] [PubMed]
- Levent, G.; Schlochtermeir, A.; Ives, S.; Norman, K.; Lawhon, S.; Loneragan, G.; Anderson, R.; Vinasco, J.; Scott, H. Population Dynamics of Salmonella enterica within Beef Cattle Cohorts Followed from Single-Dose Metaphylactic Antibiotic Treatment until Slaughter. Appl. Environ. Microbiol. 2019, 85, e01386-19. [Google Scholar] [CrossRef] [PubMed]
- Servicio de Información Agroalimentaria y Pesquera. Bovino Para Leche: Población Ganadera 2012–2021; Secretaría de Agricultura y Desarrollo Rural: Mexico City, Mexico, 2023. [Google Scholar]
- Soria, M.A.; Bueno, D.J. Culture Based Methods to Detect Salmonella from Different Poultry Products; Nova Science Publishers: New York, NY, USA, 2016. [Google Scholar]
- Fan, H.; Wu, Q.; Kou, X. Co-detection of five species of water-borne bacteria by multiplex PCR. Life Sci. J. 2008, 5, 1097–8135. [Google Scholar]
- Rodulfo, H.; De Donato, M.; Luiggi, J.; Michelli, E.; Millán, A.; Michelli, M. Molecular characterization of Salmonella strains in individuals with acute diarrhea syndrome in the State of Sucre, Venezuela. Rev. Soc. Bras. Med. Trop. 2012, 45, 329–333. [Google Scholar] [CrossRef] [PubMed]
- McDermott, P.F.; Zhao, S.; Tate, H. Antimicrobial Resistance in Nontyphoidal Salmonella. Microbiol. Spectr. 2018, 6, ARBA-0014-2017. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI supplement M100; Clinical & Laboratory Standards Institute: Wayne, PA, USA, 2019; p. 332. [Google Scholar]
- Rodulfo, H.; Horta, M.; Mata, G.; Gutiérrez, R.; González, Y.; Michelli, E.; Guzman, M.; Martinez, D.; Sharma, A.; De Donato, M. Negative correlation between virulence and multidrug resistance in intrahospital and community acquired infections by Proteus mirabilis, in Eastern Venezuela. Invest. Clin. 2021, 62, 37–51. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data; Babraham Institute: Cambridge, UK, 2010; Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 15 July 2023).
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Zhang, S.; den Bakker, H.C.; Li, S.; Chen, J.; Dinsmore, B.A.; Lane, C.; Lauer, A.C.; Fields, P.I.; Deng, X. SeqSero2: Rapid and Improved Salmonella Serotype Determination Using Whole-Genome Sequencing Data. Appl. Environ. Microbiol. 2019, 85, 23. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.; Ruppe, E.; Roberts, M.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.; Rebelo, R.; Aarestrup, F. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Aarestrup, F.; Hasman, H.A. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [PubMed]
- Roer, L.; Hendriksen, R.S.; Leekitcharoenphon, P.; Lukjancenko, O.; Kaas, R.; Hasman, H.; Aaerestrup, F. Is the Evolution of Salmonella enterica subsp. enterica Linked to Restriction-Modification Systems. mSystems 2016, 1, 9–16. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Serological Typing | Genome Based in Silico Serotyping |
|---|---|---|
| SETLX55C | B | E |
| SESLP86C | C | C |
| SEDGO167B | B | E |
| SECOA243PC | C | E |
| SECOA252B | C | B |
| SEDGO294PC | C | C |
| SEDGO298PC | B | E |
| SEDGO304PC | C | C |
| SECOA310PC | C | C |
| SEDGO318B | D | G |
| SECOA320C | C | G |
| SECOA322PC | C | C |
| Isolate | Serovar | Antibiotic Resistance Genes |
|---|---|---|
| SETLX55C | London | qnrB19, sul2, aph(6)-Id, aph(3′)-Id, aac(6′)-Iaa |
| SEBC65B | London | aph(6)-Id_1, tet(A)_6, aph(3′)-Id_1, sul2_2, qnrB19_1, aac(6′)-Iaa_1 |
| SESLP86C | Newport | aac(6′)-Iaa_1, dfrA1_8, tet(A)_6, floR_2, sul1_15, mph(A)_2, aadA2_1, blaCARB-2_1, qnrA1_1, qacE |
| SEAGS147B | Give | aac(6′)-Iaa_1 |
| SEDGO167B | Give | aac(6′)-Iaa_1 |
| SECOA225PC | Anatum | aac(6′)-Iaa_1 |
| SECOA243PC | Anatum | aac(6′)-Iaa_1 |
| SECOA246B | Anatum | aac(6′)-Iaa_1 |
| SECOA245B | Reading | tet(C)_3, fosA7_1, qnrB19_1, aac(6′)-Iaa_1 |
| SECOA252B | Agona | aac(2′)-IIa_1, fosA7_1, aac(6′)-Iaa_1 |
| SECOA255PC | Meleagridis | fosA7_1, aac(6′)-Iaa_1 |
| SEDGO269B | Meleagridis | aac(6′)-Iaa_1, fosA7_1, aac(2′)-IIa_1 |
| SEDGO298PC | Meleagridis | aac(6′)-Iaa_1, fosA7_1 |
| SEDGO276B | Havana | aac(6′)-Iaa_1 |
| SEDGO294PC | Montevideo | aac(6′)-Iaa_1 |
| SEDGO304PC | Montevideo | aac(6′)-Iaa_1 |
| SECOA310PC | Montevideo | aac(6′)-Iaa_1, blaZ_62, mph(C)_2, msr(A)_1 |
| SECOA322PC | Montevideo | aac(6′)-Iaa_1 |
| SEDGO318B | Cubana | aac(6′)-Iaa_1 |
| SECOA320C | Cubana | aac(6′)-Iaa_1 |
| Plasmid | Serovar |
|---|---|
| Col(pHAD28) | London, Reading |
| IncQ1 | London |
| ColpVC | London |
| IncR | Newport |
| Col440I | Reading |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrera, S.; Vázquez-Flores, S.; Needle, D.; Rodríguez-Medina, N.; Iglesias, D.; Sevigny, J.L.; Gordon, L.M.; Simpson, S.; Thomas, W.K.; Rodulfo, H.; et al. Serovars, Virulence and Antimicrobial Resistance Genes of Non-Typhoidal Salmonella Strains from Dairy Systems in Mexico. Antibiotics 2023, 12, 1662. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12121662
Barrera S, Vázquez-Flores S, Needle D, Rodríguez-Medina N, Iglesias D, Sevigny JL, Gordon LM, Simpson S, Thomas WK, Rodulfo H, et al. Serovars, Virulence and Antimicrobial Resistance Genes of Non-Typhoidal Salmonella Strains from Dairy Systems in Mexico. Antibiotics. 2023; 12(12):1662. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12121662
Chicago/Turabian StyleBarrera, Stephany, Sonia Vázquez-Flores, David Needle, Nadia Rodríguez-Medina, Dianella Iglesias, Joseph L. Sevigny, Lawrence M. Gordon, Stephen Simpson, W. Kelley Thomas, Hectorina Rodulfo, and et al. 2023. "Serovars, Virulence and Antimicrobial Resistance Genes of Non-Typhoidal Salmonella Strains from Dairy Systems in Mexico" Antibiotics 12, no. 12: 1662. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics12121662