Xylem Plasticity in Pinus pinaster and Quercus ilex Growing at Sites with Different Water Availability in the Mediterranean Region: Relations between Intra-Annual Density Fluctuations and Environmental Conditions



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Species and Study Site
2.2. Tree-ring Data and IADF Frequency
2.3. Microscopy and Quantitative Wood Anatomy
2.4. Climate Signal
3. Results
3.1. Tree-Ring Dating and IADFs Occurrence
3.2. Xylem Traits
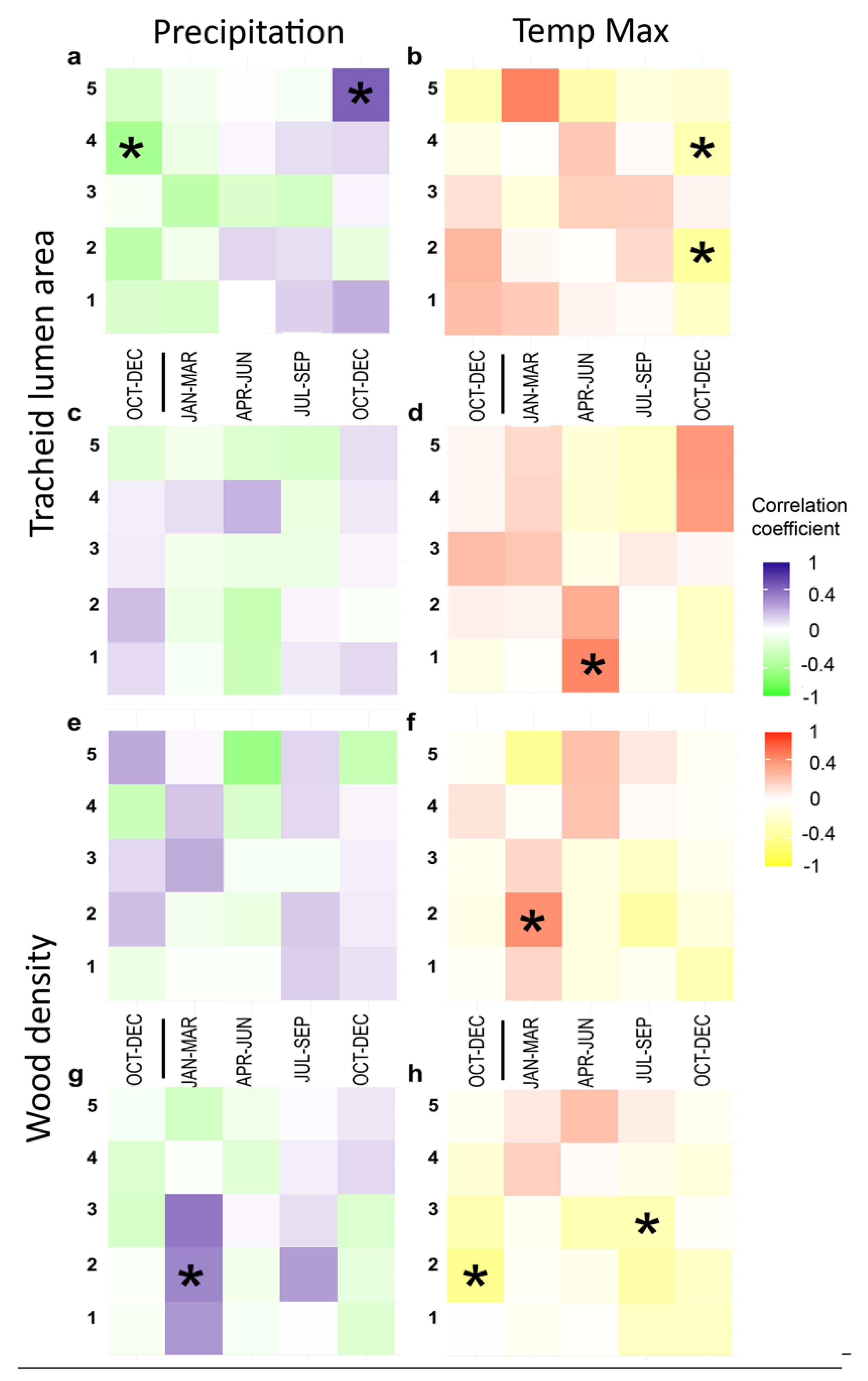
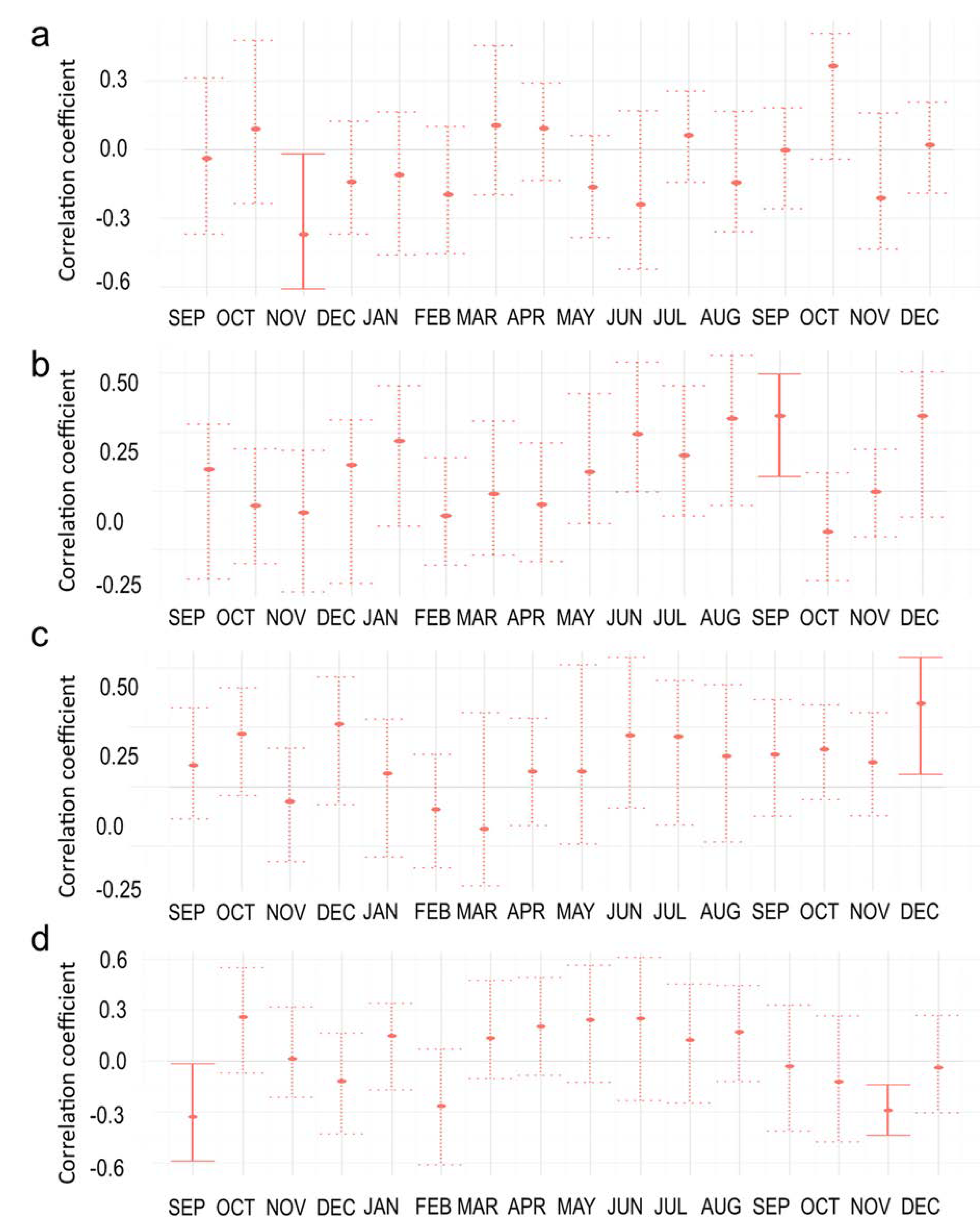
3.3. Climate Correlations
4. Discussion
4.1. The Role of Water Availability in P. pinaster
4.2. IADF Formation in Q. ilex
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sarris, D.; Christodoulakis, D.; Körner, C. Recent decline in precipitation and tree growth in the eastern Mediterranean. Glob. Chang. Biol. 2007, 13, 1187–1200. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Viguera, B.; Cabrera, M.; Cañellas, I. Drought induced decline could portend widespread pine mortality at the xeric ecotone in managed mediterranean pine-oak woodlands. For. Ecol. Manag. 2014, 320, 70–82. [Google Scholar] [CrossRef]
- De Micco, V.; Carrer, M.; Rathgeber, C.B.; Camarero, J.J.; Voltas, J.; Cherubini, P.; Battipaglia, G. From xylogenesis to tree rings: Wood traits to investigate tree response to environmental changes. IAWA J. 2019, 40, 155–1822. [Google Scholar] [CrossRef] [Green Version]
- Fahn, A.; Werker, E.; Baas, P. Wood Anatomy and Identification of Trees and Shrubs from Israel and Adjacent Regions; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1986. [Google Scholar]
- Carlquist, S. Adaptive wood anatomy of chaparral shrubs. In The California Chaparral: Paradigms re-Examine; Los Angeles Country Museum of Natural History Contributions: Los Angeles, CA, USA, 1989; pp. 25–35. [Google Scholar]
- De Micco, V.; Aronne, G.; Baas, P. Wood anatomy and hydraulic architecture of stems and twigs of some Mediterranean trees and shrubs along a mesic-xeric gradient. Trees 2008, 22, 643–655. [Google Scholar] [CrossRef]
- Tyree, M.T.; Davis, S.D.; Cochard, H. Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? IAWA J. 1994, 15, 335–360. [Google Scholar] [CrossRef] [Green Version]
- Sperry, J.S.; Meinzer, F.C.; McCulloh, K.A. Safety and efficiency conflicts in hydraulic architecture: Scaling from tissues to trees. Plant Cell Environ. 2008, 31, 632–645. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef]
- Meinzer, F.C.; McCulloh, K.A.; Lachenbruch, B.; Woodruff, D.R.; Johnson, D.M. The blind men and the elephant: The impact of context and scale in evaluating conflicts between plant hydraulic safety and efficiency. Oecologia 2010, 164, 287–296. [Google Scholar] [CrossRef]
- Vaganov, E.A.; Shiyatov, S.G.; Mazepa, V.S. Dendroclimatic Study in Ural-Siberian Subarctic; Publishing House SB RAS Nauka: Novosibirsk, Russia, 1996. [Google Scholar]
- Nicault, A.; Rathgeber, C.; Tessier, L.; Thomas, A. Intra-annual variations of radial growth and ring structure. Ann. For. Sci. 2001, 58, 769–784. [Google Scholar] [CrossRef] [Green Version]
- De Micco, V.; Saurer, M.; Aronne, G.; Tognetti, R.; Cherubini, P. Variations of wood anatomy and δ13C within-tree rings of coastal Pinus pinaster showing intra-annual density fluctuations. IAWA J. 2007, 28, 61–74. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Griçar, J.; Seo, J.W.; Rathgeber, C.B.; Anfodillo, T.; Jalkanen, R. Critical temperatures for xylogenesis in conifers of cold climates. Glob. Ecol. Biogeogr. 2008, 17, 696–707. [Google Scholar] [CrossRef]
- De Micco, V.; Campelo, F.; De Luis, M.; Bräuning, A.; Grabner, M.; Battipaglia, G.; Cherubini, P. Intra-annual density fluctuations in tree rings: How, when, where, and why? IAWA J. 2016, 37, 232–259. [Google Scholar] [CrossRef]
- Beeckman, H. Wood anatomy and trait-based ecology. IAWA J. 2016, 37, 127–151. [Google Scholar] [CrossRef]
- Rigling, A.; Waldner, P.O.; Forster, T.; Bräker, O.U.; Pouttu, A. Ecological interpretation of tree-ring width and intraannual density fluctuations in Pinus sylvestris on dry sites in the central Alps and Siberia. Can. J. For. Res. 2001, 31, 18–31. [Google Scholar] [CrossRef]
- Campelo, F.; Nabais, C.; Freitas, H.; Gutiérrez, E. Climatic significance of tree-ring width and intra-annual density fluctuations in Pinus pinea from a dry Mediterranean area in Portugal. Ann. For. Sci. 2007, 64, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Battipaglia, G.; De Micco, V.; Brand, W.A.; Linke, P.; Aronne, G.; Saurer, M.; Cherubini, P. Variations of vessel diameter and δ13C in false rings of Arbutus unedo L. reflect different environmental conditions. New Phytol. 2010, 188, 1099–1112. [Google Scholar] [CrossRef] [Green Version]
- de Luis, M.; Novak, K.; Raventós, J.; Gričar, J.; Prislan, P.; Čufar, K. Cambial activity, wood formation and sapling survival of Pinus halepensis exposed to different irrigation regimes. For. Ecol. Manag. 2011, 262, 1630–1638. [Google Scholar] [CrossRef]
- de Luis, M.; Novak, K.; Raventós, J.; Gričar, J.; Prislan, P.; Čufar, K. Climate factors promoting intra-annual density fluctuations in Aleppo pine (Pinus halepensis) from semiarid sites. Dendrochronologia 2011, 29, 163–169. [Google Scholar] [CrossRef]
- Battipaglia, G.; De Micco, V.; Brand, W.A.; Saurer, M.; Aronne, G.; Linke, P.; Cherubini, P. Drought impact on water use efficiency and intra-annual density fluctuations in Erica arborea on Elba (Italy). Plant Cell Environ. 2014, 37, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Novak, K.; de Luis, M.; Raventós, J.; Čufar, K. Climatic signals in tree-ring widths and wood structure of Pinus halepensis in contrasted environmental conditions. Trees 2013, 27, 927–936. [Google Scholar] [CrossRef]
- Novak, K.; Sánchez, M.A.S.; Čufar, K.; Raventós, J.; de Luis, M. Age, climate and intra-annual density fluctuations in in Spain. IAWA J. 2013, 34, 459–474. [Google Scholar] [CrossRef] [Green Version]
- Nabais, C.; Campelo, F.; Vieira, J.; Cherubini, P. Climatic signals of tree-ring width and intra-annual density fluctuations in Pinus pinaster and Pinus pinea along a latitudinal gradient in Portugal. Forestry 2014, 87, 598–605. [Google Scholar] [CrossRef]
- Zalloni, E.; de Luis, M.; Campelo, F.; Novak, K.; De Micco, V.; Di Filippo, A.; Battipaglia, G. Climatic signals from intra-annual density fluctuation frequency in Mediterranean pines at a regional scale. Front. Plant Sci. 2016, 7, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; Saurer, M.; De Micco, V. Contrasting physiological responses to Mediterranean climate variability are revealed by intra-annual density fluctuations in tree rings of Quercus ilex L. and Pinus pinea L. Tree Physiol. 2018, 38, 1213–1224. [Google Scholar] [CrossRef]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; De Micco, V. Site conditions influence the climate signal of intra-annual density fluctuations in tree rings of Q. ilex L. Ann. For. Sci. 2018, 75, 68. [Google Scholar] [CrossRef] [Green Version]
- Castagneri, D.; Battipaglia, G.; von Arx, G.; Pacheco, A.; Carrer, M. Tree-ring anatomy and carbon isotope ratio show both direct and legacy effects of climate on bimodal xylem formation in Pinus pinea. Tree Physiol. 2018, 38, 1098–1109. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Yepez, E.A. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Fonti, P.; von Arx, G.; García-González, I.; Eilmann, B.; Sass-Klaassen, U.; Gärtner, H.; Eckstein, D. Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol. 2010, 185, 42–53. [Google Scholar] [CrossRef]
- Battipaglia, G.; Campelo, F.; Vieira, J.; Grabner, M.; De Micco, V.; Nabais, C.; Cherubini, P.; Carrer, M.; Braeuning, A.; Cufar, K.; et al. Structure and Function of Intra–Annual Density Fluctuations: Mind the Gaps. Front. Plant Sci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Campelo, F.; Nabais, C.; Gutiérrez, E.; Freitas, H.; García-González, I. Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. Trees 2010, 24, 463–470. [Google Scholar] [CrossRef]
- De Micco, V.; Battipaglia, G.; Balzano, A.; Cherubini, P.; Aronne, G. Are wood fibres as sensitive to environmental conditions as vessels in tree rings with intra-annual density fluctuations (IADFs) in Mediterranean species? Trees 2016, 30, 971–983. [Google Scholar] [CrossRef]
- Balzano, A.; Battipaglia, G.; De Micco, V. Wood-trait analysis to understand climatic factors triggering intra-annual density-fluctuations in co-occurring Mediterranean trees. IAWA J. 2019, 40, 241–258. [Google Scholar] [CrossRef] [Green Version]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees 2004, 18, 83–92. [Google Scholar] [CrossRef]
- Sass, U.; Eckstein, D. The variability of vessel size in beech (Fagus sylvatica L.) and its ecophysiological interpretation. Trees 1995, 9, 247–252. [Google Scholar] [CrossRef]
- Abrantes, J.; Campelo, F.; García-González, I.; Nabais, C. Environmental control of vessel traits in Quercus ilex under Mediterranean climate: Relating xylem anatomy to function. Trees 2013, 27, 655–662. [Google Scholar] [CrossRef]
- Schume, H.; Grabner, M.; Eckmüllner, O. The influence of an altered groundwater regime on vessel properties of hybrid poplar. Trees 2004, 18, 184–194. [Google Scholar] [CrossRef]
- Grabner, M. Functional Tree-Ring Analysis: Wood as an Information Source to Understand Physiological, Environmental and Technological Questions. Ph.D. Thesis, University of Natural Resources and Life Sciences, Vienna, Austria, 2005. [Google Scholar]
- De Micco, V.; Battipaglia, G.; Brand, W.A.; Linke, P.; Saurer, M.; Aronne, G.; Cherubini, P. Discrete versus continuous analysis of anatomical and δ13C variability in tree rings with intra-annual density fluctuations. Trees 2012, 26, 513–524. [Google Scholar] [CrossRef]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Braeker, O.U.; Schoch, W.; Innes, J.L. Identification, measurement and interpretation of tree rings in woody species from Mediterranean climates. Biol. Rev. 2003, 78, 119–148. [Google Scholar] [CrossRef] [Green Version]
- Campelo, F.; Gutiérrez, E.; Ribas, M.; Nabais, C.; Freitas, H. Relationships between climate and double rings in Quercus ilex from northeast Spain. Can. J. For. Res. 2007, 37, 1915–1923. [Google Scholar] [CrossRef]
- Vieira, J.; Campelo, F.; Nabais, C. Age-dependent responses of tree-ring growth and intra-annual density fluctuations of Pinus pinaster to Mediterranean climate. Trees 2009, 23, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Rozas, V.; García-González, I.; Zas, R. Climatic control of intra-annual wood density fluctuations of Pinus pinaster in NW Spain. Trees 2011, 25, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Campelo, F.; Vieira, J.; Nabais, C. Tree-ring growth and intra-annual density fluctuations of Pinus pinaster responses to climate: Does size matter? Trees 2013, 27, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Wigley, T.M.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Stokes, M.; Smiley, T. An Introduction to Tree-Ring Dating; Reprinted 1996; University of Arizona Press: Tucson, AZ, USA, 1968. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Osborn, T.J.; Biffa, K.R.; Jones, P.D. Adjusting variance for sample-size in tree-ring chronologies and other regional-mean timeseries. Dendrochronologia 1997, 15, 89–99. [Google Scholar]
- Cirillo, C.; De Micco, V.; Rouphael, Y.; Balzano, A.; Caputo, R.; De Pascale, S. Morpho-anatomical and physiological traits of two Bougainvillea genotypes trained to two shapes under deficit irrigation. Trees 2017, 31, 173–187. [Google Scholar] [CrossRef] [Green Version]
- Van der Werf, G.W.; Sass-Klaassen, U.G.; Mohren, G.M.J. The impact of the 2003 summer drought on the intra-annual growth pattern of beech (Fagus sylvatica L.) and oak (Quercus robur L.) on a dry site in the Netherlands. Dendrochronologia 2007, 25, 103–112. [Google Scholar] [CrossRef]
- Venables, W.N.; Smith, D.M. R Development Core Team (2008) an Introduction to R: Notes on R, a Programming Environment for Data Analysis and Graphics; Faculty of Social Sciences—University of Ljubljana: Ljubljana, Slovenia, 2009. [Google Scholar]
- Denne, M.P. Definition of latewood according to Mork (1928). IAWA J. 1989, 10, 59–62. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Tardif, J.C.; Conciatori, F. Influence of climate on tree rings and vessel features in red oak and white oak growing near their northern distribution limit, southwestern Quebec, Canada. Can. J. For. Res. 2006, 36, 2317–2330. [Google Scholar] [CrossRef] [Green Version]
- Kames, S.; Tardif, J.C.; Bergeron, Y. Anomalous earlywood vessel lumen area in blackash (Fraxinus nigra Marsh.) tree rings as a potential indicator of forest fires. Dendrochronologia 2011, 29, 109–114. [Google Scholar] [CrossRef]
- Fonti, P.; García-González, I. Earlywood vessel size of oak as a potential proxy for spring precipitation in mesic sites. J. Biogeogr. 2008, 35, 2249–2257. [Google Scholar] [CrossRef]
- Bogino, S.; Fernandez Nieto, M.J.; Bravo, F. Climate effect on radial growth of Pinus sylvestris at its southern and western distribution limits. Silva Fennica 2009, 43, 609–623. [Google Scholar] [CrossRef] [Green Version]
- Niccoli, F.; Esposito, A.; Altieri, S.; Battipaglia, G. Fire severity influences ecophysiological responses of Pinus pinaster Ait. Front. Plant Sci. 2019. [Google Scholar] [CrossRef]
- Vieira, J.; Rossi, S.; Campelo, F.; Nabais, C. Are neighboring trees in tune? Wood formation in Pinus pinaster. Eur. J. For. Res. 2014, 133, 41–50. [Google Scholar] [CrossRef]
- Wimmer, R.; Downes, G.M.; Evans, R. High-resolution analysis of radial growth and wood density in Eucalyptus nitens, grown under different irrigation regimes. Ann. For. Sci. 2002, 59, 519–524. [Google Scholar] [CrossRef] [Green Version]
- Campelo, F.; Vieira, J.; Battipaglia, G.; de Luis, M.; Nabais, C.; Freitas, H.; Cherubini, P. Which matters most for the formation of intra-annual density fluctuations in Pinus pinaster: Age or size? Trees 2015, 29, 237–245. [Google Scholar] [CrossRef]
- Battipaglia, G.; Savi, T.; Ascoli, D.; Castagneri, D.; Esposito, A.; Mayr, S.; Nardini, A. Effects of prescribed burning on ecophysiological, anatomical and stem hydraulic properties in Pinus pinea L. Tree Physiol. 2016, 36, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Onelli, E.; Scippa, G.S.; Chiatante, D. Seasonality of fine root dynamics and activity of root and shoot vascular cambium in a Quercus ilex L. forest (Italy). For. Ecol. Manag. 2019, 431, 26–34. [Google Scholar] [CrossRef]
- Nabais, C.; Freitas, H.; Hagemeyer, J. Tree-rings to climate relationships of Quercus ilex L. in the NE-Portugal. Dendrochronologia 1998–1999, 16–17, 37–44. [Google Scholar]
- Cartenì, F.; Deslauriers, A.; Rossi, S.; Morin, H.; De Micco, V.; Mazzoleni, S.; Giannino, F. The physiological mechanisms behind the earlywood-to-latewood transition: A process-based modeling approach. Front. Plant Sci. 2018, 9, 1053. [Google Scholar] [CrossRef] [PubMed]
- Domec, J.C.; Gartner, B.L. How do water transport and water storage differ in coniferous earlywood and latewood? J. Exp. Bot. 2002, 53, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deslauriers, A.; Rossi, S.; Anfodillo, T.; Saracino, A. Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol. 2008, 28, 863–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balzano, A.; Battipaglia, G.; Cherubini, P.; De Micco, V. Xylem Plasticity in Pinus pinaster and Quercus ilex Growing at Sites with Different Water Availability in the Mediterranean Region: Relations between Intra-Annual Density Fluctuations and Environmental Conditions. Forests 2020, 11, 379. https://0-doi-org.brum.beds.ac.uk/10.3390/f11040379
Balzano A, Battipaglia G, Cherubini P, De Micco V. Xylem Plasticity in Pinus pinaster and Quercus ilex Growing at Sites with Different Water Availability in the Mediterranean Region: Relations between Intra-Annual Density Fluctuations and Environmental Conditions. Forests. 2020; 11(4):379. https://0-doi-org.brum.beds.ac.uk/10.3390/f11040379
Chicago/Turabian StyleBalzano, Angela, Giovanna Battipaglia, Paolo Cherubini, and Veronica De Micco. 2020. "Xylem Plasticity in Pinus pinaster and Quercus ilex Growing at Sites with Different Water Availability in the Mediterranean Region: Relations between Intra-Annual Density Fluctuations and Environmental Conditions" Forests 11, no. 4: 379. https://0-doi-org.brum.beds.ac.uk/10.3390/f11040379