Phenolic Profile, Antioxidant, and Antidiabetic Potential Exerted by Millet Grain Varieties

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Materials
2.3. Preparation of Ethanolic Extracts
2.4. Total Phenolic Content (TPC)
2.5. Total Flavonoid Content (TFC)
2.6. Total Condensed Tannin Content (CTC)
2.7. Antioxidant Activities Assays
2.7.1. DPPH Radical Scavenging Activity
2.7.2. ABTS Radical Scavenging Activity
2.8. Carbohydrate Digestion Enzymes and Glycation Inhibitions
2.8.1. α-Amylase Inhibitory Assay
2.8.2. α-Glucosidase Inhibitory Assay
2.8.3. Inhibition of AGEs Formation
2.9. Ultra-High PerformanceLliquid Chromatography Quadrupole Time-of-Flight Mass Spectrometry (UHPLC-Q-TOF-MS/MS) Phenolic Compounds Identification
2.10. Statistical Analysis
3. Results and Discussion
3.1. TPC, TFC, and CTC of Ethanol Extracts
3.2. Antioxidant Capacity of Ethanol Extracts
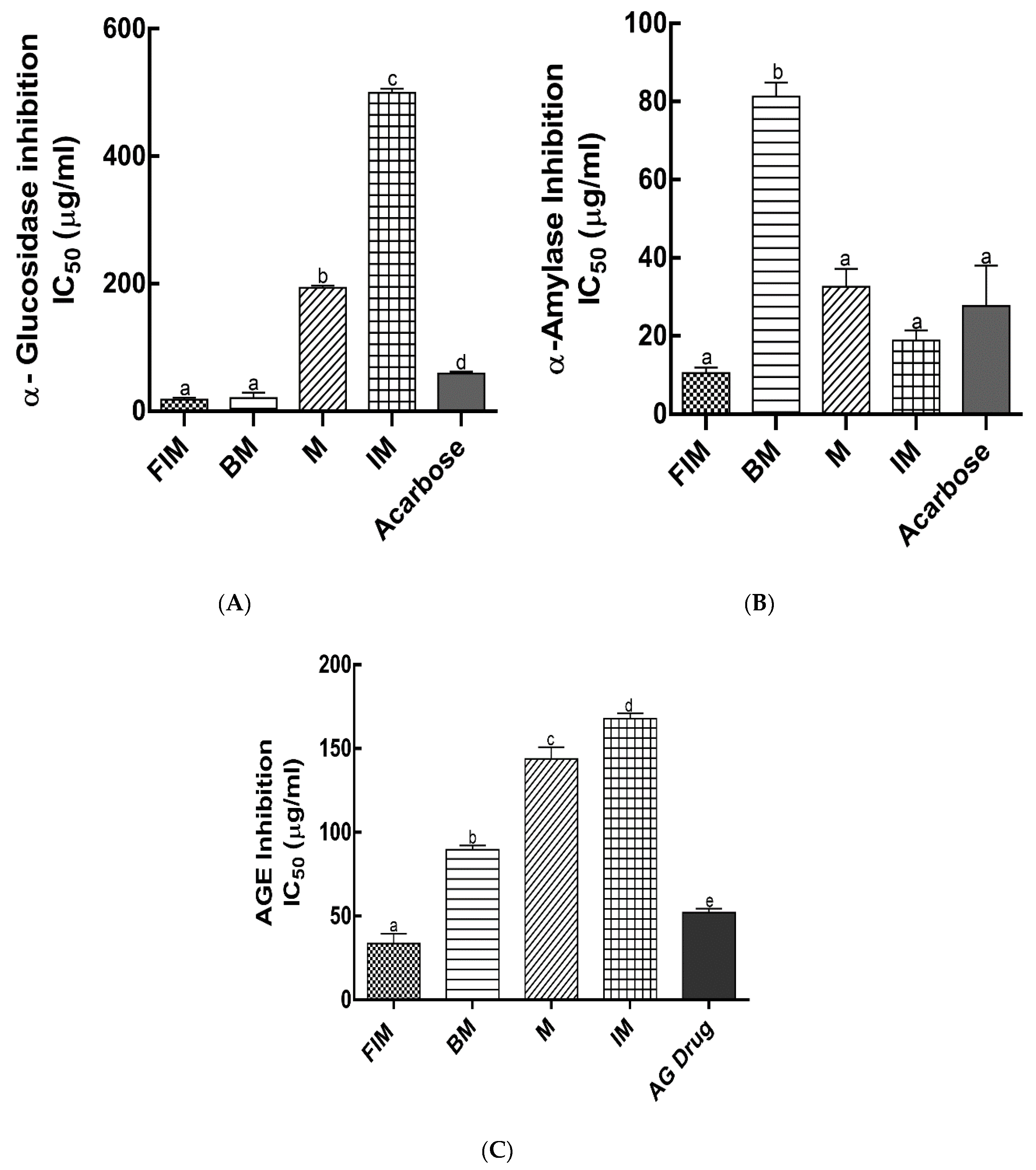
3.3. Antidiabetic Activity In Vitro
3.4. Analyses of Phenolic Compounds by UHPLC-Q-TOF-MS2
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| FIM | finger Italian millet |
| BM | barnyard millet |
| IM | Italian millet |
| M | millet |
| DPPH | 2,2’-diphenyl-1-picrylhydrazyl |
| ABTS | 2-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt |
| AGEs | advanced glycation endproducts |
| TPC | total phenolic content |
| TFC | flavonoid content |
| CTC | condensed tannin content |
| FAE | ferulic acid equivalent |
| CE | catechin equivalent |
| DW | dry weight |
| RT | retention time |
| UHPLC-Q-TOF-MS/MS | ultra-high performance liquid chromatography quadrupole time-of-flight mass spectrometry |
References
- DeFronzo, R.A.; Tripathy, D. Skeletal muscle insulin resistance is the primary defect in type 2 diabetes. Diabetes Care 2009, 32, S157–S163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, G.G.; Francis, N.; Hill, R.; Waters, D.; Blanchard, C.; Santhakumar, A.B. Dietary Polyphenols and Gene Expression in Molecular Pathways Associated with Type 2 Diabetes Mellitus: A Review. Int. J. Mol. 2020, 21, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, E.J.; LeRoith, D.; Karnieli, E. Insulin resistance in obesity as the underlying cause for the metabolic syndrome. Mt. Sinai J. Med. 2010, 77, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Migdal, C.; Serres, M. Reactive oxygen species and oxidative stress. MS Med. Sci. 2011, 27, 405–412. [Google Scholar]
- Chang, A.M.; Smith, M.J.; Bloem, C.J.; Galecki, A.T.; Halter, J.B. Effect of lowering postprandial hyperglycemia on insulin secretion in older people with impaired glucose tolerance. Am. J. Physiol. Endocrinol. 2004, 287, E906–E911. [Google Scholar] [CrossRef]
- Vinson, J.A.; Howard, T.B., III. Inhibition of protein glycation and advanced glycation end products by ascorbic acid and other vitamins and nutrients. J. Nutr. Biochem. 1996, 7, 659–663. [Google Scholar] [CrossRef]
- Pradeep, P.; Sreerama, Y.N. Soluble and bound phenolics of two different millet genera and their milled fractions: Comparative evaluation of antioxidant properties and inhibitory effects on starch hydrolysing enzyme activities. J. Funct. Foods 2017, 35, 682–693. [Google Scholar] [CrossRef]
- Pradeep, P.; Sreerama, Y.N. Phenolic antioxidants of foxtail and little millet cultivars and their inhibitory effects on α-amylase and α-glucosidase activities. Food Chem. 2018, 247, 46–55. [Google Scholar] [CrossRef]
- Sekhon-Loodu, S.; Rupasinghe, H. Evaluation of antioxidant, antidiabetic and antiobesity potential of selected traditional medicinal plants. Front. Nutr. 2019, 6, 53. [Google Scholar] [CrossRef]
- Adinortey, M.B.; Agbeko, R.; Boison, D.; Ekloh, W.; Kuatsienu, L.E.; Biney, E.E.; Affum, O.O.; Kwarteng, J.; Nyarko, A.K. Phytomedicines used for diabetes mellitus in Ghana: A systematic search and review of preclinical and clinical evidence. Evid. Based Complement. Alternat. Med. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekara, A.; Shahidi, F. Content of insoluble bound phenolics in millets and their contribution to antioxidant capacity. J. Agric. Food Chem. 2010, 58, 6706–6714. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, T.; Muthamilarasan, M.; Prasad, M. Millets for next generation climate-smart agriculture. Front. Plant Sci. 2017, 8, 1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahidi, F.; Chandrasekara, A. Millet grain phenolics and their role in disease risk reduction and health promotion: A review. J. Funct. Foods. 2013, 5, 570–581. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Shahidi, F. Antiproliferative potential and DNA scission inhibitory activity of phenolics from whole millet grains. J. Funct. Foods. 2011, 3, 159–170. [Google Scholar] [CrossRef]
- Vadivoo, A.S.; Joseph, R.; Ganesan, N.M. Genetic variability and diversity for protein and calcium contents in finger millet (Eleusine coracana (L.) Gaertn) in relation to grain color. Plant Food Hum. Nutri. 1998, 52, 353–364. [Google Scholar] [CrossRef]
- Upadhyaya, N.; Nosenko, V.; Mišković, Z.; Hou, L.-J.; Ivlev, A.; Morfill, G. A full account of compressional wave in 2D strongly coupled complex (dusty) plasmas: Theory, experiment and numerical simulation. EPL. 2011, 94, 65001. [Google Scholar] [CrossRef]
- Van Hung, P. Phenolic compounds of cereals and their antioxidant capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875. [Google Scholar] [CrossRef]
- Apea-Bah, F.B.; Minnaar, A.; Bester, M.J.; Duodu, K.G. Sorghum–cowpea composite porridge as a functional food, Part II: Antioxidant properties as affected by simulated in vitro gastrointestinal digestion. Food Chem. 2016, 197, 307–315. [Google Scholar] [CrossRef]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Li, W.; Pickard, M.D.; Beta, T. Effect of thermal processing on antioxidant properties of purple wheat bran. Food Chem. 2007, 104, 1080–1086. [Google Scholar] [CrossRef]
- Xiang, J.; Apea-Bah, F.B.; Ndolo, V.U.; Katundu, M.C.; Beta, T. Profile of phenolic compounds and antioxidant activity of finger millet varieties. Food Chem. 2019, 275, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Okarter, N.; Liu, C.-S.; Sorrells, M.E.; Liu, R.H. Phytochemical content and antioxidant activity of six diverse varieties of whole wheat. Food Chem. 2010, 119, 249–257. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Shahidi, F. Determination of antioxidant activity in free and hydrolyzed fractions of millet grains and characterization of their phenolic profiles by HPLC-DAD-ESI-MSn. J. Funct. Foods 2011, 3, 144–158. [Google Scholar] [CrossRef]
- Pradeep, P.; Sreerama, Y.N. Impact of processing on the phenolic profiles of small millets: Evaluation of their antioxidant and enzyme inhibitory properties associated with hyperglycemia. Food Chem. 2015, 169, 455–463. [Google Scholar] [CrossRef]
- Hyun, T.K.; Kim, H.-C.; Ko, Y.-J.; Kim, J.-S. Antioxidant, α-glucosidase inhibitory and anti-inflammatory effects of aerial parts extract from Korean crowberry (Empetrum nigrum var. japonicum). Saudi J. Biol. Sci. 2016, 23, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, I.M.; Li-Chan, E.C. Overview of food products and dietary constituents with antidiabetic properties and their putative mechanisms of action: A natural approach to complement pharmacotherapy in the management of diabetes. Mol. Nutri. Food Res. 2014, 58, 61–78. [Google Scholar] [CrossRef]
- Baldea, L.A.N.; Martineau, L.C.; Benhaddou-Andaloussi, A.; Arnason, J.T.; Lévy, É.; Haddad, P.S. Inhibition of intestinal glucose absorption by anti-diabetic medicinal plants derived from the James Bay Cree traditional pharmacopeia. J. Ethnopharmacol. 2010, 132, 473–482. [Google Scholar] [CrossRef]
- Wang, H.; Liu, T.; Huang, D. Starch hydrolase inhibitors from edible plants. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2013; Volume 70, pp. 103–136. [Google Scholar]
- Lim, J.; Zhang, X.; Ferruzzi, M.G.; Hamaker, B.R. Starch digested product analysis by HPAEC reveals structural specificity of flavonoids in the inhibition of mammalian α-amylase and α-glucosidases. Food Chem. 2019, 288, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Hua, F.; Zhou, P.; Wu, H.-Y.; Chu, G.-X.; Xie, Z.-W.; Bao, G.-H. Inhibition of α-glucosidase and α-amylase by flavonoid glycosides from Lu’an GuaPian tea: Molecular docking and interaction mechanism. Food Funct. 2018, 9, 4173–4183. [Google Scholar] [CrossRef]
- Wu, L.; Liu, Y.; Qin, Y.; Wang, L.; Wu, Z. HPLC-ESI-qTOF-MS/MS characterization, antioxidant activities and inhibitory ability of digestive enzymes with molecular docking analysis of various parts of raspberry (Rubus ideaus L.). Antioxidants 2019, 8, 274. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Jia, Y.; Ma, Y.; Cheng, G.; Cai, S. Phenolic composition, antioxidant properties, and inhibition toward digestive enzymes with molecular docking analysis of different fractions from Prinsepia utilis royle fruits. Molecules 2018, 23, 3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, A.; Morales, M.; Gonzalez, I.; Araya, P. Inhibition of RAGE axis signaling: A pharmacological challenge. Curr. Drug Targets 2019, 20, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Rowan, S.; Bejarano, E.; Taylor, A. Mechanistic targeting of advanced glycation end-products in age-related diseases. Biochim. Biophys. Acta. Mol. Basis Dis. 2018, 1864, 3631–3643. [Google Scholar] [CrossRef] [PubMed]
- Martini, S.; Conte, A.; Bottazzi, S.; Tagliazucchi, D. Mediterranean diet vegetable foods protect meat lipids from oxidation during in vitro gastro-intestinal digestion. Int. J. Food Sci. Nutr. 2019, 1–16. [Google Scholar] [CrossRef]
- Zhou, Q.; Gong, J.; Wang, M. Phloretin and its methylglyoxal adduct: Implications against advanced glycation end products-induced inflammation in endothelial cells. Food Chem. Toxicol. 2019, 129, 291–300. [Google Scholar] [CrossRef]
- Yılmaz, Z.; Kalaz, E.B.; Aydın, A.F.; Olgaç, V.; Doğru-Abbasoğlu, S.; Uysal, M.; Koçak-Toker, N. The effect of resveratrol on glycation and oxidation products in plasma and liver of chronic methylglyoxal-treated rats. Pharmacol. Rep. 2018, 70, 584–590. [Google Scholar] [CrossRef]
- Yeh, W.-J.; Hsia, S.-M.; Lee, W.-H.; Wu, C.-H. Polyphenols with antiglycation activity and mechanisms of action: A review of recent findings. J. Food Drug Anal. 2017, 25, 84–92. [Google Scholar] [CrossRef]
- Crascì, L.; Lauro, M.R.; Puglisi, G.; Panico, A. Natural antioxidant polyphenols on inflammation management: Anti-glycation activity vs metalloproteinases inhibition. Crit. Rev. Food Sci. Nutri. 2018, 58, 893–904. [Google Scholar] [CrossRef]
- Harris, C.S.; Cuerrier, A.; Lamont, E.; Haddad, P.S.; Arnason, J.T.; Bennett, S.A.; Johns, T. Investigating wild berries as a dietary approach to reducing the formation of advanced glycation endproducts: Chemical correlates of in vitro antiglycation activity. Plant Food Hum. Nutr. 2014, 69, 71–77. [Google Scholar] [CrossRef] [Green Version]
- González, I.; Morales, M.A.; Rojas, A. Polyphenols and AGEs/RAGE axis. Trends and challenges. Food Res. Int. 2019, 108843. [Google Scholar] [CrossRef]
- Phenol-Explorer: Comprehensive Database on Polyphenol Content in Foods. Available online: https://phenol-explorer.eu/ (accessed on 6 February 2020).
- Watanabe, M. Antioxidative Phenolic Compounds from Japanese Barnyard Millet (Echinochloa utilis) Grains. J. Agric. Food Chem. 1999, 47, 4500–4505. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Azimi-Nezhad, M.; Farkhondeh, T. Catechin Treatment Ameliorates Diabetes and Its Complications in Streptozotocin-Induced Diabetic Rats. Dose-Response 2017, 15, 1559325817691158. [Google Scholar] [CrossRef] [PubMed]
- Ueda-Wakagi, M.; Nagayasu, H.; Yamashita, Y.; Ashida, H. Green Tea Ameliorates Hyperglycemia by Promoting the Translocation of Glucose Transporter 4 in the Skeletal Muscle of Diabetic Rodents. Int. J. Mol. Sci. 2019, 20, 2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Millet Varieties | TPC (mg Ferulic Acid Equivalent/100 g, DW) | TFC (mg Catechin Equivalent/100 g, DW) | CTC (mg Catechin Equivalent/100 g, DW) |
|---|---|---|---|
| M | 125.1 ± 6.36 a | 101.9 ± 9.1 a | 50.50 ± 3.83 a |
| IM | 107.8 ± 7.02 b | 105.4 ± 9.1 a | 36.37 ± 4.56 c |
| BM | 129.5 ± 4.95 a | 101.3 ± 10.4 a | 59.54 ± 4.63 a |
| FIM | 136.4 ± 7.07 a | 115.8 ± 9.1 a | 17.65 ± 3.95 b |
| Peak No. | RT (min) | Molecular Formula | Molecular Weight | [M-H]- (m/z) | Compound Identified |
|---|---|---|---|---|---|
| 1 | 8.73 | C21H24O11 | 452.1321 | 451.1249 | (+)-Catechin 3-O-glucose |
| 2 | 9.54 | C15H14O6 | 290.0797 | 289.0721 | (+)-Catechin |
| 3 | 9.67 | C21H22O12 | 466.1113 | 465.1041 | Dihydromyricetin 3-O-rhamnoside |
| 4 | 11.06 | C15H14O6 | 290.0797 | 289.0722 | (−)-Epicatechin |
| 5 | 12.42 | C15H12O7 | 304.0585 | 303.0514 | Dihydroquercetin |
| 6 | 12.67 | C16H12O4 | 268.0739 | 267.0664 | Formononetin |
| 7 | 15.63 | C15H10O6 | 286.0482 | 285.0408 | Kaempferol |
| 8 | 16.42 | C15H10O5 | 270.0530 | 269.0457 | Apigenin |
| Peak No. | RT (min) | Molecular Formula | Molecular Weight | [M-H]− (m/z) | Compound Identified |
|---|---|---|---|---|---|
| 1 | 9.54 | C15H14O6 | 290.0797 | 289.0721 | (+)-Catechin |
| 2 | 12.68 | C16H12O4 | 268.0739 | 267.0664 | Formononetin |
| 3 | 15.63 | C15H10O6 | 286.0482 | 285.0408 | Kaempferol |
| 4 | 16.38 | C15H10O6 | 286.0482 | 285.0408 | Luteolin |
| 5 | 16.43 | C15H10O5 | 270.0530 | 269.0457 | Apigenin |
| 6 | 16.44 | C16H12O7 | 316.0586 | 315.0513 | Isorhamnetin |
| 7 | 16.50 | C17H14O7 | 330.0746 | 329.067 | 3,7-Dimethylquercetin |
| Compound | RT (min) | Finger Italian Millet (µg/100g) | Barnyard Millet (µg/100g) | Polyphenol Class |
|---|---|---|---|---|
| Gallic acid | 2.18 | ND | ND | Phenolic acid |
| Catechin | 9.55 | 577.49 | 146.1 | Flavonoid |
| Caffeic acid | 10.15 | ND | ND | Phenolic acid |
| p-Coumaric acid | 11.95 | ND | ND | Phenolic acid |
| Ferulic acid | 12.58 | ND | ND | Phenolic acid |
| Quercetin | 15.3 | ND | ND | Flavonoid |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ofosu, F.K.; Elahi, F.; Daliri, E.B.-M.; Chelliah, R.; Ham, H.J.; Kim, J.-H.; Han, S.-I.; Hur, J.H.; Oh, D.-H. Phenolic Profile, Antioxidant, and Antidiabetic Potential Exerted by Millet Grain Varieties. Antioxidants 2020, 9, 254. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9030254
Ofosu FK, Elahi F, Daliri EB-M, Chelliah R, Ham HJ, Kim J-H, Han S-I, Hur JH, Oh D-H. Phenolic Profile, Antioxidant, and Antidiabetic Potential Exerted by Millet Grain Varieties. Antioxidants. 2020; 9(3):254. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9030254
Chicago/Turabian StyleOfosu, Fred Kwame, Fazle Elahi, Eric Banan-Mwine Daliri, Ramachandran Chelliah, Hun Ju Ham, Joong-Hark Kim, Sang-Ik Han, Jang Hyun Hur, and Deog-Hwan Oh. 2020. "Phenolic Profile, Antioxidant, and Antidiabetic Potential Exerted by Millet Grain Varieties" Antioxidants 9, no. 3: 254. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox9030254