SDF-1α Gene-Activated Collagen Scaffold Restores Pro-Angiogenic Wound Healing Features in Human Diabetic Adipose-Derived Stem Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Polyplex
2.2. Cell Expansion
2.3. Cell Seeding on SDF-1α Gene-Activated Scaffold
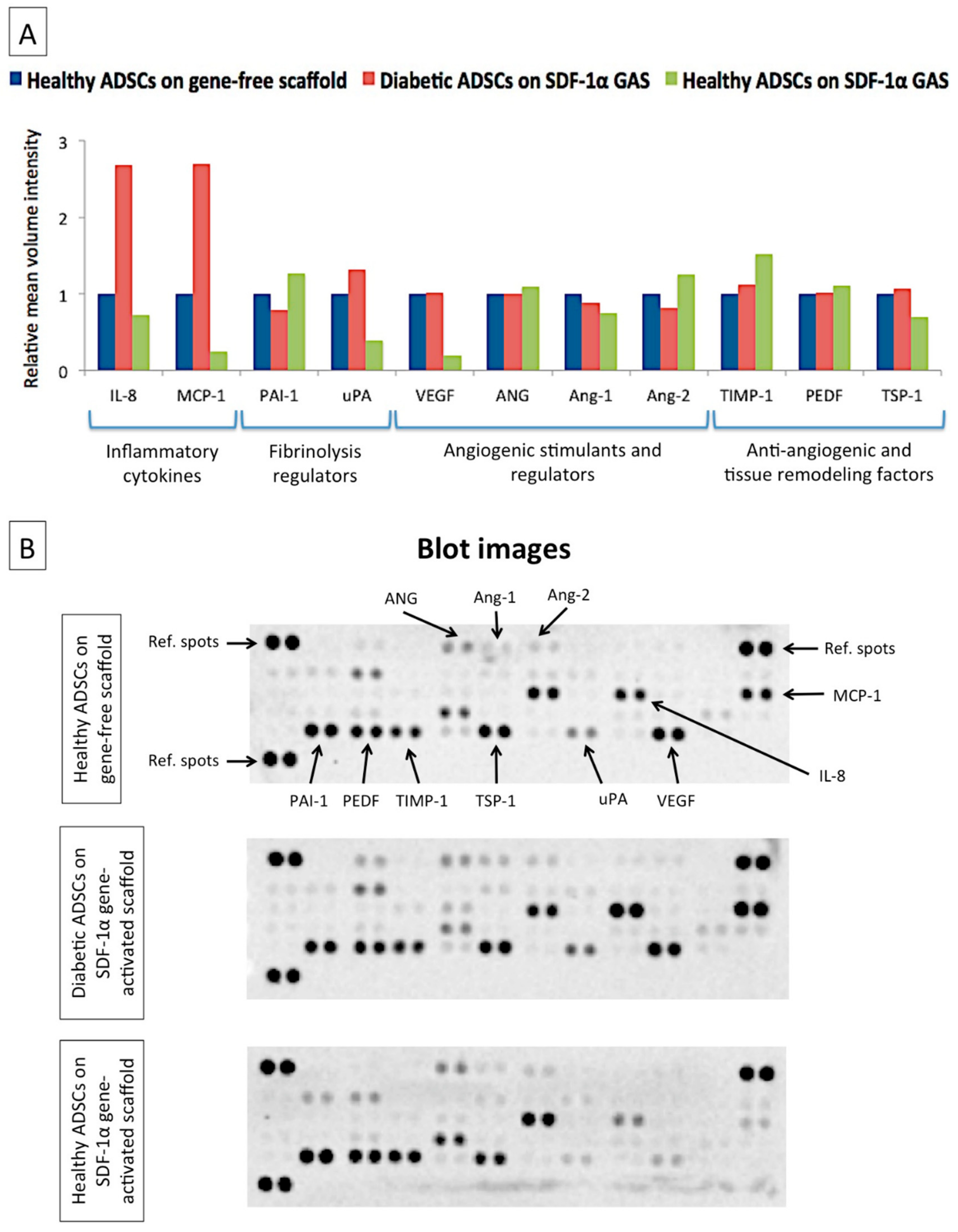
2.4. Proteome Profiling of Secreted Factors from Healthy and Diabetic ADSCs on the Gene-Free Scaffold
2.5. qRT-PCR Analysis to Determine Functional Gene Expression in ADSCs on SDF-1α Gene-Activated Scaffold
2.6. Proteome Profiling of Secreted Factors from Healthy and Diabetic ADSCs on SDF-1α Gene-Activated Scaffold
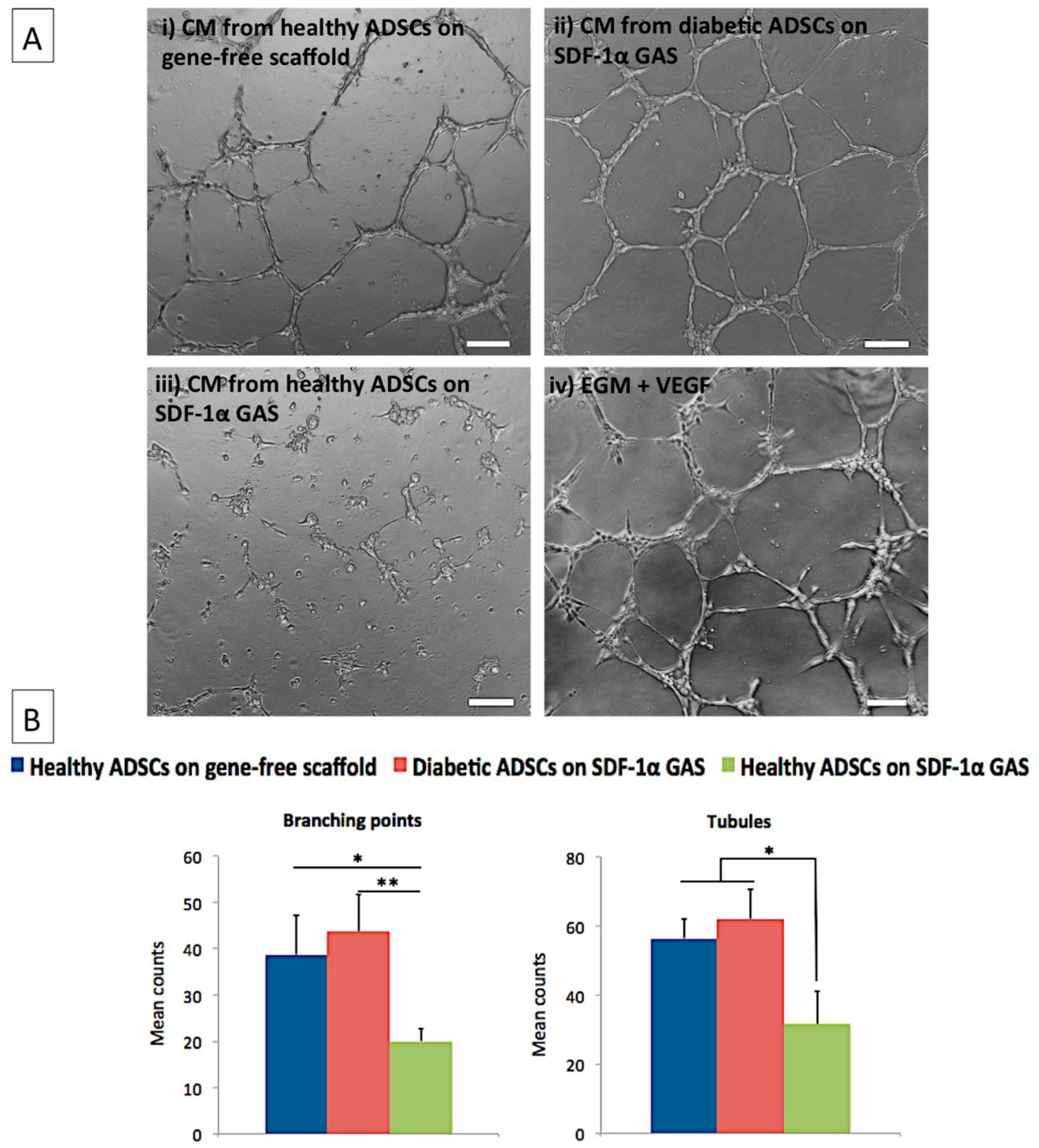
2.7. Pro-Angiogenic Bioactivity Analyses of Secreted Factors from the ADSCs on SDF-1α Gene-Activated Scaffold
2.8. Immunofluorescent Imaging
2.9. Image Analysis
2.10. Statistical Analysis
3. Results
3.1. Diabetes Impairs Signaling of Functional Factors in Human ADSCs
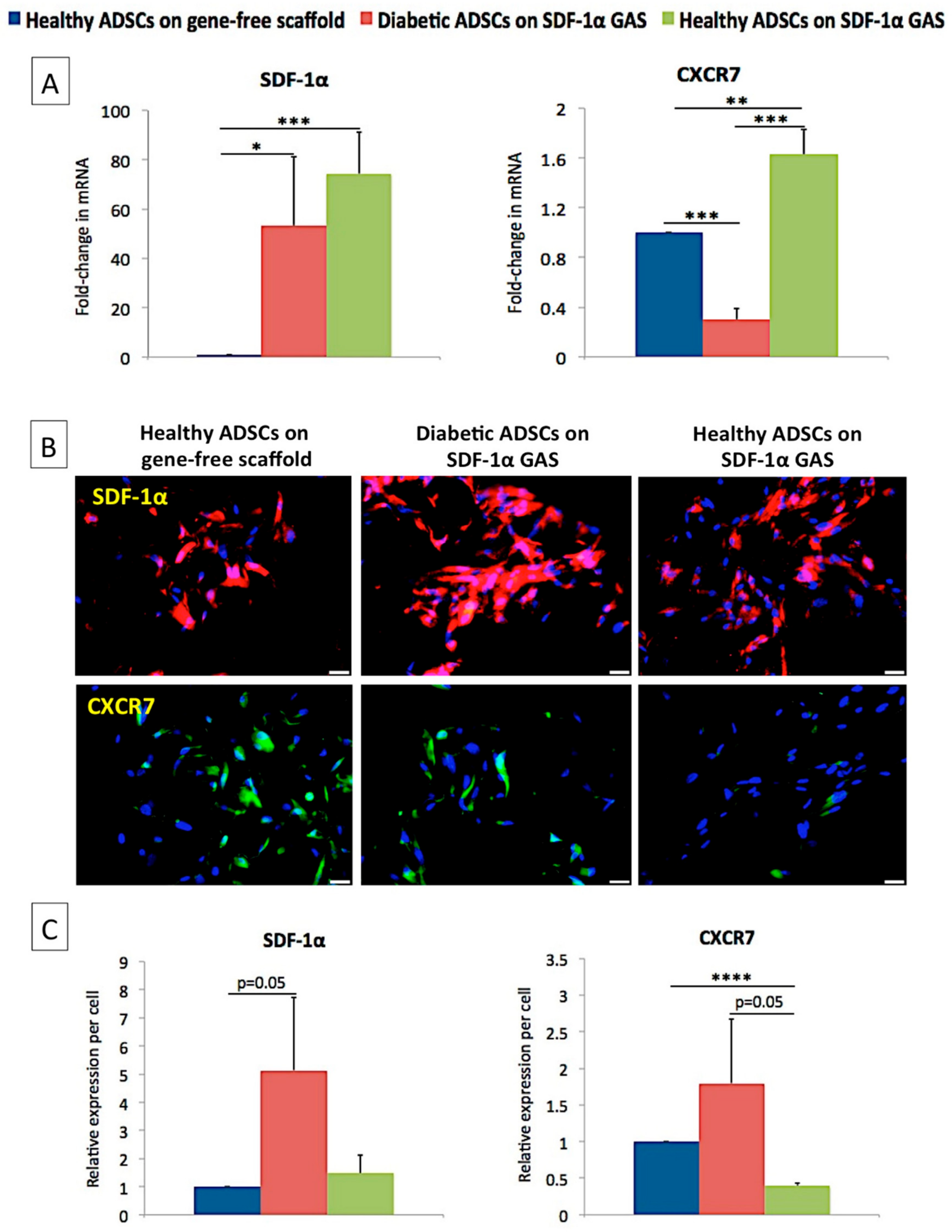
3.2. SDF-1α Gene-Activated Scaffold Promotes Overexpression of SDF-1α mRNA and Engages the CXCR7/β-Arrestin Signaling in Diabetic ADSCs
3.3. SDF-1α Gene-Activated Scaffold Restores a Healthy-Like Signaling of Functional Factors in Diabetic ADSCs
3.4. Diabetic ADSCs on the SDF-1α Gene-Activated Scaffold Effectively Enhance Angiogenesis in Endothelial Cells
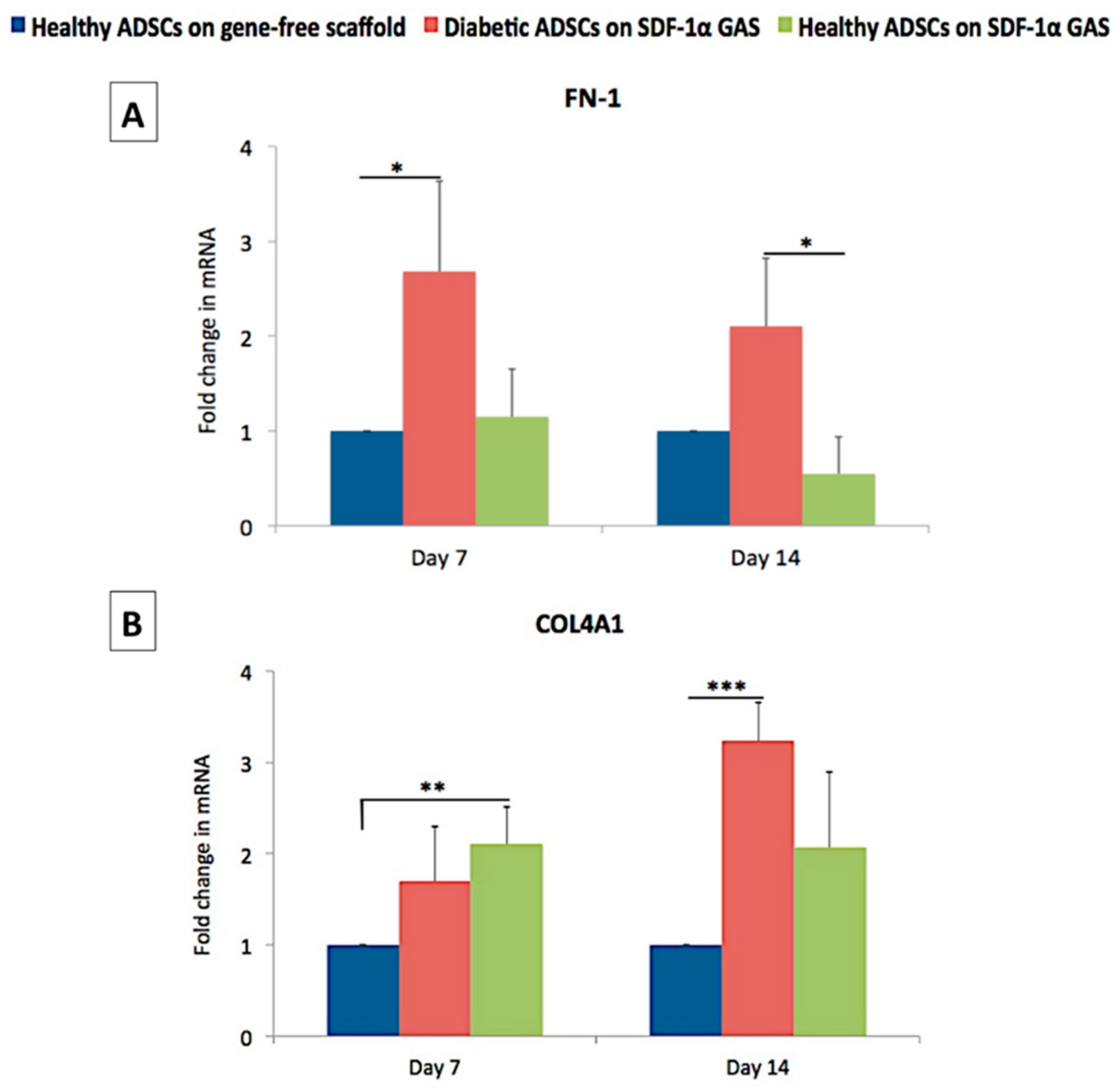
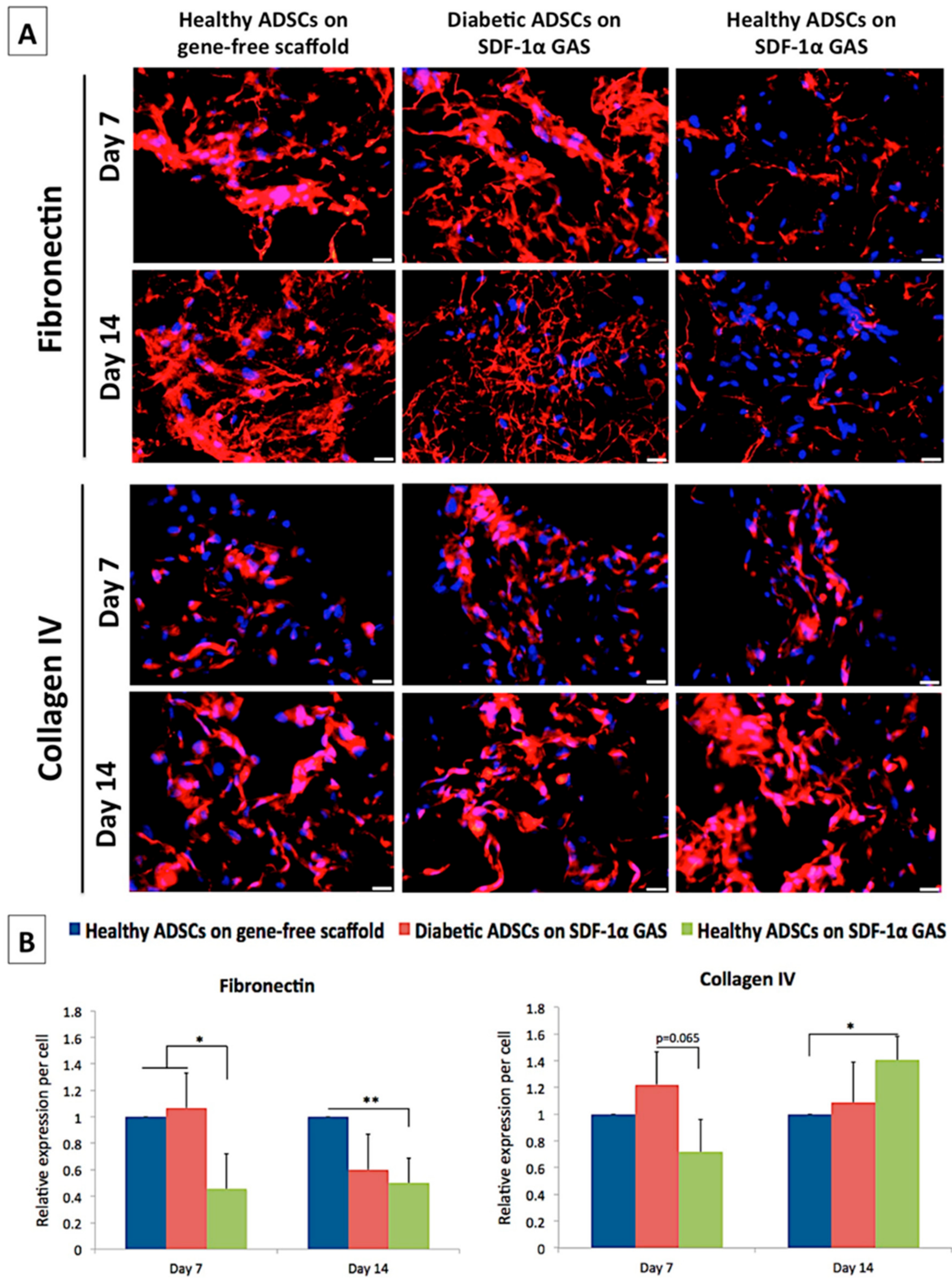
3.5. SDF-1α Gene-Activated Scaffold Promotes Pro-Wound Healing Matrix Remodeling Response in Diabetic ADSCs
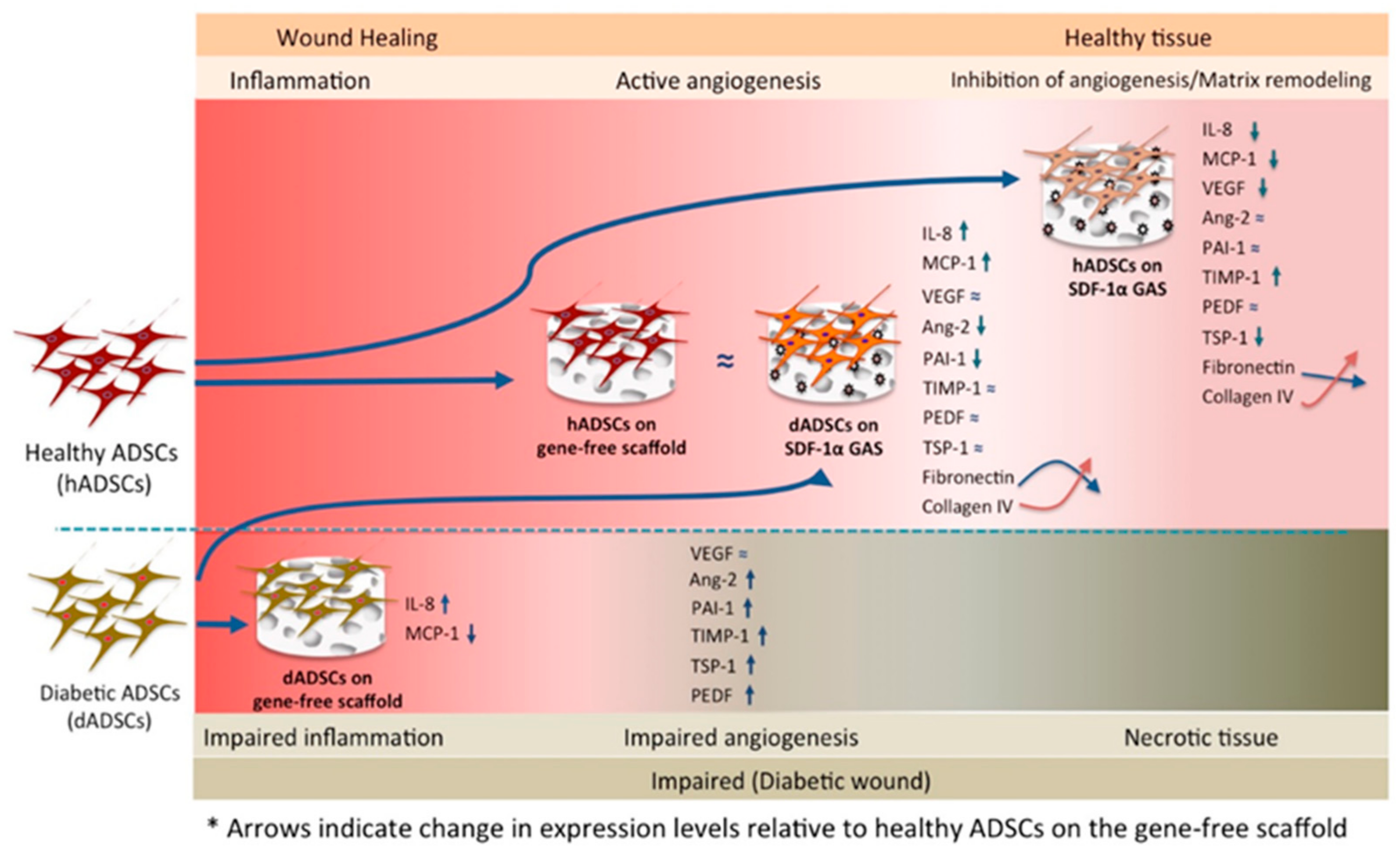
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
| ADSCs | Adipose-derived stem cells |
| Ang-2 | Angiopoeitin-2 |
| BM-MSCs | Bone marrow-mesenchymal stem cells |
| CSF | Colony stimulating factor |
| Coll–CS | Collagen–chondroitin sulfate |
| CM | Conditioned media |
| ECM | Extracellular matrix |
| EDAC/NHS | N-(3-Dimethylaminopropyl)-N’-ethylcarbodiimide hydrochloride/N-Hydroxysuccinimide |
| FDA | Food and Drug Administration |
| GAS | Gene-activated scaffold |
| HUVECs | Human umbilical vein endothelial cells |
| IGFBP-3 | Insulin growth factor binding protein-3 |
| IL-8 | Interleukin-8 |
| MCP-1 | Monocyte chemoattractant protein-1 |
| PAI-1 | Plasminogen activator inhibitor-1 |
| pDNA | Plasmid DNA |
| PEI | Polyethyleneimine |
| PEDF | Pigment epithelium-derived factor |
| SDF-1α | Stromal-derived factor-1 alpha |
| TIMP-1 | Tissue inhibitor of metalloproteinase-1 |
| TSP-1 | Thrombospondin-1 |
| VEGF | Vascular endothelial growth factor |
References
- Wang, Y.; Sun, Y.; Yang, X.Y.; Ji, S.Z.; Han, S.; Xia, Z.F. Mobilised bone marrow-derived cells accelerate wound healing. Int. Wound J. 2013, 10, 473–479. [Google Scholar] [CrossRef]
- Petit, I.; Szyper-Kravitz, M.; Nagler, A.; Lahav, M.; Peled, A.; Habler, L.; Ponomaryov, T.; Taichman, R.S.; Arenzana-Seisdedos, F.; Fujii, N.; et al. G-CSF induces stem cell mobilization by decreasing bone marrow SDF-1 and up-regulating CXCR4. Nat. Immunol. 2002, 3, 687–694. [Google Scholar] [CrossRef]
- Nassar, D.; Batteux, F.; Raymond, K.; Tharaux, P.-L.; Aractingi, S. Delayed healing of sickle cell ulcers is due to impaired angiogenesis and CXCL12 secretion in skin wounds. J. Investig. Dermatol. 2016, 136, 497–506. [Google Scholar]
- Laiva, A.L.; O’Brien, F.J.; Keogh, M.B. Innovations in gene and growth factor delivery systems for diabetic wound healing. J. Tissue Eng. Regen. Med. 2018, 12, e296–e312. [Google Scholar] [CrossRef] [Green Version]
- Dinh, T.; Tecilazich, F.; Kafanas, A.; Doupis, J.; Gnardellis, C.; Leal, E.; Tellechea, A.; Pradhan, L.; Lyons, T.E.; Giurini, J.M.; et al. Mechanisms involved in the development and healing of diabetic foot ulceration. Diabetes 2012, 61, 2937–2947. [Google Scholar] [CrossRef] [Green Version]
- Hunt, D.L. Diabetes: Foot ulcers and amputations. BMJ Clin. Evid. 2011, 2011, 0602. [Google Scholar]
- Zelen, C.M.; Orgill, D.P.; Serena, T.; Galiano, R.; Carter, M.J.; DiDomenico, L.A.; Keller, J.; Kaufman, J.; Li, W.W. A prospective, randomised, controlled, multicentre clinical trial examining healing rates, safety and cost to closure of an acellular reticular allogenic human dermis versus standard of care in the treatment of chronic diabetic foot ulcers. Int. Wound J. 2017, 14, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, B.; Liu, Y.; Zhufu, Z.; Yan, X.; Hou, X.; Dai, J.; Tan, Q. Effect of collagen scaffold with adipose-derived stromal vascular fraction cells on diabetic wound healing: A study in a diabetic porcine model. Tissue Eng. Regen. Med. 2013, 10, 192–199. [Google Scholar] [CrossRef]
- Hart, C.E.; Loewen-Rodriguez, A.; Lessem, J. Dermagraft: Use in the treatment of chronic wounds. Adv. Wound Care 2012, 1, 138–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frykberg, R.G.; Gibbons, G.W.; Walters, J.L.; Wukich, D.K.; Milstein, F.C. A prospective, multicentre, open-label, single-arm clinical trial for treatment of chronic complex diabetic foot wounds with exposed tendon and/or bone: Positive clinical outcomes of viable cryopreserved human placental membrane. Int. Wound J. 2017, 14, 569–577. [Google Scholar] [CrossRef] [PubMed]
- Veves, A.; Falanga, V.; Armstrong, D.G.; Sabolinski, M.L. Graftskin, a human skin equivalent, is effective in the management of noninfected neuropathic diabetic foot ulcers: A prospective randomized multicenter clinical trial. Diabetes Care 2001, 24, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marston, W.A.; Hanft, J.; Norwood, P.; Pollak, R. The efficacy and safety of Dermagraft in improving the healing of chronic diabetic foot ulcers: Results of a prospective randomized trial. Diabetes Care 2003, 26, 1701–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.; Kirsner, R.S.; Falanga, V.; Phillips, T. and Eaglstein, W.H. Evaluation of Apligraf® persistence and basement membrane restoration in donor site wounds: A pilot study. Wound Repair Regen. 2006, 14, 427–433. [Google Scholar] [CrossRef]
- Lopes, L.; Setia, O.; Aurshina, A.; Liu, S.; Hu, H.; Isaji, T.; Liu, H.; Wang, T.; Ono, S.; Guo, X.; et al. Stem cell therapy for diabetic foot ulcers: A review of preclinical and clinical research. Stem Cell Res. Ther. 2018, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Chen, B.; Liang, Z.; Deng, W.; Jiang, Y.; Li, S.; Xu, J.; Wu, Q.; Zhang, Z.; Xie, B.; et al. Comparison of bone marrow mesenchymal stem cells with bone marrow-derived mononuclear cells for treatment of diabetic critical limb ischemia and foot ulcer: A double-blind, randomized, controlled trial. Diabetes Res. Clin. Pract. 2011, 92, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.C.; An, S.G.; Lee, H.W.; Park, J.-S.; Cha, K.S.; Hong, T.J.; Park, J.H.; Lee, S.Y.; Kim, S.P.; Kim, Y.D.; et al. Safety and effect of adipose tissue-derived stem cell implantation in patients with critical limb ischemia. Circ. J. 2012, 1204091686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Zhang, A.; Tao, C.; Li, X.; Jin, P. The role of SDF-1-CXCR4/CXCR7 axis in biological behaviors of adipose tissue-derived mesenchymal stem cells in vitro. Biochem. Biophys. Res. Commun. 2013, 441, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Kim, Y.H.; Jun, Y.J.; Yoo, G.; Rhie, J.W. The effect of diabetes on the wound healing potential of adipose-tissue derived stem cells. Int. Wound J. 2016, 13, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Han, J.W.; Lee, J.Y.; Choi, Y.J.; Sohn, Y.-D.; Song, M.; Yoon, Y.S. Diabetic mesenchymal stem cells are ineffective for improving limb ischemia due to their impaired angiogenic capability. Cell Transplant. 2015, 24, 1571–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massee, M.; Chinn, K.; Lim, J.J.; Godwin, L.; Young, C.S.; Koob, T.J. Type I and II diabetic adipose-derived stem cells respond in vitro to dehydrated human amnion/chorion membrane allograft treatment by increasing proliferation, migration, and altering cytokine secretion. Adv. Wound Care 2016, 5, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Cianfarani, F.; Toietta, G.; Di Rocco, G.; Cesareo, E.; Zambruno, G.; Odorisio, T. Diabetes impairs adipose tissue–derived stem cell function and efficiency in promoting wound healing. Wound Repair Regen. 2013, 21, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Yang, X.; Qin, J.; Ye, K.; Wang, X.; Shi, H.; Jiang, M.; Liu, X.; Lu, X. Glyoxalase-1 overexpression reverses defective proangiogenic function of diabetic adipose-derived stem cells in streptozotocin-induced diabetic mice model of critical limb ischemia. Stem Cells Transl. Med. 2017, 6, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnoy, S.; Lee, P.-Y.; Huang, L. Intradermal injection of transforming growth factor-β1 gene enhances wound healing in genetically diabetic mice. Pharm. Res. 2003, 20, 345–350. [Google Scholar] [CrossRef]
- Ferraro, B.; Cruz, Y.L.; Coppola, D.; Heller, R. Intradermal delivery of plasmid VEGF165 by electroporation promotes wound healing. Mol. Ther. 2009, 17, 651–657. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, F.J. Biomaterials & scaffolds for tissue engineering. Mater. Today 2011, 14, 88–95. [Google Scholar]
- Wang, C.; Ma, L.; Gao, C. Design of gene-activated matrix for the repair of skin and cartilage. Polym. J. 2014, 46, 476–482. [Google Scholar] [CrossRef]
- Raftery, R.M.; Walsh, D.P.; Ferreras, L.B.; Castaño, I.M.; Chen, G.; LeMoine, M.; Osman, G.; Shakesheff, K.M.; Dixon, J.E.; O’Brien, F.J. Highly versatile cell-penetrating peptide loaded scaffold for efficient and localised gene delivery to multiple cell types: From development to application in tissue engineering. Biomaterials 2019, 216, 119277. [Google Scholar] [CrossRef]
- Walsh, D.P.; Raftery, R.M.; Castaño, I.M.; Murphy, R.; Cavanagh, B.; Heise, A.; O’Brien, F.J.; Cryan, S.A. Transfection of autologous host cells in vivo using gene activated collagen scaffolds incorporating star-polypeptides. J. Control. Release 2019, 304, 191–203. [Google Scholar] [CrossRef]
- Lackington, W.A.; Raftery, R.M.; O’Brien, F.J. In vitro efficacy of a gene-activated nerve guidance conduit incorporating non-viral PEI-pDNA nanoparticles carrying genes encoding for NGF, GDNF and c-Jun. Acta Biomater. 2018, 75, 115–128. [Google Scholar] [CrossRef]
- Lohana, P.; Hassan, S.; Watson, S. Integra™ in burns reconstruction: Our experience and report of an unusual immunological reaction. Ann. Burns Fire Disasters 2014, 27, 17. [Google Scholar]
- Dinescu, S.; Gălăţeanu, B.; Albu, M.; Lungu, A.; Radu, E.; Hermenean, A.; Costache, M. Biocompatibility assessment of novel collagen-sericin scaffolds improved with hyaluronic acid and chondroitin sulfate for cartilage regeneration. BioMed Res. Int. 2013, 2013, 598056. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Gao, C.; Mao, Z.; Zhou, J.; Shen, J.; Hu, X.; Han, C. Collagen/chitosan porous scaffolds with improved biostability for skin tissue engineering. Biomaterials 2003, 24, 4833–4841. [Google Scholar] [CrossRef]
- Liu, X.; Ma, L.; Liang, J.; Zhang, B.; Teng, J.; Gao, C. RNAi functionalized collagen-chitosan/silicone membrane bilayer dermal equivalent for full-thickness skin regeneration with inhibited scarring. Biomaterials 2013, 34, 2038–2048. [Google Scholar] [CrossRef] [PubMed]
- Laiva, A.L.; Raftery, R.M.; Keogh, M.B.; O’brien, F.J. Pro-angiogenic impact of SDF-1α gene-activated collagen-based scaffolds in stem cell driven angiogenesis. Int. J. Pharm. 2018, 544, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Raftery, R.M.; Tierney, E.G.; Curtin, C.M.; Cryan, S.-A.; O’Brien, F.J. Development of a gene-activated scaffold platform for tissue engineering applications using chitosan-pDNA nanoparticles on collagen-based scaffolds. J. Control. Release 2015, 210, 84–94. [Google Scholar] [CrossRef]
- Kolakshyapati, P.; Li, X.; Chen, C.; Zhang, M.; Tan, W.; Ma, L.; Gao, C. Gene-activated matrix/bone marrow-derived mesenchymal stem cells constructs regenerate sweat glands-like structure in vivo. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Cassidy, F.C.; Shortiss, C.; Murphy, C.G.; Kearns, S.R.; Curtin, W.; De Buitléir, C.; O’Brien, T.; Coleman, C.M. Impact of Type 2 Diabetes Mellitus on Human Bone Marrow Stromal Cell Number and Phenotypic Characteristics. Int. J. Mol. Sci. 2020, 21, 2476. [Google Scholar] [CrossRef] [Green Version]
- Nishiguchi, M.A.; Spencer, C.A.; Leung, D.H.; Leung, T.H. Aging suppresses skin-derived circulating SDF1 to promote full-thickness tissue regeneration. Cell Rep. 2018, 24, 3383–3392.e5. [Google Scholar] [CrossRef] [Green Version]
- Roura, S.; Pujal, J.-M.; Gálvez-Montón, C.; Bayes-Genis, A. The role and potential of umbilical cord blood in an era of new therapies: A review. Stem Cell Res. Ther. 2015, 6, 123. [Google Scholar] [CrossRef] [Green Version]
- Cuende, N.; Rasko, J.E.; Koh, M.B.; Dominici, M.; Ikonomou, L. Cell, tissue and gene products with marketing authorization in 2018 worldwide. Cytotherapy 2018, 20, 1401–1413. [Google Scholar] [CrossRef]
- Holm, J.S.; Toyserkani, N.M.; Sorensen, J.A. Adipose-derived stem cells for treatment of chronic ulcers: Current status. Stem Cell Res. Ther. 2018, 9, 142. [Google Scholar] [CrossRef] [Green Version]
- Moon, K.-C.; Suh, H.-S.; Kim, K.-B.; Han, S.-K.; Young, K.-W.; Lee, J.W.; Kim, M.H. Potential of allogeneic adipose-derived stem cell–hydrogel complex for treating diabetic foot ulcers. Diabetes 2019, 68, 837–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driver, V.R.; Lavery, L.A.; Reyzelman, A.M.; Dutra, T.G.; Dove, C.R.; Kotsis, S.V.; Kim, H.M.; Chung, K.C. A clinical trial of Integra Template for diabetic foot ulcer treatment. Wound Repair Regen. 2015, 23, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Murphy, C.M.; Haugh, M.G.; O’brien, F.J. The effect of mean pore size on cell attachment, proliferation and migration in collagen–glycosaminoglycan scaffolds for bone tissue engineering. Biomaterials 2010, 31, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Laiva, A.L.; O’Brien, F.J.; Keogh, M.B. SDF-1α gene-activated collagen scaffold drives functional differentiation of human Schwann cells for wound healing applications. Biotechnol. Bioeng. 2020. [Google Scholar] [CrossRef]
- Gallagher, K.A.; Liu, Z.-J.; Xiao, M.; Chen, H.; Goldstein, L.J.; Buerk, D.G.; Nedeau, A.; Thom, S.R.; Velazquez, O.C. Diabetic impairments in NO-mediated endothelial progenitor cell mobilization and homing are reversed by hyperoxia and SDF-1α. J. Clin. Investig. 2007, 117, 1249–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toksoy, A.; Müller, V.; Gillitzer, R.; Goebeler, M. Biphasic expression of stromal cell-derived factor-1 during human wound healing. Br. J. Dermatol. 2007, 157, 1148–1154. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guo, Y.; Chen, F.; Liu, J.; Jin, P. Stromal cell-derived factor-1 promotes human adipose tissue-derived stem cell survival and chronic wound healing. Exp. Ther. Med. 2016, 12, 45–50. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, F.J.; Harley, B.A.; Yannas, I.V.; Gibson, L. Influence of freezing rate on pore structure in freeze-dried collagen-GAG scaffolds. Biomaterials 2004, 25, 1077–1086. [Google Scholar] [CrossRef]
- Haugh, M.G.; Murphy, C.M.; O’Brien, F.J. Novel freeze-drying methods to produce a range of collagen–glycosaminoglycan scaffolds with tailored mean pore sizes. Tissue Eng. Part C Methods 2010, 16, 887–894. [Google Scholar] [CrossRef]
- Hoch, A.I.; Binder, B.Y.; Genetos, D.C.; Leach, J.K. Differentiation-dependent secretion of proangiogenic factors by mesenchymal stem cells. PLoS ONE 2012, 7, e35579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, S.; Koch, M.; Lee, Y.H.; Jung, F.; Blocki, A. An in vitro model of angiogenesis during wound healing provides insights into the complex role of cells and factors in the inflammatory and proliferation phase. Int. J. Mol. Sci. 2018, 19, 2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhardt, E.; Toksoy, A.; Goebeler, M.; Debus, S.; Bröcker, E.-B.; Gillitzer, R. Chemokines IL-8, GROα, MCP-1, IP-10, and Mig are sequentially and differentially expressed during phase-specific infiltration of leukocyte subsets in human wound healing. Am. J. Pathol. 1998, 153, 1849–1860. [Google Scholar] [CrossRef]
- Ju, L.; Zhou, Z.; Jiang, B.; Lou, Y.; Guo, X. Autocrine VEGF and IL-8 promote migration via Src/Vav2/Rac1/PAK1 signaling in human umbilical vein endothelial cells. Cell. Physiol. Biochem. 2017, 41, 1346–1359. [Google Scholar] [CrossRef]
- Hong, K.H.; Ryu, J.; Han, K.H. Monocyte chemoattractant protein-1–induced angiogenesis is mediated by vascular endothelial growth factor-A. Blood 2005, 105, 1405–1407. [Google Scholar] [CrossRef] [Green Version]
- Simone, T.M.; Higgins, P.J. Inhibition of SERPINE1 function attenuates wound closure in response to tissue injury: A role for PAI-1 in re-epithelialization and granulation tissue formation. J. Dev. Biol. 2015, 3, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Strawn, T.L.; Luo, M.; Wang, L.; Li, R.; Ren, M.; Xia, J.; Zhang, Z.; Ma, W.; Luo, T.; et al. Plasminogen activator inhibitor-1 inhibits angiogenic signaling by uncoupling vascular endothelial growth factor receptor-2-αVβ3 integrin cross talk. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Bugge, T.H.; Flick, M.J.; Danton, M.; Daugherty, C.C.; Romer, J.; Dano, K.; Carmeliet, P.; Collen, D.; Degen, J.L. Urokinase-type plasminogen activator is effective in fibrin clearance in the absence of its receptor or tissue-type plasminogen activator. Proc. Natl. Acad. Sci. USA 1996, 93, 5899–5904. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, V.; Jayaraman, P.-S.; Zaitsev, S.V.; Lebedeva, T.; Bdeir, K.; Kershaw, R.; Holman, K.R.; Parfyonova, Y.V.; Semina, E.V.; Beloglazova, I.B.; et al. Urokinase-type plasminogen activator (uPA) promotes angiogenesis by attenuating proline-rich homeodomain protein (PRH) transcription factor activity and de-repressing vascular endothelial growth factor (VEGF) receptor expression. J. Biol. Chem. 2016, 291, 15029–15045. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, G.; Cohen, T.; Gengrinovitch, S.; Poltorak, Z. Vascular endothelial growth factor (VEGF) and its receptors. FASEB J. 1999, 13, 9–22. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, K.; Liu, S.; Tsuji, T.; Olson, K.A.; Hu, G.-f. Endogenous angiogenin in endothelial cells is a general requirement for cell proliferation and angiogenesis. Oncogene 2005, 24, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobov, I.B.; Brooks, P.C.; Lang, R.A. Angiopoietin-2 displays VEGF-dependent modulation of capillary structure and endothelial cell survival in vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 11205–11210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akwii, R.G.; Sajib, M.S.; Zahra, F.T.; Mikelis, C.M. Role of Angiopoietin-2 in vascular physiology and pathophysiology. Cells 2019, 8, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brindle, N.P.; Saharinen, P.; Alitalo, K. Signaling and functions of angiopoietin-1 in vascular protection. Circ. Res. 2006, 98, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- Reed, M.J.; Koike, T.; Sadoun, E.; Sage, E.H.; Puolakkainen, P. Inhibition of TIMP1 enhances angiogenesis in vivo and cell migration in vitro. Microvasc. Res. 2003, 65, 9–17. [Google Scholar] [CrossRef]
- Vaalamo, M.; Leivo, T.; Saarialho-Kere, U. Differential expression of tissue inhibitors of metalloproteinases (TIMP-1,-2,-3, and-4) in normal and aberrant wound healing. Hum. Pathol. 1999, 30, 795–802. [Google Scholar] [CrossRef]
- Akahane, T.; Akahane, M.; Shah, A.; Connor, C.M.; Thorgeirsson, U.P. TIMP-1 inhibits microvascular endothelial cell migration by MMP-dependent and MMP-independent mechanisms. Exp. Cell Res. 2004, 301, 158–167. [Google Scholar] [CrossRef]
- Wietecha, M.S.; Król, M.J.; Michalczyk, E.R.; Chen, L.; Gettins, P.G.; DiPietro, L.A. Pigment epithelium-derived factor as a multifunctional regulator of wound healing. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H812–H826. [Google Scholar] [CrossRef] [Green Version]
- Streit, M.; Velasco, P.; Riccardi, L.; Spencer, L.; Brown, L.F.; Janes, L.; Lange-Asschenfeldt, B.; Yano, K.; Hawighorst, T.; Iruela-Arispe, L.; et al. Thrombospondin-1 suppresses wound healing and granulation tissue formation in the skin of transgenic mice. EMBO J. 2000, 19, 3272–3282. [Google Scholar] [CrossRef] [Green Version]
- Rohrs, J.A.; Sulistio, C.D.; Finley, S.D. Predictive model of thrombospondin-1 and vascular endothelial growth factor in breast tumor tissue. NPJ Syst. Biol. Appl. 2016, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rajagopal, S.; Kim, J.; Ahn, S.; Craig, S.; Lam, C.M.; Gerard, N.P.; Gerard, C.; Lefkowitz, R.J. β-arrestin-but not G protein-mediated signaling by the “decoy” receptor CXCR7. Proc. Natl. Acad. Sci. USA 2010, 107, 628–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitar, M.S. Diabetes Impairs Angiogenesis and Induces Endothelial Cell Senescence by Up-Regulating Thrombospondin-CD47-Dependent Signaling. Int. J. Mol. Sci. 2019, 20, 673. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, C.-T.; Cheng, H.-W.; Huang, C.-M.; Li, H.-R.; Ou, M.-H.; Huang, J.R.; Khoo, K.H.; Yu, H.W.; Chen, Y.Q.; Wang, Y.K.; et al. Fibronectin in cell adhesion and migration via N-glycosylation. Oncotarget 2017, 8, 70653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos-Lewis, W.; LaFever, K.S.; Page-McCaw, A. A scar-like lesion is apparent in basement membrane after wound repair in vivo. Matrix Biol. 2018, 74, 101–120. [Google Scholar] [CrossRef] [PubMed]
- Rennert, R.C.; Sorkin, M.; Januszyk, M.; Duscher, D.; Kosaraju, R.; Chung, M.T.; Lennon, J.; Radiya-Dixit, A.; Raghvendra, S.; Maan, Z.N.; et al. Diabetes impairs the angiogenic potential of adipose-derived stem cells by selectively depleting cellular subpopulations. Stem Cell Res. Ther. 2014, 5, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzhoyashvili, N.A.; Efimenko, A.Y.; Kochegura, T.N.; Kalinina, N.I.; Koptelova, N.V.; Sukhareva, O.Y.; Shestakova, M.V.; Akchurin, R.S.; Tkachuk, V.A.; Parfyonova, Y.V. Disturbed angiogenic activity of adipose-derived stromal cells obtained from patients with coronary artery disease and diabetes mellitus type 2. J. Transl. Med. 2014, 12, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Rebalka, I.A.; Raleigh, M.J.; D’Souza, D.M.; Coleman, S.K.; Rebalka, A.N.; Hawke, T.J. Inhibition of PAI-1 via PAI-039 improves dermal wound closure in diabetes. Diabetes 2015, 64, 2593–2602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, W.; Yang, C.; Dai, Z.; Che, D.; Feng, J.; Mao, Y.; Cheng, R.; Wang, Z.; He, X.; Zhou, T.; et al. High Levels of Pigment Epithelium–Derived Factor in Diabetes Impair Wound Healing Through Suppression of Wnt Signaling. Diabetes 2015, 64, 1407–1419. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.S.; Lip, G.Y.; Blann, A.D. Angiopoietin-1 and angiopoietin-2 in diabetes mellitus: Relationship to VEGF, glycaemic control, endothelial damage/dysfunction and atherosclerosis. Atherosclerosis 2005, 180, 113–118. [Google Scholar] [CrossRef]
- Lim, H.S.; Blann, A.D.; Chong, A.Y.; Freestone, B.; Lip, G.Y. Plasma vascular endothelial growth factor, angiopoietin-1, and angiopoietin-2 in diabetes: Implications for cardiovascular risk and effects of multifactorial intervention. Diabetes Care 2004, 27, 2918–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-R.; Kim, J.-W.; Jun, H.-S.; Roh, J.Y.; Lee, H.-Y.; Hong, I.S. Stem cell secretome and its effect on cellular mechanisms relevant to wound healing. Mol. Ther. 2018, 26, 606–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, L.; Landau, S.; Flugelman, M.Y.; Levenberg, S. Genetically engineered human muscle transplant enhances murine host neovascularization and myogenesis. Commun. Biol. 2018, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.; Jayaraman, V.; Huelsmann, E.J.; Bonish, B.; Burgad, D.; Sivaramakrishnan, G.; Qin, S.; DiPietro, L.A.; Zloza, A.; Zhang, C.; et al. Pro-inflammatory chemokine CCL2 (MCP-1) promotes healing in diabetic wounds by restoring the macrophage response. PLoS ONE 2014, 9, e91574. [Google Scholar] [CrossRef] [Green Version]
- Galkowska, H.; Wojewodzka, U.; Olszewski, W.L. Low recruitment of immune cells with increased expression of endothelial adhesion molecules in margins of the chronic diabetic foot ulcers. Wound Repair Regen. 2005, 13, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Ellison, D.D.; Suhail, Y.; Afzal, J.; Woo, L.; Kilic, O.; Spees, J.; Levchenko, A. Dynamic secretome of bone marrow-derived stromal cells reveals a cardioprotective biochemical cocktail. Proc. Natl. Acad. Sci. USA 2019, 116, 14374–14383. [Google Scholar]
- Kowalski, K.; Brzoska, E.; Ciemerych, M.A. The role of CXC receptors signaling in early stages of mouse embryonic stem cell differentiation. Stem Cell Res. 2019, 41, 101636. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, S.; Li, Y.; Wang, X.; Xue, W.; Ge, G.; Luo, X. The role of SDF-1-CXCR4/CXCR7 axis in the therapeutic effects of hypoxia-preconditioned mesenchymal stem cells for renal ischemia/reperfusion injury. PLoS ONE 2012, 7, e34608. [Google Scholar] [CrossRef]
- Wynn, R.F.; Hart, C.A.; Corradi-Perini, C.; O’Neill, L.; Evans, C.A.; Wraith, J.; Fairbairn, L.J.; Bellantuono, I. A small proportion of mesenchymal stem cells strongly expresses functionally active CXCR4 receptor capable of promoting migration to bone marrow. Blood 2004, 104, 2643–2645. [Google Scholar] [CrossRef]
- Wang, E.; Jarrah, A.; Benard, L.; Chen, J.; Schwarzkopf, M.; Hadri, L.; Tarzami, S.T. Deletion of CXCR4 in cardiomyocytes exacerbates cardiac dysfunction following isoproterenol administration. Gene Ther. 2014, 21, 496–506. [Google Scholar] [CrossRef] [Green Version]
- Albersen, M.; Berkers, J.; Dekoninck, P.; Deprest, J.; Lue, T.F.; Hedlund, P.; Lin, C.S.; Bivalacqua, T.J.; Van Poppel, H.; De Ridder, D.; et al. Expression of a distinct set of chemokine receptors in adipose tissue-derived stem cells is responsible for in vitro migration toward chemokines appearing in the major pelvic ganglion following cavernous nerve injury. Sex. Med. 2013, 1, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, F.; Müller, W.; Schütz, D.; Penfold, M.E.; Wong, Y.H.; Schulz, S.; Stumm, R. Rapid uptake and degradation of CXCL12 depend on CXCR7 carboxyl-terminal serine/threonine residues. J. Biol. Chem. 2012, 287, 28362–28377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisignoli, G.; Cristino, S.; Piacentini, A.; Cavallo, C.; Caplan, A.I.; Facchini, A. Hyaluronan-based polymer scaffold modulates the expression of inflammatory and degradative factors in mesenchymal stem cells: Involvement of Cd44 and Cd54. J. Cell. Physiol. 2006, 207, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Goessler, U.R.; Bugert, P.; Bieback, K.; Stern-Straeter, J.; Bran, G.; Hörmann, K.; Riedel, F. Integrin expression in stem cells from bone marrow and adipose tissue during chondrogenic differentiation. Int. J. Mol. Med. 2008, 21, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Popov, C.; Radic, T.; Haasters, F.; Prall, W.; Aszodi, A.; Gullberg, D.; Schieker, M.; Docheva, D. Integrins α2 β1 and α11 β1 regulate the survival of mesenchymal stem cells on collagen I. Cell Death Dis. 2011, 2, e186. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Xue, W.; Ge, G.; Luo, X.; Li, Y.; Xiang, H.; Ding, X.; Tian, P.; Tian, X. Hypoxic preconditioning advances CXCR4 and CXCR7 expression by activating HIF-1α in MSCs. Biochem. Biophys. Res. Commun. 2010, 401, 509–515. [Google Scholar] [CrossRef]
- Lee, Y.J.; Baek, S.E.; Lee, S.; Jeong, Y.J.; Kim, K.J.; Jun, Y.J.; Rhie, J.W. Wound-healing effect of adipose stem cell-derived extracellular matrix sheet on full-thickness skin defect rat model: Histological and immunohistochemical study. Int. Wound J. 2019, 16, 286–296. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.B.; Choi, J.; Cho, S.B.; Chung, J.Y.; Moon, E.S.; Kim, N.S.; Han, H.J. Topical embryonic stem cells enhance wound healing in diabetic rats. J. Orthop. Res. 2011, 29, 1554–1562. [Google Scholar] [CrossRef]
- Roy, D.C.; Mooney, N.A.; Raeman, C.H.; Dalecki, D.; Hocking, D.C. Fibronectin matrix mimetics promote full-thickness wound repair in diabetic mice. Tissue Eng. Part A 2013, 19, 2517–2526. [Google Scholar] [CrossRef] [Green Version]
- Hocking, D.C.; Brennan, J.R.; Raeman, C.H. A Small Chimeric Fibronectin Fragment accelerates dermal Wound repair in diabetic mice. Adv. Wound Care 2016, 5, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Z.; Kwon, A.-H.; Kamiyama, Y. Effects of plasma fibronectin on the healing of full-thickness skin wounds in streptozotocin-induced diabetic rats. J. Surg. Res. 2007, 138, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Damås, J.K.; Wæhre, T.; Yndestad, A.; Ueland, T.; Müller, F.; Eiken, H.G.; Holm, A.M.; Halvorsen, B.; Frøland, S.S.; Gullestad, L.; et al. Stromal Cell–Derived Factor-1α in Unstable Angina: Potential Antiinflammatory and Matrix-Stabilizing Effects. Circulation 2002, 106, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blommestein, H.; Verelst, S.; Huijgens, P.; Blijlevens, N.; Cornelissen, J.; Uyl-de Groot, C.A. Real-world costs of autologous and allogeneic stem cell transplantations for haematological diseases: A multicentre study. Ann. Hematol. 2012, 91, 1945–1952. [Google Scholar] [CrossRef] [PubMed]
- Crop, M.; Baan, C.; Korevaar, S.; Ijzermans, J.; Pescatori, M.; Stubbs, A.P.; Van IJcken, W.F.J.; Dahlke, M.H.; Eggenhofer, E.; Weimar, W.; et al. Inflammatory conditions affect gene expression and function of human adipose tissue-derived mesenchymal stem cells. Clin. Exp. Immunol. 2010, 162, 474–486. [Google Scholar] [CrossRef]
- Daina, L.G.; Sabău, M.; Daina, C.M.; Neamțu, C.; Tit, D.M.; Buhaș, C.L.; Bungau, C.; Aleya, L.; Bungau, S. Improving performance of a pharmacy in a Romanian hospital through implementation of an internal management control system. Sci. Total Environ. 2019, 675, 51–61. [Google Scholar] [CrossRef]
- Gofrit, O.N.; Benjamin, S.; Halachmi, S.; Leibovitch, I.; Dotan, Z.; Lamm, D.L.; Ehrlich, N.; Yutkin, V.; Ben-Am, M.; Hochberg, A. DNA based therapy with diphtheria toxin-A BC-819: A phase 2b marker lesion trial in patients with intermediate risk nonmuscle invasive bladder cancer. J. Urol. 2014, 191, 1697–1702. [Google Scholar] [CrossRef]
- Shishehbor, M.H.; Rundback, J.; Bunte, M.; Hammad, T.A.; Miller, L.; Patel, P.D.; Sadanandan, S.; Fitzgerald, M.; Pastore, J.; Kashyap, V.; et al. SDF-1 plasmid treatment for patients with peripheral artery disease (STOP-PAD): Randomized, double-blind, placebo-controlled clinical trial. Vasc. Med. 2019, 24, 200–207. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soluble Factor | Full Name | Function |
|---|---|---|
| IL-8 | Interleukin-8 | Recruitment of neutrophils; component of early inflammatory phase; angiogenic [53,54] |
| MCP-1 | Monocyte chemoattractant protein-1 | Recruitment of monocytes; essential for progression of healing; angiogenic [53,55] |
| PAI-1 | Plasminogen activator inhibitor-1 | Serine protease inhibitor; inhibits fibrinolysis; promotes re-epithelialization; anti-angiogenic [56,57] |
| uPA | Urokinase-type plasminogen activator | Mediates fibrinolysis; promotes angiogenesis [58,59] |
| VEGF | Vascular endothelial growth factor | Pro-angiogenic factor; stimulates vessel sprouting [60] |
| ANG | Angiogenin | Exerts ribonuclease activity; regulates VEGF-induced endothelial proliferation and angiogenesis [61] |
| Ang-2 | Angiopoietin-2 | Promotes vessel sprouting with VEGF; exerts vascular destabilizing effects [62,63] |
| Ang-1 | Angiopoietin-1 | Exerts vascular protective effects; essential for vessel stabilization [64] |
| TIMP-1 | Tissue inhibitor of metalloproteinase-1 | Anti-angiogenic; involved in the regeneration of epidermis and matrix remodeling [65,66,67] |
| PEDF | Pigment epithelium-derived factor | Potent anti-angiogenic factor; expression increases at late stage of wound healing; promotes resolution of angiogenesis [68] |
| TSP-1 | Thrombospondin-1 | Matricellular protein; inhibits angiogenesis; elevated expression suppresses wound healing [69,70] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laiva, A.L.; O’Brien, F.J.; Keogh, M.B. SDF-1α Gene-Activated Collagen Scaffold Restores Pro-Angiogenic Wound Healing Features in Human Diabetic Adipose-Derived Stem Cells. Biomedicines 2021, 9, 160. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020160
Laiva AL, O’Brien FJ, Keogh MB. SDF-1α Gene-Activated Collagen Scaffold Restores Pro-Angiogenic Wound Healing Features in Human Diabetic Adipose-Derived Stem Cells. Biomedicines. 2021; 9(2):160. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020160
Chicago/Turabian StyleLaiva, Ashang L., Fergal J. O’Brien, and Michael B. Keogh. 2021. "SDF-1α Gene-Activated Collagen Scaffold Restores Pro-Angiogenic Wound Healing Features in Human Diabetic Adipose-Derived Stem Cells" Biomedicines 9, no. 2: 160. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9020160