Application of Bacteriophages to Control Vibrio alginolyticus Contamination in Oyster (Saccostrea glomerata) Larvae


Abstract
:1. Introduction
2. Results
2.1. Vibrio spp. Isolation and Pathogenicity Test
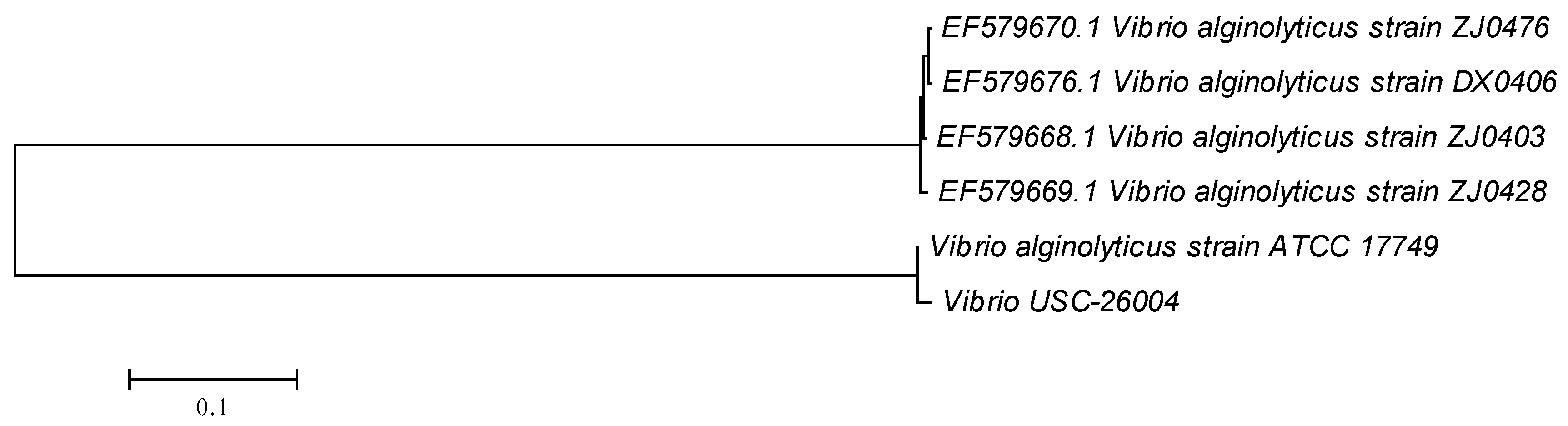
2.2. Prophage Detection in Bacterial Isolates and Antibiotic Susceptibility
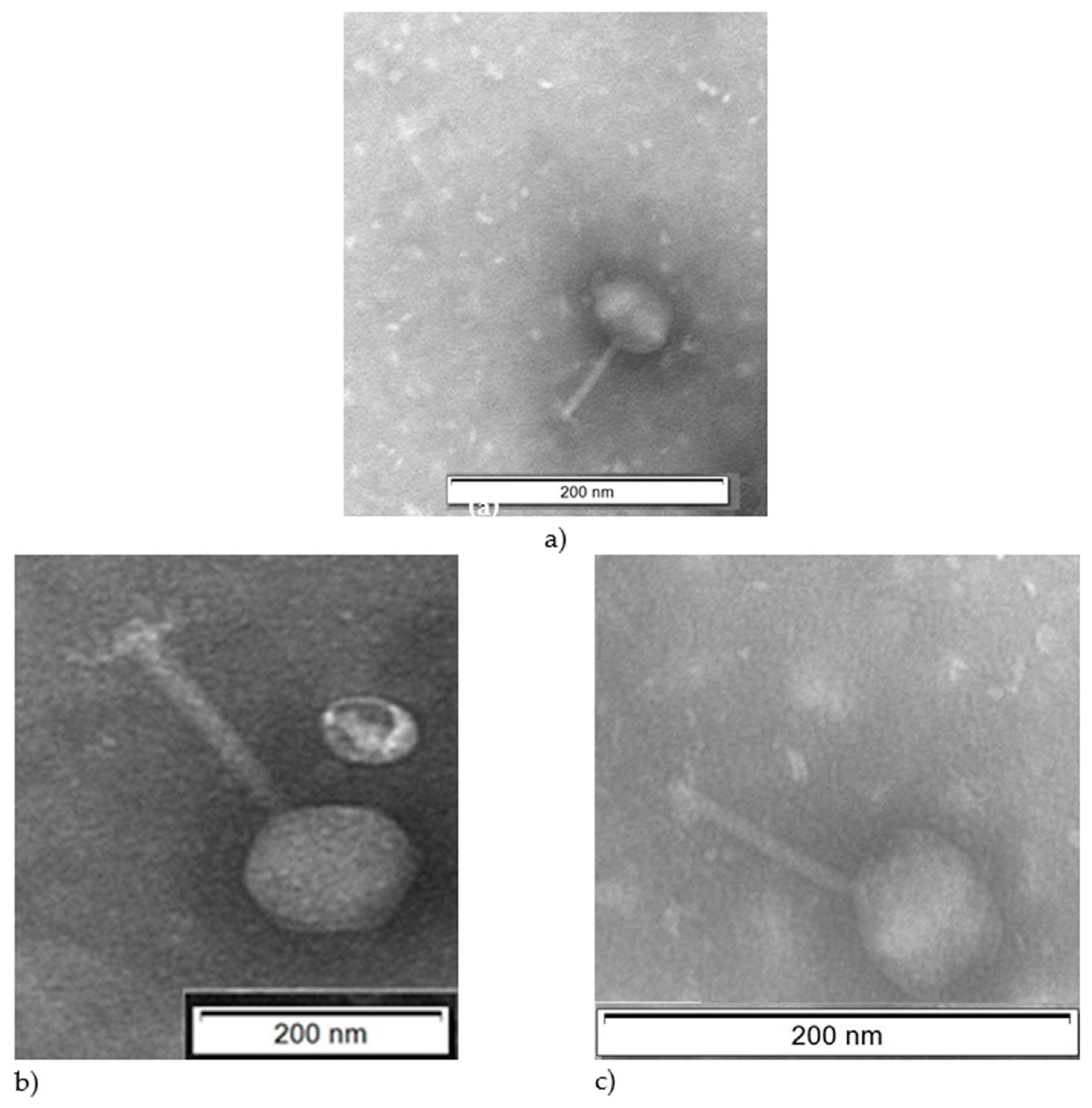
2.3. Isolation and Characterization of Bacteriophages
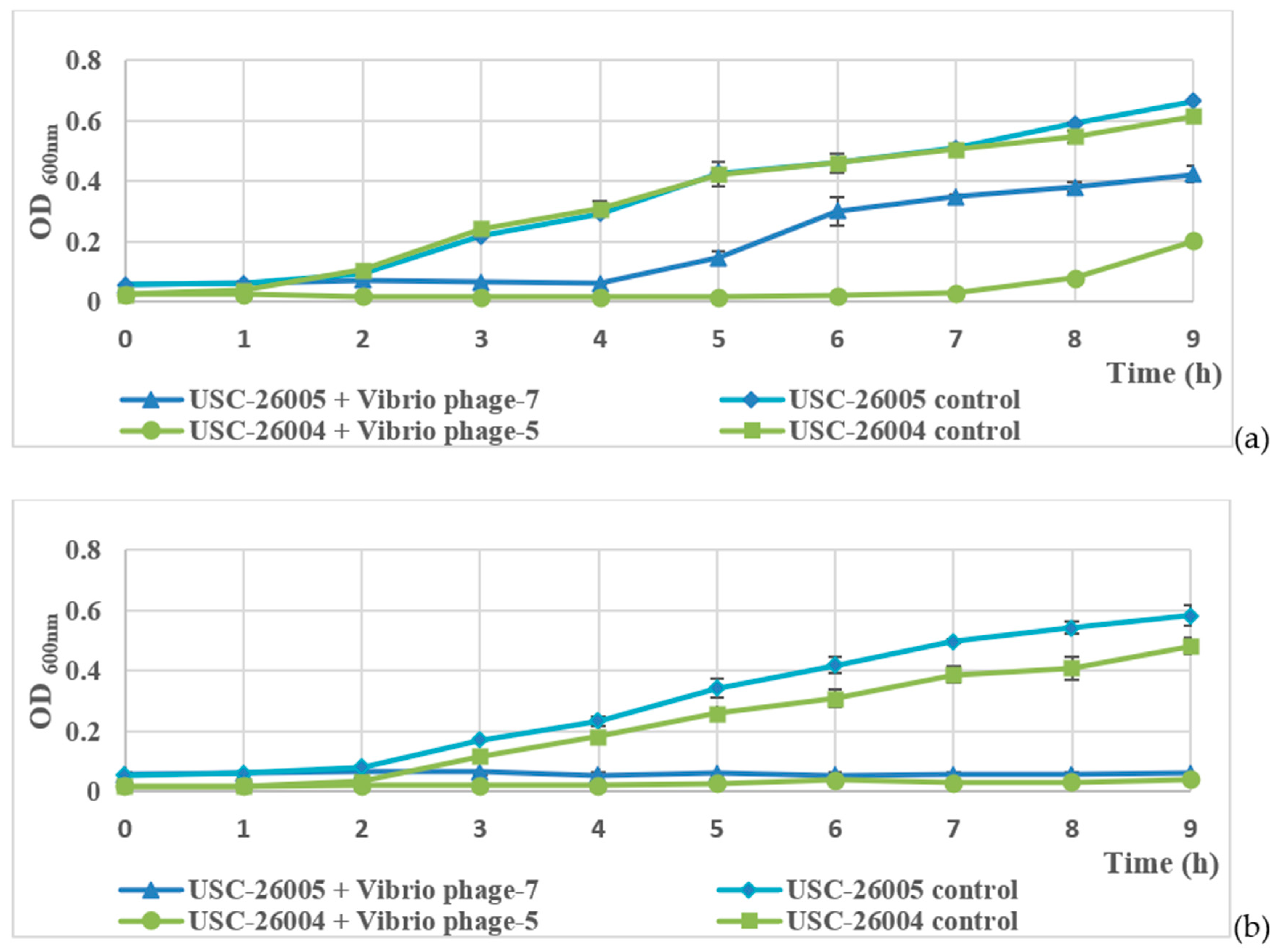
2.4. Host Range and Bacteriophage Lytic Ability
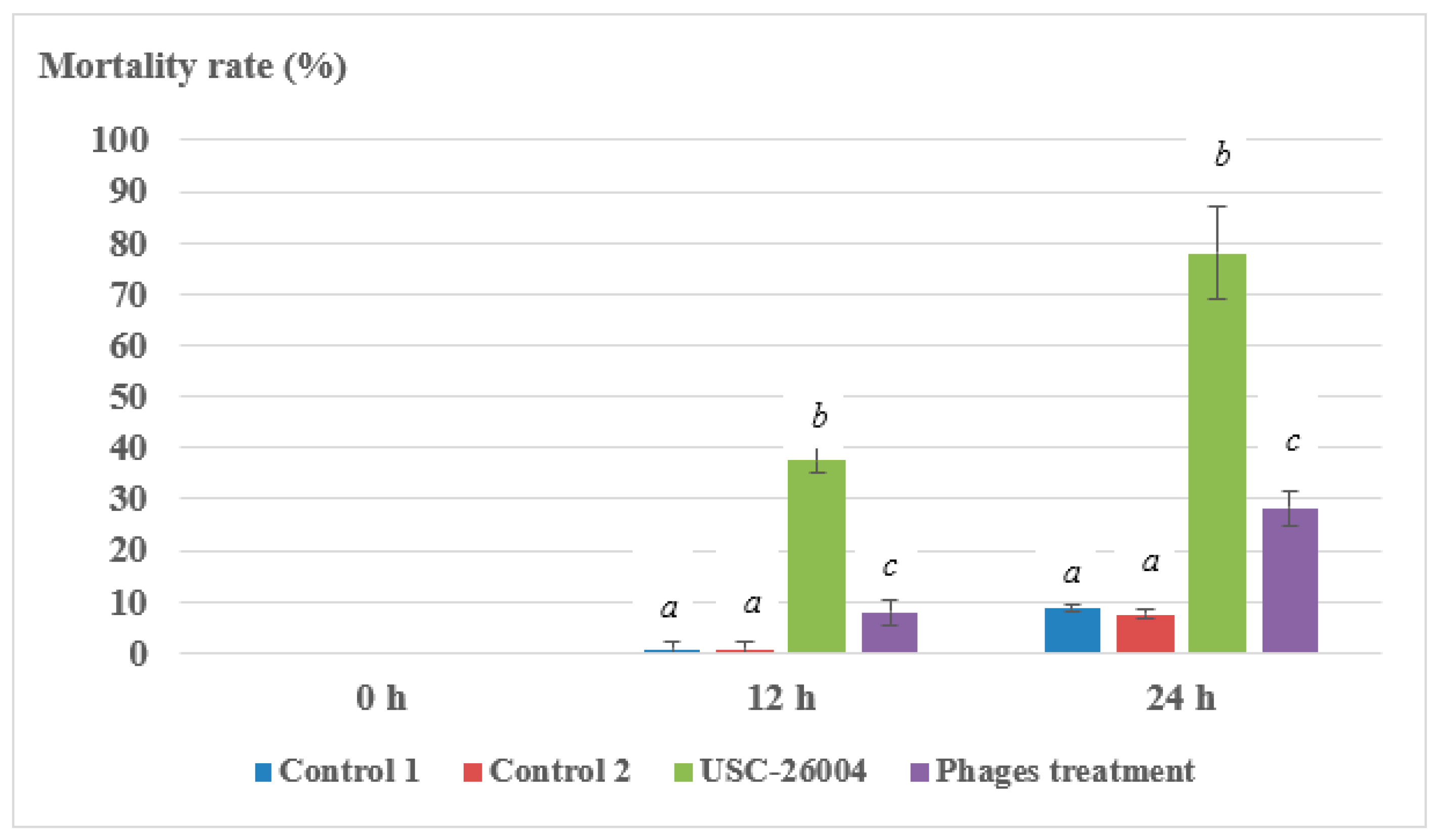
2.5. Laboratory Trial of Bacteriophages Against Vibrio spp.
3. Discussion
4. Materials and Methods
4.1. Isolation and Characterization of Vibrio spp. Isolated from Oyster Hatcheries
4.2. Pathogenicity Test and Prophage Induction
4.3. Isolation and Characterization of Bacteriophages
4.4. Bacteriophage Lytic Activity
4.5. Bacteriophage Control of Vibrio spp. with Oyster Larvae
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Lucas, J.S. Bivalve molluscs. In Aquaculture: Farming Aquatic Animals and Plants, 3rd ed.; Lucas, J.S., Southgate, P.C., Eds.; Wiley-Blackwell Publishing Ltd.: Oxford, UK, 2019; pp. 541–566. [Google Scholar]
- Elston, R.A.; Hasegawa, H.; Humphrey, K.L.; Polyak, I.K.; Häse, C.C. Re-emergence of Vibrio tubiashii in bivalve shellfish aquaculture: Severity, environmental drivers, geographic extent and management. Dis. Aquat. Organ. 2008, 82, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Romalde, J.; Barja, J. Bacteria in molluscs: Good and bad guys. Curr. Res. Technol. Educ. Top. Appl. Microbiol. Microb. Biotechnol. 2010, 1, 136–147. [Google Scholar]
- Tubiash, H.S.; Chanley, P.E.; Leifson, E. Bacillary necrosis, a disease of larval and juvenile bivalve mollusks I. etiology and epizootiology. J. Bacteriol. 1965, 90, 1036–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugumar, G.; Nakai, T.; Hirata, Y.; Matsubara, D.; Muroga, K. Vibrio splendidus biovar II as the causative agent of bacillary necrosis of Japanese oyster Crassostrea gigas larvae. Dis. Aquat. Organ. 1998, 33, 111–118. [Google Scholar] [CrossRef]
- Prado, S.; Dubert, J.; Romalde, J.L.; Toranzo, A.E.; Barja, J.L. Vibrio ostreicida sp. nov., a new pathogen of bivalve larvae. Int. J. Syst. Evol. Microbiol. 2014, 64, 1641–1646. [Google Scholar] [CrossRef]
- Dubert, J.; Nelson, D.R.; Spinard, E.J.; Kessner, L.; Gomez-Chiarri, M.; da Costa, F.; Prado, S.; Barja, J.L. Following the infection process of vibriosis in manila clam (Ruditapes philippinarum) larvae through gfp-tagged pathogenic Vibrio species. J. Invertebr. Pathol. 2016, 133, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Southgate, P.C. Foods and feeding. In Aquaculture: Farming Aquatic Animals and Plants, 3rd ed.; Lucas, J.S., Southgate, P.C., Eds.; Wiley-Blackwell Publishing Ltd.: Oxford, UK, 2019; pp. 188–213. [Google Scholar]
- Dubert, J.; Barja, J.L.; Romalde, J.L. New insights into pathogenic Vibrios affecting bivalves in hatcheries: Present and future prospects. Front. Microbiol. 2017, 8, 762. [Google Scholar] [CrossRef] [Green Version]
- Defoirdt, T.; Boon, N.; Sorgeloos, P.; Verstraete, W.; Bossier, P. Alternatives to antibiotics to control bacterial infections: Luminescent vibriosis in aquaculture as an example. Trends Biotechnol. 2007, 25, 472–479. [Google Scholar] [CrossRef] [Green Version]
- Karunasagar, I.; Pai, R.; Malathi, G.; Karunasagar, I. Mass mortality of Penaeus monodon larvae due to antibiotic-resistant Vibrio harveyi infection. Aquaculture 1994, 128, 203–209. [Google Scholar] [CrossRef]
- Igbinosa, E.O. Detection and antimicrobial resistance of Vibrio isolates in aquaculture environments: Implications for public health. Microb. Drug Resist. 2016, 22, 238–245. [Google Scholar] [CrossRef]
- Kang, C.-H.; Shin, Y.; Jang, S.; Jung, Y.; So, J.-S. Antimicrobial susceptibility of Vibrio alginolyticus isolated from oyster in Korea. Environ. Sci. Pollut. R. 2016, 23, 21106–21112. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Barton, M.; Elliott, L.; Li, X.; Abraham, S.; O’Dea, M.; Munro, J. Bacteriophage therapy for the control of Vibrio harveyi in Greenlip abalone (Haliotis laevigata). Aquaculture 2017, 473, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Rong, R.; Lin, H.; Wang, J.; Khan, M.N.; Li, M. Reductions of Vibrio parahaemolyticus in oysters after bacteriophage application during depuration. Aquaculture 2014, 418, 171–176. [Google Scholar] [CrossRef]
- Le, T.S.; Nguyen, T.H.; Vo, H.P.; Doan, V.C.; Nguyen, H.L.; Tran, M.T.; Tran, T.T.; Southgate, P.C.; Kurtböke, D.İ. Protective effects of bacteriophages against Aeromonas hydrophila causing motile aeromonas septicemia (MAS) in striped catfish. Antibiotics 2018, 7, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Jun, J.W.; Giri, S.S.; Chi, C.; Yun, S.; Kim, S.G.; Oh, W.T. Application of the bacteriophage pVco-14 to prevent Vibrio coralliilyticus infection in Pacific oyster (Crassostrea gigas) larvae. J. Invertebr. Pathol. 2019, 167, 107244. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.-N.; Smith, D.L.; Young, R. Holins: The protein clocks of bacteriophage infections. Annu. Rev. Microbiol. 2000, 54, 799–825. [Google Scholar] [CrossRef]
- Oliveira, H.; Melo, L.D.; Santos, S.B.; Nóbrega, F.L.; Ferreira, E.C.; Cerca, N.; Azeredo, J.; Kluskens, L.D. Molecular aspects and comparative genomics of bacteriophage endolysins. J. Virol. 2013, 87, 4558–4570. [Google Scholar] [CrossRef] [Green Version]
- Fischetti, V. Development of phage lysins as novel therapeutics: A historical perspective. Viruses 2018, 10, 310. [Google Scholar] [CrossRef] [Green Version]
- Witzenrath, M.; Schmeck, B.; Doehn, J.M.; Tschernig, T.; Zahlten, J.; Loeffler, J.M.; Zemlin, M.; Müller, H.; Gutbier, B.; Schütte, H. Systemic use of the endolysin cpl-1 rescues mice with fatal pneumococcal pneumonia. Crit. Care Med. 2009, 37, 642–649. [Google Scholar] [CrossRef]
- Resch, G.; Moreillon, P.; Fischetti, V.A. A stable phage lysin (cpl-1) dimer with increased antipneumococcal activity and decreased plasma clearance. Int. J. Antimicrob. Agents 2011, 38, 516–521. [Google Scholar] [CrossRef]
- Gilmer, D.B.; Schmitz, J.E.; Euler, C.W.; Fischetti, V.A. Novel bacteriophage lysin with broad lytic activity protects against mixed infection by Streptococcus pyogenes and methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemoter. 2013, 57, 2743–2750. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Euler, C.W.; Delaune, A.; Fischetti, V.A. Using a novel lysin to help control Clostridium difficile infections. Antimicrob. Agents Chemoter. 2015, 59, 7447–7457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larpin, Y.; Oechslin, F.; Moreillon, P.; Resch, G.; Entenza, J.M.; Mancini, S. In vitro characterization of plye146, a novel phage lysin that targets gram-negative bacteria. PLoS ONE 2018, 13, e0192507. [Google Scholar] [CrossRef]
- Lood, R.; Winer, B.Y.; Pelzek, A.J.; Diez-Martinez, R.; Thandar, M.; Euler, C.W.; Schuch, R.; Fischetti, V.A. Novel phage lysin capable of killing the multidrug-resistant gram-negative bacterium Acinetobacter baumannii in a mouse bacteremia model. Antimicrob. Agents Chemoter. 2015, 59, 1983–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, H.; Vilas Boas, D.; Mesnage, S.; Kluskens, L.D.; Lavigne, R.; Sillankorva, S.; Secundo, F.; Azeredo, J. Structural and enzymatic characterization of abgp46, a novel phage endolysin with broad anti-gram-negative bacterial activity. Front. Microbiol. 2016, 7, 208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chibani-Chennoufi, S.; Bruttin, A.; Dillmann, M.-L.; Brüssow, H. Phage-host interaction: An ecological perspective. J. Bacteriol. 2004, 186, 3677–3686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado, S.; Romalde, J.L.; Montes, J.; Barja, J.L. Pathogenic bacteria isolated from disease outbreaks in shellfish hatcheries. First description of Vibrio neptunius as an oyster pathogen. Dis. Aquat. Organ. 2005, 67, 209–215. [Google Scholar] [CrossRef]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S.J.B. Viptree: The viral proteomic tree server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef]
- Watts, J.E.; Schreier, H.J.; Lanska, L.; Hale, M.S. The rising tide of antimicrobial resistance in aquaculture: Sources, sinks and solutions. Mar. Drugs 2017, 15, 158. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, H.-W. 5500 phages examined in the electron microscope. Arch. Virol. 2007, 152, 227–243. [Google Scholar] [CrossRef]
- Ackermann, H.W. Bacteriophage classification. In Bacteriophages: Biology and Applications; Kutter, E., Sulakvelidze, A., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 67–89. [Google Scholar]
- Sasikala, D.; Srinivasan, P. Characterization of potential lytic bacteriophage against Vibrio alginolyticus and its therapeutic implications on biofilm dispersal. Microb. Pathog. 2016, 101, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, Z.; Xu, Y.; Li, X.; Li, H.; Wu, F.; Wang, L.; Cao, F.; Li, Z.; Li, S. Complete genomic sequence of the Vibrio alginolyticus lytic bacteriophage pva1. Arch. Virol. 2014, 159, 3447–3451. [Google Scholar] [CrossRef] [PubMed]
- Carrias, A. Evaluation of biological agents for controlling enteric septicemia of catfish. Ph.D. Thesis, Auburn University, Auburn, AL, USA, 2011. [Google Scholar]
- Jacquemot, L.; Bettarel, Y.; Monjol, J.; Corre, E.; Halary, S.; Desnues, C.; Bouvier, T.; Ferrier-Pagès, C.; Baudoux, A.C. Therapeutic potential of a new jumbo phage that infects Vibrio coralliilyticus, a widespread coral pathogen. Front. Microbiol. 2018, 9, 2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, P.; Yun, L.; Li, Y.; Tian, Y.; Liu, Q.; Huang, W.; Hu, C. Complete genomic sequence of the Vibrio alginolyticus bacteriophage Vp670 and characterization of the lysis-related genes, cwlQ and hola. BMC Genom. 2018, 19, 741. [Google Scholar] [CrossRef]
- Yuan, Y.; Gao, M. Jumbo bacteriophages: An overview. Front. Microbiol. 2017, 8, 403. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Feng, C.; Ren, J.; Zhuang, X.; Zhang, Y.; Zhu, Y.; Dong, K.; He, P.; Guo, X.; Qin, J. A novel antimicrobial endolysin, lyspa26, against Pseudomonas aeruginosa. Front. Micriobiol. 2017, 8, 293. [Google Scholar] [CrossRef] [Green Version]
- Elliman, J. Bacteriophage of Burkholderia pseudomallei; Friend or Foe? Ph.D. Thesis, James Cook University, Queensland, Australia, 2006. [Google Scholar]
- Hertwig, S.; Bockelmann, W.; Teuber, M. Purification and characterization of the lytic activity induced by the prolate-headed bacteriophage p001 in Lactococcus lactis. J. Appl. Microbiol. 1997, 82, 233–239. [Google Scholar] [CrossRef]
- Pastagia, M.; Schuch, R.; Fischetti, V.A.; Huang, D.B. Lysins: The arrival of pathogen-directed anti-infectives. J. Med. Microbiol. 2013, 62, 1506–1516. [Google Scholar] [CrossRef]
- Crothers-Stomps, C.; Høj, L.; Bourne, D.; Hall, M.; Owens, L. Isolation of lytic bacteriophage against Vibrio harveyi. J. Appl. Microbiol. 2010, 108, 1744–1750. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16s ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Pang, L.; Zhang, X.H.; Zhong, Y.; Chen, J.; Li, Y.; Austin, B. Identification of Vibrio harveyi using PCR amplification of the toxr gene. Lett. Appl. Microbiol. 2006, 43, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing: Twentieth Informational Supplement m100-s20; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2010. [Google Scholar]
- Clokie, M.R.; Kropinski, A. Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions; Humana Press: Totowa, NJ, USA, 2009. [Google Scholar]
- Estes, R.M.; Friedman, C.S.; Elston, R.A.; Herwig, R.P. Pathogenicity testing of shellfish hatchery bacterial isolates on pacific oyster Crassostrea gigas larvae. Dis. Aquat. Organ. 2004, 58, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Ma, J. The global catalogue of microorganisms (GCM) 10k type strain sequencing project: Providing services to taxonomists for standard genome sequencing and annotation. Int. J. Syst. Evol. Microbiol. 2019, 69, 895–898. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vibrio Species | BLAST (Identity %) | Pathogenicity Rate (%) |
|---|---|---|
| USC-26004 | V. alginolyticus (99%) | 17.7 |
| USC-26005 | V. alginolyticus (99%) | 2.5 |
| ATCC 14126 | V. harveyi | 16.6 |
| ATCC 17749 | V. alginolyticus | 12.9 |
| Bacteriophage | Head (nm) | Nucleic Acid | Neck (nm) | Tail Sheath (nm) | Family | |||
|---|---|---|---|---|---|---|---|---|
| L | W | L | W | L | W | |||
| Φ-5 | 131.8 | 122.7 | DNA, ds | 11.4 | 6.8 | 129.5 | 18.2 | Myoviridae |
| Φ-6 | NM | 96.0 | DNA, ds | NM | NM | 162.2 | 16.1 | Myoviridae |
| Φ-7 | 68.6 | 59.1 | DNA, ds | 4.76 | 3.8 | 81.9 | 8.6 | Myoviridae |
| Vibrio spp. | Vibrio Φ-5 | Vibrio Φ-6 | Vibrio Φ-7 |
|---|---|---|---|
| USC-26004 | + | + | + |
| USC-26005 | PH | PH | PH |
| ATCC 14126 | - | + | - |
| ATCC 17749 | - | - | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le, T.S.; Southgate, P.C.; O’Connor, W.; Vu, S.V.; Kurtböke, D.İ. Application of Bacteriophages to Control Vibrio alginolyticus Contamination in Oyster (Saccostrea glomerata) Larvae. Antibiotics 2020, 9, 415. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070415
Le TS, Southgate PC, O’Connor W, Vu SV, Kurtböke Dİ. Application of Bacteriophages to Control Vibrio alginolyticus Contamination in Oyster (Saccostrea glomerata) Larvae. Antibiotics. 2020; 9(7):415. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070415
Chicago/Turabian StyleLe, Tuan Son, Paul C. Southgate, Wayne O’Connor, Sang V. Vu, and D. İpek Kurtböke. 2020. "Application of Bacteriophages to Control Vibrio alginolyticus Contamination in Oyster (Saccostrea glomerata) Larvae" Antibiotics 9, no. 7: 415. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9070415