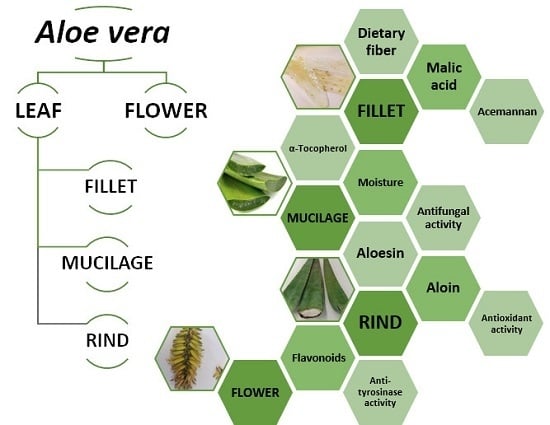
3.1. Nutritional Composition of Fillet
Aloe vera fillet has been used in the food industry to develop functional foods such as beverages, milk, yogurt, jam, jellies, ice cream, and food supplements, as well as in edible fruit coatings. It can also be used to improve the quality of meat products [
36] and is often commercialized as concentrated dry powder. In this study, the fillet sample corresponded to 58 ± 4% of the total leaf weight, while 31 ± 2% consisted of green rind.
The nutritional composition of
Aloe vera fillet is presented in
Table 1. This inner part of the leaf consists of 98 ± 1 g/100 g of moisture, the same amount that was found in the mucilage. Similar moisture contents (98–99 g/100 g) were previously reported [
5,
9,
37]. Lower values were found in the rind and flower samples (87 ± 1 and 84 ± 1, respectively;
Table S1).
Dietary fibre was a predominant macronutrient in
Aloe vera fillet, with 50.1 ± 0.3 g/100 g dw, followed by available carbohydrates, which corresponded to 37.4 ± 0.3 g/100 g dw (
Table 1). A slightly higher dietary fibre content (57.64 g/100 g dw) was described by Femenia et al. [
5] in fillet samples of
Aloe vera cultivated in Ibiza, Spain. In turn, the fraction corresponding to crude fibre was isolated through acid and alkaline digestion of the sample and may consist of cellulose and small amounts of hemicellulose and lignin [
38], which do not dissolve in the used solutions of sulphuric acid and potassium hydroxide. This small amount of insoluble fibre may correspond to the cell walls of the parenchyma cells that contain the gel. A higher crude fibre content (12.95 g/100 g dw) was previously reported in
Aloe vera samples from Coquimbo, Chile [
39].
The fresh fillet revealed reduced levels of protein and fat and slightly higher amounts of ash (minerals) (
Table 1). This sample was, therefore, characterized by a low energy value (269 ± 3 kcal/100 g dw). These values are lower than that previously reported for fat (4.21 g/100 g dw) and ash (15.37–17.64 g/100 g dw) but slightly higher for the protein content (3.72–7.26 g/100 g dw) [
5,
39]. Potassium and calcium were previously found as major minerals in fillet samples [
5] and may contribute to the wound healing capacity of this medicinal plant. Such compositional variations can be justified by the different geographical and edaphoclimatic conditions where the
Aloe samples were grown.
As shown in
Table 1, oxalic, quinic, and malic acids were detected in the fillet. Malic acid was the most abundant, with a concentration of 97 ± 1 mg/100 g of fresh fillet and 5.75 ± 0.07 g/100 g of dried powder. This acid is a natural component of aloe gel and an excellent freshness indicator. It was also detected along with the other two acids in the mucilage sample (
Table S1). In fact, it was more abundant in the mucilage collected from the vascularized layer of the leaf, but the amount of total organic acids found in fillet and mucilage did not differ significantly. Bozzi et al. [
9] also detected malic acid in fresh
Aloe vera gel and others, like citric, lactic, and succinic acids, in commercial gel powders. However, citric acid (which can be found in the rind;
Table S1) is added to the concentrated powders as a natural preservative by adjusting the gel pH prior to its concentration and drying in order to improve flavour and prevent oxidation. In turn, lactic and succinic acids should be absent from these concentrates, since they are indicators of bacterial fermentation and enzymatic degradation [
9]. As presented in
Table S1, ascorbic acid was not detected in the fillet and mucilage samples, but it was found in the green rind. Fumaric acid, in turn, was detected in rind and flower.
Tocopherols are important fat-soluble chain-breaking antioxidants. As shown in
Table 1, the four isoforms were detected in the fillet, and α-tocopherol (4.8 ± 0.1 mg/100 g dw) was the most abundant, followed by γ- and β-tocopherols. Therefore, a 100 g portion of dried fillet provides about 69%, 44%, and 32% of the recommended dietary allowances of vitamin E for children from 4–8 and 9–13 years old and individuals with 14 or more years old, respectively (values calculated based on the α-tocopherol content) [
40]. Bashipour and Ghoreishi [
41] obtained a lower amount (1.53 mg/100 g dw) of α-tocopherol from
Aloe vera samples grown in Isfahan, Iran, when applying optimized supercritical CO
2 extraction conditions. Comparable α-tocopherol levels (4.70 mg/100 g dw) were reported by López-Cervantes et al. [
42] in
Aloe vera flowers harvested in south Sonora, México.
The fatty acids composition of
Aloe vera fillet is presented in
Table 2. Eighteen fatty acids were detected, with predominance of palmitic (C16:0, 32.1 ± 0.6%), stearic (C18:0, 16.4 ± 0.2%), linoleic (C18:2n6, 15.0 ± 0.2%), and oleic (C18:1n9, 12.9 ± 0.1%) acids. Thus, 67 ± 1% of the lipid fraction is constituted by saturated fatty acids (SFA) and 32.9 ± 0.4% corresponds to mono- and polyunsaturated (MUFA and PUFA) fatty acids. Essential PUFA such as C18:2n6 and linolenic acid (C18:3n3) play important biological functions and are involved in the modulation of inflammatory and chronic degenerative diseases [
43]. Despite this, its concentration in the fillet is very low compared to other compounds identified with possible health-promoting effects. Odd-chain SFA, including pentadecanoic (C15:0) and heptadecanoic (C17:0) acids, were also found (
Table 2). These two fatty acids have been gaining research interest because they are important as quantitative internal standards and biomarkers for assessing dietary food intake and the risk of coronary heart disease and type II diabetes mellitus [
44]. A previous study also reported C18:2n6, C18:3n3, lauric acid (C12:0) and myristic acid (C14:0) in
Aloe vera gel [
45]. Regarding other
Aloe species, the fatty acids C18:2n6, C16:0, lignoceric (C24:0), C18:0, tricosanoic (C23:0), behenic (C22:0), and C18:3n3, among others, were described in
Aloe ferox gel [
46]; and C18:3n3, C18:2n6, C16:0, C18:0, C18:1n9, and C14:0 were found in a whole leaf extract of
Aloe arborescence [
47].
3.2. Sugars and Glycosidic-linkage Composition of Fillet
Table 3 shows that the fillet sample contained 64 g/100 g dw of sugars, mainly glucose, uronic acids, and mannose. In this sample, 35 g/100 g dw of the sugars were found in the free form, determined as glucose and mannose (
Table 1). The identification of the alditol acetate corresponding to mannose is probably due to the presence of fructose, as the methodology used converts fructose in glucitol (57%) and mannitol (43%) [
48]. These results show that fillet sample contained 8 g/100 g dw of fructose and 27 g/100 g dw of glucose. A previous study reported fructose and glucose contents ranging from 0.56 to 9.62 g/100 g dw and 4.57 to 28.27 g/100 g dw, respectively, in fresh gel and powdered gel concentrates of
Aloe vera [
9], thus comprising the value quantified in this study.
Upon dialysis (12–14 kDa cut off), the weight of the obtained high molecular weight (HMW) material accounted for 39.9%. The sample obtained after dialysis (HMW) was composed by 77% of sugars, mainly mannose (65%), although glucose was also present (16%). To disclose the polysaccharides composition of the HMW sample, a methylation analysis was performed (
Table 4).
The glycosidic-linkage composition of dialysed
Aloe vera fillet shows a mannan composed by a backbone of (β1→4)-mannose residues [
4,
6], as observed by the high amount of the (1→4)-linked mannose residues (74.0%). Although (β1→4)-glucose residues may be present as insertions of the mannan backbone [
49], these can have also indicate the presence of a glucan, possibly cellulose.
Aloe vera mannan is also reported to be acetylated at the C-2 and C-3 positions and containing some side chains, mainly of galactose attached to C-6 [
4,
6]. The analysis performed using alkali conditions does not allow to maintain the acetyl groups. Nevertheless, the presence of (1→2,4)- and (1→3,4)-linked mannose residues are probably resultant from resistant acetylation positions of the mannan (
Table 4). The branching percentage (3.2%), which can be estimated by the ratio between the (1→4,6)- mannose and the total amount of mannose, is in accordance with the presence of terminally-linked galactose and arabinose residues, identified in both pyranose (0.5%) and furanose (0.4%) forms [
6]. The ratio calculated by the relative amount of total mannose divided by the amount of terminally linked mannose shows that this polysaccharide had a higher molecular weight than those previously reported [
6].
3.3. Phenolic Composition of Fillet, Mucilage, Rind, and Flower
Data related to the phenolic compounds identification in the obtained
Aloe vera extracts are presented in
Table 5, namely the retention time, λ
max in the UV-vis region, pseudomolecular ion, ions of major fragments in MS
2, and tentative identification (the obtained extraction yields and the phenolic contents that can be found in fresh and dried samples are shown in
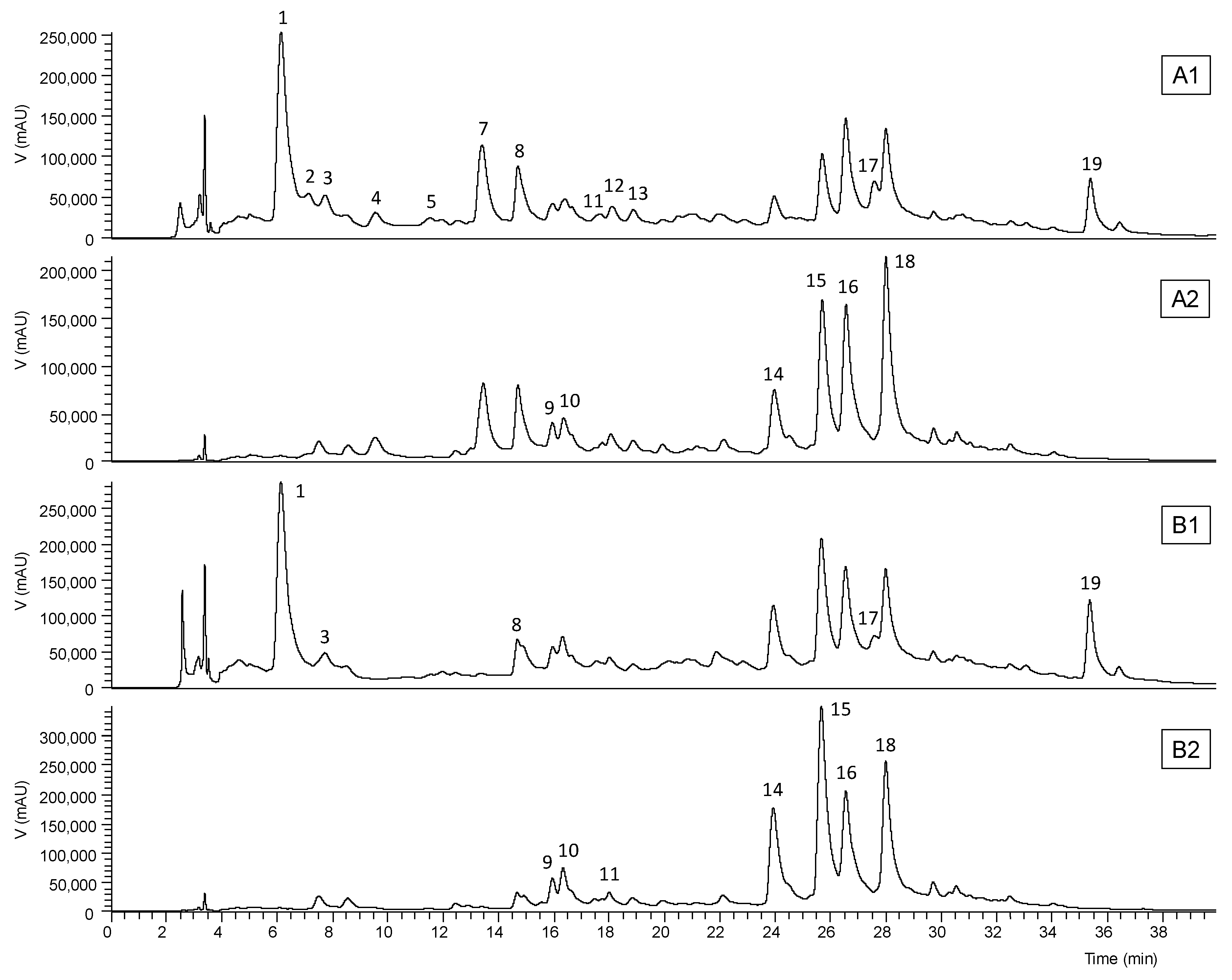
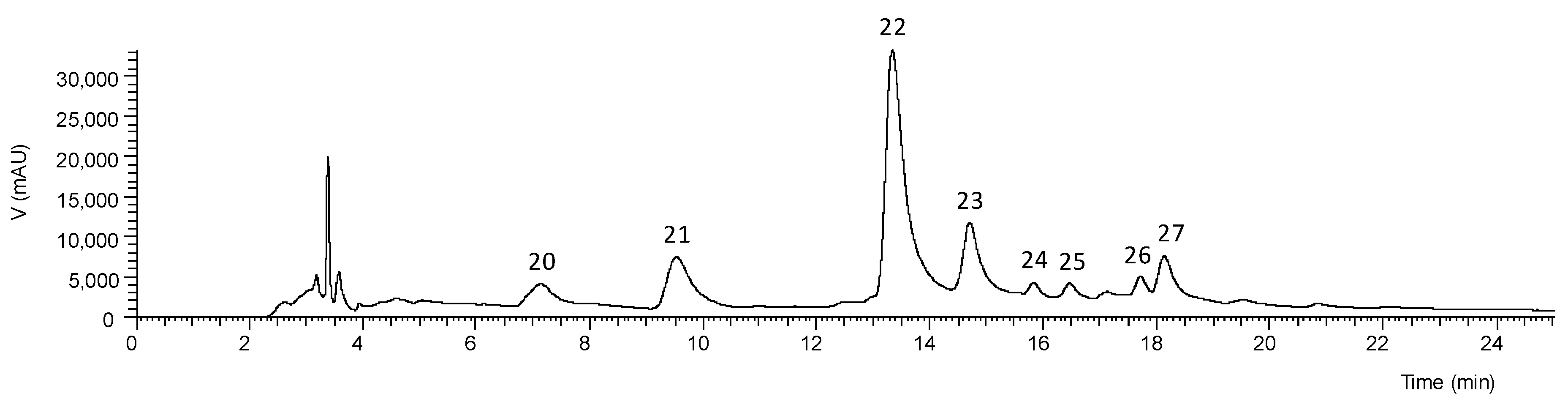
Table S1). The chromatographic profiles recorded at 280 and 370 nm are shown in
Figure 1 and
Figure 2. Up to 17 phenolic compounds were identified in the leaf extracts and eight in the flower extract, which were classified into four groups—phenolic acids, flavonoids, chromones, and anthrones. Most of these compounds have already been previously reported in
Aloe vera [
50,
51,
52,
53], so that their identities were attributed by interpreting data acquired from HPLC-DAD-ESI/MS
n with those of literature.
The chromones aloesin or aloeresin B (peak 1) and 2’-
p-methoxycoumaroylaloresin (peak 17) and the anthrones 10-hydroxyaloin B (peak 9), 10-hydroxyaloin A (peak 10), aloin B or isobarbaloin (peak 14), aloin A or barbaloin (peak 15), malonyl aloin B (peak 16), and malonyl aloin A (peak 18) were detected in the three studied parts of the
Aloe vera leaf (
Table 6). The mucilage contained the highest content (131 ± 3 mg/g extract) of phenolic compounds, mostly anthrones (62.1%) and chromones (34.6%), followed by two luteolin glucosides (3.3%). This result is in accordance with the literature, which states that the vascularized layer that covers the inner fillet is rich in anthraquinone glycosides and anthrone derivatives [
3]. The rind was ranked second, with 105 ± 3 mg/g extract of phenolic compounds, of which 44.9% anthrones and 43.8% chromones; it also contained luteolin and apigenin glucosides and the phenolic acid
p-coumaroylquinic acid. However, the chromone levels found in the rind did not differ statistically from those of the mucilage (
Table 6). Although the phenolic profiles of the fillet and mucilage were similar, a significantly lower concentration (11.2 ± 0.2 mg/g extract) of these secondary metabolites was found in the fillet. In addition, this leaf part had an equal ratio of anthrones and chromones (
Table 6).
Variations in the phenolic profiles of
Aloe species have been reported. According to Fan et al. [
50], aloesin is more abundant in
A. barbadensis and
A. ferox than in
A. chinensis and
A. arborescens. In these species, aloin A predominated over aloin B (according to our results), and lower concentrations were also found in
A. chinensis. In general, higher contents and more complex phenolic compounds were reported in
A. barbadensis. Kanama et al. [
54] found minimal qualitative variations in the phenolic profiles of
A. ferox exudate samples obtained from different regions of South Africa. Despite this, aloin B content varied from 18.4 to 149.7 mg/g, aloin A ranged from 21.3 to 133.4 mg/g, and aloesin from 111.8 to 561.8 mg/g of dried exudate. This result corroborates the data presented in
Table 6, since aloesin predominated over both aloins, despite lower levels have been quantified in our samples.
6′-Malonylnataloin (peak 13), a malonylated derivative of the rare anthrone nataloin, was detected in the rind extract (
Table 6). This anthrone
C-glycoside is considered of great importance in systematic discrimination of different
Aloe species and has been reported in
A. vera,
A. arborescens,
A. ellenbeckii,
A. eru,
A. grandidentata,
A. brevifolia, and
A. ferox [
52,
55].
The quantification of aloins (as hydroxyanthracene derivatives) is recommended in routine quality control analyses of
Aloe samples. These compounds are highly valorised in the pharmaceutical industry, allowed in dietary supplements, and used in small quantities as a bittering agent in alcoholic beverages. However, because of their laxative properties, levels of aloin A and B in
Aloe leaf preparations intended for oral consumption were limited by the International Aloe Science Council to 10 ppm (10 mg/kg) or less [
56]. These levels can be controlled and limited by adding purification steps in the manufacturing process.
The phenolic profile of
Aloe vera flower (
Figure 2) was different from that of leaf (
Figure 1), being constituted mainly by the flavonoids apigenin-6,8-
C-diglucoside, apigenin-2’’-
O-pentoxide-
C-hexoside, apigenin-6-
C-glucoside, and traces of luteolin glucoside derivatives (accounting for 93.4% of the extract), and by the phenolic acid 5-
O-caffeoylquinic acid (
Table 5). As shown in
Table 6, this extract had the lowest levels (4.78 ± 0.05 mg/g extract) of phenolic compounds. As far as we know, it is the first time that some of these compounds are described in
Aloe vera flower. No anthraquinone glycosides were detected in this part of the plant as previously stated by Keyhanian and Stahl-Biskup [
51].


 ,
, 






{kind=link}
{kind=link}
{kind=link}