
Hypoxia Enhances Oxidative Stress in Neutrophils from ZZ Alpha-1 Antitrypsin Deficiency Patients
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Anamnesis and Physical Examination
2.3. Isolation and Culture of Peripheral Blood Neutrophils
2.4. Priming and Stimulation of Neutrophils
2.5. Oxidative Stress Assessment
2.6. Expression of Antioxidant Enzymes Assessment
2.7. Statistical Analysis
3. Results
3.1. Anthropometric and Clinical Data

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | MM (n = 7) | MZ (n = 31) | SZ (n = 8) | ZZ (n = 15) | p-Value |
|---|---|---|---|---|---|
| Age (years) | 11.00 ± 5.41 | 7.61 ± 3.89 | 10.13 ± 1.25 | 6.69 ± 2.87 | 0.06 |
| Male/Female (%) | 43/57 | 52/48 | 75/25 | 54/46 | 0.69 |
| AAT (mg·dL−1) | 148.50 ± 18.56 | 85.27 ± 25.08 | 57.38 ± 8.89 | 26.22 ± 8.03 | <0.0001 |
| BMI (kg·m−2) | 16.00 ± 2.42 | 17.47 ± 2.48 | 17.24 ± 1.41 | 17.50 ± 3.43 | 0.99 |
| FEV1 % | - | 102.50 ± 14.01 | 107.9 ± 5.67 | 102.6 ± 12.47 | 0.57 |
| FVC % | - | 98.69 ± 9.18 | 100.1 ± 7.60 | 98.44 ± 16.53 | 0.59 |
| FEV1/FVC % | - | 90.86 ± 6.64 | 94.71 ± 6.69 | 91.79 ± 5.22 | 0.22 |
| AST (U·L−1) | 20.00 ± 4.61 | 29.79 ± 6.57 | 35.00 ± 1.00 | 35.20 ± 6.00 | 0.14 |
| ALT (U·L−1) | 23.17 ± 7.65 | 22.76 ± 9.25 | 22.14 ± 4.38 | 32.33 ± 11.02 | 0.11 |
| GGT (U·L−1) | 18.46 ± 2.75 | 15.8 ± 2.69 | 17.38 ± 3.42 | 19.00 ± 1.00 | 0.14 |
| Vitamin A (µg·dL−1) | 40.10 ± 12.57 | 34.83 ± 11.11 | 32.50 ± 7.09 | 39.80 ± 13.94 | 0.69 |
| Vitamin D (ng·ml−1) | 36.50 ± 10.88 | 27.43 ± 9.81 | 35.50 ± 7.27 | 32.40 ± 10.46 | 0.18 |
| Vitamin E (mg·L−1) | 9.25 ± 3.79 | 10.87 ± 9.73 | 10.40 ± 1.14 | 11.72 ± 2.05 | 0.14 |
3.2. Oxidative Stress Profile
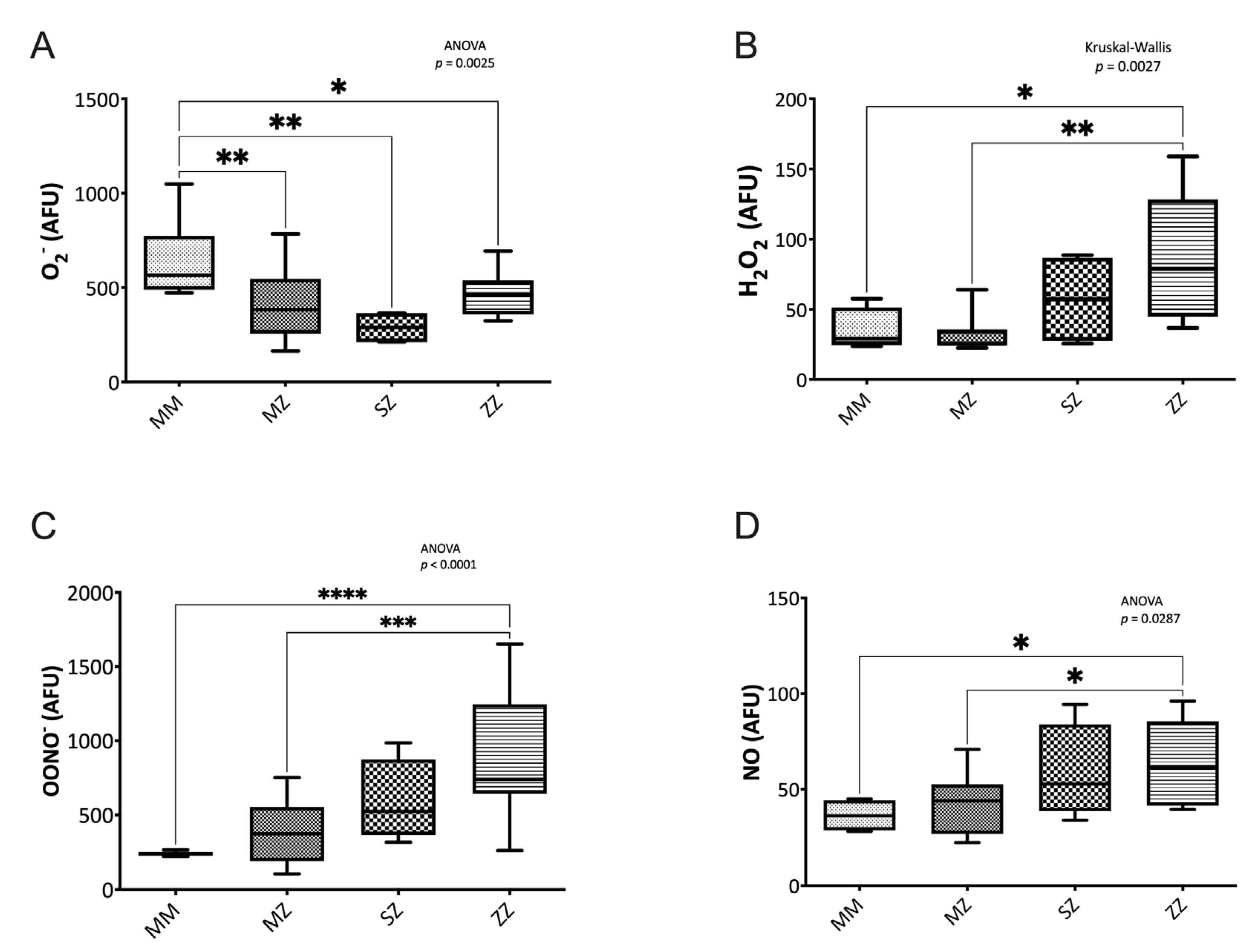
3.2.1. Nitrogen Species Assessment
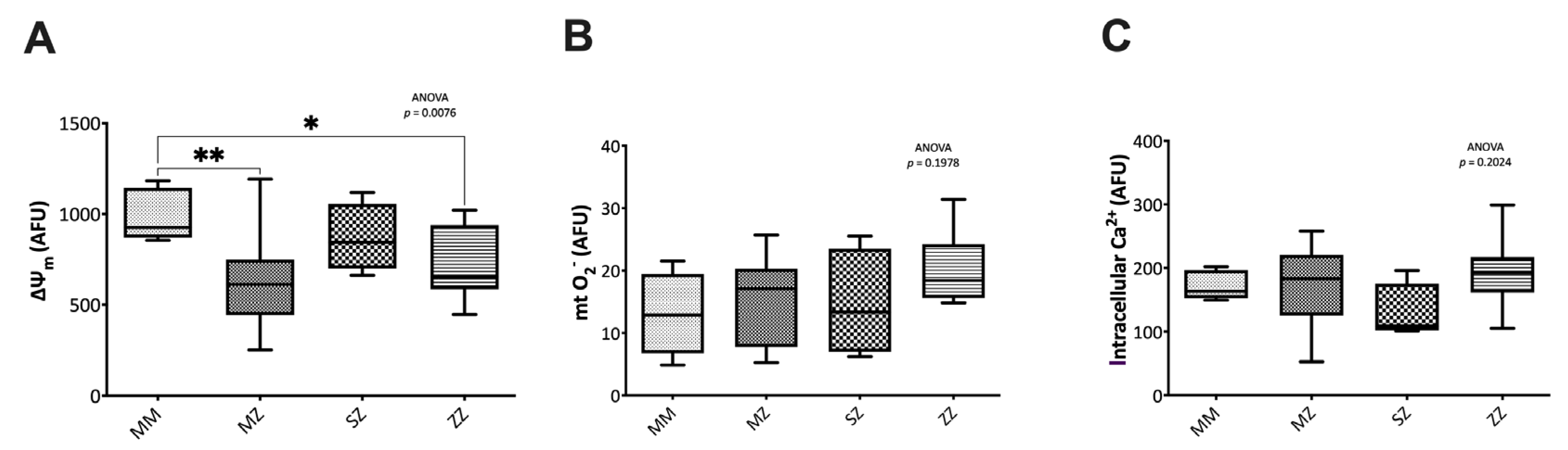
3.2.2. Mitochondrial Function and Intracellular Calcium Assessment
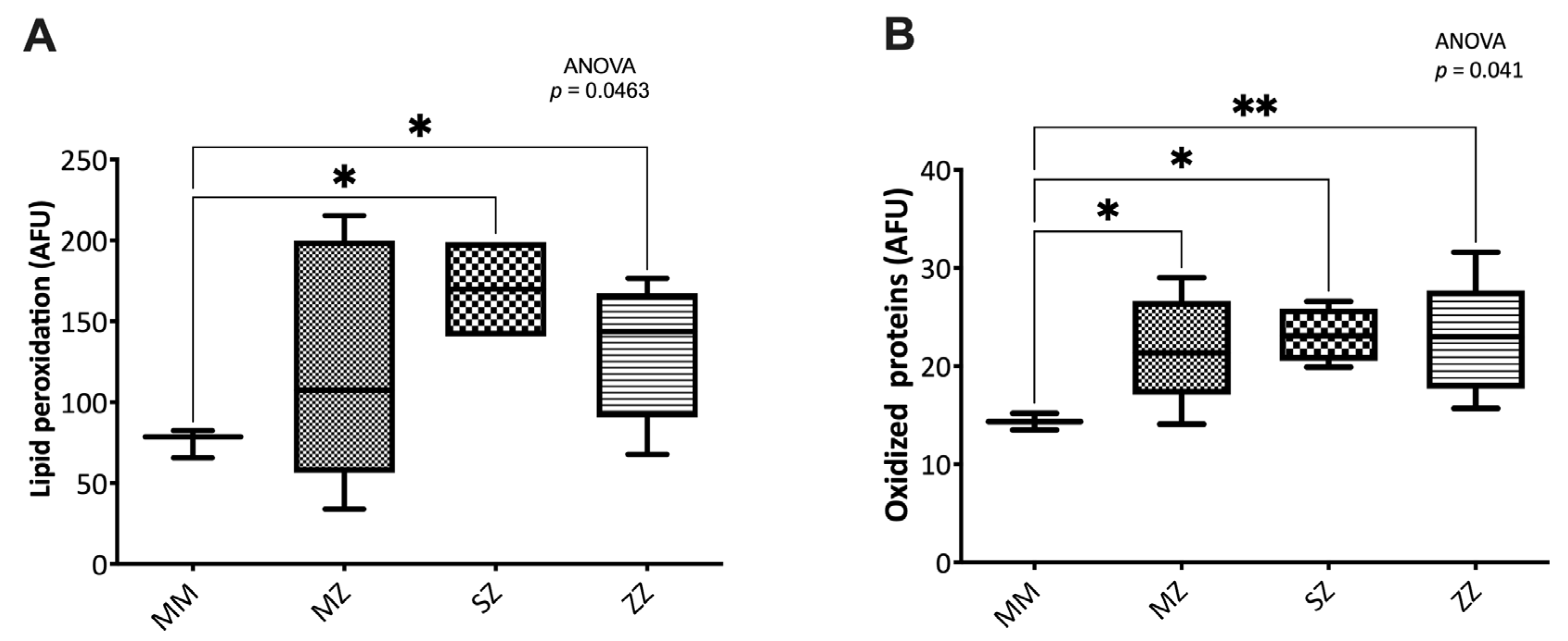
3.2.3. Oxidative Damage to Biomolecules
3.3. Antioxidant Defense Systems in AATD
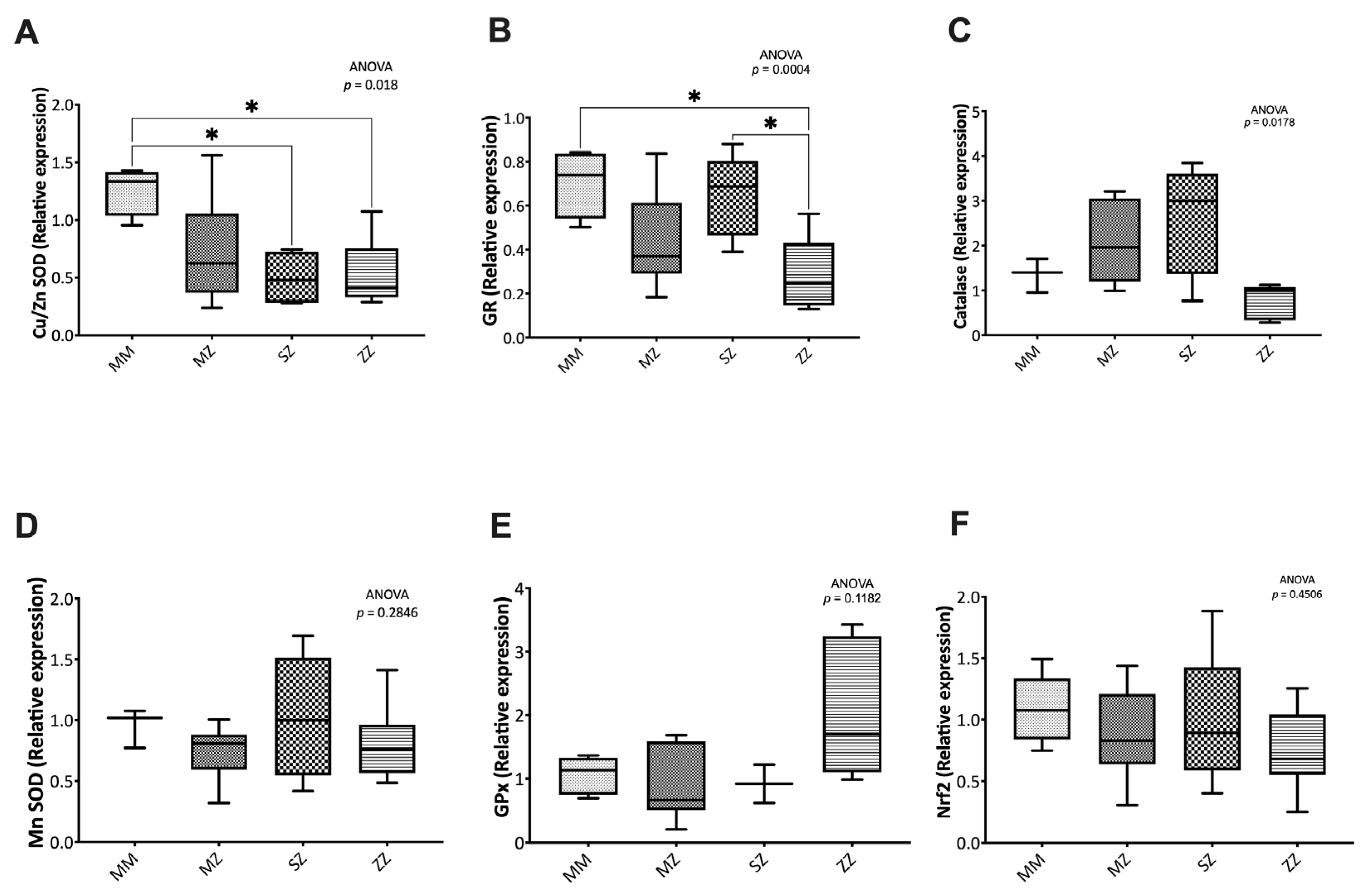
3.3.1. Expression of Antioxidant Enzymes
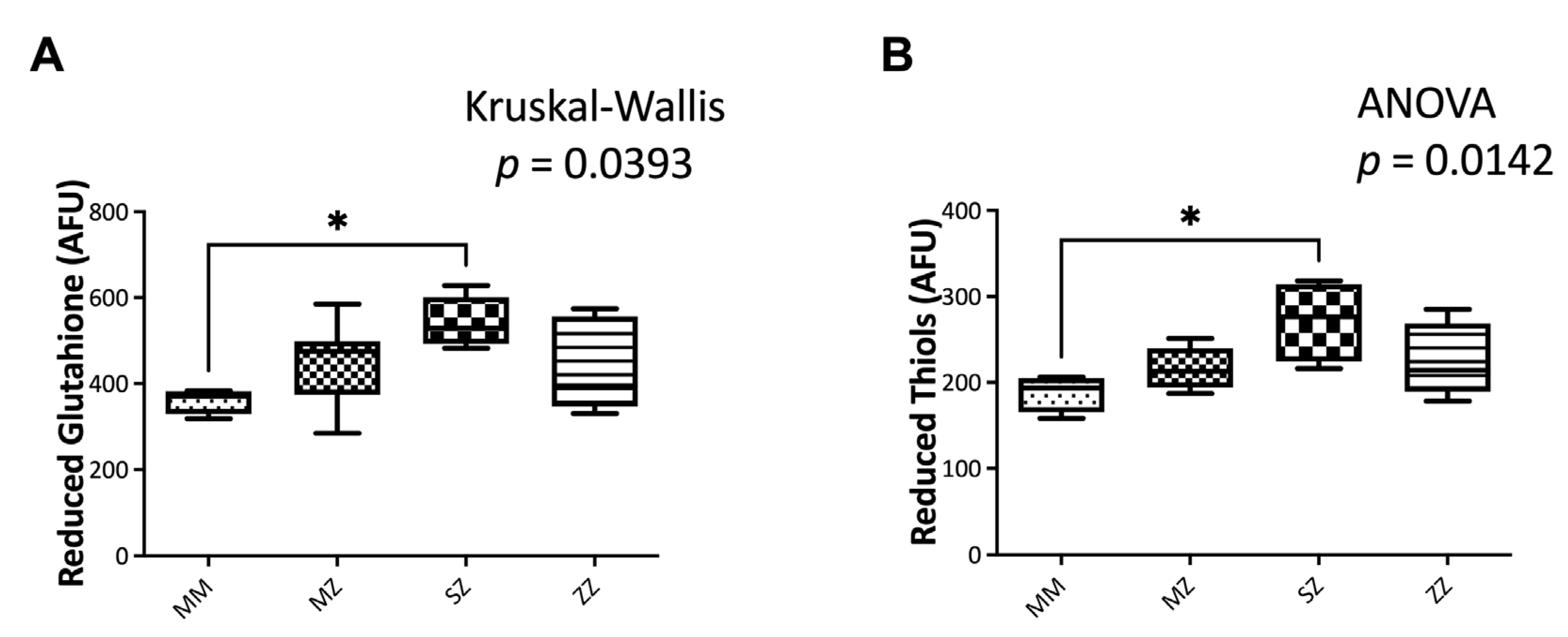
3.3.2. Non-Enzymatic Antioxidant Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torres-Durán, M.; Lopez-Campos, J.L.; Barrecheguren, M.; Miravitlles, M.; Martinez-Delgado, B.; Castillo, S.; Escribano, A.; Baloira, A.; Navarro-Garcia, M.M.; Pellicer, D.; et al. Alpha-1 Antitrypsin Deficiency: Outstanding Questions and Future Directions. Orphanet. J. Rare. Dis. 2018, 13, 114. [Google Scholar] [CrossRef] [Green Version]
- Strnad, P.; McElvaney, N.G.; Lomas, D.A. Alpha1-Antitrypsin Deficiency. N. Engl. J. Med. 2020, 382, 1443–1455. [Google Scholar] [CrossRef] [PubMed]
- Palacio, P.L.; Godoy, J.R.; Aktas, O.; Hanschmann, E.-M. Changing Perspectives from Oxidative Stress to Redox Signaling—Extracellular Redox Control in Translational Medicine. Antioxidants 2022, 11, 1181. [Google Scholar] [CrossRef]
- Belchamber, K.B.R.; Walker, E.M.; Stockley, R.A.; Sapey, E. Monocytes and Macrophages in Alpha-1 Antitrypsin Deficiency. Int. J. Chronic. Obstr. 2020, 15, 3183–3192. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, C.; Reeves, E.P.; McElvaney, N.G. The Role of Neutrophils in Alpha-1 Antitrypsin Deficiency. Ann. Am. Thorac. Soc. 2016, 13, S297–S304. [Google Scholar] [CrossRef]
- Williams, A.E.; Chambers, R.C. Neutrophils and Tissue Damage: Is Hypoxia the Key to Excessive Degranulation? Thorax 2016, 71, 977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stuart, J.A.; Fonseca, J.; Moradi, F.; Cunningham, C.; Seliman, B.; Worsfold, C.R.; Dolan, S.; Abando, J.; Maddalena, L.A. How Supraphysiological Oxygen Levels in Standard Cell Culture Affect Oxygen-Consuming Reactions. Oxid. Med. Cell. Longev. 2018, 2018, 8238459. [Google Scholar] [CrossRef] [Green Version]
- Alva, R.; Gardner, G.L.; Liang, P.; Stuart, J.A. Supraphysiological Oxygen Levels in Mammalian Cell Culture: Current State and Future Perspectives. Cells 2022, 11, 3123. [Google Scholar] [CrossRef]
- Miravitlles, M.; Dirksen, A.; Ferrarotti, I.; Koblizek, V.; Lange, P.; Mahadeva, R.; McElvaney, N.G.; Parr, D.; Piitulainen, E.; Roche, N.; et al. European Respiratory Society Statement: Diagnosis and Treatment of Pulmonary Disease in A1-Antitrypsin Deficiency. Eur. Respir. J. 2017, 50, 1700610. [Google Scholar] [CrossRef] [Green Version]
- Hoenderdos, K.; Lodge, K.M.; Hirst, R.A.; Chen, C.; Palazzo, S.G.C.; Emerenciana, A.; Summers, C.; Angyal, A.; Porter, L.; Juss, J.K.; et al. Hypoxia Upregulates Neutrophil Degranulation and Potential for Tissue Injury. Thorax 2016, 71, 1030–1038. [Google Scholar] [CrossRef] [Green Version]
- Beltrán, B.; Nos, P.; Dasí, F.; Iborra, M.; Bastida, G.; Martínez, M.; O’Connor, J.; Sáez, G.; Moret, I.; Ponce, J. Mitochondrial Dysfunction, Persistent Oxidative Damage, and Catalase Inhibition in Immune Cells of Naïve and Treated Crohn’s Disease. Inflamm. Bowel. Dis. 2010, 16, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Reula, A.; Pellicer, D.; Castillo, S.; Magallón, M.; Armengot, M.; Herrera, G.; O’Connor, J.-E.; Bañuls, L.; Navarro-García, M.M.; Escribano, A.; et al. New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry. J. Clin. Med. 2021, 10, 1172. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An Improvement of the 2ˆ(-Delta Delta CT) Method for Quantitative Real-Time Polymerase Chain Reaction Data Analysis. Biostat Bioinform Biomath. 2013, 3, 71–85. [Google Scholar]
- Manda-Handzlik, A.; Demkow, U. Neutrophils: The Role of Oxidative and Nitrosative Stress in Health and Disease. Adv. Exp. Med. Biol. 2015, 857, 51–60. [Google Scholar] [CrossRef]
- Janciauskiene, S.; Welte, T. Well-Known and Less Well-Known Functions of Alpha-1 Antitrypsin. Its Role in Chronic Obstructive Pulmonary Disease and Other Disease Developments. Ann. Am. Thorac. Soc. 2016, 13, S280–S288. [Google Scholar] [CrossRef]
- Ertel, W.; Morrison, M.H.; Ayala, A.; Chaudry, I.H. Hypoxemia in the Absence of Blood Loss or Significant Hypotension Causes Inflammatory Cytokine Release. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1995, 269, R160–R166. [Google Scholar] [CrossRef]
- Hartmann, G.; Tschöp, M.; Fischer, R.; Bidlingmaier, C.; Riepl, R.; Tschöp, K.; Hautmann, H.; Endres, S.; Toepfer, M. High altitude increases circulating interleukin-6, interleukin-1 receptor antagonist and C-reactive protein. Cytokine 2000, 12, 246–252. [Google Scholar] [CrossRef]
- Han, S.; Mallampalli, R.K. The Acute Respiratory Distress Syndrome: From Mechanism to Translation. J. Immunol. Baltim. Md. 1950 2015, 194, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Hoenderdos, K.; Condliffe, A. The Neutrophil in Chronic Obstructive Pulmonary Disease. Am. J. Resp. Cell. Mol. 2013, 48, 531–539. [Google Scholar] [CrossRef]
- Iannitti, R.G.; Casagrande, A.; Luca, A.D.; Cunha, C.; Sorci, G.; Riuzzi, F.; Borghi, M.; Galosi, C.; Massi-Benedetti, C.; Oury, T.D.; et al. Hypoxia Promotes Danger-Mediated Inflammation via Receptor for Advanced Glycation End Products in Cystic Fibrosis. Am. J. Resp. Crit. Care. 2013, 188, 1338–1350. [Google Scholar] [CrossRef]
- Thompson, A.A.R.; Binham, J.; Plant, T.; Whyte, M.K.B.; Walmsley, S.R. Hypoxia, the HIF Pathway and Neutrophilic Inflammatory Responses. BiolChem 2013, 394, 471–477. [Google Scholar] [CrossRef]
- Walmsley, S.R.; Print, C.; Farahi, N.; Peyssonnaux, C.; Johnson, R.S.; Cramer, T.; Sobolewski, A.; Condliffe, A.M.; Cowburn, A.S.; Johnson, N.; et al. Hypoxia-Induced Neutrophil Survival Is Mediated by HIF-1alpha-Dependent NF-KappaB Activity. J. Exp. Med. 2005, 201, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Beckman, J.S.; Koppenol, W.H. Nitric Oxide, Superoxide, and Peroxynitrite: The Good, the Bad, and Ugly. Am. J. Physiol-cell Ph. 1996, 271, C1424–C1437. [Google Scholar] [CrossRef] [Green Version]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papp, E.; Száiraz, P.; Korcsmáiros, T.; Csermely, P.; Papp, E.; Száiraz, P.; Korcsmáiros, T.; Csermely, P. Changes of Endoplasmic Reticulum Chaperone Complexes, Redox State, and Impaired Protein Disulfide Reductase Activity in Misfolding Ai-antitrypsin Transgenic Mice. Faseb. J. 2006, 20, 1018–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzone, G.R.; Liberona, L.F.; Bustamante, A.P.; Saez, C.G.; Olmos, P.R.; Vecchiola, A.; Villagrán, A.; Serrano, C.; Reyes, T.P. Differences in Lung Glutathione Metabolism May Account for Rodent Susceptibility in Elastase-Induced Emphysema Development. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2009, 296, R1113–R1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcus, N.Y.; Blomenkamp, K.; Ahmad, M.; Teckman, J.H. Oxidative Stress Contributes to Liver Damage in a Murine Model of Alpha-1-Antitrypsin Deficiency. Exp. Biol. Med. 2012, 237, 1163–1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escribano, A.; Amor, M.; Pastor, S.; Castillo, S.; Sanz, F.; Codoñer-Franch, P.; Dasí, F. Decreased Glutathione and Low Catalase Activity Contribute to Oxidative Stress in Children with α-1 Antitrypsin Deficiency. Thorax 2015, 70, 82. [Google Scholar] [CrossRef] [Green Version]
- Escribano, A.; Pastor, S.; Reula, A.; Castillo, S.; Vicente, S.; Sanz, F.; Casas, F.; Torres, M.; Fernández-Fabrellas, E.; Codoñer-Franch, P.; et al. Accelerated Telomere Attrition in Children and Teenagers with A1-Antitrypsin Deficiency. Eur. Respir. J. 2016, 48, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Dasi, F.; Pastor, S.; Reula, A.; Castillo, S.; Escribano, A. Augmentation Therapy Increases Hydrogen Peroxide Accumulation in Peripheral Blood Mononuclear Cells of AATD Patients. In Proceedings of the American Thoracic Society Annual Meeting, Washington, DC, USA, 19 May–24 May 2017; Volume 195, p. A6337. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magallón, M.; Castillo-Corullón, S.; Bañuls, L.; Pellicer, D.; Romero, T.; Martínez-Ferraro, C.; Navarro-García, M.M.; Herrejón, A.; González, C.; Dasí, F. Hypoxia Enhances Oxidative Stress in Neutrophils from ZZ Alpha-1 Antitrypsin Deficiency Patients. Antioxidants 2023, 12, 872. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12040872
Magallón M, Castillo-Corullón S, Bañuls L, Pellicer D, Romero T, Martínez-Ferraro C, Navarro-García MM, Herrejón A, González C, Dasí F. Hypoxia Enhances Oxidative Stress in Neutrophils from ZZ Alpha-1 Antitrypsin Deficiency Patients. Antioxidants. 2023; 12(4):872. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12040872
Chicago/Turabian StyleMagallón, María, Silvia Castillo-Corullón, Lucía Bañuls, Daniel Pellicer, Teresa Romero, Carlos Martínez-Ferraro, María Mercedes Navarro-García, Alberto Herrejón, Cruz González, and Francisco Dasí. 2023. "Hypoxia Enhances Oxidative Stress in Neutrophils from ZZ Alpha-1 Antitrypsin Deficiency Patients" Antioxidants 12, no. 4: 872. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox12040872