Melatonin Alleviates Hypoxia-Induced Apoptosis of Granulosa Cells by Reducing ROS and Activating MTNR1B–PKA–Caspase8/9 Pathway

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Reagents and Antibodies
2.3. Cell Culture and Treatments
2.4. Western Blotting
2.5. Apoptosis by Flow Cytometry Analysis
2.6. Cell Viability Assay
2.7. Detection of ROS by Immunofluorescence-Confocal Microscopy and Flow Cytometry Analysis
2.8. Real-Time Quantitative RT-PCR Analysis
2.9. PKA Detection
2.10. Statistics Analysis
3. Results
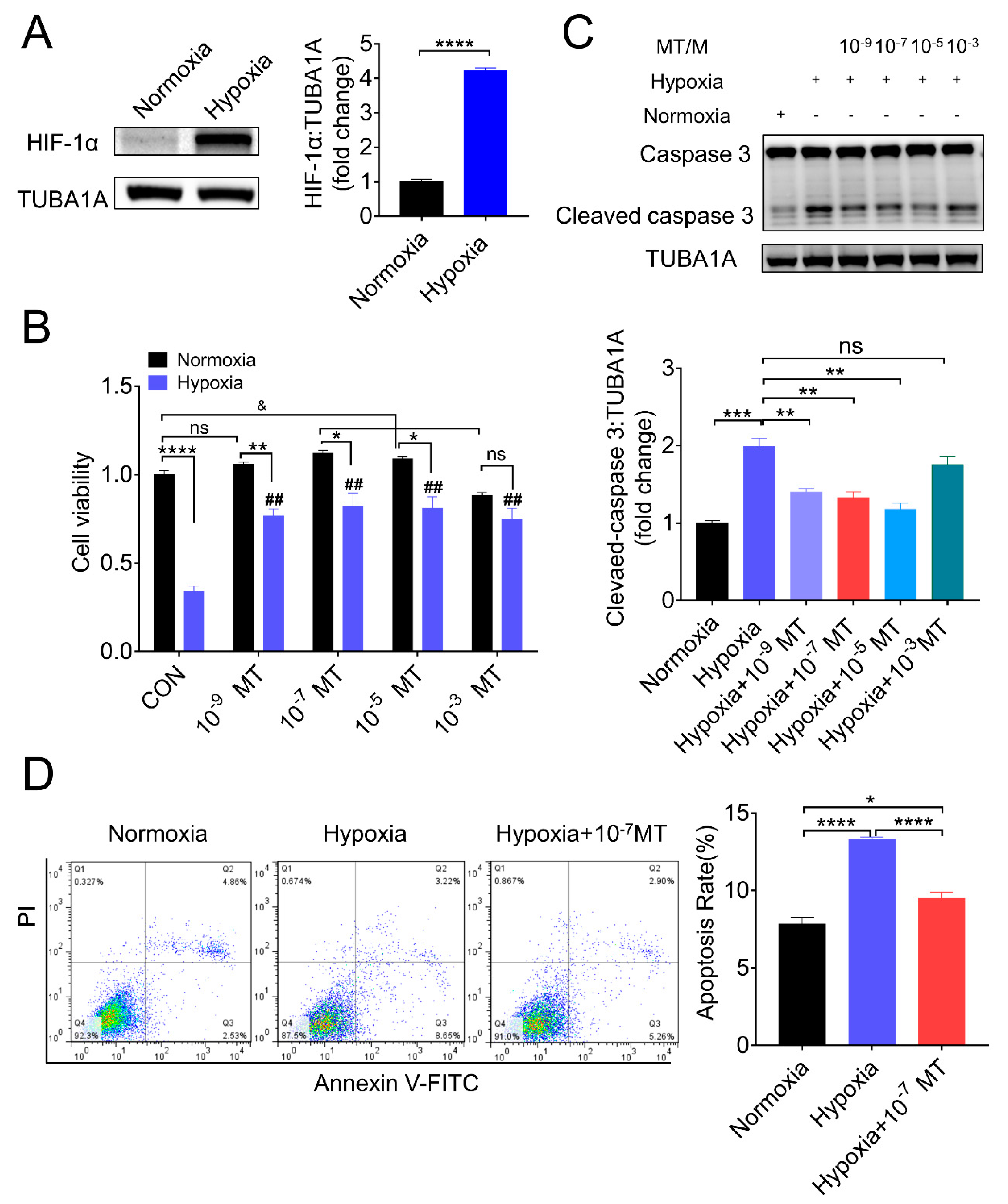
3.1. Melatonin Reduced Apoptosis and Rescued Cell Viability of Porcine Granulosa Cells (GCs) in the Setting of Hypoxia
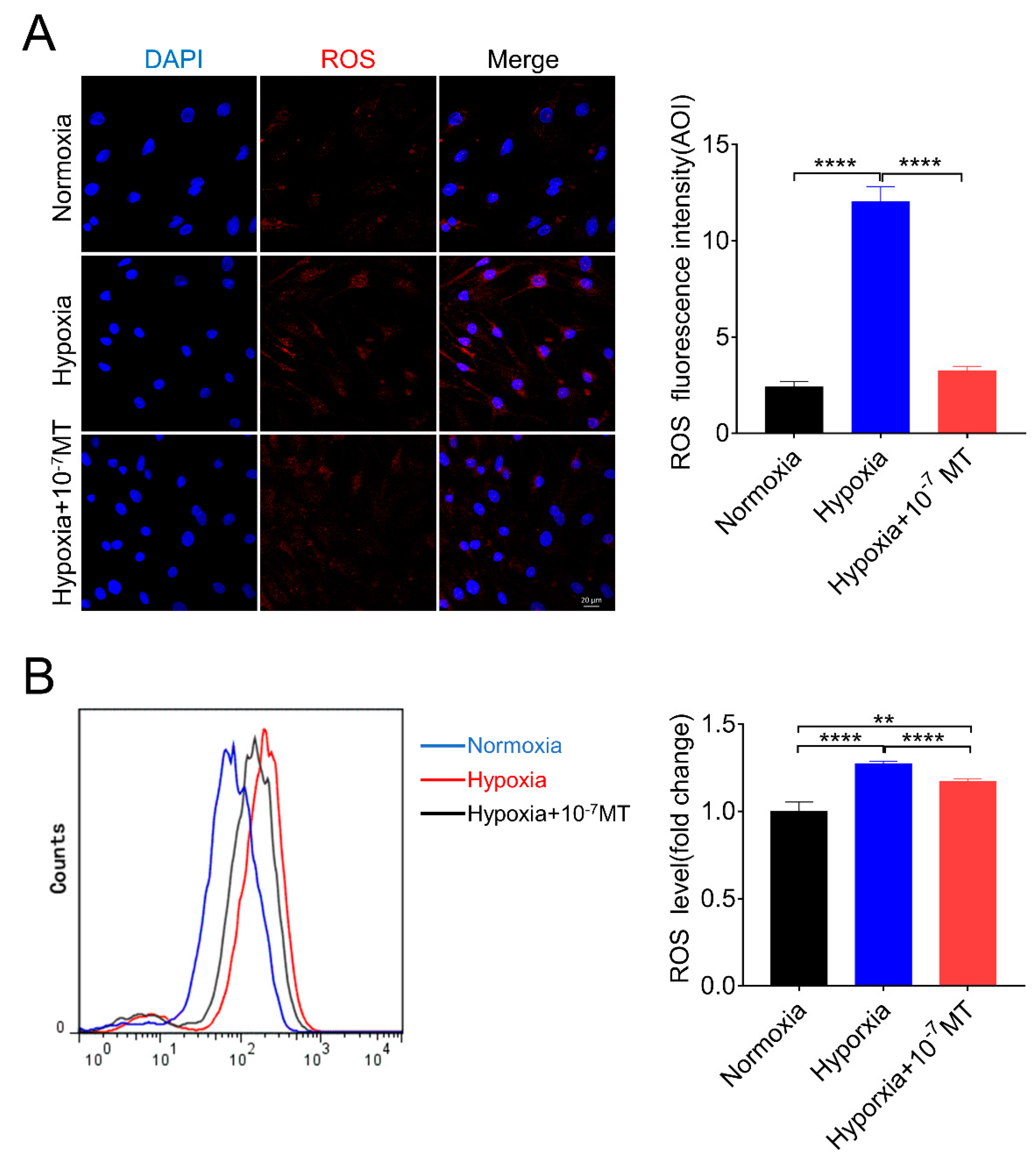
3.2. Melatonin Lowered ROS Level Caused by Hypoxia in Porcine GCs
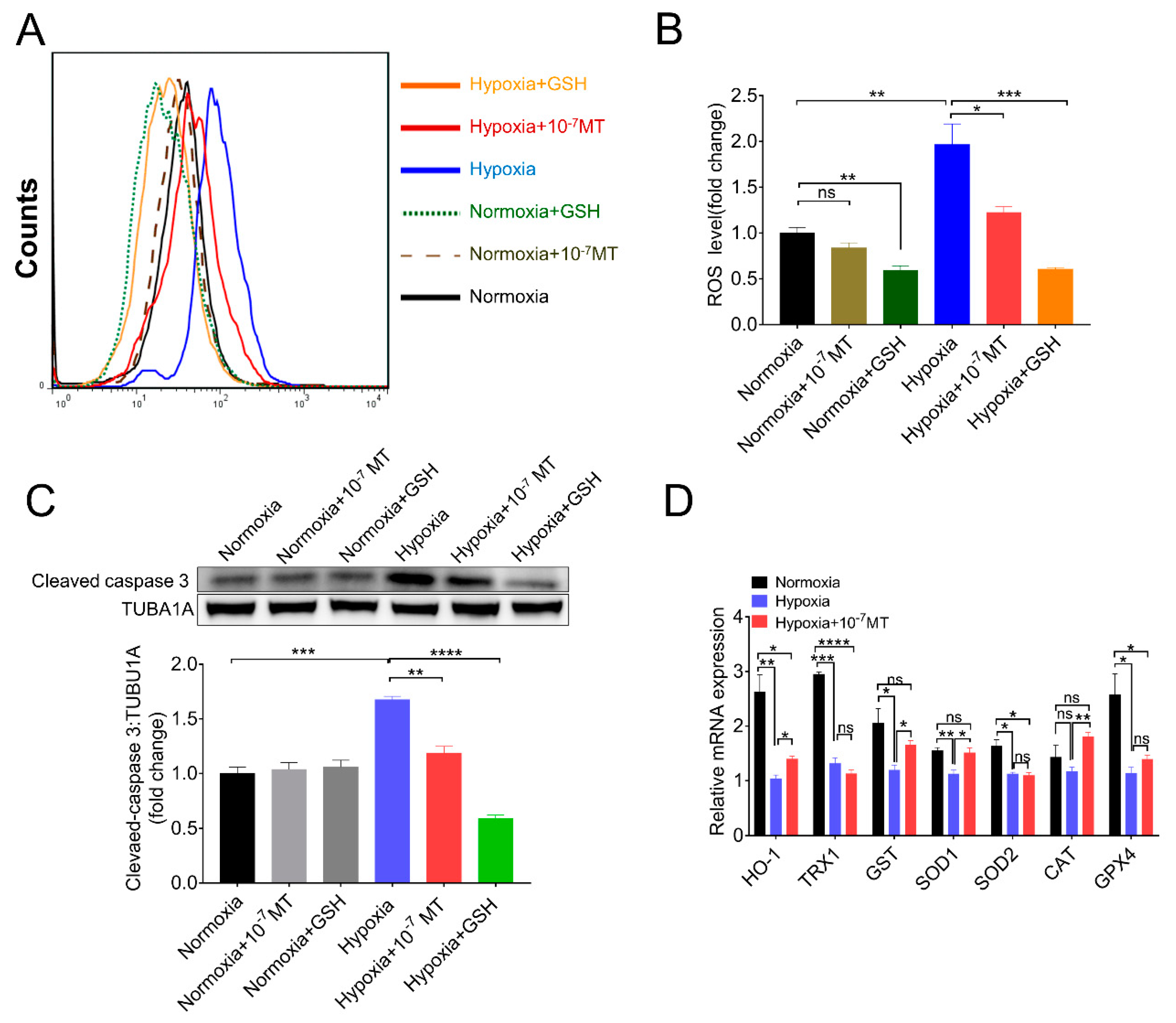
3.3. Melatonin Suppressed Hypoxia-Induced Apoptosis of Porcine GCs by Eliminating ROS and Increasing the Expression of Antioxidant Enzymes
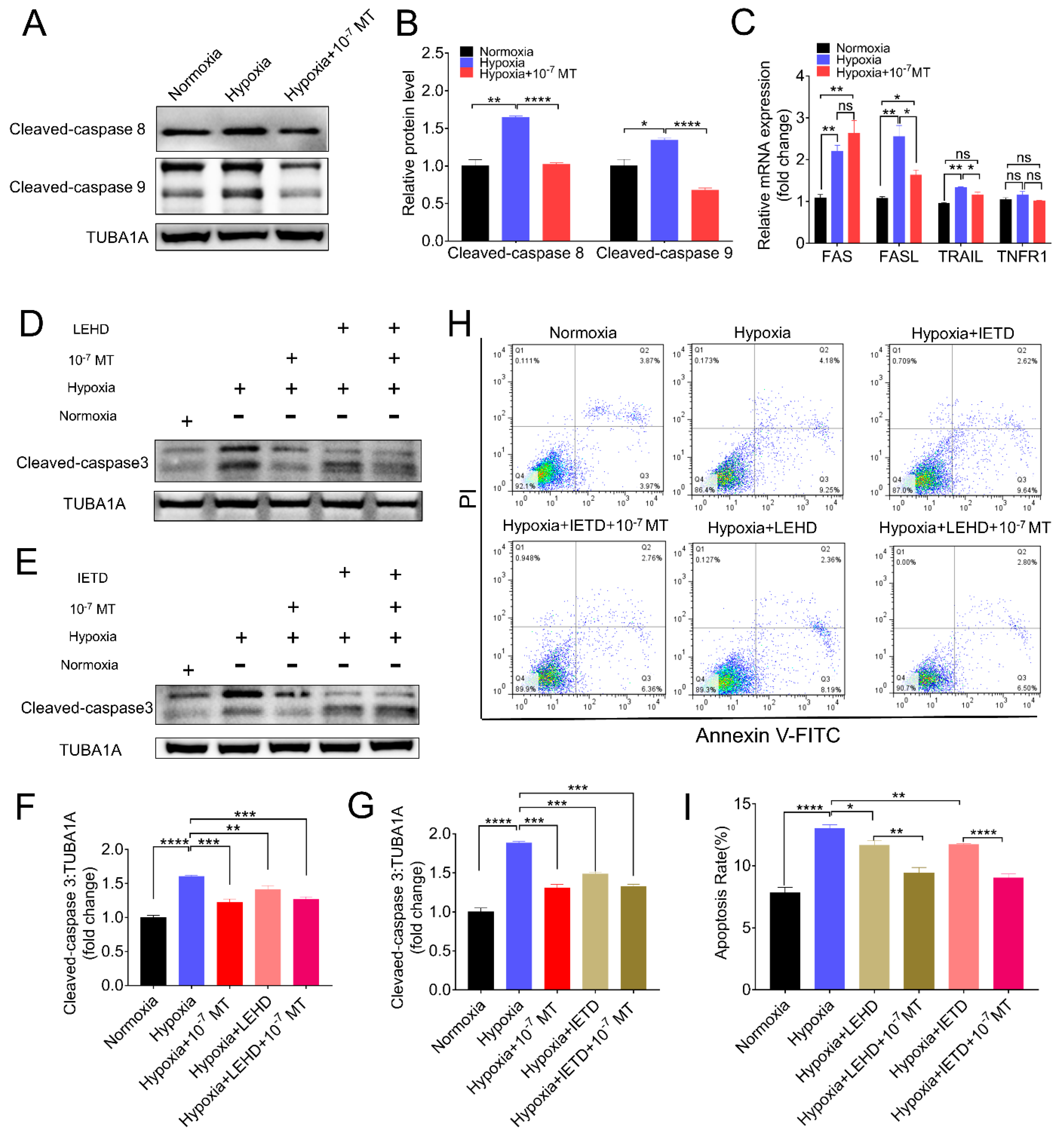
3.4. Melatonin Alleviated Hypoxia-Induced Apoptosis by Deactivating Caspase-8 and Caspase-9
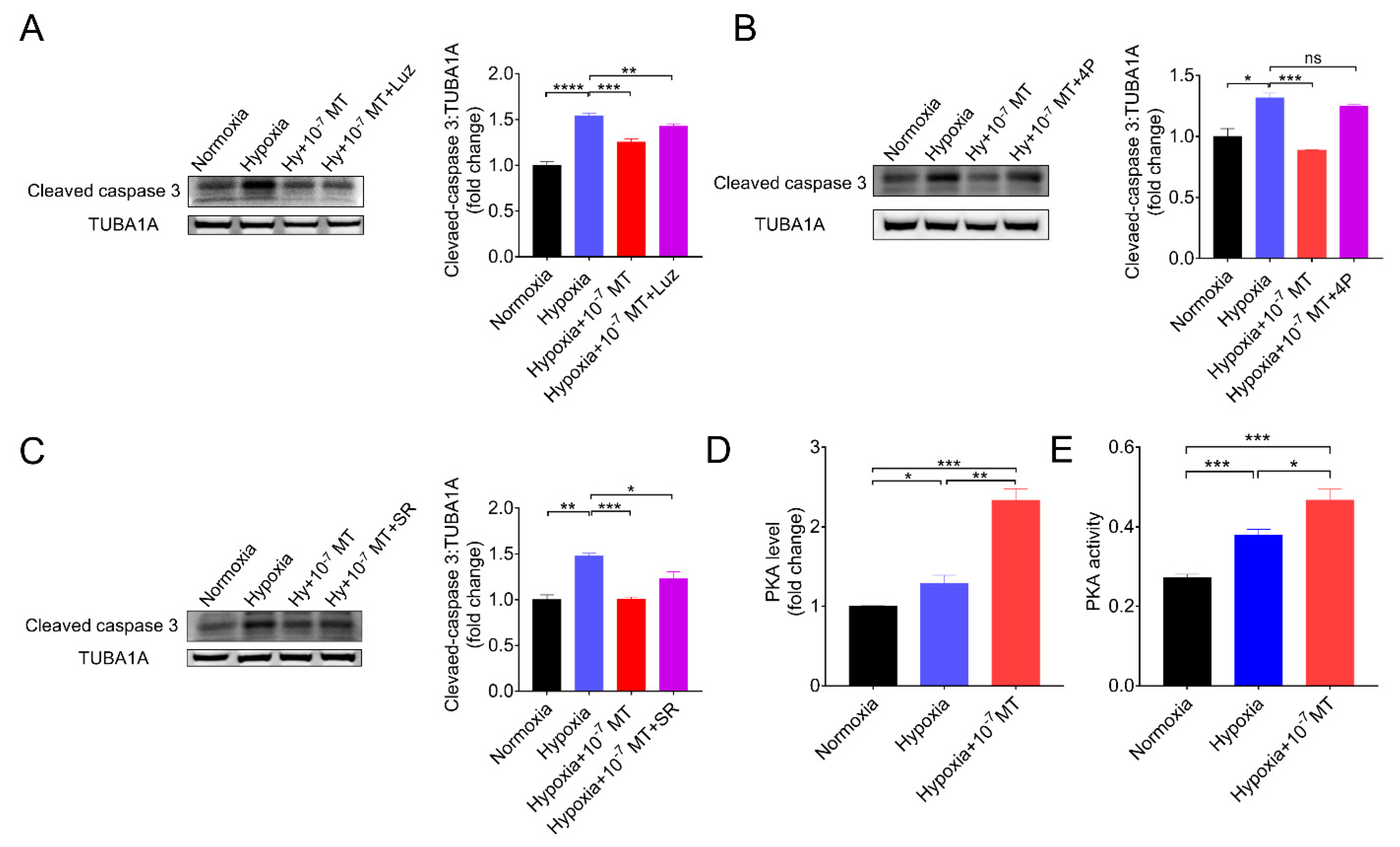
3.5. Melatonin Inhibited Hypoxia-Induced Apoptosis via MTNR1B-PKA Pathway
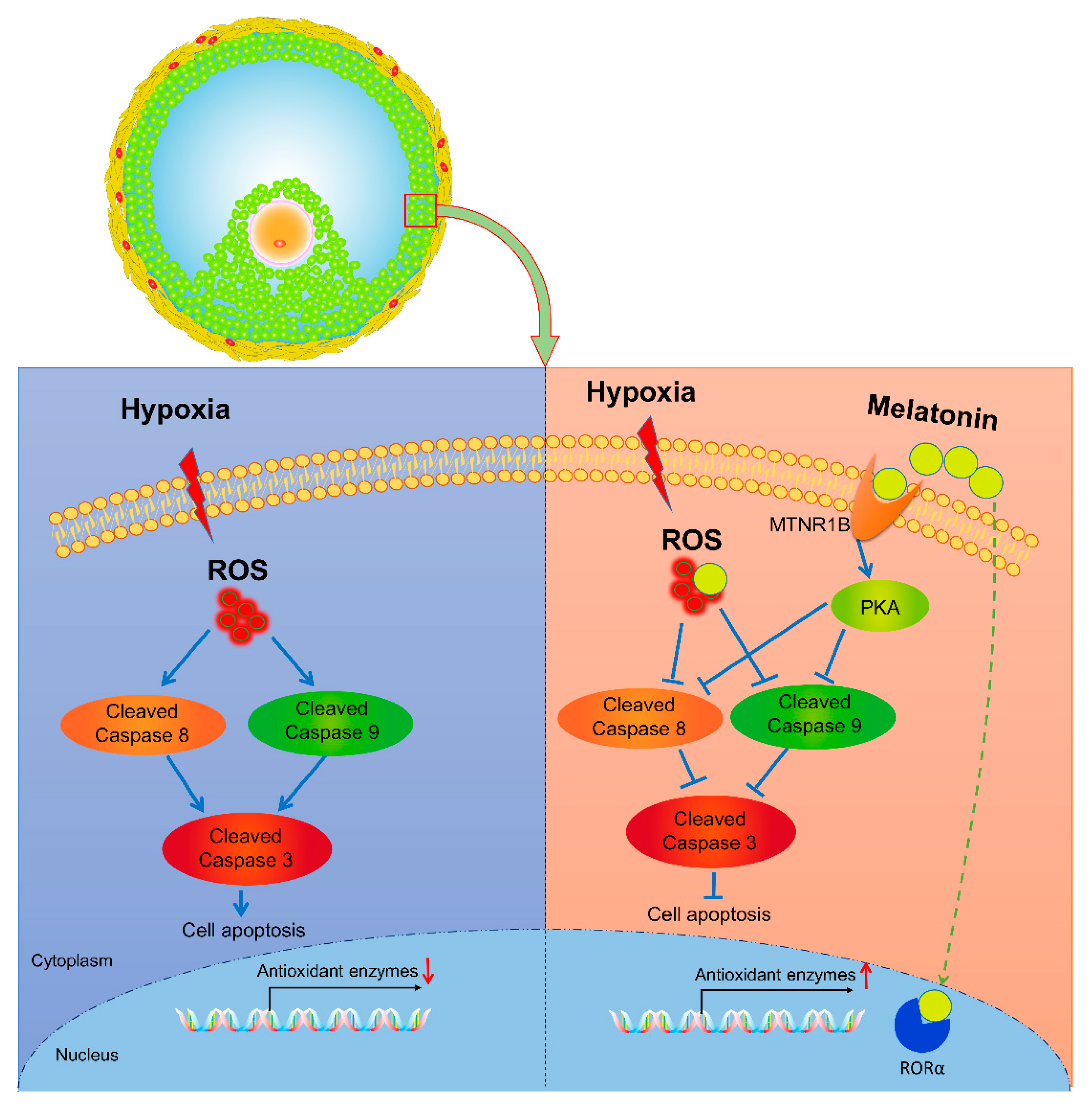
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kerr, J.B.; Myers, M.; Anderson, R.A. The dynamics of the primordial follicle reserve. Reproduction 2013, 146, R205–R215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Liu, Y.; Yao, W.; Li, Q.; Liu, H.; Pan, Z. Initiation of follicular atresia: Gene networks during early atresia in pig ovaries. Reproduction 2018, 156, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaipia, A.; Hsueh, A.J. Regulation of ovarian follicle atresia. Annu. Rev. Physiol. 1997, 59, 349–363. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.Y.; Cheung, C.K.; Wang, Y.; Tsang, B.K. Regulation of cell death and cell survival gene expression during ovarian follicular development and atresia. Front. Biosci. 2003, 8, d222–d237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilly, J.L.; Kowalski, K.I.; Johnson, A.L.; Hsueh, A.J. Involvement of apoptosis in ovarian follicular atresia and postovulatory regression. Endocrinology 1991, 129, 2799–2801. [Google Scholar] [CrossRef]
- Jolly, P.D.; Tisdall, D.J.; Heath, D.A.; Lun, S.; McNatty, K.P. Apoptosis in bovine granulosa cells in relation to steroid synthesis, cyclic adenosine 3′,5′-monophosphate response to follicle-stimulating hormone and luteinizing hormone, and follicular atresia. Biol. Reprod. 1994, 51, 934–944. [Google Scholar] [CrossRef] [Green Version]
- McRae, R.S.; Johnston, H.M.; Mihm, M.; O’Shaughnessy, P.J. Changes in mouse granulosa cell gene expression during early luteinization. Endocrinology 2005, 146, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Hickey, T.E.; Marrocco, D.L.; Gilchrist, R.B.; Norman, R.J.; Armstrong, D.T. Interactions between androgen and growth factors in granulosa cell subtypes of porcine antral follicles. Biol. Reprod. 2004, 71, 45–52. [Google Scholar] [CrossRef]
- Murdoch, W.J. Programmed cell death in preovulatory ovine follicles. Biol. Reprod. 1995, 53, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Basini, G.; Bianco, F.; Grasselli, F.; Tirelli, M.; Bussolati, S.; Tamanini, C. The effects of reduced oxygen tension on swine granulosa cell. Regul. Pept. 2004, 120, 69–75. [Google Scholar] [CrossRef]
- Lopez-Barneo, J.; Pardal, R.; Ortega-Saenz, P. Cellular mechanism of oxygen sensing. Annu. Rev. Physiol. 2001, 63, 259–287. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Choi, A.; Yu, H.Y.; Czerniak, S.M.; Holick, E.A.; Paolella, L.J.; Agarwal, A.; Combelles, C.M. Fluctuations in total antioxidant capacity, catalase activity and hydrogen peroxide levels of follicular fluid during bovine folliculogenesis. Reprod. Fertil. Dev. 2011, 23, 673–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Chattopadhyay, R.; Chakravarty, B.; Chaudhury, K. Markers of oxidative stress in follicular fluid of women with endometriosis and tubal infertility undergoing IVF. Reprod. Toxicol. 2013, 42, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Matzuk, M.M.; Dionne, L.; Guo, Q.; Kumar, T.R.; Lebovitz, R.M. Ovarian function in superoxide dismutase 1 and 2 knockout mice. Endocrinology 1998, 139, 4008–4011. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Nakamura, B.N.; Mohar, I.; Kavanagh, T.J.; Luderer, U. Glutamate Cysteine Ligase Modifier Subunit (Gclm) Null Mice Have Increased Ovarian Oxidative Stress and Accelerated Age-Related Ovarian Failure. Endocrinology 2015, 156, 3329–3343. [Google Scholar] [CrossRef]
- Ortega-Camarillo, C.; Gonzalez-Gonzalez, A.; Vergara-Onofre, M.; Gonzalez-Padilla, E.; Avalos-Rodriguez, A.; Gutierrez-Rodriguez, M.E.; Arriaga-Pizano, L.; Cruz, M.; Baiza-Gutman, L.A.; Diaz-Flores, M. Changes in the glucose-6-phosphate dehydrogenase activity in granulosa cells during follicular atresia in ewes. Reproduction 2009, 137, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Lin, F.; Zhang, J.; Tang, Y.; Chen, W.K.; Liu, H. Involvement of the up-regulated FoxO1 expression in follicular granulosa cell apoptosis induced by oxidative stress. J. Biol. Chem. 2012, 287, 25727–25740. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Q.; Shen, M.; Zhu, C.C.; Yu, F.X.; Liu, Z.Q.; Ally, N.; Sun, S.C.; Li, K.; Liu, H.L. 3-Nitropropionic acid induces ovarian oxidative stress and impairs follicle in mouse. PLoS ONE 2014, 9, e86589. [Google Scholar] [CrossRef]
- Tilly, J.L.; Tilly, K.I.; Kenton, M.L.; Johnson, A.L. Expression of members of the bcl-2 gene family in the immature rat ovary: Equine chorionic gonadotropin-mediated inhibition of granulosa cell apoptosis is associated with decreased bax and constitutive bcl-2 and bcl-xlong messenger ribonucleic acid levels. Endocrinology 1995, 136, 232–241. [Google Scholar] [CrossRef]
- Gupta, R.K.; Miller, K.P.; Babus, J.K.; Flaws, J.A. Methoxychlor inhibits growth and induces atresia of antral follicles through an oxidative stress pathway. Toxicol. Sci. 2006, 93, 382–389. [Google Scholar] [CrossRef] [Green Version]
- Brzezinski, A.; Seibel, M.M.; Lynch, H.J.; Deng, M.H.; Wurtman, R.J. Melatonin in human preovulatory follicular fluid. J. Clin. Endocrinol. Metab. 1987, 64, 865–867. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Nakamura, Y.; Korkmaz, A.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and the ovary: Physiological and pathophysiological implications. Fertil. Steril. 2009, 92, 328–343. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.M.; Tian, X.Z.; Zhou, G.B.; Wang, L.; Gao, C.; Zhu, S.E.; Zeng, S.M.; Tian, J.H.; Liu, G.S. Melatonin exists in porcine follicular fluid and improves in vitro maturation and parthenogenetic development of porcine oocytes. J. Pineal Res. 2009, 47, 318–323. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Deng, H.; Jiang, Z.; Li, Q.; Shi, M.; Chen, H.; Han, Z. Effects of melatonin on follicular atresia and granulosa cell apoptosis in the porcine. Mol. Reprod. Dev. 2016, 83, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.H.; Leal, C.L.; Da, C.J.; Tan, D.X.; Reiter, R.J. Role of melatonin on production and preservation of gametes and embryos: A brief review. Anim. Reprod. Sci. 2014, 145, 150–160. [Google Scholar] [CrossRef]
- Zhao, X.M.; Hao, H.S.; Du, W.H.; Zhao, S.J.; Wang, H.Y.; Wang, N.; Wang, D.; Liu, Y.; Qin, T.; Zhu, H.B. Melatonin inhibits apoptosis and improves the developmental potential of vitrified bovine oocytes. J. Pineal Res. 2016, 60, 132–141. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.; He, C.; Zhu, K.; Xu, Z.; Ma, T.; Tao, J.; Liu, G. Melatonin protects porcine oocyte in vitro maturation from heat stress. J. Pineal Res. 2015, 59, 365–375. [Google Scholar] [CrossRef]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Asghari, M.H.; Abdollahi, M.; de Oliveira, M.R.; Nabavi, S.M. A review of the protective role of melatonin during phosphine-induced cardiotoxicity: Focus on mitochondrial dysfunction, oxidative stress and apoptosis. J. Pharm. Pharmacol. 2017, 69, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhou, J.; Liu, Z.; Zhou, J.; Yao, W.; Tao, J.; Shen, M.; Liu, H. FSH prevents porcine granulosa cells from hypoxia-induced apoptosis via activating mitophagy through the HIF-1alpha-PINK1-Parkin pathway. FASEB J. 2020, 34, 3631–3645. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, C.; Wu, G.; Li, W.; Zhang, X.; Zhou, J.; Zhang, L.; Tao, J.; Shen, M.; Liu, H. Involvement of JNK/FOXO1 pathway in apoptosis induced by severe hypoxia in porcine granulosa cells. Theriogenology 2020, 154, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Allegra, M.; Reiter, R.J.; Tan, D.X.; Gentile, C.; Tesoriere, L.; Livrea, M.A. The chemistry of melatonin’s interaction with reactive species. J. Pineal Res. 2003, 34, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef]
- Venegas, C.; García, J.A.; Escames, G.; Ortiz, F.; López, A.; Doerrier, C.; García-Corzo, L.; López, L.C.; Reiter, R.J.; Acuña-Castroviejo, D. Extrapineal melatonin: Analysis of its subcellular distribution and daily fluctuations. J. Pineal Res. 2012, 52, 217–227. [Google Scholar] [CrossRef]
- Sakaguchi, K.; Itoh, M.T.; Takahashi, N.; Tarumi, W.; Ishizuka, B. The rat oocyte synthesises melatonin. Reprod. Fertil. Dev. 2013, 25, 674. [Google Scholar] [CrossRef]
- El-Raey, M.; Geshi, M.; Somfai, T.; Kaneda, M.; Hirako, M.; Abdel-Ghaffar, A.E.; Sosa, G.A.; El-Roos, M.E.; Nagai, T. Evidence of melatonin synthesis in the cumulus oocyte complexes and its role in enhancing oocyte maturation in vitro in cattle. Mol. Reprod. Dev. 2011, 78, 250–262. [Google Scholar] [CrossRef]
- Amireault, P.; Dubé, F. Serotonin and Its Antidepressant-Sensitive Transport in Mouse Cumulus-Oocyte Complexes and Early Embryos1. Biol. Reprod. 2005, 73, 358–365. [Google Scholar] [CrossRef]
- Muller, F.L.; Lustgarten, M.S.; Jang, Y.; Richardson, A.; Van Remmen, H. Trends in oxidative aging theories. Free Radic. Bio. Med. 2007, 43, 477–503. [Google Scholar] [CrossRef]
- Elmorsy, E.; Al-Ghafari, A.; Aggour, A.M.; Khan, R.; Amer, S. The role of oxidative stress in antipsychotics induced ovarian toxicity. Toxicol. In Vitro 2017, 44, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, E.R. Importance of individuality in oxidative stress and aging. Free Radic. Biol. Med. 2002, 33, 597–604. [Google Scholar] [CrossRef]
- Malpaux, B.; Thiery, J.C.; Chemineau, P. Melatonin and the seasonal control of reproduction. Reprod. Nutr. Dev. 1999, 39, 355–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olcese, J. The mammalian pineal gland and reproduction. Controversies and strategies for future research. Adv. Exp. Med. Biol. 1995, 377, 1–14. [Google Scholar] [CrossRef]
- Badness, T.J.; Powers, J.B.; Hastings, M.H.; Bittman, E.L.; Goldman, B.D. The timed infusion paradigm for melatonin delivery: What has it taught us about the melatonin signal, its reception, and the photoperiodic control of seasonal responses? J. Pineal Res. 1993, 15, 161–190. [Google Scholar] [CrossRef]
- ViviD, D.; Bentley, G.E.; Dax, V.; George, B. Seasonal Reproduction in Vertebrates: Melatonin Synthesis, Binding, and Functionality Using Tinbergen’s Four Questions. Molecules 2018, 23, 652. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, R.A.; Reiter, R.J. Pineal Gland: Influence on Gonads of Male Hamsters. Science 1965, 148, 1609–1611. [Google Scholar] [CrossRef]
- Luo, G.P.; Jian, Z.; Ma, R.Y.; Cao, Z.Z.; Zhu, Y.; Zhu, Y.; Tang, F.Q.; Xiao, Y.B. Melatonin alleviates hypoxia-induced cardiac apoptosis through PI3K/Akt pathway. Int. J. Clin. Exp. Pathol. 2018, 11, 5840–5849. [Google Scholar]
- Zhao, S.; Wang, Y.; Zhang, X.; Zheng, L.; Zhu, B.; Yao, S.; Yang, L.; Du, J. Melatonin Protects against Hypoxia/Reoxygenation-Induced Dysfunction of Human Umbilical Vein Endothelial Cells through Inhibiting Reactive Oxygen Species Generation. Acta Cardiol. Sin. 2018, 34, 424–431. [Google Scholar] [CrossRef]
- Zhao, X.M.; Wang, N.; Hao, H.S.; Li, C.Y.; Zhao, Y.H.; Yan, C.L.; Wang, H.Y.; Du, W.H.; Wang, D.; Liu, Y.; et al. Melatonin improves the fertilization capacity and developmental ability of bovine oocytes by regulating cytoplasmic maturation events. J. Pineal Res. 2018, 64. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, M.; Tamura, H.; Taketani, T.; Okada, M.; Lee, L.; Tamura, I.; Maekawa, R.; Asada, H.; Yamagata, Y.; Sugino, N. Melatonin protects the integrity of granulosa cells by reducing oxidative stress in nuclei, mitochondria, and plasma membranes in mice. J. Reprod. Dev. 2015, 61, 35–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Yang, Y.; Li, W.; Ao, H.; Zhang, Y.; Zhou, R.; Li, K. Effects of melatonin on the synthesis of estradiol and gene expression in pig granulosa cells. J. Pineal Res. 2019, 66, e12546. [Google Scholar] [CrossRef] [PubMed]
- Janes, K.A. An analysis of critical factors for quantitative immunoblotting. Sci. Signal. 2015, 8, s2. [Google Scholar] [CrossRef] [Green Version]
- Butler, T.A.J.; Paul, J.W.; Chan, E.; Smith, R.; Tolosa, J.M. Misleading Westerns: Common Quantification Mistakes in Western Blot Densitometry and Proposed Corrective Measures. Biomed. Res. Int. 2019, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L.; Prabhakar, N.R. The role of hypoxia-inducible factors in carotid body (patho) physiology. J. Physiol. 2018, 596, 2977–2983. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.A.; Waypa, G.B.; Schumacker, P.T. Redox signaling during hypoxia in mammalian cells. Redox Biol. 2017, 13, 228–234. [Google Scholar] [CrossRef]
- Coimbra-Costa, D.; Alva, N.; Duran, M.; Carbonell, T.; Rama, R. Oxidative stress and apoptosis after acute respiratory hypoxia and reoxygenation in rat brain. Redox Biol. 2017, 12, 216–225. [Google Scholar] [CrossRef]
- Niizuma, K.; Yoshioka, H.; Chen, H.; Kim, G.S.; Jung, J.E.; Katsu, M.; Okami, N.; Chan, P.H. Mitochondrial and apoptotic neuronal death signaling pathways in cerebral ischemia. Biochim. Biophys. Acta 2010, 1802, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Lochner, A.; Marais, E.; Huisamen, B. Melatonin and cardioprotection against ischaemia/reperfusion injury: What’s new? A review. J. Pineal Res. 2018, 65, e12490. [Google Scholar] [CrossRef] [Green Version]
- Ferlazzo, N.; Andolina, G.; Cannata, A.; Costanzo, M.G.; Rizzo, V.; Curro, M.; Ientile, R.; Caccamo, D. Is Melatonin the Cornucopia of the 21st Century? Antioxidants 2020, 9, 1088. [Google Scholar] [CrossRef]
- He, Y.M.; Deng, H.H.; Shi, M.H.; Bodinga, B.M.; Chen, H.L.; Han, Z.S.; Jiang, Z.L.; Li, Q.W. Melatonin modulates the functions of porcine granulosa cells via its membrane receptor MT2 in vitro. Anim. Reprod. Sci. 2016, 172, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Liu, W.; Pang, X.; Dai, S.; Liu, G. The Mechanism of Melatonin and Its Receptor MT2 Involved in the Development of Bovine Granulosa Cells. Int. J. Mol. Sci. 2018, 19, 2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.; Wang, Y.; Chen, J.; Zhang, J.; Yu, P.; Zhang, R.; Li, S.; Tao, B.; Wang, Y.; Qiu, Y.; et al. Activation of melatonin receptor 2 but not melatonin receptor 1 mediates melatonin-conferred cardioprotection against myocardial ischemia/reperfusion injury. J. Pineal Res. 2019, 67, e12571. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zeng, S. Melatonin Promotes Ubiquitination of Phosphorylated Pro-Apoptotic Protein Bcl-2-Interacting Mediator of Cell Death-Extra Long (BimEL) in Porcine Granulosa Cells. Int. J. Mol. Sci. 2018, 19, 3431. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequence (5′–3′) | Product Size (bp) | Accession No. |
|---|---|---|---|
| GAPDH | Forward: GTCGGTTGTGGATCTGACCT | 207 | XM_021091114.1 |
| Reverse: TTGACGAAGTGGTCGTTGAG | |||
| ACTB | Forward: TGCGGGACATCAAGGAGAAGC | 273 | XM_021086047.1 |
| Reverse: ACAGCACCGTGTTGGCGTAGAG | |||
| HO-1 | Forward: CCGAGAAGGCTTTAAGCTGGT | 110 | NM_001004027.1 |
| Reverse: GGAAGTAGAGGGGCGTGTAG | |||
| TRX1 | Forward: CTTTACCTTATTGCCCGGGT | 162 | NM_214313.2 |
| Reverse: GTTCACCGATTTTGTTGGCC | |||
| GST | Forward: CAACCCAGAAGACTGCTCAAG | 159 | NM_214300.1 |
| Reverse: GGACCACTCAAGGAATACAGAAG | |||
| SOD1 | Forward: TCCATGTCCATCAGTTTGGA | 248 | NM_001190422.1 |
| Reverse: AGTCACATTGGCCCAGGTCTC | |||
| SOD2 | Forward: AGGCGCTGAAAAAGGGTGAT | 163 | NM_214127.2 |
| Reverse: AAGTCGCGTTTGATGGCTTC | |||
| CAT | Forward: CACACATACCCATTCGTCACT | 157 | NM_214301.2 |
| Reverse: CAGCCCTAACCTTCACTTACC | |||
| GPX4 | Forward: ATTCTCAGCCAAGGACATCG | 93 | NM_214407.1 |
| Reverse: CCTCATTGAGAGGCCACATT | |||
| FAS | Forward: CCTGCCATCCCTGATGCTATT | 156 | XM_021072073.1 |
| Reverse: AGCGATACAGGTAAATACATGGC | |||
| FASL | Forward: GAGAGTCTGCCAGCCAAAGG | 125 | NM_213806.1 |
| Reverse: TCTTGAGTTAGGCTTGCCTGTT | |||
| TNFR1 | Forward: ACCGGGAGAAGAGAGAGAGTT | 102 | NM_213969.1 |
| Reverse: TGTGTAGGTAGGTGCCTTTGTG | |||
| TRAIL | Forward: GGGCAGACCTGTGTGTTGAT | 114 | NM_001024696.1 |
| Reverse: GGAGTACTTGTCCTGCATCTGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, J.-L.; Zhang, X.; Zhou, J.-Q.; Li, C.-Y.; Yang, M.-H.; Liu, Z.-J.; Zhang, L.-L.; Deng, S.-L.; Zhang, L.; Shen, M.; et al. Melatonin Alleviates Hypoxia-Induced Apoptosis of Granulosa Cells by Reducing ROS and Activating MTNR1B–PKA–Caspase8/9 Pathway. Antioxidants 2021, 10, 184. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10020184
Tao J-L, Zhang X, Zhou J-Q, Li C-Y, Yang M-H, Liu Z-J, Zhang L-L, Deng S-L, Zhang L, Shen M, et al. Melatonin Alleviates Hypoxia-Induced Apoptosis of Granulosa Cells by Reducing ROS and Activating MTNR1B–PKA–Caspase8/9 Pathway. Antioxidants. 2021; 10(2):184. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10020184
Chicago/Turabian StyleTao, Jing-Li, Xuan Zhang, Jia-Qi Zhou, Cheng-Yu Li, Ming-Hui Yang, Zhao-Jun Liu, Liang-Liang Zhang, Shou-Long Deng, Lu Zhang, Ming Shen, and et al. 2021. "Melatonin Alleviates Hypoxia-Induced Apoptosis of Granulosa Cells by Reducing ROS and Activating MTNR1B–PKA–Caspase8/9 Pathway" Antioxidants 10, no. 2: 184. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10020184