
Application of the OECT-Based In Vivo Biosensor Bioristor in Fruit Tree Monitoring to Improve Agricultural Sustainability
 , , , ,
, , , ,  and
and 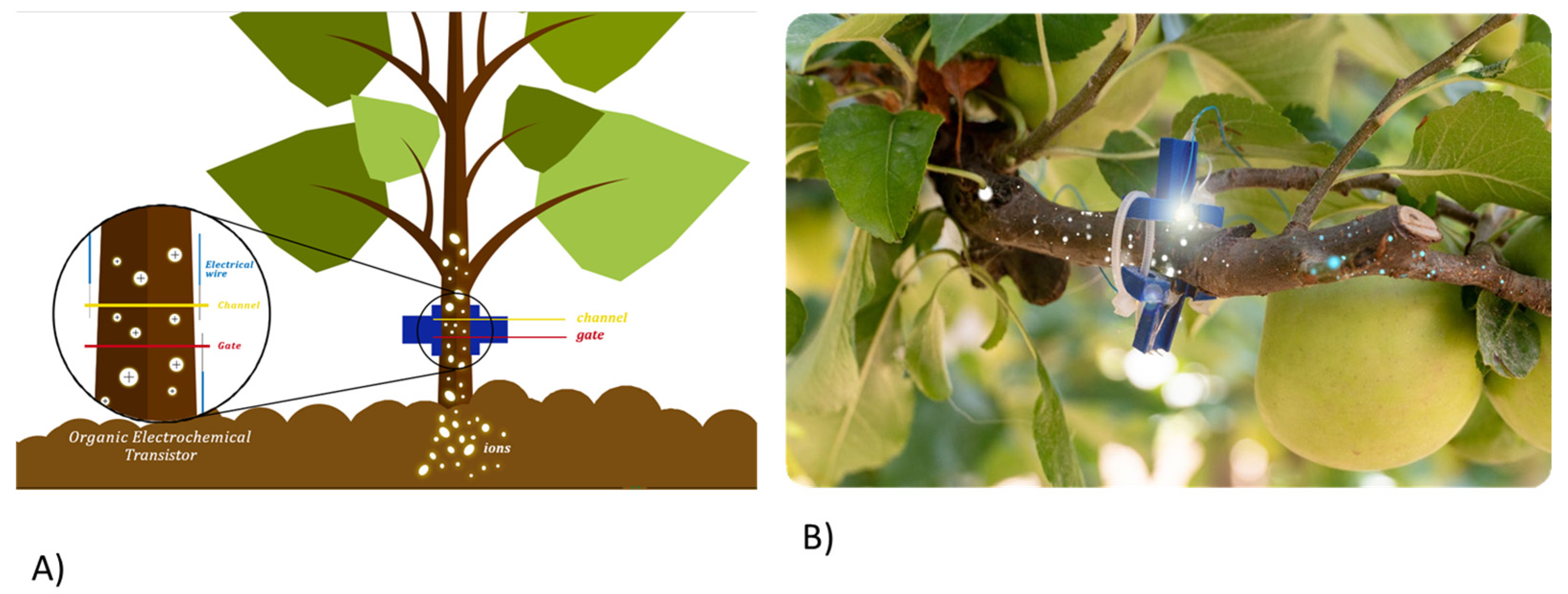{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. OECT Sensor Device: Bioristor Preparation and Insertion in Plant Trees
2.2. Measuring the Electric Activity of the Plants Using OECT
2.3. Bioristor Setup in Tree Crops
2.4. Bioristor Data Analysis
3. Results
Day/Night Modulation Transpiration and Ion Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devin, S.R.; Prudencio, Á.S.; Mahdavi, S.M.E.; Rubio, M.; Martínez-García, P.J.; Martínez-Gómez, P. Orchard Management and Incorporation of Biochemical and Molecular Strategies for Improving Drought Tolerance in Fruit Tree Crops. Plants 2023, 12, 773. [Google Scholar] [CrossRef]
- Fader, M.; Shi, S.; von Bloh, W.; Bondeau, A.; Cramer, W. Mediterranean Irrigation under Climate Change: More Efficient Irrigation Needed to Compensate for Increases in Irrigation Water Requirements. Hydrol. Earth Syst. Sci. 2016, 20, 953–973. [Google Scholar] [CrossRef] [Green Version]
- Tombesi, S.; Frioni, T.; Poni, S.; Palliotti, A. Effect of Water Stress “Memory” on Plant Behavior during Subsequent Drought Stress. Environ. Exp. Bot. 2018, 150, 106–114. [Google Scholar] [CrossRef]
- Wang, X.; Xing, Y. Evaluation of the Effects of Irrigation and Fertilization on Tomato Fruit Yield and Quality: A Principal Component Analysis. Sci. Rep. 2017, 7, 350. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.G.; Girona, J.; Gucci, R. Using Water for Best Product Quality in Fruit and Nut Trees and Vines. Irrig. Sci. 2023, 41, 449–452. [Google Scholar] [CrossRef]
- Du, T.; Kang, S.; Zhang, J.; Davies, W.J. Deficit Irrigation and Sustainable Water-Resource Strategies in Agriculture for China’s Food Security. J. Exp. Bot. 2015, 66, 2253–2269. [Google Scholar] [CrossRef]
- Chai, Q.; Gan, Y.; Zhao, C.; Xu, H.-L.; Waskom, R.M.; Niu, Y.; Siddique, K.H.M. Regulated Deficit Irrigation for Crop Production under Drought Stress. A Review. Agron. Sustain. Dev. 2016, 36, 3. [Google Scholar] [CrossRef] [Green Version]
- Scalisi, A. Continuous Determination of Fruit Tree Water-Status by Plant-Based Sensors. Italus Hortus 2018, 24, 39–50. [Google Scholar] [CrossRef]
- Costa, C.; Schurr, U.; Loreto, F.; Menesatti, P.; Carpentier, S. Plant Phenotyping Research Trends, a Science Mapping Approach. Front. Plant Sci. 2019, 9, 1933. [Google Scholar] [CrossRef] [Green Version]
- Dufil, G.; Bernacka-Wojcik, I.; Armada-Moreira, A.; Stavrinidou, E. Plant Bioelectronics and Biohybrids: The Growing Contribution of Organic Electronic and Carbon-Based Materials. Chem. Rev. 2022, 122, 4847–4883. [Google Scholar] [CrossRef]
- Valentini, R.; Belelli Marchesini, L.; Gianelle, D.; Sala, G.; Yarovslavtsev, A.; Vasenev, V.; Castaldi, S. New Tree Monitoring Systems: From Industry 4.0 to Nature 4.0. Ann. Silvic. Res. 2019, 43, 84–88. [Google Scholar] [CrossRef]
- Blanco, V. Microtensiometers: A New Tool to Monitor Your Apple Trees for Deciding When and How Much to Irrigate. Ph.D. Thesis, WSU Tree Fruit, Washington State University, Pullman, WA, USA, 2023. [Google Scholar]
- Gontia, N.K.; Tiwari, K.N. Development of Crop Water Stress Index of Wheat Crop for Scheduling Irrigation Using Infrared Thermometry. Agric. Water Manag. 2008, 95, 1144–1152. [Google Scholar] [CrossRef]
- Gracia-Romero, A.; Kefauver, S.C.; Fernandez-Gallego, J.A.; Vergara-Díaz, O.; Nieto-Taladriz, M.T.; Araus, J.L. UAV and Ground Image-Based Phenotyping: A Proof of Concept with Durum Wheat. Remote Sens. 2019, 11, 1244. [Google Scholar] [CrossRef] [Green Version]
- Blaya-Ros, P.J.; Blanco, V.; Domingo, R.; Soto-Valles, F.; Torres-Sánchez, R. Feasibility of Low-Cost Thermal Imaging for Monitoring Water Stress in Young and Mature Sweet Cherry Trees. Appl. Sci. 2020, 10, 5461. [Google Scholar] [CrossRef]
- Ahmad, U.; Alvino, A.; Marino, S. A Review of Crop Water Stress Assessment Using Remote Sensing. Remote Sens. 2021, 13, 4155. [Google Scholar] [CrossRef]
- Alizadeh, A.; Toudeshki, A.; Ehsani, R.; Migliaccio, K.; Wang, D. Detecting Tree Water Stress Using a Trunk Relative Water Content Measurement Sensor. Smart Agric. Technol. 2021, 1, 100003. [Google Scholar] [CrossRef]
- Noun, G.; Lo Cascio, M.; Spano, D.; Marras, S.; Sirca, C. Plant-Based Methodologies and Approaches for Estimating Plant Water Status of Mediterranean Tree Species: A Semi-Systematic Review. Agronomy 2022, 12, 2127. [Google Scholar] [CrossRef]
- Simbeye, D.S.; Mkiramweni, M.E.; Karaman, B.; Taskin, S. Plant Water Stress Monitoring and Control System. Smart Agric. Technol. 2023, 3, 100066. [Google Scholar] [CrossRef]
- Simon, D.T.; Gabrielsson, E.O.; Tybrandt, K.; Berggren, M. Organic Bioelectronics: Bridging the Signaling Gap between Biology and Technology. Chem. Rev. 2016, 116, 13009–13041. [Google Scholar] [CrossRef] [Green Version]
- Coppedè, N.; Janni, M.; Bettelli, M.; Maida, C.L.; Gentile, F.; Villani, M.; Ruotolo, R.; Iannotta, S.; Marmiroli, N.; Marmiroli, M.; et al. An in Vivo Biosensing, Biomimetic Electrochemical Transistor with Applications in Plant Science and Precision Farming. Sci. Rep. 2017, 7, 16195. [Google Scholar] [CrossRef] [Green Version]
- Janni, M.; Coppede, N.; Bettelli, M.; Briglia, N.; Petrozza, A.; Summerer, S.; Vurro, F.; Danzi, D.; Cellini, F.; Marmiroli, N.; et al. In Vivo Phenotyping for the Early Detection of Drought Stress in Tomato. Plant Phenomics 2019, 2019, 6168209. [Google Scholar] [CrossRef] [Green Version]
- Janni, M.; Claudia, C.; Federico, B.; Sara, P.; Filippo, V.; Nicola, C.; Manuele, B.; Davide, C.; Loreto, F.; Zappettini, A. Real-Time Monitoring of Arundo Donax Response to Saline Stress through the Application of in Vivo Sensing Technology. Sci. Rep. 2021, 11, 18598. [Google Scholar] [CrossRef]
- Amato, D.; Montanaro, G.; Vurro, F.; Coppedé, N.; Briglia, N.; Petrozza, A.; Janni, M.; Zappettini, A.; Cellini, F.; Nuzzo, V. Towards In Vivo Monitoring of Ions Accumulation in Trees: Response of an in Planta Organic Electrochemical Transistor Based Sensor to Water Flux Density, Light and Vapor Pressure Deficit Variation. Appl. Sci. 2021, 11, 4729. [Google Scholar] [CrossRef]
- Gentile, F.; Vurro, F.; Janni, M.; Manfredi, R.; Cellini, F.; Petrozza, A.; Zappettini, A.; Coppedè, N. A Biomimetic, Biocompatible OECT Sensor for the Real-Time Measurement of Concentration and Saturation of Ions in Plant Sap. Adv. Electron. Mater. 2022, 8, 2200092. [Google Scholar] [CrossRef]
- Vurro, F.; Janni, M.; Coppedè, N.; Gentile, F.; Manfredi, R.; Bettelli, M.; Zappettini, A. Development of an In Vivo Sensor to Monitor the Effects of Vapour Pressure Deficit (VPD) Changes to Improve Water Productivity in Agriculture. Sensors 2019, 19, 4667. [Google Scholar] [CrossRef] [Green Version]
- Manfredi, R.; Vurro, F.; Janni, M.; Bettelli, M.; Gentile, F.; Zappettini, A.; Coppedè, N. Long-Term Stability in Electronic Properties of Textile Organic Electrochemical Transistors for Integrated Applications. Materials 2023, 16, 1861. [Google Scholar] [CrossRef]
- Coppedè, N.; Villani, M.; Gentile, F. Diffusion Driven Selectivity in Organic Electrochemical Transistors. Sci. Rep. 2014, 4, 4297. [Google Scholar] [CrossRef] [Green Version]
- Bernards, D.A.; Malliaras, G.G. Steady-State and Transient Behavior of Organic Electrochemical Transistors. Adv. Funct. Mater. 2007, 17, 3538–3544. [Google Scholar] [CrossRef]
- Gentile, F.; Vurro, F.; Picelli, F.; Bettelli, M.; Zappettini, A.; Coppedè, N. A Mathematical Model of OECTs with Variable Internal Geometry. Sens. Actuators Phys. 2020, 304, 111894. [Google Scholar] [CrossRef]
- Finco, A.; Bentivoglio, D.; Chiaraluce, G.; Alberi, M.; Chiarelli, E.; Maino, A.; Mantovani, F.; Montuschi, M.; Raptis, K.G.C.; Semenza, F.; et al. Combining Precision Viticulture Technologies and Economic Indices to Sustainable Water Use Management. Water 2022, 14, 1493. [Google Scholar] [CrossRef]
- Montanaro, G.; Dichio, B.; Xiloyannis, C.; Lang, A. Fruit Transpiration in Kiwifruit: Environmental Drivers and Predictive Model. AoB Plants 2012, 2012, pls036. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.; Manfrini, L.; Venturi, M.; Corelli Grappadelli, L.; Morandi, B. Fruit Transpiration Drives Interspecific Variability in Fruit Growth Strategies. Hortic. Res. 2022, 9, uhac036. [Google Scholar] [CrossRef]
- Oliveira, C.P.M.D.; Simões, W.L.; Silva, J.A.B.D.; Faria, G.A.; Lopes, P.R.C.; Amorim, M.D.N. Physiological and Biochemical Responses of Apple Trees to Irrigation Water Depth in a Semiarid Region of Brazil. Ciênc. E Agrotecnol. 2020, 44, e015620. [Google Scholar] [CrossRef]
- Tombesi, S.; Nardini, A.; Frioni, T.; Soccolini, M.; Zadra, C.; Farinelli, D.; Poni, S.; Palliotti, A. Stomatal Closure Is Induced by Hydraulic Signals and Maintained by ABA in Drought-Stressed Grapevine. Sci. Rep. 2015, 5, 12449. [Google Scholar] [CrossRef] [Green Version]
- Danzi, D.; Briglia, N.; Petrozza, A.; Summerer, S.; Povero, G.; Stivaletta, A.; Cellini, F.; Pignone, D.; De Paola, D.; Janni, M. Can High Throughput Phenotyping Help Food Security in the Mediterranean Area? Front. Plant Sci. 2019, 10, 15. [Google Scholar] [CrossRef]
- Liu, H.; Song, S.; Zhang, H.; Li, Y.; Niu, L.; Zhang, J.; Wang, W. Signaling Transduction of ABA, ROS, and Ca2+ in Plant Stomatal Closure in Response to Drought. Int. J. Mol. Sci. 2022, 23, 14824. [Google Scholar] [CrossRef]
- Boini, A.; Cavallina, L.; Perulli, G.; Bresilla, K.; Bortolotti, G.; Morandi, B.; Corelli Grappadelli, L.; Manfrini, L. Actinidia Chinensis: Physiological and Productive Performance under Water Stress Condition. Acta Hortic. 2022, 43–50. [Google Scholar] [CrossRef]
- Dayer, S.; Herrera, J.C.; Dai, Z.; Burlett, R.; Lamarque, L.J.; Delzon, S.; Bortolami, G.; Cochard, H.; Gambetta, G.A. Nighttime Transpiration Represents a Negligible Part of Water Loss and Does Not Increase the Risk of Water Stress in Grapevine. Plant Cell Environ. 2021, 44, 387–398. [Google Scholar] [CrossRef]
- Morandi, B.; Manfrini, L.; Losciale, P.; Zibordi, M.; Corelli Grappadelli, L. Changes in Vascular and Transpiration Flows Affect the Seasonal and Daily Growth of Kiwifruit (Actinidia deliciosa) Berry. Ann. Bot. 2010, 105, 913–923. [Google Scholar] [CrossRef] [Green Version]
- Morandi, B.; Losciale, P.; Manfrini, L.; Zibordi, M.; Anconelli, S.; Pierpaoli, E.; Corelli Grappadelli, L. Leaf Gas Exchanges and Water Relations Affect the Daily Patterns of Fruit Growth and Vascular Flows in Abbé Fétel Pear (Pyrus communis L.) Trees. Sci. Hortic. 2014, 178, 106–113. [Google Scholar] [CrossRef]
- Ortega-Farias, S.; Meza, S.E.; López-Olivari, R.; Araya-Alman, M.; Carrasco-Benavides, M. Effects of Four Irrigation Regimes on Yield, Fruit Quality, Plant Water Status, and Water Productivity in a Furrow-Irrigated Red Raspberry Orchard. Agric. Water Manag. 2022, 273, 107885. [Google Scholar] [CrossRef]
- Jones, H.G. Monitoring Plant and Soil Water Status: Established and Novel Methods Revisited and Their Relevance to Studies of Drought Tolerance. J. Exp. Bot. 2007, 58, 119–130. [Google Scholar] [CrossRef] [Green Version]
- Canaj, K.; Parente, A.; D’Imperio, M.; Boari, F.; Buono, V.; Toriello, M.; Mehmeti, A.; Montesano, F.F. Can Precise Irrigation Support the Sustainability of Protected Cultivation? A Life-Cycle Assessment and Life-Cycle Cost Analysis. Water 2022, 14, 6. [Google Scholar] [CrossRef]
- Buono, V.; Mastroleo, M.; Lucchi, C.; D’Amato, G.; Manfrini, L.; Morandi, B. Field-Testing of a Decision Support System (DSS) to Optimize Irrigation Management of Kiwifruit in Italy: A Comparison with Current Farm Management. Acta Hortic. 2022, 355–362. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vurro, F.; Marchetti, E.; Bettelli, M.; Manfrini, L.; Finco, A.; Sportolaro, C.; Coppedè, N.; Palermo, N.; Tommasini, M.G.; Zappettini, A.; et al. Application of the OECT-Based In Vivo Biosensor Bioristor in Fruit Tree Monitoring to Improve Agricultural Sustainability. Chemosensors 2023, 11, 374. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors11070374
Vurro F, Marchetti E, Bettelli M, Manfrini L, Finco A, Sportolaro C, Coppedè N, Palermo N, Tommasini MG, Zappettini A, et al. Application of the OECT-Based In Vivo Biosensor Bioristor in Fruit Tree Monitoring to Improve Agricultural Sustainability. Chemosensors. 2023; 11(7):374. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors11070374
Chicago/Turabian StyleVurro, Filippo, Edoardo Marchetti, Manuele Bettelli, Luigi Manfrini, Adele Finco, Carlo Sportolaro, Nicola Coppedè, Nadia Palermo, Maria Grazia Tommasini, Andrea Zappettini, and et al. 2023. "Application of the OECT-Based In Vivo Biosensor Bioristor in Fruit Tree Monitoring to Improve Agricultural Sustainability" Chemosensors 11, no. 7: 374. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors11070374