

Evaluation of Mutagenicity and Anti-Mutagenicity of Various Bean Milks Using Drosophila with High Bioactivation
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Preparation
2.3. Somatic Mutation and Recombination Test (SMART) or Wing Spot Test
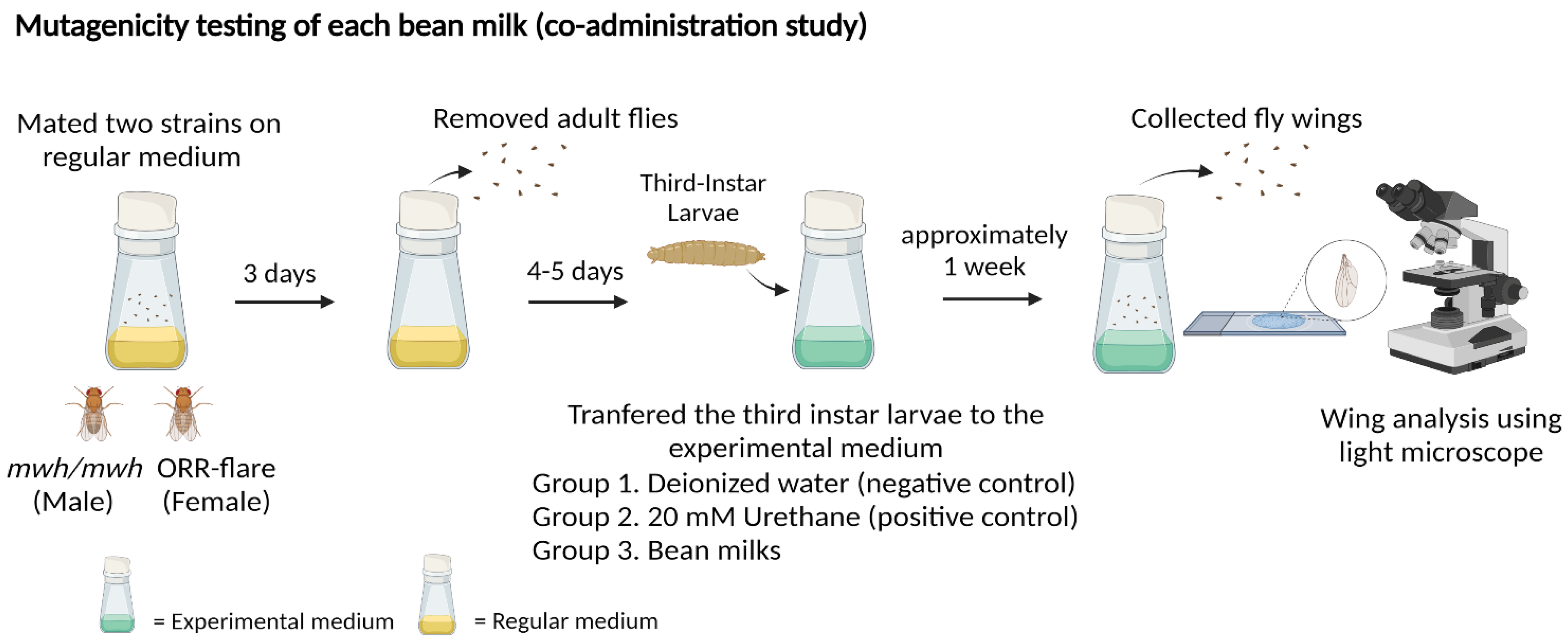
2.3.1. Mutagenic Evaluation of Bean Milks Using the Co-Administration Study
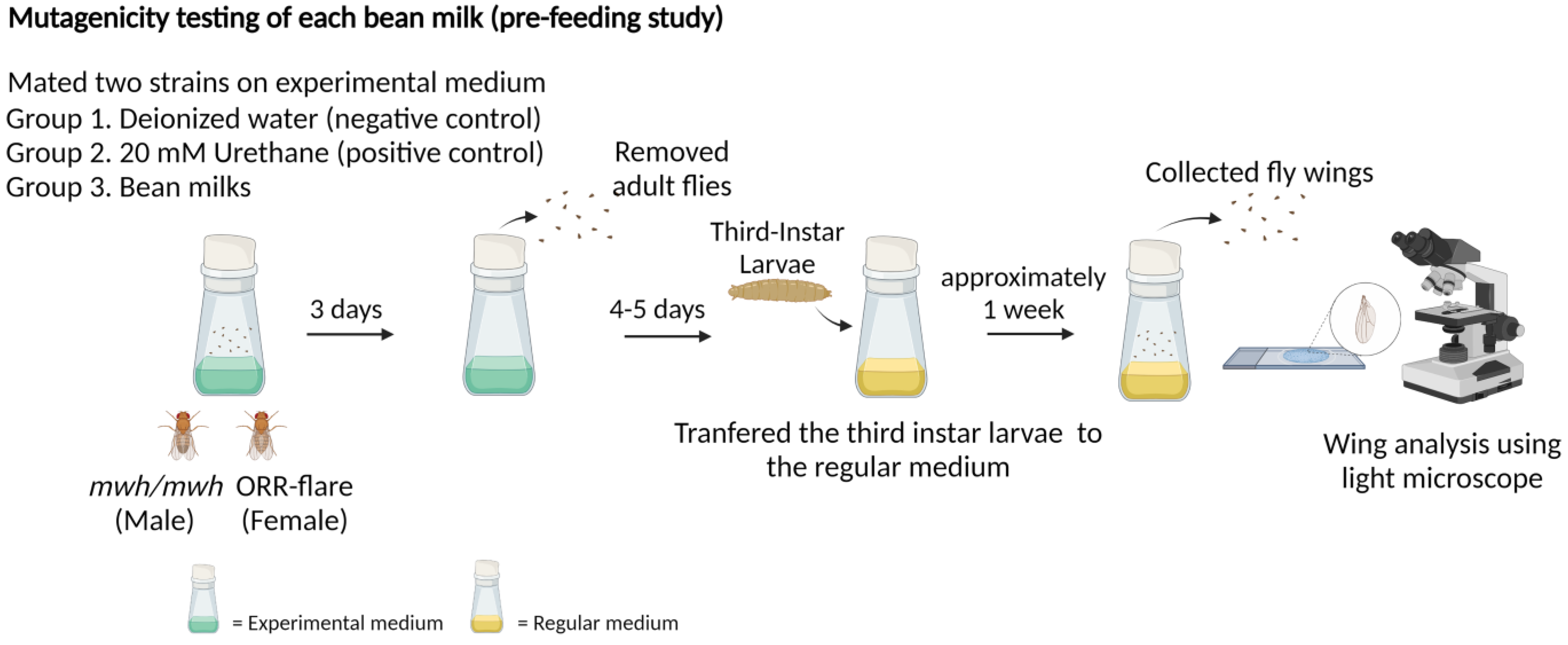
2.3.2. Mutagenic Evaluation of Bean Milks Using the Pre-Feeding Study
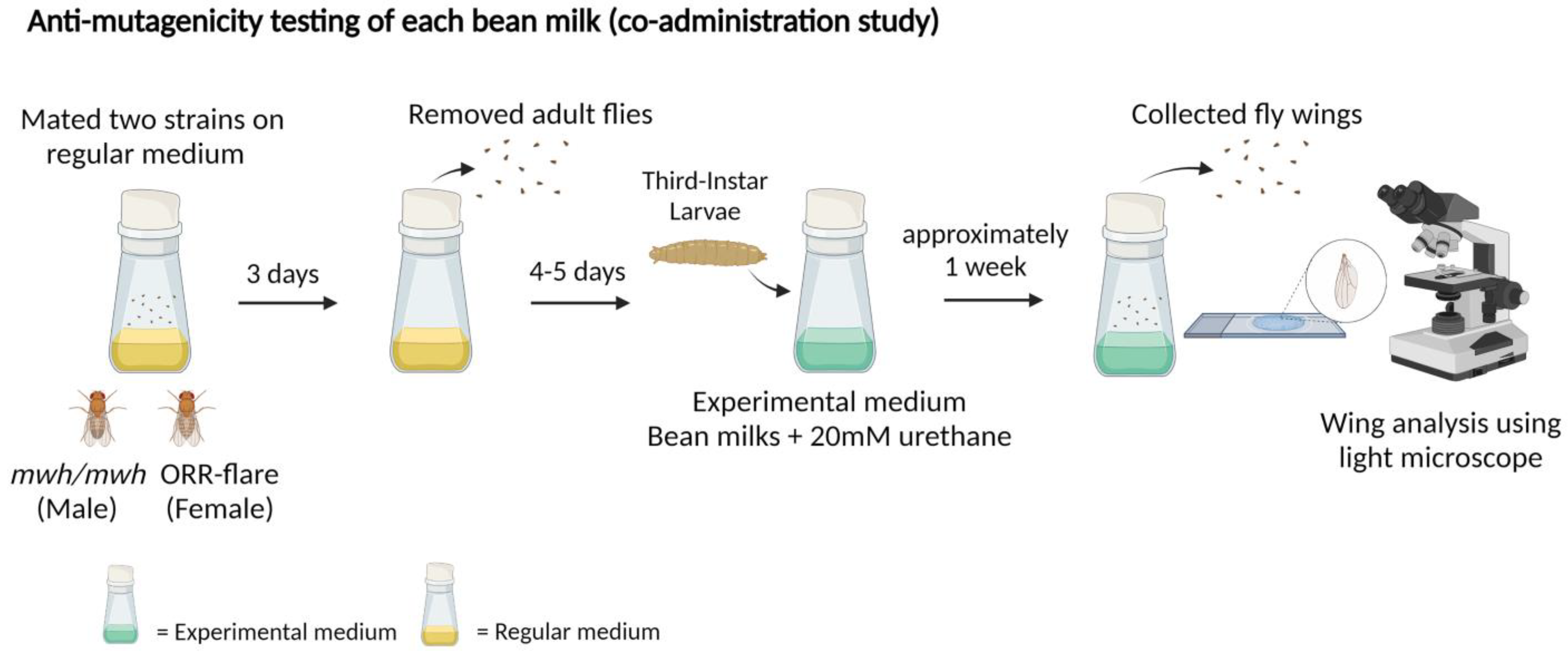
2.3.3. Anti-Mutagenic Evaluation of Bean Extracts Using the Co-Administration Study
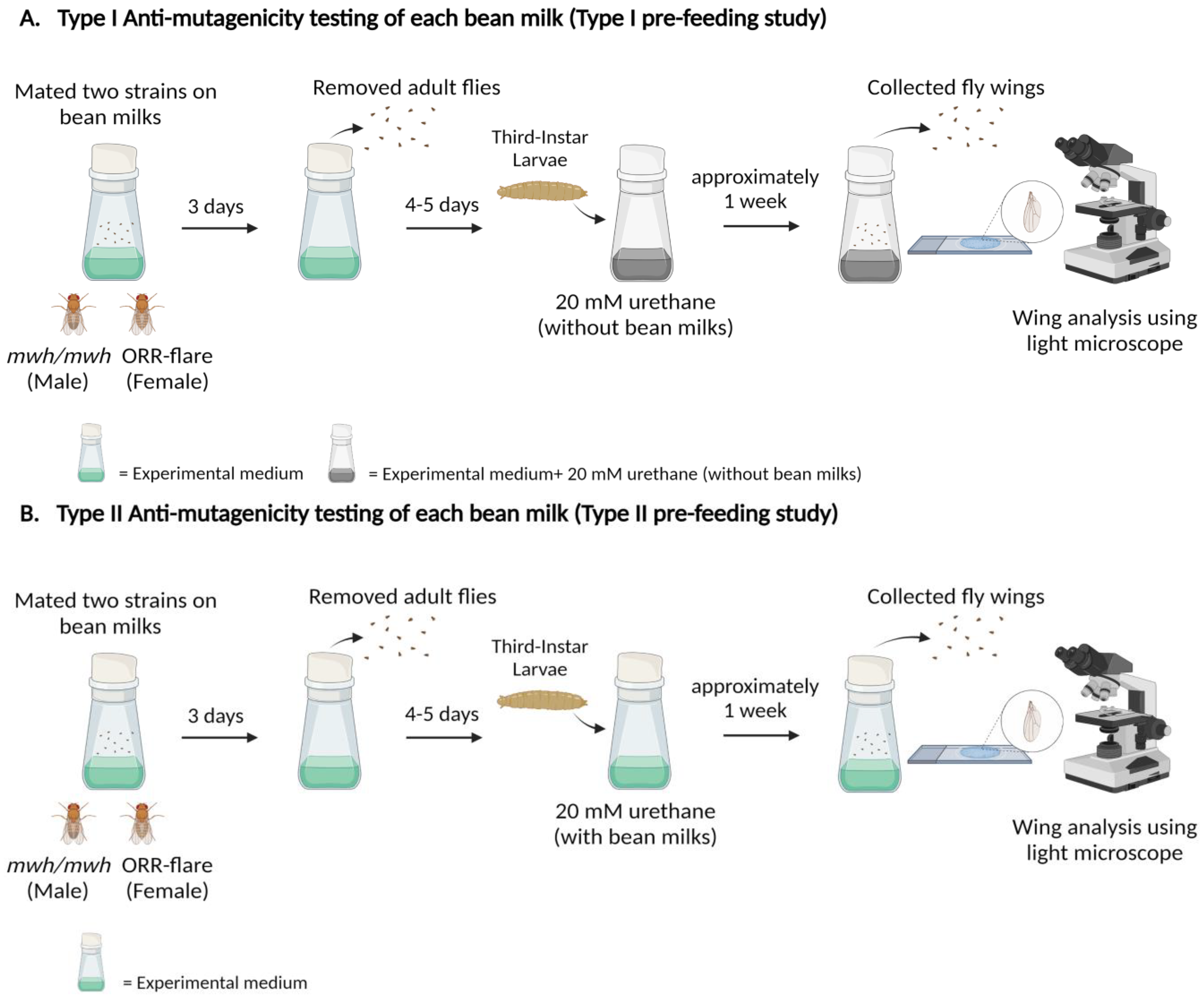
2.3.4. Anti-Mutagenic Evaluation of Bean Milks Using the Type I Pre-Feeding Study
2.3.5. Anti-Mutagenic Evaluation of Bean Milks Using the Type II Pre-Feeding Study
2.4. Ferric Reducing Antioxidant Power (FRAP) Assay
2.5. Total Phenolic Content Assay
2.6. Statistical Analysis
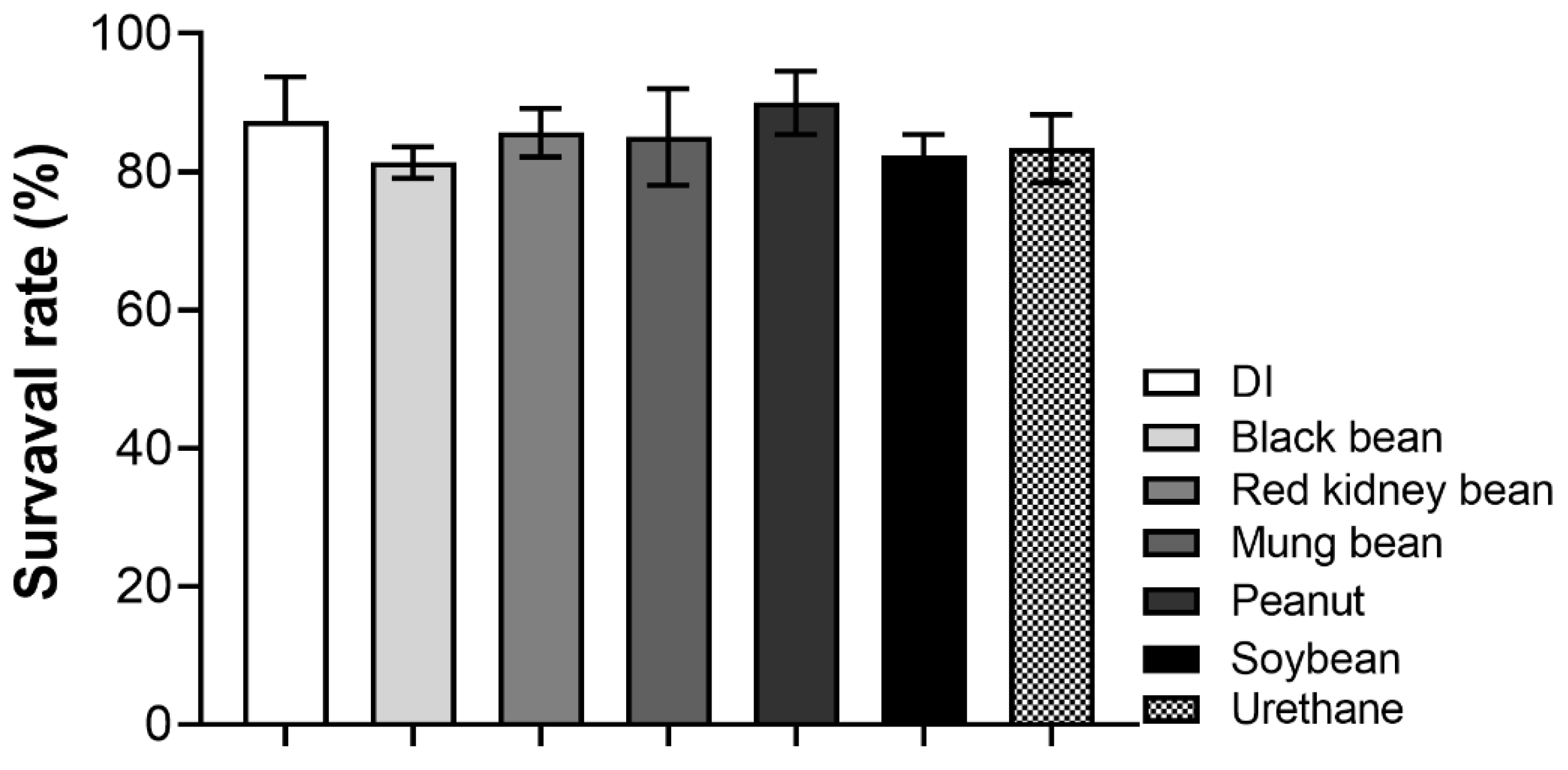
3. Results
3.1. Mutagenic Evaluation of Bean Milks Using the Co-Administration Study
3.2. Mutagenic Evaluation of Bean Milks Using the Pre-Feeding Study
3.3. Anti-Mutagenic Evaluation of Bean Extracts Using the Co-Administration Study
3.4. Anti-Mutagenic Evaluation of Bean Extracts Using the Pre-Feeding Study
3.5. Antioxidant Activities and Total Phenolic Contents of Bean Milks
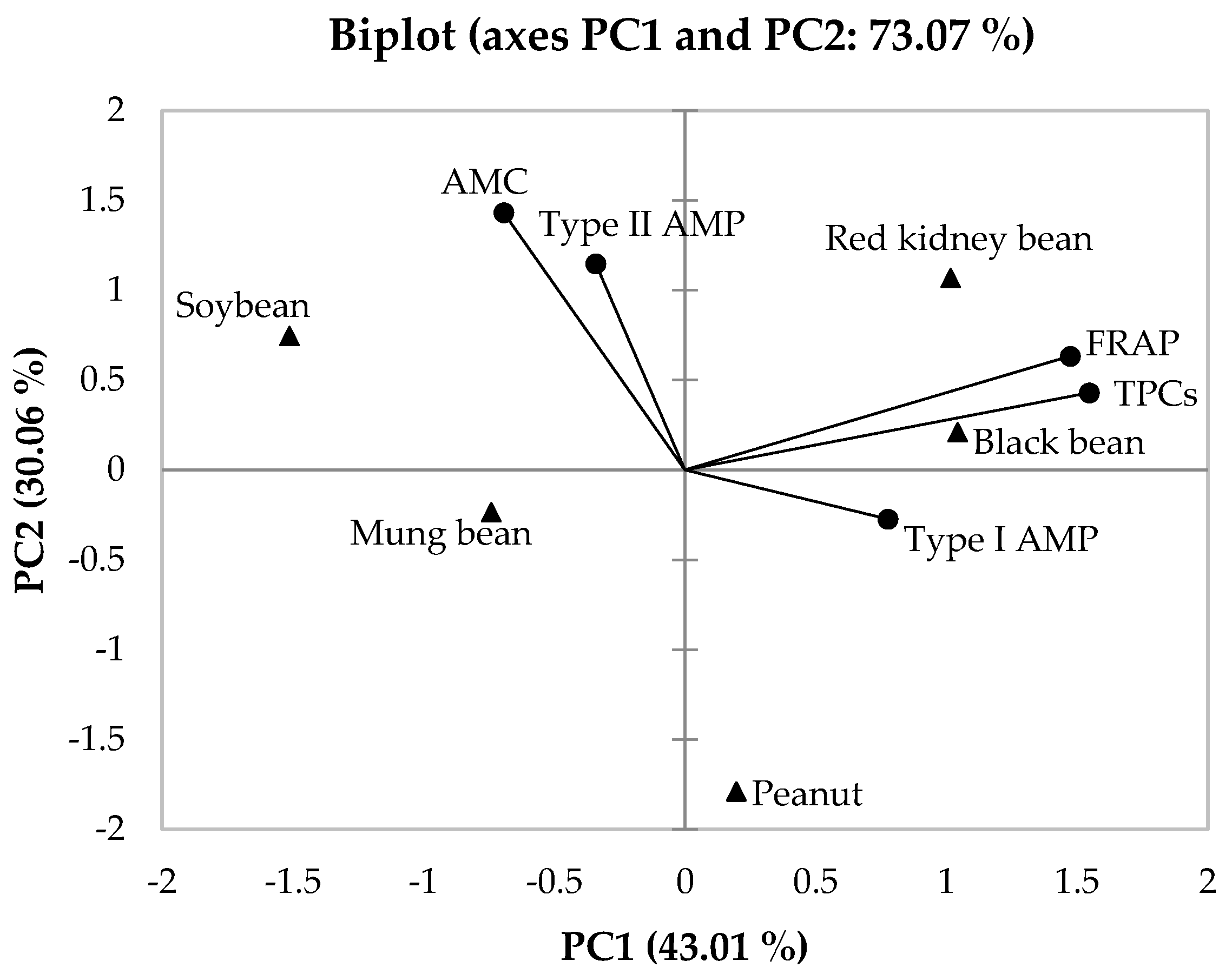
3.6. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pich, O.; Muiños, F.; Sabarinathan, R.; Reyes-Salazar, I.; Gonzalez-Perez, A.; Lopez-Bigas, N. Somatic and Germline Mutation Periodicity Follow the Orientation of the DNA Minor Groove around Nucleosomes. Cell 2018, 175, 1074–1087.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muir, B.; Nunney, L. The expression of tumour suppressors and proto-oncogenes in tissues susceptible to their hereditary cancers. Br. J. Cancer 2015, 113, 345–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, F.O.; Aigner, K.R.; Allen-Mersh, T.G. Basics of Oncology; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer Res. 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Henson, K.E.; Elliss-Brookes, L.; Coupland, V.H.; Payne, E.; Vernon, S.; Rous, B.; Rashbass, J. Data Resource Profile: National Cancer Registration Dataset in England. Int. J. Epidemiol. 2020, 49, 16. [Google Scholar] [CrossRef] [Green Version]
- Kerschbaum, E.; Nüssler, V. Cancer prevention with nutrition and lifestyle. Visc. Med. 2019, 35, 204–209. [Google Scholar] [CrossRef]
- Donaldson, M.S. Nutrition and cancer: A review of the evidence for an anti-cancer diet. Nutr. J. 2004, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Turati, F.; Rossi, M.; Pelucchi, C.; Levi, F.; La Vecchia, C. Fruit and vegetables and cancer risk: A review of southern European studies. Br. J. Nutr. 2015, 113 (Suppl. S2), S102–S110. [Google Scholar] [CrossRef]
- Aune, D.; De Stefani, E.; Ronco, A.; Boffetta, P.; Deneo-Pellegrini, H.; Acosta, G.; Mendilaharsu, M. Legume intake and the risk of cancer: A multisite case–control study in Uruguay. Cancer Causes Control 2009, 20, 1605–1615. [Google Scholar] [CrossRef]
- Murphy, N.; Norat, T.; Ferrari, P.; Jenab, M.; Bueno-de-Mesquita, B.; Skeie, G.; Dahm, C.C.; Overvad, K.; Olsen, A.; Tjønneland, A.; et al. Dietary Fibre Intake and Risks of Cancers of the Colon and Rectum in the European Prospective Investigation into Cancer and Nutrition (EPIC). PLoS ONE 2012, 7, e39361. [Google Scholar] [CrossRef]
- Sahasakul, Y.; Aursalung, A.; Thangsiri, S.; Wongchang, P.; Sangkasa-Ad, P.; Wongpia, A.; Polpanit, A.; Inthachat, W.; Temviriyanukul, P.; Suttisansanee, U. Nutritional Compositions, Phenolic Contents, and Antioxidant Potentials of Ten Original Lineage Beans in Thailand. Foods 2022, 11, 2062. [Google Scholar] [CrossRef] [PubMed]
- Messina, V. Nutritional and health benefits of dried beans. Am. J. Clin. Nutr. 2014, 100 (Suppl. S1), 437s–442s. [Google Scholar] [CrossRef] [Green Version]
- Chávez-Mendoza, C.; Sánchez, E. Bioactive compounds from Mexican varieties of the common bean (Phaseolus vulgaris): Implications for health. Molecules 2017, 22, 1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guajardo-Flores, D.; García-Patiño, M.; Serna-Guerrero, D.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Characterization and quantification of saponins and flavonoids in sprouts, seed coats and cotyledons of germinated black beans. Food Chem. 2012, 134, 1312–1319. [Google Scholar] [CrossRef] [PubMed]
- Takeoka, G.R.; Dao, L.T.; Full, G.H.; Wong, R.Y.; Harden, L.A.; Edwards, R.H.; Berrios, J.D.J. Characterization of black bean (Phaseolus vulgaris L.) anthocyanins. J. Agric. Food Chem. 1997, 45, 3395–3400. [Google Scholar] [CrossRef]
- Hou, D.; Yousaf, L.; Xue, Y.; Hu, J.; Wu, J.; Hu, X.; Feng, N.; Shen, Q. Mung Bean (Vigna radiata L.): Bioactive Polyphenols, Polysaccharides, Peptides, and Health Benefits. Nutrients 2019, 11, 1238. [Google Scholar] [CrossRef] [Green Version]
- Makhafola, T.J.; Elgorashi, E.E.; McGaw, L.J.; Verschaeve, L.; Eloff, J.N. The correlation between antimutagenic activity and total phenolic content of extracts of 31 plant species with high antioxidant activity. BMC Complement. Altern. Med. 2016, 16, 490. [Google Scholar] [CrossRef] [Green Version]
- Pitchakarn, P.; Inthachat, W.; Karinchai, J.; Temviriyanukul, P. Human Hazard Assessment Using Drosophila Wing Spot Test as an Alternative In Vivo Model for Genotoxicity Testing—A Review. Int J. Mol. Sci. 2021, 22, 9932. [Google Scholar] [CrossRef]
- Karinchai, J.; Budluang, P.; Temviriyanukul, P.; Ting, P.; Nuchuchua, O.; Wongnoppavich, A.; Imsumran, A.; Pitchakarn, P. Bioassay-guided study of the anti-inflammatory effect of Anoectochilus burmannicus ethanolic extract in RAW 264.7 cells. J. Ethnopharmacol. 2021, 280, 114452. [Google Scholar] [CrossRef]
- Reyes-Rodríguez, M.; Santos-Cruz, L.F.; García-Castro, C.; Durán-Díaz, Á.; Castañeda-Partida, L.; Dueñas-García, I.E.; Heres-Pulido, M.E.; Rodríguez-Mercado, J.J. Genotoxicity and cytotoxicity evaluation of two thallium compounds using the Drosophila wing somatic mutation and recombination test. Heliyon 2021, 7, e07087. [Google Scholar] [CrossRef]
- Jiang, S.; Cai, W.; Xu, B. Food Quality Improvement of Soy Milk Made from Short-Time Germinated Soybeans. Foods 2013, 2, 198–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graf, U.; van Schaik, N. Improved high bioactivation cross for the wing somatic mutation and recombination test in Drosophila melanogaster. Mutat Res. 1992, 271, 59–67. [Google Scholar] [CrossRef]
- Turna, F.; Aksakal, S.; Demir, E.; Kaya, B. Antigenotoxic effects of Resveratrol in somatic cells of Drosophila melanogaster. Fresenius Environ. Bull. 2014, 23, 2116–2125. [Google Scholar]
- Abraham, S.K. Antigenotoxicity of coffee in the Drosophila assay for somatic mutation and recombination. Mutagenesis 1994, 9, 383–386. [Google Scholar] [CrossRef]
- Griffin, S.P.; Bhagooli, R. Measuring antioxidant potential in corals using the FRAP assay. J. Exp. Mar. Biol. Ecol. 2004, 302, 201–211. [Google Scholar] [CrossRef]
- Amarowicz, R.; Pegg, R.; Rahimi-Moghaddam, P.; Barl, B.; Weil, J. Free-radical scavenging capacity and antioxidant activity of selected plant species from the Canadian prairies. Food Chem. 2004, 84, 551–562. [Google Scholar] [CrossRef]
- Frei, H.; Würgler, F.E. Statistical methods to decide whether mutagenicity test data from Drosophila assays indicate a positive, negative, or inconclusive result. Mutat Res. 1988, 203, 297–308. [Google Scholar] [CrossRef]
- Schlatter, J.; Lutz, W.K. The carcinogenic potential of ethyl carbamate (urethane): Risk assessment at human dietary exposure levels. Food Chem. Toxicol. 1990, 28, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Payne, A.C.; Mazzer, A.; Clarkson, G.J.; Taylor, G. Antioxidant assays—Consistent findings from FRAP and ORAC reveal a negative impact of organic cultivation on antioxidant potential in spinach but not watercress or rocket leaves. Food Sci. Nutr. 2013, 1, 439–444. [Google Scholar] [CrossRef]
- Maisuthisakul, P.; Suttajit, M.; Pongsawatmanit, R. Assessment of phenolic content and free radical-scavenging capacity of some Thai indigenous plants. Food Chem. 2007, 100, 1409–1418. [Google Scholar] [CrossRef]
- Osaba, L.; Aguirre, A.; Alonso, A.; Graf, U. Genotoxicity testing of six insecticides in two crosses of the Drosophila wing spot test. Mutat Res. 1999, 439, 49–61. [Google Scholar] [CrossRef]
- Kalyarat, K.; Linna, T.; Kaew, K. Antimutagenic and co-mutagenic activities of some legume seeds and their seed coats. J. Med. Plants Res. 2012, 6, 3845–3851. [Google Scholar] [CrossRef]
- Azevedo, L.; Gomes, J.C.; Stringheta, P.C.; Gontijo, A.M.; Padovani, C.R.; Ribeiro, L.R.; Salvadori, D.M. Black bean (Phaseolus vulgaris L.) as a protective agent against DNA damage in mice. Food Chem. Toxicol. 2003, 41, 1671–1676. [Google Scholar] [CrossRef]
- Menis Candela, F.; Giordano, W.F.; Quiroga, P.L.; Escobar, F.M.; Mañas, F.; Roma, D.A.; Larrauri, M.; Comini, L.R.; Soria, E.A.; Sabini, M.C. Evaluation of cellular safety and the chemical composition of the peanut (Arachis hypogaea L.) ethanolic extracts. Heliyon 2020, 6, e05119. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.H.; Collins, A.R. Effects of a soy milk supplement on plasma cholesterol levels and oxidative DNA damage in men—A pilot study. Eur. J. Nutr. 1999, 38, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Berhow, M.A.; Wagner, E.D.; Vaughn, S.F.; Plewa, M.J. Characterization and antimutagenic activity of soybean saponins. Mutat Res. 2000, 448, 11–22. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Sheen, L.-Y.; Chou, C.-C. Storage effects on the content of anthocyanin, mutagenicity and antimutagenicity of black soybean koji. LWT-Food Sci. Technol. 2010, 43, 702–707. [Google Scholar] [CrossRef]
- Cardador-MartÍNez, A.; Albores, A.; Bah, M.; CalderÓN-Salinas, V.; CastaÑO-Tostado, E.; Guevara-GonzÁlEz, R.; Shimada-Miyasaka, A.; Loarca-PiÑA, G. Relationship Among Antimutagenic, Antioxidant and Enzymatic Activities of Methanolic Extract from Common Beans (Phaseolus vulgaris L). Plant. Foods Hum. Nutr. 2006, 61, 161–168. [Google Scholar] [CrossRef]
- Cardador-Martínez, A.; Castaño-Tostado, E.; Loarca-Piña, G. Antimutagenic activity of natural phenolic compounds present in the common bean (Phaseolus vulgaris) against aflatoxin B1. Food Addit. Contam. 2002, 19, 62–69. [Google Scholar] [CrossRef]
- Słoczyńska, K.; Powroźnik, B.; Pękala, E.; Waszkielewicz, A.M. Antimutagenic compounds and their possible mechanisms of action. J. Appl. Genet. 2014, 55, 273–285. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez Madrera, R.; Campa Negrillo, A.; Suárez Valles, B.; Ferreira Fernández, J.J. Phenolic Content and Antioxidant Activity in Seeds of Common Bean (Phaseolus vulgaris L.). Foods 2021, 10, 864. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-Z.; Harnly, J.M.; Pastor-Corrales, M.S.; Luthria, D.L. The polyphenolic profiles of common bean (Phaseolus vulgaris L.). Food Chem. 2008, 107, 399–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymczak, G.; Wójciak-Kosior, M.; Sowa, I.; Zapała, K.; Strzemski, M.; Kocjan, R. Evaluation of isoflavone content and antioxidant activity of selected soy taxa. J. Food Compos. Anal. 2017, 57, 40–48. [Google Scholar] [CrossRef]
- Rodríguez-Roque, M.J.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. Soymilk phenolic compounds, isoflavones and antioxidant activity as affected by in vitro gastrointestinal digestion. Food Chem. 2013, 136, 206–212. [Google Scholar] [CrossRef]
- Zimmerli, B.; Schlatter, J. Ethyl carbamate: Analytical methodology, occurrence, formation, biological activity and risk assessment. Mutat Res. 1991, 259, 325–350. [Google Scholar] [CrossRef]
- Ribovich, M.L.; Miller, J.A.; Miller, E.C.; Timmins, L.G. Labeled 1, N 6-ethenoadenosine and 3, N 4-ethenocytidine in hepatic RNA of mice given [ethyl-1, 2-3 H or ethyl-1-14 C] ethyl carbamate (urethan). Carcinogenesis 1982, 3, 539–546. [Google Scholar] [CrossRef]
- Idaomar, M.; El Hamss, R.; Bakkali, F.; Mezzoug, N.; Zhiri, A.; Baudoux, D.; Munoz-Serrano, A.; Liemans, V.; Alonso-Moraga, A. Genotoxicity and antigenotoxicity of some essential oils evaluated by wing spot test of Drosophila melanogaster. Mutat Res. 2002, 513, 61–68. [Google Scholar] [CrossRef]
- Kopečná-Zapletalová, M.; Krasulová, K.; Anzenbacher, P.; Hodek, P.; Anzenbacherová, E. Interaction of isoflavonoids with human liver microsomal cytochromes P450: Inhibition of CYP enzyme activities. Xenobiotica 2017, 47, 324–331. [Google Scholar] [CrossRef]
- Lepri, S.R.; Sartori, D.; Semprebon, S.C.; Baranoski, A.; Coatti, G.C.; Mantovani, M.S. Genistein affects expression of cytochrome P450 (CYP450) genes in hepatocellular carcinoma (HEPG2/C3A) cell line. Drug Metab. Lett. 2018, 12, 138–144. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Cho, H.J.; Moon, S.; Choi, J.; Lee, S.; Ahn, C.; Yoo, K.Y.; Kim, I.; Ko, K.P.; Lee, J.E.; et al. Pickled Vegetable and Salted Fish Intake and the Risk of Gastric Cancer: Two Prospective Cohort Studies and a Meta-Analysis. Cancers 2020, 12, 996. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Number of Wings | Frequency of Mutant Spots per Individual (Number of Spots) # | |||
|---|---|---|---|---|---|
| Small Single (1–2 Cells) | Large Single (>2 Cells) | Twin | Total Spots | ||
| DI (negative control) | 40 | 0.13 (5) | 0.10 (4) | 0.00 (0) | 0.23 (9) |
| Urethane | 40 | 9.03 (361)+ | 1.30 (52)+ | 1.03 (41)+ | 11.35 (454)+ |
| Black bean | 40 | 0.15 (6)i | 0.06 (2)i | 0.00 (0)− | 0.20 (8)i |
| Red kidney bean | 40 | 0.03 (1)− | 0.00 (0)− | 0.00 (0)− | 0.03 (1)− |
| Mung bean | 40 | 0.13 (5)− | 0.00 (0)− | 0.03 (1)− | 0.15 (6)i |
| Peanut | 40 | 0.08 (3)− | 0.03 (1)− | 0.00 (0)− | 0.10 (4)− |
| Soybean | 40 | 0.10 (4)− | 0.00 (0)− | 0.00 (0)− | 0.10 (4)− |
| Samples | Number of Wings | Frequency of Mutant Spots per Individual (Number of Spots/Wing) # | |||
|---|---|---|---|---|---|
| Small Single (1–2 Cells) | Large Single (>2 Cells) | Twin | Total Spots | ||
| DI (negative control) | 40 | 0.08 (3) | 0.08 (3) | 0.00 (0) | 0.15 (6) |
| Urethane | 40 | 11.05 (442)+ | 2.13 (85)+ | 1.58 (63)+ | 14.75 (590)+ |
| Black bean | 40 | 0.18 (7)i | 0.08 (3)− | 0.00 (0)− | 0.25 (10)i |
| Red kidney bean | 40 | 0.08 (3)− | 0.00 (0)− | 0.03 (1)− | 0.08 (3)− |
| Mung bean | 40 | 0.30 (10)+ | 0.05 (2)− | 0.00 (0)− | 0.30 (12)i |
| Peanut | 40 | 0.13 (5)i | 0.03 (1)− | 0.00 (0)− | 0.15 (6)− |
| Soybean | 40 | 0.10 (4)− | 0.00 (0)− | 0.03 (1)− | 0.10 (4)− |
| Samples | Number of Wings | Frequency of Mutant Spots per Individual (Number of Spots/Wing) | Anti-Mutagenicity (%) | |||
|---|---|---|---|---|---|---|
| Small Single (1–2 Cells) | Large Single (>2 Cells) | Twin | Total Spots | |||
| Urethane | 38 | 5.421 (206) | 2.789 (106) | 0.894 (34) | 9.110 (346) | |
| Black bean | 39 | 4.589 (179) | 1.897 (74) | 0.846 (33) | 7.333 (286) | 17.34 |
| Red kidney bean | 40 | 3.325 (133) | 2.425 (97) | 0.775 (31) | 6.525 (261) | 24.57 |
| Mung bean | 40 | 2.500 (100) | 2.825 (113) | 1.250 (50) | 6.575 (263) | 23.99 |
| Peanut | 40 | 4.275 (171) | 2.425 (97) | 1.300 (52) | 8.000 (320) | 7.51 |
| Soybean | 40 | 3.500 (140) | 1.850 (74) | 0.900 (36) | 6.250 (250) | 27.75 |
| Samples | Number of Wings | Frequency of Mutant Spots per Individual (Number of Spots/Wing) | Anti-Mutagenicity (%) | |||
|---|---|---|---|---|---|---|
| Small Single (1–2 Cells) | Large Single (>2 Cells) | Twin | Total Spots | |||
| Type I | ||||||
| Urethane (positive control) | 37 | 6.189 (229) | 2.108 (78) | 1.054 (39) | 9.351 (346) | |
| Black bean | 37 | 5.756 (213) | 1.324 (49) | 0.648 (24) | 7.730 (286) | 17.34 |
| Red kidney bean | 40 | 5.080 (203) | 1.430 (57) | 1.100 (44) | 7.600 (304) | 12.14 |
| Mung bean | 40 | 6.450 (258) | 1.330 (53) | 1.400 (56) | 9.180 (367) | −6.07 |
| Peanut | 40 | 5.375 (215) | 1.100 (44) | 0.525 (21) | 7.000 (280) | 19.08 |
| Soybean | 40 | 6.250 (250) | 1.850 (74) | 0.675 (27) | 7.650 (306) | 11.56 |
| Type II | ||||||
| Urethane (positive control) | 40 | 6.150 (246) | 2.200 (88) | 1.480 (59) | 9.830 (393) | |
| Black bean | 40 | 3.730 (149) | 1.280 (51) | 0.550 (22) | 6.050 (222) | 43.51 |
| Red kidney bean | 40 | 4.130 (165) | 0.800 (32) | 0.380 (15) | 5.300 (212) | 46.06 |
| Mung bean | 40 | 5.975 (239) | 2.300 (92) | 0.875 (35) | 9.150 (366) | 6.87 |
| Peanut | 40 | 5.450 (218) | 1.925 (77) | 0.975 (39) | 8.350 (334) | 15.01 |
| Soybean | 40 | 1.800 (72) | 0.350 (14) | 0.150 (6) | 2.300 (92) | 76.59 |
| Samples | FRAP Values (mM FeSO4 Equivalent/g DW) | Total Phenolic Contents (mg GAE/g DW) |
|---|---|---|
| Black bean | 0.207 ± 0.006 B | 93.12 ± 0.25 B |
| Red kidney bean | 0.241 ± 0.015 A | 103.73 ± 2.27 A |
| Mung bean | 0.100 ± 0.004 C | 43.69 ± 0.72 C |
| Peanut | 0.081 ± 0.007 D | 43.45 ± 3.56 C |
| Soybean | 0.029 ± 0.001 E | 6.20 ± 1.43 D |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inthachat, W.; Suttisansanee, U.; Kruawan, K.; On-Nom, N.; Chupeerach, C.; Temviriyanukul, P. Evaluation of Mutagenicity and Anti-Mutagenicity of Various Bean Milks Using Drosophila with High Bioactivation. Foods 2022, 11, 3090. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11193090
Inthachat W, Suttisansanee U, Kruawan K, On-Nom N, Chupeerach C, Temviriyanukul P. Evaluation of Mutagenicity and Anti-Mutagenicity of Various Bean Milks Using Drosophila with High Bioactivation. Foods. 2022; 11(19):3090. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11193090
Chicago/Turabian StyleInthachat, Woorawee, Uthaiwan Suttisansanee, Kalyarat Kruawan, Nattira On-Nom, Chaowanee Chupeerach, and Piya Temviriyanukul. 2022. "Evaluation of Mutagenicity and Anti-Mutagenicity of Various Bean Milks Using Drosophila with High Bioactivation" Foods 11, no. 19: 3090. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11193090