
RNA Epigenetics: Fine-Tuning Chromatin Plasticity and Transcriptional Regulation, and the Implications in Human Diseases


Department of Pathology, Hematopathology Section, University of Chicago, Chicago, IL 60637, USA
*
Author to whom correspondence should be addressed.
Genes 2021, 12(5), 627; https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050627
Submission received: 1 March 2021
/
Revised: 13 April 2021
/
Accepted: 14 April 2021
/
Published: 22 April 2021
(This article belongs to the Special Issue Epigenetic Control of Chromatin Organization and Plasticity)
Abstract
:Chromatin structure plays an essential role in eukaryotic gene expression and cell identity. Traditionally, DNA and histone modifications have been the focus of chromatin regulation; however, recent molecular and imaging studies have revealed an intimate connection between RNA epigenetics and chromatin structure. Accumulating evidence suggests that RNA serves as the interplay between chromatin and the transcription and splicing machineries within the cell. Additionally, epigenetic modifications of nascent RNAs fine-tune these interactions to regulate gene expression at the co- and post-transcriptional levels in normal cell development and human diseases. This review will provide an overview of recent advances in the emerging field of RNA epigenetics, specifically the role of RNA modifications and RNA modifying proteins in chromatin remodeling, transcription activation and RNA processing, as well as translational implications in human diseases.
1. Introduction
In eukaryotes, genomic DNA and its associated proteins, primarily histones, are organized into chromatin structural domains that affect the binding of transcription factors (TFs), recruitment of RNA polymerases (RNAPs) and the initiation and elongation of transcription [1]. The advent of new genome-wide sequencing technologies has identified chromatin-associated RNAs (ChrRNAs) [2], including nascent RNAs, coding RNAs and various regulatory non-coding RNAs (ncRNAs). RNAs can act either in cis by targeting genomic DNA or in trans through RNAP complexes and interactions with DNA-binding proteins, such as TFs and DNA/histone modifiers, to regulate chromatin structure and gene expression. There are extensive studies and reviews of the roles of ncRNAs in the regulation of DNA/histone modifications, chromatin structure, and gene expression [3,4,5,6,7,8,9,10]. In contrast, the role of RNA modifications and RNA modifying proteins (RMPs) in chromatin plasticity and transcription regulation remains largely unknown. This review will discuss recent studies, which suggest RNA modifications and RMPs function to fine-tune chromatin structure, in turn facilitating transcription activation or repression.
2. Co- and Post-Transcriptional RNA Modifications and RNA Modifying Proteins
2.1. Overview of RNA Modifications and RMPs
Over 170 chemical modifications have been identified in RNA [11]. The associated RNA modifying proteins (RMPs) are classified into three groups based on their roles in RNA modification: (1) writers: enzymes that catalyze specific RNA modifications, (2) readers: enzymes that recognize and selectively bind to RNA modifications and (3) erasers: enzymes that remove specific RNA modifications. These RMPs are distributed throughout the nuclei, cytoplasm and mitochondria and are involved in nearly all essential bioprocesses (Figure 1) [11,12,13]. In this section, we will provide a brief update on the common RNA modifications and their associated RMPs as well as their impact on chromatin structure and transcription regulation. The common RNA modifications and their writers, readers and erasers are shown in Figure 2 and Figure 3; N6-methyladenosine (m6A) and its RMPs are summarized in Figure 2A; 5-methylcytosine (m5C) and its RMPs in Figure 2B; Adenosine to inosine (A-to-I) RNA editing and its RMPs in Figure 3A; Cytosine to uracil RNA editing and its RMPs in Figure 3B.
2.2. 5′Cap RNA Modifications and RMPs
In eukaryotes, a newly synthesized mRNA (pre-mRNA) must undergo three steps to become a mature mRNA, which include (1) capping at the 5′ end, (2) adding a poly(A) tail to the 3′ end and (3) splicing to remove introns [14]. The 5′ cap addition is a complex process, illustrated in Figure 4, concluding with a final m7G0pppN1 structure [15]. Loaded by RNA guanine-N7 methyltransferase (RGMT), 7-methylguanosine (m7G) protects the pre-mRNA from degradation and is recognized by 5′ cap-binding complex (CBC). CBC binds the 5′ cap and interacts with transcription machinery and the RNA splicing complex as well as translation initiation factors and the 40S ribosomal subunit to co-ordinate the functions of transcription, splicing, RNA export and translation [15,16].
2′-O-methylation (Nm) is one of the most abundant RNA modifications and can occur on any base in all RNA species [17]. In about half of all capped and polyadenylated RNA molecules, mRNA and small nuclear RNAs (snRNAs) with a 5′ cap structure are further modified by cap methyltransferases 1 and 2 (CMTR1 and CMTR2) to add Nm on the first and second transcribed nucleosides, respectively [18,19] (Figure 4). Cytoplasmic sensors MDA5 and RIG-I, which recognize non-2′-O-methylated viral RNAs, highlight an important function of Nm in the mRNA 5′ cap as a distinction between self and non-self [20,21,22,23].
N6-methyladenosine (m6A) is one of the most prevalent modifications in the 5′ cap of mRNAs [24], and the 5′ cap m6A stabilizes mRNA transcripts through resistance to the mRNA-decapping enzyme DCP2 [25]. The previously-known phosphorylated carboxyl-terminal domain (CTD) of RNA polymerase II (RNAP II) interacting factor (PCIF1) was recently found to be a 5′ cap-specific adenosine N6-methyltransferase (CAPAM), which catalyzes m6A of the first transcribed nucleoside adenosine to form the cap structure m7G(5′)ppp(5′)m6Am [26] (Figure 4). CAPAM/PCIF1 negatively regulates RNAPII-dependent transcription [27]. CAPAM/PCIF1 is the only known cap-specific adenosine N6-methyltransferase that does not methylate adenosine residues in the RNA body and its function is the subject of ongoing research [28].
2.3. Internal m6A and Associated RMPs
m6A is one of the most abundant modifications on mRNA and was first discovered in 1965 [29]. m6A exhibits tissue-specific distribution and is enriched near stop codons and in 3′ untranslated region (3’-UTRs) [30]. Found in all RNA types, including mRNA, tRNA, rRNA and non-coding RNA (ncRNA), m6A is involved in nearly every aspect of the mRNA life cycle and cellular processes, as well as cell/organ development and disease pathogenesis [11,13]. The m6A associated RMPs, i.e., writers, readers and erasers, are listed in Figure 2A. Methyltransferase 3 (METTL3) and methyltransferase 14 (METTL14) form a stable heterodimer that can efficiently catalyze methyl group transferring from S-adenosylmethionine (SAM) to the N6-amine of adenosine in the sequence motif RRACH (where R = G/A, H = A/C/U) of RNA [31,32,33]. The Wilms tumor-associated protein (WTAP) complex can interact with the METTL3/14 complex to promote mRNA methylation [31,34] (Figure 2A). Of note, recent studies showed that METTL16 binds to U6 snRNA and other ncRNAs as well as numerous lncRNAs and pre-mRNAs at a sequence motif, which is different from the METTL3/METTL14 binding motif [35,36], to regulate splicing and mouse embryonic development [35,37].
Readers of RNA m6A modifications primarily include members of the YTH family proteins and heterogeneous nuclear ribonucleoproteins (hnRNPs) (Figure 2A) [38,39,40,41]. YTH family proteins are mainly involved in translation regulation and degradation, either promoting m6A-modified RNA translation (YTHDF1 and 3, YTHDC2) or regulating m6A-modified RNA splicing and degradation (YTHDF2 and 3, YTHDC1) [38,42,43,44]. Similarly, hnRNPs are involved in regulating RNA processing/alternative splicing, and HNRNPC and HNRNPG facilitate m6A binding by targeting m6A-induced structural changes in RNA structure [40]. Additional m6A readers have been identified, such as IGF2BP1-2 and Prrc2a, which stabilize and promote storage of m6A-modified mRNA, further affecting gene expression [45]. Finally, fat mass and obesity associated (FTO) and ALKBH5 have been identified as m6A erasers [46,47,48]. FTO and ALKBH5 are known to regulate pre-mRNA alternative splicing and subsequent mRNA processing, metabolism, and export, respectively [48,49].
2.4. m5C and Associated RMPs
5-methylcytosine (m5C) has been well-characterized in tRNA and rRNA [50,51] and is now increasingly recognized as a widespread RNA modification in coding and non-coding eukaryotic mRNA as well [52,53,54]. However, m5C levels in mRNA remain low, reportedly ~0.43% by one early study [52]. The low abundance of m5C in mRNA is an obstacle in determining m5C function in transcription regulation [50], but the abundance of m5C methyltransferases and readers has revealed some insight into this concept. RNA m5C is catalyzed by RNA cytosine methyltransferases (RCMTs) (Figure 2B). This group is comprised of NOL1/NOP2/sun (NSUN) family members 1–7 and DNA methyltransferase 2 (DNMT2) [55]. RCMTs are known to play a wide variety of roles, notably ribosomal assembly, transcription regulation, mRNA nuclear export and tRNA cleavage and charging [55]. NSUN2 is known to be responsible for the majority of m5C in mRNA [56,57]. NSUN6 has been shown to be implicated in the formation of m5C in mRNA [58].
ALYREF has been shown to selectively bind m5C and is critical for exporting mRNAs that are methylated by NSUN2 [59]. Other proteins, which function as m5C readers, include YTHDF2, and Y-box binding protein 1 (YBX1) [60,61]. Together, these m5C readers are known to selectively bind m5C to regulate transportation, ribosomal biogenesis, and mRNA stability (Figure 2B).
Ten–eleven translocation (TET) enzymes are traditionally known to “reverse” DNA cytosine methylation by oxidizing m5Cs to 5-hydroxymethylcytosines (5hmCs). Recently TET1/2 enzymes have been shown to share partial responsibility for RNA m5C demethylation to 5-hydroxymethylcytosine (hm5C) in tRNA to promote translation [62] and to regulate embryonic stem cell (ESC) self-renewal network regulation [63].
2.5. RNA Editing: A-To-I and C-To-U
RNA editing is another common RNA modification. There are two common forms of RNA editing in higher eukaryotes, known as adenosine-to-inosine (A-to-I) and cytosine to uracil (C-to-U) editing [64]. Both A-to-I and C-to-U are important for codon alteration, as well as implicated in RNA splicing and mRNA processing, stability, localization and translation [64].
A-to-I editing is carried out by adenosine deaminase acting on RNA (ADAR) enzymes 1–3 (Figure 3A), and the vast majority of A-to-I sites are in noncoding sequences, 5′ and 3′ UTRs, and intronic retrotransposon elements, such as Alu [64]. Globally, ADAR1 is the primary editor of repetitive sites and ADAR2 is the primary editor of non-repetitive coding sites, whereas ADAR3 predominantly acts as an inhibitor of editing [65].
Occurring within regions of double-stranded RNA (dsRNA), A-to-I editing causes disruption and destabilization of dsRNA base pairing and affects the RNA interference pathway (RNAi), RNA splicing, and RNA secondary structures in a tissue-specific manner [66,67,68].
ADAR1-mediated A-to-I editing of endogenous dsRNA is needed for normal development [66] and essential in the prevention of cytosolic innate immune system activation [69]. ADAR3-mediated editing occurs primarily in the brain and is found to be overexpressed in glioma cells [70], as well as involved in mouse learning and memory [71]. Interestingly, A-to-I editing has been found to occur in a negative correlation with m6A [72].
C-to-U editing involves the hydrolytic deamination of a cytosine to a uracil base by a member of the AID/APOBEC enzyme family (Figure 3B). All members (apart from APOBEC2 and 4) demonstrate C-to-U deaminase activity on single-stranded DNA or RNA [73]. Genome-wide sequencing studies have shown that C-to-U editing is widespread, predominantly in 3′ UTR and coincident with splicing and polyA sites [74]. When in coding regions, such editing can create start or stop codons or alter amino acid codes and splice sites, leading to varied genetic information on the RNA level [75,76]. Finally, APOBEC1 and ACF regulate nonsense-mediated RNA decay [77]. Taken together, these studies suggest C-to-U editing functions in the regulation of mRNA processing, stability, localization and translation.
3. Transcription and Nascent RNA-Associated Chromatin Structure
3.1. Transcription and Nascent RNA Synthesis
Transcription is the first step of an essential multistep bioprocess and consists of three major steps: initiation, elongation and termination [78]. In eukaryotes, the activity between transcription initiation and elongation can be further described in five major steps: (1) chromatin remodeling; (2) formation of the preinitiation complex (PIC); (3) formation of an open complex and abortive synthesis of short (2–8) nt RNAs; (4) promoter pausing and escaping by RNAPII; and (5) pause releasing and productive elongation (Figure 5). In this section, we will describe these steps in detail.
Chromatin remodeling begins with the interaction between sequence-specific transcription factors (TFs) and chromatin modifiers to “open” condensed chromatin (heterochromatin) [79] (Figure 5, step 1), allowing RNAPII/Mediator complex to bind specific DNA sequences and form a PIC at the gene promoters and enhancers [79] (Figure 5, step 2). The mediator of RNAPII transcription (mediator) complex is critical in transcription activation and is assembled at the PIC. Interactions between RNAPII and mediator complex are facilitated by the C-terminal domain (CTD) of the largest unit, RPB1, of RNAPII, which contains a highly conserved region with up to 52 repeats of the heptad of Y1S2P3T4S5P6S7 (YSPTSPS) largely unphosphorylated during initiation [80] (Figure 5, step 2). Open complex formation is initiated by the multiprotein complex TFIIH. TFIIH can not only unwind DNA to form the transcription “bubble” through its associated helicases (Figure 5, step 3), but can also phosphorylate the YSPTSPS CTD serine 5 residue through its associated CDK7 and cyclin H kinases, stimulating mediator complex and PIC general TF dissociation and allowing RNAPII escape from the promoter [81,82] (Figure 5, step 3). Two other factors, DRB sensitivity inducing factor (DSIF) and negative elongation factor (NELF) contribute to the transition from transcriptional initiation to elongation and RNAPII promoter escape [83]. DSIF is a heterodimer composed of Spt4 and Spt5, and NELF consists of four subunits (A, B, C/D, and E) [84,85]. Spt5 interacts with the 5′ cap of nascent RNA (pre-mRNA), and then NELF recognizes the Spt5 interface [86]. TFIIH interacts with DSIF and NELF, which then cooperatively induce RNAPII promoter escape and subsequent pausing (Figure 5, step 4). Finally, transcription elongation is promoted by P-TEFb [87], which consists of a kinase/cyclin pair of CDK9 and CCNT1 (or CCNT2). P-TEFb phosphorylates DSIF and RNAPII CTD serine 2, inducing NELF release from RNAPII [88] and allowing RNAPII interaction with other elongation factors and chromatin modifying complexes to ensure productive transcription elongation, respectively [89,90,91] (Figure 5, step 5).
3.2. R-Loops as the Regulators of Transcription and Chromatin
3.2.1. Mechanisms of R-Loop Formation
R-loops are three-strand nucleic acid structures consisting of an RNA-DNA duplex and an unpaired DNA strand generated during transcription upon nascent RNA “threadback” invasion into the DNA duplex to displace the non-template strand [92] (Figure 5). Different from the RNA: DNA hybrids occurring inside the active sites of RNAP during replication and transcription, R-loops span a much longer range (~100–200 base pairs) and form outside of RNAP during transcription [93]. R-loops are found ubiquitously from bacteria to mammals and frequently at gene promoters and enhancers, as well as terminators. Functionally, R-loops can positively and negatively regulate transcription and affect the differentiation and lineage plasticity of pluripotent progenitors/stem cells [94,95,96].
3.2.2. Distribution and Function of R-Loops
R-loops prevent DNA methylation by DNMT3B1 and are found to be clustered at the CpG islands (CGIs) of gene promoters and enhancers in the human genome [97]. In transcription activation, R-loops are found to promote recruitment of epigenetic modifiers and RNAPII to CGIs [98]. As shown in Figure 5, R-loops at gene promoters and enhancers are associated with active chromatin markers, such as H3 lysine 27 acetylation (H3K27ac) and H3 lysine 4 trimethylation (H3K4me3), which suggests the involvement of R-loops in enhancer–promoter looping and high-order chromatin architecture [95,99].
Antisense RNA transcripts promote the formation of R-loops, creating a local chromatin environment which facilitates the recruitment of transcription factors and chromatin modifiers to promote gene transcription [100,101]. For example, the LncRNA TARID is reportedly involved in TCF21 transcriptional activation by generating an R-loop at the CpG-rich promoter of TCF21, enabling GADD45A binding to the R-loop and recruitment of the DNA demethylase TET1 [102]. On the other hand, it is also well-documented that R-loops can trigger chromatin condensation and heterochromatin formation to repress gene expression [103]. Co-transcriptional R-loops have been shown to impede the movement of RNAP in vitro and exert a negative impact on transcription elongation [104]. R-loops were reported to regulate the chromatin structure of specific genes involved in control of mouse ESC differentiation by differentially interacting with the active Tip60-p400 histone acetyltransferase complex and repressive PRC2 H3K27me3 complex [95]. Mechanistically, higher Tip60-p400 and lower PRC2 levels are associated with promoter-proximal R-loops and active transcription to promote ESC differentiation, while decreased Tip60-p400 and increased PRC2 disrupts R-loop formation and impairs ESC differentiation [95].
In addition to gene promoters and enhancers, R-loops are highly enriched in retrotransposons (i.e., LINE-1), ribosomal RNA (rRNA), tRNA loci and mitochondria, and are crucial for regulating genome stability, stress response and mitochondrial function [105,106,107]. R-loops can cause hypermutation, hyperrecombination, chromosomal rearrangements or chromosome loss, leading to genome instability and chromosome fragility [108]. Genome-wide mapping of R-loops demonstrates an enrichment at the 3′ end of genes with GC skew patterns in mammalian cells, and R-loop accumulation at the 3′ end of genes inhibits transcription [109]. R-loops were found to be associated with a subset of developmental regulator genes, which are targeted by Polycomb complexes PCR1 and PCR2 [110]. Removal of the R-loop causes decreased PRC1 and PRC2 binding and increased RNAPII recruitment, resulting in activation of these PRC1/2 targeting genes [110]. Arginine methyltransferases PRMT1/5 and CARM1 interact with tudor domain-containing protein 3 (TDRD3) to facilitate recruitment of topoisomerase IIIB (TOP3B) to prevent such R-loop accumulation [111], and release RNAPII for successive transcription by methylating the arginine residues in histone tails, as well as the RNAPII CTD [112].
It is increasingly clear that the formation, distribution and removal of R-loops is tightly regulated during DNA transcription and replication.R-loops are involved in regulation of all stages of gene expression, from transcription initiation to termination, and can act as transcriptional activators or repressors at specific gene loci in different cell types and differentiation states. Regulatory R-loops are important for transcription elongation and termination, as well as telomere stability and DNA repair in normal physiological conditions. In contrast, dysregulated R-loops are associated with transcription elongation defects, DNA damage and genome instability in pathophysiological conditions, such as cancer [92,105].
3.3. MYC, 7SK snRNP and BRD4 Transcription and Chromatin Structure Regulation
3.3.1. MYC-Mediated RNA 5’ Capping, Transcription Elongation and Active Chromatin
MYC is well-known to activate Myc-dependent transcription, but its underlying mechanisms remain incompletely understood [113,114]. As shown in Figure 6, MYC is involved in m7G 5′ capping of pre-mRNAs by binding to the E-box in the promoter of active genes, which recruits the capping enzyme complex composed of RNA guanylyltransferase and 5’-phosphatase (RNGTT) and CMTR1 [114]. Upon binding to the E-box, MYC also recruits CDK7/TFIIH and CDK9/P-TEFb to gene promoters, which phosphorylate RNAPII CTD serine 5 and 2 residues, respectively [115].
Finally, MYC appears to be implicated in regulation of R-loop formation as well. Recent studies have shown that MYC can suppress R-loop formation in promoter-proximal regions through recruitment of BRCA1, which stabilizes the DCP2 de-capping enzyme complex and enables RNAPII escape from promoter pausing, leading to increased transcription elongation and gene expression [116] (Figure 6).
3.3.2. SK snRNP Complex as a Negative Regulator of Transcription Elongation
The 7SK small nuclear ribonucleoprotein (7SK snRNP) complex is composed of the core 7SK snRNPs and other proteins, including AFF1/4, HEXIM1/HEXIM2, LARP7, MePCE and hnRNPs [117]. This complex functions as a major negative regulator of CDK9/P-TEFb by inhibiting CDK9/P-TEFb-mediated RNAPII escape and transcription elongation [118,119]. 7SK snRNP interacts with HEXIMI that directly binds CDK9/P-TEFb, inactivating its kinase activity and inhibiting transcription elongation [120] (Figure 6).
3.3.3. BRD4-Mediated Transcription Elongation and Active Chromatin
BRD4 is a member of the Bromodomains and Extraterminal (BET) family and contains two bromodomains that recognize acetylated lysine residues of histone tails [121,122]. BRD4 not only promotes the establishment of chromatin, favoring transcriptional activation, but also directly competes with release of CDK9/P-TEFb sequestrated by the 7SK snRNP complex, which promotes CTD-S2 phosphorylation and activates RNAPII-dependent transcription elongation [123] (Figure 6). BRD4 was initially reported to interact with CDK9/P-TEFb through its bromodomain, resulting in CDK9/P-TEFb-dependent phosphorylation of RNAPII CTD serine 2 (CTD-S2) and transcription activation in vivo [124,125]. However, later studies demonstrated that BRD4 functions as an atypical kinase by directly binding and phosphorylating CTD-S2 both in vitro and in vivo under conditions where other CTD kinases are inactive [126]. Our study demonstrated that RNA cytosine methyltransferase NOP2/NSUN1 and NSUN2 interact with BRD4 and the elongating form of RNAP (CTD-S2P) to form a transcriptionally active chromatin structure in leukemia cells [127] (Figure 6). BRD4 was also found to prevent R-loop accumulation and to protect against transcription-replication collision [128]. Loss of BRD4 in cancer cells leads to replication stress, DNA damage and eventually apoptotic cell death [128].
4. Co-Transcriptional RNA Splicing and Its Associated Chromatin Structure
4.1. RNA 5′ Capping Enzyme-Coupled Spliceosome Assembly
Over 90% of human genes undergo alternative splicing, which is carried out by splicing machinery called the spliceosome. The spliceosome contains more than 150 associated proteins and five uridine-rich small nuclear RNAs (snRNAs)—U1, U2, U4, U5 and U6 (RNU1, RNU2, RNU4, RNU5 and RNU6) [129,130]. Nascent RNA splicing is regulated by multiple cis-elements and trans-factors through extensive interactions between splicing factors and the core transcription machinery [131,132,133,134,135,136]. At the earliest stages of spliceosome assembly (Figure 7A), the CTD serine 5 phosphorylation (S5P) of RNAPII facilitates recruitment of the 5′ capping enzyme complex (CE/RGMT). The U1 snRNP binds to the GU sequence at the 5′ splice site (5′SS) of the first intron of mRNA through base pairing, and splicing factors U2AF1 and U2AF2 bind at the 3′SS and the branch point sequence (BPS), respectively [137]. The U2 snRNP complex interacts with the BPS and the pre-mRNA 3′SS site of pre-mRNA, and the SF3A and SF3B proteins in the U2 snRNPP complex stabilize these interactions [138]. The U5/U4/U6 snRNP trimer is recruited with the U5 binding to the 5’SS and the U6 binding to U2 (Figure 7A). Then the U1 snRNP is released, U5 shifts from exon to intron, and the U6 binds at the 5’ splice site (Figure 7b). The splicing factor SF2/ASF binds the S2-phosphorylated CTD of RNAPII to facilitate the interactions between the spliceosomal U1 and U2 snRNPs complexes as well as RNAPII during the splicing process [139,140].
4.2. Alternative Spliceosome Assembly and Determination of Internal Exon/Intron Junctions
Determination of the internal exons and introns of pre-mRNAs, i.e., alternative splicing, is carried out by the spliceosome through extensive interactions between trans-acting factors/complexes and cis-acting spicing elements/sequences, as well as transcription machinery and chromatin modifying complexes [132,135,141,142,143,144,145,146,147]. Serine and arginine rich splicing factors (SRSFs) selectively bind to cis exonic and intronic splicing elements, promoting spliceosome formation [148], while hnRNPs, such as hnRNPA1 (HNRNPA), negatively regulate spliceosome formation by binding to exonic or intronic splicing silencers [149] (Figure 7B).
4.3. Alternative RNA Splicing Regulation by Transcription Elongation Complex
Transcription elongation rate and chromatin structure have a large impact on splice site identification and definition of exons and introns by the spliceosome [143]. Recent studies show depletion of the pre-exon junction complex (EJC), a protein complex formed at the junction of two exons of a pre-messenger RNA during RNA splicing, led to a global decrease in RNAPII pausing and premature entry into elongation by CDK9/P-TEFb, suggesting that the pre-EJC serves as an early transcriptional checkpoint to prevent premature entry into elongation and proper exon definition [150].
Other trans-acting factors, such as MYC, BRD4 and many RNA-binding proteins (RBPs) regulate transcription elongation and alternative splicing in tissue-specific patterns and sizes [151,152]. MYC and BRD4 have been shown to physically interact with RNA splicing factors to contribute to spliceosome formation and alternative splicing regulation at the co- and post-transcriptional levels [153,154,155,156,157], as shown in Figure 7.
4.4. Nascent Cis-Acting Elements and Degradation Machinery Regulate Alternative RNA Splicing
Intronic RNAs constitute most pre-mRNAs in vertebrate cells and were traditionally considered transcriptional “junk” or “waste” due to their complete degradation first by a unique debranching endonuclease, followed by exoribonucleases after splicing [158]. However, recent studies show that many spliced introns can escape complete degradation and are further processed to generate regulatory RNAs that can regulate chromatin structure, gene expression, and genomic stability [159] (Figure 7).
Terminal exons are defined by the cleavage and polyadenylation complex (CPA), which is composed of the cleavage and polyadenylation specificity factor (CPSF) complex, cleavage factors (CFs) and the cleavage stimulation factor (CstF) complex. The CPSF complex recognizes the polyadenylation signal sequence AAUAAA and other nearby sequences [160] (Figure 7B). The poly(A) tail is added to nascent RNA when the nascent RNA is cleaved to produce mature RNA by the polyadenylate polymerase complex, which is also physically linked to the spliceosome [161]. After introns are spliced out from pre-mRNA, exons are ligated together to yield multiple mature mRNAs through alternative splicing, significantly increasing transcriptomic/proteomic complexity [162].
4.5. The Impact of Chromatin Structure and R-Loops on Alternative RNA Splicing
Earlier studies showed that nucleosome positioning contributes to definition of exons and introns, and the ratio between nucleosome occupancy within and upstream from the exons correlates with exon-inclusion levels [163,164,165]. Recent studies have demonstrated that splicing machinery interacts with histone and DNA modifications and their modifiers, such as histone acetyltransferases (HAT), histone deacetylases (HDACs), histone methyltransferases (HMTs), DNA methylation, and DNA methyltransferases (DNMTs) [166,167] (Figure 7A,B). Using HDAC inhibitors (HDACIs), such as trichostatin A (TSA) or knockdown of HDAC1 or HDAC2 increased H4 acetylation, specifically at the exon 2 of MCL1, a gene encoding a longer form of antiapoptotic protein (MCL1L) and a shorter form of proapoptotic protein (Mcl-1S), resulting in exon 2 skipping and production of the short form of proapoptotic MCL1S [168,169]. Altered histone methylations and methyltransferases are associated with differential exon inclusion and intron retention [170,171]. ZMYND11, aka BS69, is a reader for histone H3.3 lysine 36 trimethylation (H3K36me3) and involved in transcription elongation. It is known to interact with U5 snRNP in the spliceosome, and depletion of BS69/ZMYND11 strongly decreases intron retention [172].
Other studies have shown that DNA methylation and DNA modifiers are also involved in RNA splicing [173]. Higher levels of DNA methylation have been identified in exons, and especially splice sites, than those found in flanking introns [174]. The zinc-finger protein CCCTC-binding factor (CTCF) negatively regulates RNAPII elongation rate through transient obstruction of RNAPII and promotes splicing by favoring spliceosome assembly at weak splice sites. DNA 5-methylcytosine (5mC) promotes exon exclusion through 5mC-mediated CTCF eviction; however, TET1 and TET2-mediated 5-hydroxymethylcytosine (5hmC) has an opposite effect on CTCF-dependent splicing [175] (Figure 7). The methyl-CpG binding protein 2 (MeCP2) was shown to interact with the RNA-binding protein Y box-binding protein 1 (YBX1) and to regulate alternative splicing in Rett syndrome (RTT) mouse models [176]. Additionally, a recent study demonstrated that R-loops are able to activate gene promoters by recruiting the DNA methylcytosine dioxygenase TET1 to demethylate CpG islands [102].
5. Chromatin Associated Non-Coding RNAs and Acting Modes in Chromatin Remodeling
5.1. Non-Coding RNAs (ncRNAs) Subtypes
The advent of genome-wide sequencing technologies has revealed that 75% of the human genome is transcribed into RNA, but only a small percentage of transcribed RNA encodes proteins [177,178]. Noncoding RNAs play an important role in regulating gene expression and chromatin structure and can be functionally or structurally divided into several subtypes [179,180,181,182,183,184], as shown in Table 1.
As stated previously, ncRNAs were traditionally considered transcriptional “junk” or “waste”, but are now widely accepted to play an important role in regulation of transcription elongation and chromatin structure [185]. It is well-known that ncRNAs can be generated from gene bodies, extragenic regions (particularly enhancers) [186,187], and promoter upstream sequences to generate a class of short, polyadenylated, and highly unstable RNAs called promoter upstream transcripts (PROMPTs) [188,189]. Expression levels of enhancer RNAs (eRNAs) are correlated with signal-regulated transcriptional activity of gene enhancers. Furthermore, the heterogeneity of eRNAs may also be correlated with their functional diversity in the mammalian genome [187,190].
5.2. Non-Coding RNAs Regulate Chromatin Structure and Transcription in Cis
The cis-acting models of ncRNAs include formation of R-loops and DNA-RNA triple helixes through Hoogsteen base-pairing interactions between the major groove of double-stranded DNA and single stranded RNA [191,192,193]. Particularly, the cis-acting symmetrical triplex-forming motifs in lncRNAs favor the formation of RNA-DNA triplexes, which can affect RNA-RNA, RNA-protein, and RNA-DNA interactions and modulate chromatin structure [191]. By forming RNA: DNA: DNA triplexes/R-loops, lncRNAs can target specific DNA sites, such as enhancers, to contribute to local chromatin organization [194,195].
5.3. Non-Coding RNAs Regulate Chromatin Structure and Transcription in Trans
The most common mode of ncRNA-mediated chromatin organization is through in trans RNA-protein interactions. The role of RBPs in regulating chromatin structure and gene expression has been studied extensively [196,197]. RBPs may have single or multiple previously characterized canonical RNA binding domains (RBDs), including zinc finger domains [198,199], RNA recognition motif (RRM) [200], double-strand RNA binding domain (dsRBD) [201] and hnRNPK homology domain (aka KH) [202,203]. As one of the best-characterized RBPs, hnRNPK is known to play a role in regulating transcription and splicing, as well as chromatin structure and DNA damage and repair [204,205,206,207]. Other RBPs are known to be involved in chromatin remodeling through interaction with DNA methyltransferases (DNMTs) or methyl-DNA-binding domain (MBD)-containing proteins, such as Methyl-CpG binding protein 2 (MeCP2) [208,209]. MeCP2 is a multifunctional epigenetic reader that can bind RNA and methylated CpGs in DNA to orchestrate chromatin compartmentalization and higher order genome architecture [209].
Recent genome-wide proteomic studies have identified many new RBPs [2,210]. Those newly identified RBPs are conserved from yeast to humans. Despite lacking the conventional or canonical RBDs, those new RBPs can modulate chromatin structure and gene expression in response to various environmental and intrinsic stimuli [2,210,211,212,213]. Both canonical and non-canonical RBPs are reported to function together to regulate transcription and chromatin structure [214,215].
6. Fine-Tuning Chromatin and Transcription through RNA Modifications and RNA-Modifying Proteins
6.1. m6A and RMPs Modulate Interactions between Non-Coding RNAs, Transcription Factors and Chromatin Modifiers
The METTL3/14 methyltransferases, YTH-domain-containing proteins (YTHDs)/HNRNPs, and FTO/ALKB family proteins function as m6A writers, readers and erasers, respectively, to regulate the level and distribution of m6A in a tissue-/differentiation-specific manner [31,34,38,46,216] (Figure 2A). Recent studies have shown widespread m6A in human tissues, which is significantly enriched in long intergenic non-coding RNA (lincRNA) and CpG-rich gene promoters, suggesting that m6A is positively correlated with gene expression homeostasis and has broad involvement in human development and disease [217].
Increasing evidence suggests that m6A and its associated RMPs, more specifically m6A writers and readers, modulate chromatin structure and transcriptional activation through interactions with TFs and chromatin modifying proteins/complexes [218,219]. METTL14 was reported to recognize the active histone mark H3K36me3, resulting in METTL3-METTL14-WTAP m6A methyltransferase complex recruitment to the vicinity of H3K36me3 peaks, promoting m6A and positively regulating transcription elongation [220] (Figure 8A).
Zinc finger protein 217 (ZFP217), a chromatin-associated oncogenic TF important for embryonic stem cell (ESC) differentiation [221], is associated with repressive chromatin marked by histone 3 lysine 9 trimethylation (H3K9me3) and histone 3 lysine 27 trimethylation (H3K27me3) through interactions with G9a and EZH2 methyltransferases [8]. ZFP217 also negatively affects m6A deposition at its own target gene mRNAs by sequestering METTL3 in an inactive status, in which METTL3 is dissociated from the METTL14/WTAP co-factor complex [221,222] (Figure 8A).
YTHDC1 can de-repress (activate) gene expression by physically interacting with and recruiting KDM3B demethylase to hyper-m6A in nascent RNAs, leading to demethylation of the repressive transcriptional histone modification H3K9me2 [223]. YTHDC1 can also facilitate the decay of a subset of these m6A-modified RNAs, especially LINE-1, through nuclear exosome targeting-mediated nuclear degradation, while METTL3 deposits m6A modifications on chromosome-associated regulatory RNAs (carRNAs), including promoter-associated RNAs, enhancer RNAs and repeat RNAs [224].
m6A was reported to promote translation of the SETD1A/B components, which increases the transcriptional activation histone mark H3K4me3 and recruitment of the erythroid-specific zinc finger transcription factor KLF1 to its target gene promoters [225]. Inhibition of the METTL3/METTL14/WTAP m6A methyltransferase complex blocks erythropoiesis in human bone marrow hematopoietic stem/progenitor cells (HSPCs). The METTL3-METTL14-WTAP complex also functionally interacts with SMAD2/3 proteins, the key signal transducers and transcriptional modulators of the TGFβ pathway. SMAD2/3 promotes binding of the m6A methyltransferase complex to a subset of transcripts involved in early cell fate decisions [226]. In addition, a recent study showed that RBM15, a subunit of the m6A methyltransferase complex, mediates the degradation of the pre-BAF155 mRNA to affect the chromatin remodeling function of the BAF (SWI/SNF-like) complex [227].
6.2. The Impact of m5C and Its RMPs on Chromatin and Transcription
In eukaryotes, RNA m5C is catalyzed by RCMTs, including NOL1/NOP2/SUN domain (NSUN) family enzymes and DNMT2 (Figure 2B). NSUN2 is responsible for catalyzing the majority of m5C in mRNAs and non-coding RNAs [57,228,229]. Our study demonstrated a marked increase in NOP2/NSUN1 and NSUN2-mediated m5C in drug (5-azacitidine) resistant myeloid leukemia cells [127]. NSUN7 is reported to promote transcription of genes labeled with enriched m5C eRNAs in mouse hepatoma cells and primary hepatocytes for adaptive metabolic responses [226]. Mechanistically, the methyltransferase SET7/9 methylates histone 3 K4 residues [227] and PGC-1α K779, a transcriptional co-activator important for regulating adapted metabolic responses [230]. Methylated PGC-1α interacts with Spt-Ada-Gcn5-acetyltransferase (SAGA) and mediator complexes, as well as NSUN7, at the m5C-marked enhancer RNAs of PGC-1α target genes to promote recruitment of transcription machinery and reinforce transcription of those genes (Figure 8).
Heterozygous mutations in the X-linked gene encoding MeCP2 cause the neurological disorder Rett syndrome. MeCP2 interacts with more than 40 binding partners, including transcriptional regulators, chromatin modifiers and RNA- splicing factors [231]. MeCP2 chromatin binding is controlled by m5C, and recent studies show enrichment of MeCP2 on long non-coding RNAs (lncRNAs), such as retinal noncoding RNA3 (RNCR3) [232], muscle-specific long non-coding RNA (ChRO1) [233], and major Satellite Forward RNAs [234]. This MeCP2 enrichment on lncRNAs promotes deposition of H3K9me3 and H4K20me3 and heterochromatin formation. Additionally, m5C is present in a wide variety of RNA species, including cytoplasmic and mitochondrial ribosomal RNAs (rRNAs) and tRNAs, as well as mRNAs, eRNAs and other types of non-coding RNAs [55]. There are widespread and dynamic changes in RNA m5C in response to stress and stimuli, which suggests an important role of m5C and its RMPs in epigenetic gene regulation [55]. However, the detailed mechanisms underlying m5C and its RMP-mediated chromatin remodeling and gene regulation are yet to be elucidated.
6.3. RNA Modifications and RMPs in R-Loop Formation and Transcription Regulation
R-loops can be formed both in cis by a nascent RNA folding back to its DNA template at the same transcription locus and in trans by ncRNAs at distant enhancers (Figure 5). R-loops are prevalent in highly transcribed genes, and occur most frequently at conserved genetic hotspots, such as unmethylated CpG island promoters, G-rich terminators and other regulatory chromosomal loci [98,104]. R-loops play an important role in organizing chromatin structure and maintaining genome stability [99], and RNA modifications and their associated RMPs can modulate the interactions between chromatin-associated RNAs and chromatin modifiers, as well as transcription machinery, to regulate chromatin structure and transcription in a position and context-dependent manner [235].
6.3.1. m6A and Its RMPs Promote R-Loop Formation at Transcription Termination Sites
A recent study showed that R-loops accumulate in m6A-rich transcription termination sites (TTSs) [236]. Depletion of m6A methyltransferase complex components dramatically reduces R-loop accumulation in m6A marked genes around TTSs, leading to disruption of proper transcription termination [236]. In contrast, depletion of the nuclear m6A reader protein YTHDC1 does not affect R-loop levels [236].
6.3.2. m6A and RMPs Associated R-Loop Formation on DNA Damage-Associated RNAs
ATM serine/threonine kinase (ATM) is recruited to DNA double-strand break (DSB) sites to phosphorylate several key proteins, leading to cell cycle arrest, DNA repair or apoptosis [237]. Specifically, in response to DSBs, ATM phosphorylates METTL3 serine 43, and the phosphorylated METTL3 is then recruited to DNA-damage sites to catalyze m6A in the DNA damage-associated RNAs. This increased m6A deposition leads to YTHDC1 recruitment and R-loop accumulation, promoting homologous recombination-mediated DNA repair [238]. In human pluripotent stem cells, m6A-containing R-loops are reported to accumulate during G2/M and are depleted at G0/G1 phases of the cell cycle [239]. Knockout of the m6A reader YTHDF2 leads to increased R-loop levels, cell growth retardation, and accumulation of the DNA double-strand break marker γH2AX in mammalian cells, which suggests that YTHDF2 negatively regulates m6A and R-loop accumulation [239]. Taken together, these results suggest that m6A and its RMPs regulate accumulation of R-loops and genomic stability in a position and context-dependent manner [96,240,241].
6.3.3. RNA Cytosine Methylations and RMPs in R-Loop Formation
R-loops have been found to be required for transcription-coupled homologous recombination, as they recruit Cockayne Syndrome Protein B (CSB) and RAD52 proteins, but not the canonical HR proteins BRCA1 and BRCA2, to sites of reactive oxygen species (ROS)-induced DNA damage [242]. The formation of R-loops at break sites is dependent upon Drosha, an RNase III enzyme in the microRNA (miRNA) biogenesis apparatus [243]. DNMT2 is predominantly located in the cytoplasm and mitochondria, where it contributes to m5C in tRNAs, consequently affecting protein translation [244,245,246]. Little is known about the role of DNMT2 in regulating chromatin structure and R-loop formation. A recent report showed that DNMT2 (Figure 3B) is recruited to DNA damage sites, where it deposits m5C onto the DNA damage-associated mRNAs and contributes to R-loop formation. The m5C and DNMT2-mediated R-loops facilitate the homologous recombination (HR) and DNA repair of DSBs [247].
Loss of TRDMT1 in cancer cells confers sensitivity to PARP inhibitors in vitro and in vivo, suggesting that transcription-coupled RNA modifications may serve as DNA damage markers to regulate DNA repair [247]. METTL8 was previously reported to contribute m3C in mRNA, while METTL2 and METTL6 contribute m3C in tRNAs [248]. Recently, METTL8 was found to form a large SUMOylated nuclear RNA-binding protein complex that is associated with R-loops, and knockout of METTL8 resulted in reduction of m3C and R-loops in the nucleolus [249]. The nucleolar-enriched SUMOylated METTL8 complex regulates R-loop formation and promotes tumorigenesis [249]. Retention of R-loops at their transcription sites of nascent RNAs is an important and intrinsic part of chromatin structure in human cells, and R-loop dysregulation is associated with DNA damage, transcription elongation defects, hyper-recombination and genome instability [105,250].
6.4. RNA Editing in Chromatin Remodeling
6.4.1. A-To-I Editing and ADAR-Mediated Heterochromatin and Gene Silencing
A-to-I editing is one of the most abundant RNA modifications in mammalian cells and is catalyzed by ADAR family proteins ADAR1 and ADAR2 (Figure 3A). A-to-I editing occurs most frequently in non-coding RNAs within repetitive elements in the genome, mainly Alu repeats [251,252,253], and has a critical role in immune regulation [69,254], and alternative splicing [67]. Vigilin is the largest protein in the KH domain-containing RNA-binding protein family [255]. Vigilin binds to promiscuously A-to-I-edited RNAs, predominantly in inverted Alu repeats, and is involved in the formation of heterochromatin [256], which is marked with the repressive chromatin marker H3K9me3 by histone H3-specific methyltransferase SUV39H1 [257,258]. Mechanistically, vigilin binds to A-to-I-edited RNAs in a complex that contains ADAR1, as well as RNA helicase A and Ku86/70 [259]. The C-terminal domain of human vigilin interacts with SUV39H1 and recruits it to the A-to-I-edited RNAs, leading to formation of heterochromatin and gene silencing [259]. This suggests a mechanistic link between DNA-repair machinery and the A-to-I RNA editing complex. Interestingly, the levels of global A-to-I editing and m6A in mammalian cells were shown to be negatively correlated, and depletion of m6A modification increases the association of m6A-depleted transcripts with ADAR enzymes, resulting in upregulated A-to-I editing on the same m6A-depleted transcripts [72,260].
6.4.2. C-To-U Editing and Associated RMPs in Chromatin Remodeling and Gene Regulation
C-to-U editing is carried out by the APOBECs family proteins (Figure 3B). APOBECs function to restrict retroviruses and retrotransposons [261], and have been identified as a prominent source of mutations in cancers [262]. APOBEC3B (A3B) was shown to regulate expression of the estrogen receptor α (ERα)-target gene by causing C-to-U deamination at ER binding regions [263]. Further, transient cytidine deamination by A3B promotes chromatin modification and remodeling at regulatory regions of ER target genes that increase ER expression levels, and elevated A3B expression is associated with poor patient survival in ER+ breast cancer [263]. However, more studies are needed to understand the role of C-to-U editing and the APOBECs in regulation of chromatin structure and gene expression.
7. Implications of RNA Epigenetics in Human Diseases
The advent of new genome-wide sequencing technologies has uncovered abnormal RNA modifications and their associated RMPs in a variety of human diseases. Here we will briefly summarize the recent progress surrounding the roles of some major RNA modifications and their associated RMPs in four major disease categories: neuronal disorders, metabolic disorders, immune disorders/infections and cancer/leukemia (Table 2).
7.1. RNA Epigenetics in Neural Disorders
7.1.1. m6A and Its RMPs
m6A is highly enriched in neural tissue and plays an important role in neurogenesis and neuronal disorders [264,265,266,267] (Table 1). Knockout of METTL3 or METTL14 prolongs the cell cycle of embryonic neural stem cells (NSCs)/radial glia cells (RGCs) and inhibits proliferation of NSCs/RGCs, resulting in delay of cortical neurogenesis into postnatal stages in embryonic mouse brains [268,269]. The effects of m6A and its writers on NSC self-renewal and neurogenesis have been linked to histone modifications [270,271]. METTL3/METTL14 and the reader protein YTHDF1 are also important for maintaining the stemness of adult neural stem cells (aESCs) and axon regeneration in adult mice [272]. The m6A eraser protein FTO also plays a critical role in regulation of the proliferation and differentiation of adult neural stem cells (aESCs) [273,274].
Recent studies show that m6A and its RMPs, specifically METTL3, YTHDF1 and FTO, play an important role in regulation of stress responses, formation of hippocampal memories, and development of neuropsychiatric diseases [275,276,277]. Furthermore, the FTO-induced hypo-m6A levels have been shown to activate the TSC1-mTOR-Tau signaling pathway and increased apoptosis of dopaminergic neurons, which contributes to the pathogenesis of Alzheimer’s disease (AD) and Parkinson’s disease (PD), respectively [278,279,280].
7.1.2. RNA Editing and Its RMPs
RNA editing can alter codons and/or splicing sites in pre-mRNAs, resulting in non-functional transcripts or proteins. Earlier studies demonstrated that knockout (KO) of ADAR2 expression could substantially reduce the functional mRNA transcripts from the mutated AMPA receptor gene and rescue mice from mutations-induced lethality [281,282]. More recent studies demonstrate that A-to-I and C-to-U RNA editing enzymes can affect the development and function of neurons and neurotransmitters through modulating mRNA abundance and miRNA binding of neuronal development-related genes and neurotransmitters in neurons [283,284,285] (Table 2).
7.1.3. RNA 2’-O-Methylation (Nm) and Its RMPs
7.1.4. m5C and Its RMPs
Eukaryotic m5C and its RMPs regulate the function of three RNA species: rRNA, tRNA and mRNA [55]. m5C and its writer proteins, NSUNs, are involved in early mouse embryogenesis, especially neurogenesis [290], and the embryonic NSCs of mouse brains shows a distinct m5C pattern in mRNA [53]. NSUN2, a major m5C writer protein, is highly expressed in early neuroepithelial progenitors of the developing human brain, and loss of NSUN2-mediated tRNA:m5C impairs differentiation and motility of NES cells in mouse models [291]. Mutations in the NSUN2 gene cause aberrant tRNA:m5C and autosomal-recessive intellectual disability, called Dubowitz syndrome [292,293,294,295,296] (Table 2).
7.2. RNA Epigenetics in Metabolic Disorders
m6A and Its RMPs
Recent studies show that m6A and its RMPs play a critical role in Type 2 Diabetes (T2D) pathogenesis, and ablation of m6A levels by targeting METTL3 or METTL14 in mouse models recapitulates human T2D by inhibiting the Insulin/IGF1-AKT-PDX1 pathway in pancreatic β-cells, resulting in cell-cycle arrest and impaired glucose-stimulated insulin secretion (GSIS) [297]. METTL14 deletion in β-cells of adult mice results in glucose intolerance [298,299]. WTAP, coupled with METTL3 and METTL14, is increased and distributed in the nucleus through upregulation of cyclin A2, which positively regulates mitotic clonal expansion of adipocytes [300]. The zinc finger transcription factor ZFP217 promotes mitotic clonal expansion (MCE) of adipocytes and adipogenesis by inhibiting CCND1 mRNA expression, which is positively and negatively regulated by METTL3 and YTHDF2, respectively [301] (Table 2).
FTO is well-known to promote adipogenesis, and inactivation of the gene encoding FTO protects mice from obesity [302]. The underlying mechanism has been linked to IRX3, a member of the Iroquois homeobox protein family [303]. FTO-induced adipogenesis is achieved by MCE regulation via the adipogenic isoform of RUNX1T1 [304]. Other molecular mechanisms by which FTO promotes adipogenesis are inhibition of the Wnt/β-catenin signaling in intramuscular preadipocytes [305] and activation of ATG5 and ATG77-mediated autophagy and adipogenesis [306]. Finally, since FTO negatively regulates m6A levels and positively regulates adipogenesis, FTO inhibitors may provide a novel therapeutic strategy for metabolic diseases [307].
7.3. RNA Epigenetics in Immune Disorders and Viral Infections
7.3.1. m6A and Its RMPs
Recent studies demonstrate a critical role of m6A in the regulation of immune disorders and infections [308,309]. The innate immune system detects the nucleic acids, i.e., DNA and RNAs, of various pathogenic organisms via pattern-recognition receptors (PRRs). PRRs include several endosomal members of the Toll-like receptor (TLR) family and cytosolic sensors, such as retinoic acid inducible gene-I (RIG-I) and its homolog melanoma differentiation associated gene 5 (MDA5), as well as RIG-I-like receptors (RLR) [310,311]. RNA modifications, such as m5C, m6A, m5U, s2U or pseudouridine (Ψ), are known to ablate activation of dendritic cells (DCs) to inhibit innate immune system through TLRs [312]. Specifically, METTL3-mediated mRNA:m6A was recently shown to promote dendritic cell activation [313]. METTL3 has also been shown to be involved in T cell homeostasis and differentiation [314]. Depletion of METTL3 in naive T cells decreased m6A in the mRNAs of SOCS family genes encoding the STAT signaling inhibitory proteins SOCS1, SOCS3 and CISH, leading to slower mRNA decay and increased protein expression, consequently inhibiting IL-7-mediated STAT5 activation in the Mettl3-deficient T cells [314]. Another study showed that deletion of METTL3/m6A leads to suppression of IL-2-STAT5 signaling pathway, which controls Treg cell functions and regulates the tumor-killing functions of CD8 T cells [315]. Yet further studies show that METTL3 or YTHDF2 deletion leads to an increase in type I interferon β (IFNB1) mRNA, thus suppressing the propagation of different viruses [316].
Depletion of METTL14 showed a similar effect, stimulating accumulation of dsDNA- or virus-induced IFNB1 mRNA and inhibition of virus reproduction. Alternatively, ALKBH5 had the opposite effect [317]. Since the viral Rev protein preferentially interacts with methylated RRE, decreased or increased m6A levels in HIV-1 RRE due to silencing of METTL3/METTL14 or ALKBH5, respectively, lead to reduced or increased viral RNA export and replication suppression [318] (Table 2).
7.3.2. RNA Editing and Its RMPs
Most A-to-I editing is predominantly located within mobile elements in non-coding parts of the human genome [65] and plays a critical role in innate immunity and protection against self-transcript activation [69,319]. ADAR1 knockout in neuronal progenitor cells increases MDA5 (dsRNA sensor)-dependent spontaneous interferon production, PKR activation, and cell death [320]. Therefore, human ADAR1 regulates sensing of self vs non-self RNA, enabling pathogen detection and autoimmune/inflammation prevention. Loss-of-function mutations in the ADAR1 gene cause Aicardi-Goutières syndrome, a congenital autoimmune disease characterized by encephalopathy and IFN signature [321] (Table 2).
7.3.3. 5′ Cap Nm and CMTR
The 5′ cap structure with m7G (cap 0) and Nm of the first residue (cap 1) is vital for pre-mRNA processing and translation in multicellular eukaryotes and some viruses (Figure 4). The human coronavirus mutants lacking 2’-O-methyltransferase activity induce IFN expression in an MDA5–dependent manner, and these mutants are more sensitive to the antiviral action of IFN [21,322]. IFIT1 (IFN-induced protein with tetratricopeptide repeats-1) is an important effector of innate immune and antiviral response. Its antiviral activity relies on its ability to selectively bind the viral mRNA 5’ cap that lacks Nm in order to compete with translation initiation factor eIF4F binding to the same mRNAs, while remaining inactive against host mRNAs with Nm at the cap 1 and 2 residues [323,324,325,326] (Figure 4). A recent study showed that the SARS-CoV-2 nsp16 and nsp10 complex can carry out the 5′ cap Nm on virally encoded mRNAs to mimic cellular mRNAs, thus protecting the virus from host innate immune restriction [327]. This may provide a potential strategy to develop coronavirus vaccines and antiviral drugs [328].
7.4. RNA Epigenetics in Cancer and Leukemia
7.4.1. m6A and Its RMPs in Cancer
Recent studies have demonstrated that RNA modifications and RMPs play a critical role in controlling cancer cell proliferation, metastasis, immune evasion and drug resistance [329,330,331]. m6A and its RMPs have a bi-faceted role, i.e., acting as either an oncogene or a tumor suppressor in the pathogenesis of solid tumors and leukemia. METTL3 upregulation increases translation of MYC, BCL2 and PTEN mRNAs in acute myeloid leukemia (AML), and METTL3 loss leads to increased levels of phosphorylated AKT, promoting differentiation of hematopoietic stem/progenitor cells (HSPCs) [332] (Table 2). METTL3, independently of METTL14, interacts with the transcription factor CEBPZ and binds the gene promoters occupied by CEBPZ to promote m6A in the CEBPZ-targeting gene transcript, enhancing translation and thereby promoting leukemogenesis [333].
m6A and its RMPs are essential for mRNA processing and miRNA biogenesis [334]. Upregulation of METTTL3 and m6A has been reported to promote tumorigenesis or metastasis through promotion or inhibition of oncogenic or tumor suppressive miRNAs biogenesis, respectively, in various solid tumors, including bladder [335], lung [336], breast [337], gastrointestinal and liver cancers [338,339], and glioblastoma [340]. However, elevated METTL3 expression inhibits self-renewal of bladder tumor initiating cells and bladder tumorigenesis through Notch 1 mRNA:m6A [341].
In contrast to increased m6A/METTL3 in most tumor types, decreased METTL14/m6A has been reported to act as a tumor suppressor, and elevated METTL14 expression has been reported in a variety of solid tumors and leukemia through various mechanisms (Table 2). m6A was reported to be reduced in about 70% of endometrial carcinoma cases. Endometrial cancer is associated with a METTL14 mutation (R298P), which results in decreased expression of the negative AKT regulator PHLPP2, increased expression of the positive AKT regulator mTORC2 and tumor cell proliferation [342]. Increased m6A by METTL14 suppresses renal cancer cell migration and invasion through down-regulation of P2RX6 protein translation and ATP-P2RX6-Ca2+-p-ERK1/2-MMP9 signaling [343]. METTL14 can suppress proliferation and metastasis of colorectal cancer and liver cancer through down-regulating oncogenic long non-coding RNA XIST and miRNAs, respectively [344,345]. METTL3 and METTL14 expression was reported to be downregulated in pediatric acute lymphoblastic leukemia (ALL) with ETV6/RUNX1 gene rearrangement [346]. However, paradoxically, METTL14 was reported to be highly expressed in normal hematopoietic stem/progenitor cells (HSPCs) and AML with t (11q23), t (15;17), or t (8;21) and necessary for development and maintenance of AML and self-renewal of leukemia stem/initiation cells (LSCs/LICs) [347]. Mechanistically, METTL14 exerts its oncogenic role by regulating the SPI1-METTL14-MYB/MYC signaling axis in myelopoiesis and leukemogenesis [347].
Like the m6A writer proteins METTL3 and METTL14, the m6A reader proteins YTHDF1 and YTHDF2 can act as either an oncogene or a tumor suppressor. YTHDF1 promotes cancer cell growth through upregulation of MYC [348] (Table 2). YTHDF1 also increases Wnt/β-catenin signaling and EIF3C-mediated mRNA translation, promoting tumor cell growth in colon cancer and ovarian cancer, respectively [349,350]. Furthermore, YTHDF1 inhibits neoantigen-specific immunity by binding to m6A-masked transcripts of lysosomal proteases. This binding increases lysosomal cathepsins translation, leading to inhibition of antigen presentation of wild-type dendritic cells and CD8+ T cell infiltration, and immune evasion by tumor cells in vivo [351]. YTHDF2 orchestrates epithelial-mesenchymal transition/proliferation dichotomy in pancreatic cancer cells [352] through YAP/TAZ signaling [353]. YTHDF2 regulates the lncRNA LINC00470/METTL3-mediated degradation of the tumor suppressor PTEN mRNA, which promotes cell growth and metastasis in gastric cancer [354]. YTHDC1 has been shown to promote m6A/METTL3-mediated tumor cell growth in glioblastoma (GBM) via regulation of nonsense-mediated mRNA decay (NMD) of serine- and arginine-rich splicing factors (SRSF) [355]. Tumor cell growth and metastasis is mediated by YTHDC2 through increased translation of c-Jun and ATF-2 in liver cancer [356], and HIF-1α in colon cancer [357]. However, reports have also shown that YTHDF1 and YTHDF2 can act as tumor suppressors. For example, YTHDF1 promotes the translation of m6A-marked HINT2 mRNA, a tumor suppressor, which inhibits ocular melanoma progression [358], and YTHDF2 suppresses cell proliferation and growth via destabilizing the EGFR mRNA in hepatocellular carcinoma [359].
In contrast to the plethora of studies on m6A writers and readers in cancer, less is known about the m6A demethylase, FTO. In breast cancer, FTO has been shown to enhance the PI3K/AKT signaling pathway and degrade BNIP3 (a mitochondrial pro-apoptotic protein) mRNA transcripts, promoting cancer cell growth [360,361]. Paradoxically, FTO can also suppress renal cancer cell growth by reducing m6A levels in its mRNA transcripts, which in turn increase PGC-1α expression [362].
7.4.2. Nm and Its RMPs in Cancer
Nm is one of the most common RNA modifications and plays a role in the pathogenic processes of various human diseases [17]. Rare cases of non-small cell lung cancer (NSCLC) harboring CMTR1-ALK fusion have been reported, and do not respond to ALK inhibitor crizotinib [363]. Fibrillarin (FBL), the methyltransferase in the C/D box small nucleolar RNAs (SNORDs)/small nucleolar ribonucleoprotein complexes (snoRNPs), carries out Nm in rRNA [364]. FBL Nm promotion in rRNA increases internal ribosome entry site (IRES)-dependent translation initiation of key cancer genes and tumorigenesis. Meanwhile, p53 acts as a safeguard of protein synthesis by regulating FBL-mediated translation initiation [365]. Long noncoding RNA ZFAS1 was reported to promote small nucleolar RNA-mediated Nm in rRNA via NOP58 recruitment in colorectal cancer [366]. Increased Nm in 18S rRNA has been linked to the pathogenesis of AML [367,368]. Recently, HENMT1 was identified as the methyltransferase responsible for 3’-terminal Nm of mammalian miRNAs, which promotes 3’-terminal Nm in human non-small cell lung cancer (NSCLC) [369] (Table 2).
7.4.3. RNA Editing and Its RMPs in Cancer
A-to-I editing and ADARs are known to contribute to tumorigenesis [370]. Like m6A and its RMPs, the roles of A-to-I editing and ADARs are also multifaceted. ADAR1 is overexpressed in many cancer types, including lung, liver and esophageal cancers and chronic myeloid leukemia (CML), and it acts predominantly as an oncogene. In contrast, ADAR2 expression is reduced in glioblastoma, in which ADAR2 acts as a tumor suppressor [371,372,373]. ADAR1 edits an intronic splicing silencer, leading to recruitment of SRSF7 and repression of exon inclusion, as well as cancer progression [374]. In antizyme inhibitor 1 (AZIN1), ADAR1-induced A-to-I editing causes the serine (S) → glycine (G) substitution at residue 367, leading to a cytoplasmic-to-nuclear translocation and “gain-of-function” phenotypes manifested by augmented tumor initiating potential and more aggressive cancer cell behavior [375]. ADAR1 promotes A-to-I editing in the 3’UTRs of ATM, GINS4 and POLH mRNA transcripts, resulting in increased expression of those oncogenic proteins and cancer cell proliferation in breast cancer [376]. Recently, ADAR1 was identified as a critical factor contributing to drug resistance in cancer cells, and the aberrant expression of ADAR1 promotes resistance to BET inhibitors in pancreatic cancer [377,378,379] (Table 2).
ADAR2 is essential for editing several miRNAs, such as miR-222/221 and the miR-21 precursor in vivo and in vitro, most of them oncogenic miRNAs [380]. However, its editing activity is impaired in glioblastoma cells, and restoring its activity inhibits tumor cell proliferation [380]. ADAR2 editing in tumor suppressor miRNAs, such as miR-589-3p and miRNA-379-5p, leads to increased expression of these tumor suppressor miRNAs [381,382]. Together, the A-to-I editing of these miRNAs by ADAR2 leads to inhibition of glioblastoma cell proliferation, migration and invasion (Table 2).
7.4.4. m5C and Its RMPs in Cancer
RNA:m5C is mediated by NSUN RNA methyltransferase family proteins (Figure 2B) [51]. Upregulation of RNA:m5C and its writer proteins, especially NSUN1 and NSUN2, has been reported in a variety of solid tumors and leukemia and contributes to tumor cell proliferation, metastasis and drug resistance [55,57,331]. However, compared to a large body of studies on the pathophysiological function of m6A and its RMPs, much less is known about m5C and its RMPs.
NSUN1 (aka NOL1, NOP2 and p120) is a well-known tumor proliferation associated nucleolar protein [383,384] and its expression levels predict breast cancer disease progress and patient survival [385]. Our recent study demonstrates that NSUN1 expression is elevated in leukemia cells, and NSUN1 interacts with BRD4 to recruit elongating RNA polymerase II (eRNAPII) to nascent RNA, forming an active chromatin structure and transcription complex that mediates drug resistance in myeloid leukemia [127]. NSUN1 increases expression of oncogenic lncRNAs PVT1 and/or LINC00963 and inhibits expression of tumor suppressor miRNAs, leading to prostate cancer cell growth and metastasis [386,387]. Finally, NSUN1 can also interact with RPL6 to promote gallbladder cancer progression [388].
NSUN2 is responsible for the majority of mRNA:m5C in mammalian cells and is upregulated in various cancer types [57,389]. NSUN2 was found to be a downstream regulator of MYC and promotes cancer cell proliferation by RNAPIII methylation [390]. In bladder cancer, NSUN2 is involved in pathogenesis promotion through m5C deposition in the 3’ UTR of hepatoma-derived growth factor (HDGF). HDGF stabilizes the mRNA transcripts marked with m5C, which is selectively bound by YBX1 protein [391,392]. NSUN2-induced m5C in CDK1 mRNA promotes CDK1 translation, leading to tumor cell proliferation [392,393,394].
8. Conclusions and Future Directions
RNA modifications and RMPs are distributed widely in various subcellular organelles, such as nuclear particles, ribosomes and mitochondria, and identifying the role of RNA epigenetics in coordinating the functions of these subcellular organelles is likely to play an important part in more accurately characterizing multiple cellular functions and disease processes. It is increasingly clear that the role of RNA epigenetics is multifaceted and context-dependent in chromatin remodeling and gene expression. Although great progress has been made in elucidating the many roles of RNA epigenetics physiologically, the detailed mechanisms by which RNA modifications and RMPs control cancer progression and drug resistance remain largely unknown. There is still little known about how selective hyper- or hypo-regulation of RNA modifications in various RNA species contributes to tumor progression and therapeutic response/resistance. Great potential exists for the use of RNA modifications and RMPs as diagnostic/prognostic tools and therapeutic targets in future cancer therapy. However, given the large number of RNA modifications, an enormous amount of work lies ahead to fully understand the pathophysiological function of RNA modifications and their RMPs in cancer.
Author Contributions
J.X.C. conceived the review article; J.X.C., A.W. and S.W. contributed to writing and reviewing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The Taub Foundation Grants Program for MDS, the Cancer Research Foundation, the Michael Reese Bench to Bedside Award, and the CTSA-ITA Core Subsidies from the University of Chicago ITM grant (#UL1TR002389).
Acknowledgments
This work was sponsored by the grants from the Taub Foundation Grants Program for MDS, the Cancer Research Foundation, the Michael Reese Bench to Bedside Award, and the CTSA-ITA Core Subsidies from the University of Chicago ITM grant (#UL1TR002389) to J.X.C. The authors gratefully acknowledge the funding support from these resources.
Conflicts of Interest
No conflicts of interest declared by the authors.
References
- Li, B.; Carey, M.; Workman, J.L. The Role of Chromatin during Transcription. Cell 2007, 128, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Kato, M.; Carninci, P. Genome-Wide Technologies to Study RNA–Chromatin Interactions. Non Coding RNA 2020, 6, 20. [Google Scholar] [CrossRef]
- Holoch, D.; Moazed, D. RNA-mediated epigenetic regulation of gene expression. Nat. Rev. Genet. 2015, 16, 71–84. [Google Scholar] [CrossRef]
- Ipsaro, J.J.; Joshua-Tor, L. From guide to target: Molecular insights into eukaryotic RNA-interference machinery. Nat. Struct. Mol. Biol. 2015, 22, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Martienssen, R.; Moazed, D. RNAi and heterochromatin assembly. Cold Spring Harb. Perspect. Biol. 2015, 7, a019323. [Google Scholar] [CrossRef] [PubMed]
- Holoch, D.; Moazed, D. Small-RNA loading licenses Argonaute for assembly into a transcriptional silencing complex. Nat. Struct. Mol. Biol. 2015, 22, 328–335. [Google Scholar] [CrossRef]
- Thakur, J.; Henikoff, S. Architectural RNA in chromatin organization. Biochem. Soc. Trans. 2020, 48, 1967–1978. [Google Scholar] [CrossRef]
- Dykes, I.M.; Emanueli, C. Transcriptional and Post-transcriptional Gene Regulation by Long Non-coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fu, X.-D. Chromatin-associated RNAs as facilitators of functional genomic interactions. Nat. Rev. Genet. 2019, 20, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Frye, M.; Harada, B.T.; Behm, M.; He, C. RNA modifications modulate gene expression during development. Science 2018, 361, 1346–1349. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, W.V.; Bell, T.A.; Schaening, C. Messenger RNA modifications: Form, distribution, and function. Science 2016, 352, 1408–1412. [Google Scholar] [CrossRef] [Green Version]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA Modifications in Gene Expression Regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hocine, S.; Singer, R.H.; Grünwald, D. RNA Processing and Export. Cold Spring Harb. Perspect. Biol. 2010, 2, a000752. [Google Scholar] [CrossRef] [Green Version]
- Gonatopoulos-Pournatzis, T.; Cowling, V.H. Cap-binding complex (CBC). Biochem. J. 2014, 457, 231–242. [Google Scholar] [CrossRef] [Green Version]
- Thompson, S.R. Tricks an IRES uses to enslave ribosomes. Trends Microbiol. 2012, 20, 558–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitrova, D.G.; Teysset, L.; Carré, C. RNA 2’-O-Methylation (Nm) Modification in Human Diseases. Genes 2019, 10, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bélanger, F.; Stepinski, J.; Darzynkiewicz, E.; Pelletier, J. Characterization of hMTr1, a Human Cap1 2′-O-Ribose Methyltransferase*. J. Biol. Chem. 2010, 285, 33037–33044. [Google Scholar] [CrossRef] [Green Version]
- Werner, M.; Purta, E.; Kaminska, K.H.; Cymerman, I.A.; Campbell, D.A.; Mittra, B.; Zamudio, J.R.; Sturm, N.R.; Jaworski, J.; Bujnicki, J.M. 2′-O-ribose methylation of cap2 in human: Function and evolution in a horizontally mobile family. Nucleic Acids Res. 2011, 39, 4756–4768. [Google Scholar] [CrossRef] [Green Version]
- Daffis, S.; Szretter, K.J.; Schriewer, J.; Li, J.; Youn, S.; Errett, J.; Lin, T.-Y.; Schneller, S.W.; Zust, R.; Dong, H.; et al. 2′-O methylation of the viral mRNA cap evades host restriction by IFIT family members. Nat. Cell Biol. 2010, 468, 452–456. [Google Scholar] [CrossRef]
- Züst, R.; Cervantes-Barragan, L.; Habjan, M.; Maier, R.; Neuman, B.W.; Ziebuhr, J.; Szretter, K.J.; Baker, S.C.; Barchet, W.; Diamond, M.S.; et al. Ribose 2′-O-methylation provides a molecular signature for the distinction of self and non-self mRNA dependent on the RNA sensor Mda5. Nat. Immunol. 2011, 12, 137–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuberth-Wagner, C.; Ludwig, J.; Bruder, A.K.; Herzner, A.-M.; Zillinger, T.; Goldeck, M.; Schmidt, T.; Schmid-Burgk, J.L.; Kerber, R.; Wolter, S.; et al. A Conserved Histidine in the RNA Sensor RIG-I Controls Immune Tolerance to N1-2′O-Methylated Self RNA. Immunity 2015, 43, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devarkar, S.C.; Wang, C.; Miller, M.T.; Ramanathan, A.; Jiang, F.; Khan, A.G.; Patel, S.S.; Marcotrigiano, J. Structural basis for m7G recognition and 2’-O-methyl discrimination in capped RNAs by the in-nate immune receptor RIG-I. Proc. Natl. Acad. Sci. USA 2016, 113, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.-M.; Gershowitz, A.; Moss, B. Methylated nucleotides block 5′ terminus of HeLa cell messenger RNA. Cell 1975, 4, 379–386. [Google Scholar] [CrossRef]
- Mauer, J.; Luo, X.; Blanjoie, A.; Jiao, X.; Grozhik, A.V.; Patil, D.P.; Linder, B.; Pickering, B.F.; Vasseur, J.-J.; Chen, Q.; et al. Reversible methylation of m6Am in the 5′ cap controls mRNA stability. Nat. Cell Biol. 2017, 541, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akichika, S.; Hirano, S.; Shichino, Y.; Suzuki, T.; Nishimasu, H.; Ishitani, R.; Sugita, A.; Hirose, Y.; Iwasaki, S.; Nureki, O.; et al. Cap-specific terminal N (6)-methylation of RNA by an RNA polymerase II-associated methyltrans-ferase. Science 2019, 363. [Google Scholar] [CrossRef] [PubMed]
- Hirose, Y.; Iwamoto, Y.; Sakuraba, K.; Yunokuchi, I.; Harada, F.; Ohkuma, Y. Human phosphorylated CTD-interacting protein, PCIF1, negatively modulates gene expression by RNA polymerase II. Biochem. Biophys. Res. Commun. 2008, 369, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Cowling, V.H. CAPAM: The mRNA Cap Adenosine N6-Methyltransferase. Trends Biochem. Sci. 2019, 44, 183–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holley, R.W.; Everett, G.A.; Madison, J.T.; Zamir, A. Nucleotide sequences in the yeast alanine transfer ribonucleic acid. J. Biol. Chem. 1965, 240, 2122–2128. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive Analysis of mRNA Methylation Reveals Enrichment in 3′ UTRs and near Stop Codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3–METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Feng, J.; Xue, Y.; Guan, Z.; Zhang, D.; Liu, Z.; Gong, Z.; Huang, J.; Tang, Z.G.C.; Zou, T.; et al. Structural basis of N6-adenosine methylation by the METTL3–METTL14 complex. Nat. Cell Biol. 2016, 534, 575–578. [Google Scholar] [CrossRef]
- Zaccara, S.; Ries, R.J.; Jaffrey, S.R. Reading, writing and erasing mRNA methylation. Nat. Rev. Mol. Cell Biol. 2019, 20, 608–624. [Google Scholar] [CrossRef]
- Schöller, E.; Weichmann, F.; Treiber, T.; Ringle, S.; Treiber, N.; Flatley, A.; Feederle, R.; Bruckmann, A.; Meister, G. Interactions, localization, and phosphorylation of the m6A generating METTL3–METTL14–WTAP complex. RNA 2018, 24, 499–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pendleton, K.E.; Chen, B.; Liu, K.; Hunter, O.V.; Xie, Y.; Tu, B.P.; Conrad, N.K. The U6 snRNA m 6 A Methyltransferase METTL16 Regulates SAM Synthetase Intron Retention. Cell 2017, 169, 824–835.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruszkowska, A.; Ruszkowski, M.; Dauter, Z.; Brown, J.A. Structural insights into the RNA methyltransferase domain of METTL16. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendel, M.; Chen, K.-M.; Homolka, D.; Gos, P.; Pandey, R.R.; McCarthy, A.A.; Pillai, R.S. Methylation of Structured RNA by the m(6)A Writer METTL16 Is Essential for Mouse Embryonic Development. Mol. Cell 2018, 71, 986–1000.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, S.; Sun, H.; Xu, C. YTH Domain: A Family of N 6 -methyladenosine (m 6 A) Readers. Genom. Proteom. Bioinform. 2018, 16, 99–107. [Google Scholar] [CrossRef]
- Wu, B.; Su, S.; Patil, D.P.; Liu, H.; Gan, J.; Jaffrey, S.R.; Ma, J. Molecular basis for the specific and multivariant recognitions of RNA substrates by human hnRNP A2/B1. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Dai, Q.; Zheng, G.; He, C.; Parisien, M.; Pan, T. N6-methyladenosine-dependent RNA structural switches regulate RNA–protein interactions. Nat. Cell Biol. 2015, 518, 560–564. [Google Scholar] [CrossRef] [Green Version]
- Zarnack, K.; König, J.; Tajnik, M.; Martincorena, I.; Eustermann, S.; Stévant, I.; Reyes, A.; Anders, S.; Luscombe, N.M.; Ule, J. Direct Competition between hnRNP C and U2AF65 Protects the Transcriptome from the Exonization of Alu Elements. Cell 2013, 152, 453–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 facilitates translation and decay of N6-methyladenosine-modified RNA. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasowitz, S.D.; Ma, J.; Anderson, S.J.; Leu, N.A.; Xu, Y.; Gregory, B.D.; Schultz, R.M.; Wang, P.J. Nuclear m6A reader YTHDC1 regulates alternative polyadenylation and splicing during mouse oocyte development. PLoS Genet. 2018, 14, e1007412. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Wang, X.; Liu, K.; Roundtree, I.A.; Tempel, W.; Li, Y.; Lu, Z.; He, C.; Min, J. Structural basis for selective binding of m6A RNA by the YTHDC1 YTH domain. Nat. Chem. Biol. 2014, 10, 927–929. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Gerken, T.; Girard, C.A.; Tung, Y.-C.L.; Webby, C.J.; Saudek, V.; Hewitson, K.S.; Yeo, G.S.H.; McDonough, M.A.; Cunliffe, S.; McNeill, L.A.; et al. The Obesity-Associated FTO Gene Encodes a 2-Oxoglutarate-Dependent Nucleic Acid Demethylase. Science 2007, 318, 1469–1472. [Google Scholar] [CrossRef] [Green Version]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.-G.; et al. N6-Methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef]
- Tang, C.; Klukovich, R.; Peng, H.; Wang, Z.; Yu, T.; Zhang, Y.; Zheng, H.; Klungland, A.; Yan, W. ALKBH5-dependent m6A demethylation controls splicing and stability of long 3′-UTR mRNAs in male germ cells. Proc. Natl. Acad. Sci. USA 2018, 115, E325–E333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosovic, M.; Molares, H.C.; Gregorova, P.; Hrossova, D.; Kudla, G.; Vanacova, S. N6-methyladenosine demethylase FTO targets pre-mRNAs and regulates alternative splicing and 3’-end processing. Nucleic Acids Res. 2017, 45, 11356–11370. [Google Scholar] [CrossRef]
- Motorin, Y.; Lyko, F.; Helm, M. 5-methylcytosine in RNA: Detection, enzymatic formation and biological functions. Nucleic Acids Res. 2009, 38, 1415–1430. [Google Scholar] [CrossRef] [Green Version]
- Trixl, L.; Lusser, A. The dynamic RNA modification 5-methylcytosine and its emerging role as an epitranscriptomic mark. Wiley Interdiscip. Rev. RNA 2018, 10, e1510. [Google Scholar] [CrossRef] [Green Version]
- Squires, J.E.; Patel, H.R.; Nousch, M.; Sibbritt, T.; Humphreys, D.T.; Parker, B.J.; Suter, C.M.; Preiss, T. Widespread occurrence of 5-methylcytosine in human coding and non-coding RNA. Nucleic Acids Res. 2012, 40, 5023–5033. [Google Scholar] [CrossRef]
- Amort, T.; Rieder, D.; Wille, A.; Khokhlova-Cubberley, D.; Riml, C.; Trixl, L.; Jia, X.-Y.; Micura, R.; Lusser, A. Distinct 5-methylcytosine profiles in poly(A) RNA from mouse embryonic stem cells and brain. Genome Biol. 2017, 18, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, T.; Chen, W.; Liu, J.; Gu, N.; Zhang, R. Genome-wide identification of mRNA 5-methylcytosine in mammals. Nat. Struct. Mol. Biol. 2019, 26, 380–388. [Google Scholar] [CrossRef]
- Bohnsack, K.E.; Höbartner, C.; Bohnsack, M.T. Eukaryotic 5-methylcytosine (m5C) RNA Methyltransferases: Mechanisms, Cellular Functions, and Links to Disease. Genes 2019, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Wang, W. mRNA methylation by NSUN2 in cell proliferation. Wiley Interdiscip. Rev. RNA 2016, 7, 838–842. [Google Scholar] [CrossRef] [PubMed]
- Chellamuthu, A.; Gray, S.G. The RNA Methyltransferase NSUN2 and Its Potential Roles in Cancer. Cells 2020, 9, 1758. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.J.; Long, T.; Li, J.; Li, H.; Wang, E.D. Structural basis for substrate binding and catalytic mechanism of a human RNA:m5C methyltransferase NSun6. Nucleic Acids Res. 2017, 45, 6684–6697. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yang, Y.; Sun, B.-F.; Chen, Y.-S.; Xu, J.-W.; Lai, W.-Y.; Li, A.; Wang, X.; Bhattarai, D.P.; Xiao, W.; et al. 5-methylcytosine promotes mRNA export—NSUN2 as the methyltransferase and ALYREF as an m5C reader. Cell Res. 2017, 27, 606–625. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Gonzalez, G.; Li, L.; Li, J.; You, C.; Miao, W.; Hu, J.; Fu, L.; Zhao, Y.; Li, R.; et al. YTHDF2 Binds to 5-Methylcytosine in RNA and Modulates the Maturation of Ribosomal RNA. Anal. Chem. 2019, 92, 1346–1354. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Han, X.; Yang, W.-L.; Zhang, M.; Ma, H.-L.; Sun, B.-F.; Li, A.; Xia, J.; Chen, J.; et al. RNA 5-Methylcytosine Facilitates the Maternal-to-Zygotic Transition by Preventing Maternal mRNA Decay. Mol. Cell 2019, 75, 1188–1202.e11. [Google Scholar] [CrossRef]
- Shen, H.; Ontiveros, R.J.; Owens, M.C.; Liu, M.Y.; Ghanty, U.; Kohli, R.M.; Liu, K.F. TET-mediated 5-methylcytosine oxidation in tRNA promotes translation. J. Biol. Chem. 2021, 296, 100087. [Google Scholar] [CrossRef]
- Lan, J.; Rajan, N.; Bizet, M.; Penning, A.; Singh, N.K.; Guallar, D.; Calonne, E.; Greci, A.L.; Bonvin, E.; Deplus, R.; et al. Functional role of Tet-mediated RNA hydroxymethylcytosine in mouse ES cells and during differentiation. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. Functions and Regulation of RNA Editing by ADAR Deaminases. Annu. Rev. Biochem. 2010, 79, 321–349. [Google Scholar] [CrossRef] [Green Version]
- Tan, M.H.; Li, Q.; Shanmugam, R.; Piskol, R.; Kohler, J.; Young, A.N.; Liu, K.I.; Zhang, R.; Ramaswami, G.; Ariyoshi, K.; et al. Dynamic landscape and regulation of RNA editing in mammals. Nat. Cell Biol. 2017, 550, 249–254. [Google Scholar] [CrossRef]
- Ganem, N.S.; Lamm, A.T. A-to-I RNA editing—Thinking beyond the single nucleotide. RNA Biol. 2017, 14, 1690–1694. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.-H.E.; Bahn, J.H.; Yang, Y.; Lin, X.; Tran, S.; Yang, E.-W.; Quinones-Valdez, G.; Xiao, X. RNA editing in nascent RNA affects pre-mRNA splicing. Genome Res. 2018, 28, 812–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, O.; Di Segni, A.; Cesarkas, K.; Porath, H.T.; Marcu-Malina, V.; Mizrahi, O.; Stern-Ginossar, N.; Kol, N.; Farage-Barhom, S.; Glick-Saar, E.; et al. RNA editing by ADAR1 leads to context-dependent transcriptome-wide changes in RNA secondary structure. Nat. Commun. 2017, 8, 1–14. [Google Scholar] [CrossRef]
- Eisenberg, E.; Levanon, E.Y. A-to-I RNA editing—Immune protector and transcriptome diversifier. Nat. Rev. Genet. 2018, 19, 473–490. [Google Scholar] [CrossRef]
- Oakes, E.; Anderson, A.; Cohen-Gadol, A.; Hundley, H.A. Adenosine Deaminase That Acts on RNA 3 (ADAR3) Binding to Glutamate Receptor Subunit B Pre-mRNA Inhibits RNA Editing in Glioblastoma. J. Biol. Chem. 2017, 292, 4326–4335. [Google Scholar] [CrossRef] [Green Version]
- Mladenova, D.; Barry, G.; Konen, L.M.; Pineda, S.S.; Guennewig, B.; Avesson, L.; Zinn, R.; Schonrock, N.; Bitar, M.; Jonkhout, N.; et al. Adar3 Is Involved in Learning and Memory in Mice. Front. Neurosci. 2018, 12, 243. [Google Scholar] [CrossRef] [Green Version]
- Xiang, J.-F.; Yang, Q.; Liu, C.-X.; Wu, M.; Chen, L.-L.; Yang, L. N6-Methyladenosines Modulate A-to-I RNA Editing. Mol. Cell 2018, 69, 126–135.e36. [Google Scholar] [CrossRef]
- Holland, S.J.; Berghuis, L.M.; King, J.J.; Iyer, L.M.; Sikora, K.; Fifield, H.; Peter, S.; Quinlan, E.M.; Sugahara, F.; Shingate, P.; et al. Expansions, diversification, and interindividual copy number variations of AID/APOBEC family cytidine deaminase genes in lampreys. Proc. Natl. Acad. Sci. USA 2018, 115, E3211–E3220. [Google Scholar] [CrossRef] [Green Version]
- Blanc, V.; Park, E.; Schaefer, S.; Miller, M.; Lin, Y.; Kennedy, S.; Billing, A.M.; Ben Hamidane, H.; Graumann, J.; Mortazavi, A.; et al. Genome-wide identification and functional analysis of Apobec-1-mediated C-to-U RNA editing in mouse small intestine and liver. Genome Biol. 2014, 15, R79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wedekind, J.E.; Dance, G.S.; Sowden, M.; Smith, H.C. Messenger RNA editing in mammals: New members of the APOBEC family seeking roles in the family business. Trends Genet. 2003, 19, 207–216. [Google Scholar] [CrossRef]
- Vu, L.T.; Tsukahara, T. C-to-U editing and site-directed RNA editing for the correction of genetic mutations. Biosci. Trends 2017, 11, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Chester, A.; Weinreb, V.; CARTER, C.W.; Navaratnam, N. Optimization of apolipoprotein B mRNA editing by APOBEC1 apoenzyme and the role of its auxil-iary factor, ACF. RNA 2004, 10, 1399–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wade, J.T.; Struhl, K. The transition from transcriptional initiation to elongation. Curr. Opin. Genet. Dev. 2008, 18, 130–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soutourina, J. Transcription regulation by the Mediator complex. Nat. Rev. Mol. Cell Biol. 2018, 19, 262–274. [Google Scholar] [CrossRef]
- Meinhart, A.; Cramer, P. Recognition of RNA polymerase II carboxy-terminal domain by 3′-RNA-processing factors. Nat. Cell Biol. 2004, 430, 223–226. [Google Scholar] [CrossRef]
- Wong, K.H.; Jin, Y.; Struhl, K. TFIIH Phosphorylation of the Pol II CTD Stimulates Mediator Dissociation from the Preinitiation Complex and Promoter Escape. Mol. Cell 2014, 54, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Haberle, V.; Stark, A. Eukaryotic core promoters and the functional basis of transcription initiation. Nat. Rev. Mol. Cell Biol. 2018, 19, 621–637. [Google Scholar] [CrossRef] [PubMed]
- Missra, A.; Gilmour, D.S. Interactions between DSIF (DRB sensitivity inducing factor), NELF (negative elongation factor), and the Drosophila RNA polymerase II transcription elongation complex. Proc. Natl. Acad. Sci. USA 2010, 107, 11301–11306. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, Y.; Shibata, H.; Handa, H. Transcription elongation factors DSIF and NELF: Promoter-proximal pausing and beyond. Biochim. Biophys. Acta 2013, 1829, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Vos, S.M.; Farnung, L.; Urlaub, H.; Cramer, P. Structure of paused transcription complex Pol II-DSIF-NELF. Nature 2018, 560, 601–606. [Google Scholar] [CrossRef]
- Bernecky, C.; Plitzko, J.M.; Cramer, P. Cramer, Structure of a transcribing RNA polymerase II-DSIF complex reveals a multi-dentate DNA-RNA clamp. Nat. Struct. Mol. Biol. 2017, 24, 809–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterlin, B.M.; Price, D.H. Controlling the Elongation Phase of Transcription with P-TEFb. Mol. Cell 2006, 23, 297–305. [Google Scholar] [CrossRef]
- Wada, T.; Takagi, T.; Yamaguchi, Y.; Watanabe, D.; Handa, H. Evidence that P-TEFb alleviates the negative effect of DSIF on RNA polymerase II-dependent transcrip-tion in vitro. EMBO J. 1998, 17, 7395–7403. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Lin, C.; Shilatifard, A. The super elongation complex (SEC) family in transcriptional control. Nat. Rev. Mol. Cell Biol. 2012, 13, 543–547. [Google Scholar] [CrossRef]
- Kizer, K.O.; Phatnani, H.P.; Shibata, Y.; Hall, H.; Greenleaf, A.L.; Strahl, B.D. A novel domain in Set2 mediates RNA polymerase II interaction and couples histone H3 K36 methyl-ation with transcript elongation. Mol. Cell. Biol. 2005, 25, 3305–3316. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, S.; Workman, J.L. Histone exchange, chromatin structure and the regulation of transcription. Nat. Rev. Mol. Cell Biol. 2015, 16, 178–189. [Google Scholar] [CrossRef]
- Santos-Pereira, J.M.; Aguilera, A. R loops: New modulators of genome dynamics and function. Nat. Rev. Genet. 2015, 16, 583–597. [Google Scholar] [CrossRef]
- Wongsurawat, T.; Jenjaroenpun, P.; Kwoh, C.K.; Kuznetsov, V. Quantitative model of R-loop forming structures reveals a novel level of RNA-DNA interac-tome complexity. Nucleic Acids Res. 2012, 40, e16. [Google Scholar] [CrossRef] [Green Version]
- Fazzio, T.G. Regulation of chromatin structure and cell fate by R-loops. Transcription 2016, 7, 121–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.B.; Chen, H.V.; Acharya, D.; Rando, O.J.; Fazzio, T.G. R loops regulate promoter-proximal chromatin architecture and cellular differentiation. Nat. Struct. Mol. Biol. 2015, 22, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- García-Muse, T.; Aguilera, A. R Loops: From Physiological to Pathological Roles. Cell 2019, 179, 604–618. [Google Scholar] [CrossRef] [PubMed]
- Ginno, P.A.; Lott, P.L.; Christensen, H.C.; Korf, I.; Chédin, F. R-Loop Formation Is a Distinctive Characteristic of Unmethylated Human CpG Island Promoters. Mol. Cell 2012, 45, 814–825. [Google Scholar] [CrossRef] [Green Version]
- Sanz, L.A.; Hartono, S.R.; Lim, Y.W.; Steyaert, S.; Rajpurkar, A.; Ginno, P.A.; Xu, X.; Chédin, F. Prevalent, Dynamic, and Conserved R-Loop Structures Associate with Specific Epigenomic Signatures in Mammals. Mol. Cell 2016, 63, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Chédin, F. Nascent Connections: R-Loops and Chromatin Patterning. Trends Genet. 2016, 32, 828–838. [Google Scholar] [CrossRef] [Green Version]
- Boque-Sastre, R.; Soler, M.; Oliveira-Mateos, C.; Portela, A.; Moutinho, C.; Sayols, S.; Villanueva, A.; Esteller, M.; Guil, S. Head-to-head antisense transcription and R-loop formation promotes transcriptional activation. Proc. Natl. Acad. Sci. USA 2015, 112, 5785–5790. [Google Scholar] [CrossRef] [Green Version]
- Rondón, A.G.; Aguilera, A. R-Loops as Promoters of Antisense Transcription. Mol. Cell 2019, 76, 529–530. [Google Scholar] [CrossRef] [PubMed]
- Arab, K.; Karaulanov, E.; Musheev, M.; Trnka, P.; Schäfer, A.; Grummt, I.; Niehrs, C. GADD45A binds R-loops and recruits TET1 to CpG island promoters. Nat. Genet. 2019, 51, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Castellano-Pozo, M.; Santos-Pereira, J.M.; Rondón, A.G.; Barroso, S.; Andújar, E.; Pérez-Alegre, M.; García-Muse, T.; Aguilera, A. R Loops Are Linked to Histone H3 S10 Phosphorylation and Chromatin Condensation. Mol. Cell 2013, 52, 583–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tous, C.; Aguilera, A. Impairment of transcription elongation by R-loops in vitro. Biochem. Biophys. Res. Commun. 2007, 360, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Niehrs, C.; Luke, B. Regulatory R-loops as facilitators of gene expression and genome stability. Nat. Rev. Mol. Cell Biol. 2020, 21, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Allison, D.F.; Wang, G.G. R-loops: Formation, function, and relevance to cell stress. Cell Stress 2019, 3, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hage, A.; Webb, S.; Kerr, A.; Tollervey, D. Genome-Wide Distribution of RNA-DNA Hybrids Identifies RNase H Targets in tRNA Genes, Retrotransposons and Mitochondria. PLoS Genet. 2014, 10, e1004716. [Google Scholar] [CrossRef]
- Aguilera, A.; García-Muse, T. R Loops: From Transcription Byproducts to Threats to Genome Stability. Mol. Cell 2012, 46, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Ginno, P.A.; Lim, Y.W.; Lott, P.L.; Korf, I.; Chédin, F. GC skew at the 5’ and 3’ ends of human genes links R-loop formation to epigenetic regulation and transcription termination. Genome Res. 2013, 23, 1590–1600. [Google Scholar] [CrossRef] [Green Version]
- Skourti-Stathaki, K.; Triglia, E.T.; Warburton, M.; Voigt, P.; Bird, A.; Pombo, A. R-Loops Enhance Polycomb Repression at a Subset of Developmental Regulator Genes. Mol. Cell 2019, 73, 930–945.e4. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; McBride, K.M.; Hensley, S.; Lu, Y.; Chedin, F.; Bedford, M.T. Arginine Methylation Facilitates the Recruitment of TOP3B to Chromatin to Prevent R Loop Accumulation. Mol. Cell 2014, 53, 484–497. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.Y.; Gish, G.; Braunschweig, U.; Li, Y.; Ni, Z.; Schmitges, F.W.; Zhong, G.; Liu, K.; Li, W.; Moffat, J.; et al. SMN and symmetric arginine dimethylation of RNA polymerase II C-terminal domain control termination. Nature 2016, 529, 48–53. [Google Scholar] [CrossRef]
- Rahl, P.B.; Lin, C.Y.; Seila, A.C.; Flynn, R.A.; McCuine, S.; Burge, C.B.; Sharp, P.A.; Young, R.A. c-Myc Regulates Transcriptional Pause Release. Cell 2010, 141, 432–445. [Google Scholar] [CrossRef] [Green Version]
- Rahl, P.B.; Young, R.A. MYC and Transcription Elongation. Cold Spring Harb. Perspect. Med. 2014, 4, a020990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, S.; Cowling, V.H. Myc and mRNA capping. Biochim. Biophys. Acta 2015, 1849, 501–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herold, S.; Kalb, J.; Büchel, G.; Ade, C.P.; Baluapuri, A.; Xu, J.; Koster, J.; Solvie, D.; Carstensen, A.; Klotz, C.; et al. Recruitment of BRCA1 limits MYCN-driven accumulation of stalled RNA polymerase. Nat. Cell Biol. 2019, 567, 545–549. [Google Scholar] [CrossRef] [PubMed]
- C. Quaresma, A.J.; Bugai, A.; Barboric, M. Cracking the control of RNA polymerase II elongation by 7SK snRNP and P-TEFb. Nucleic Acids Res. 2016, 44, 7527–7539. [Google Scholar]
- McNamara, R.P.; Bacon, C.W.; D’Orso, I. Transcription elongation control by the 7SK snRNP complex: Releasing the pause. Cell Cycle 2016, 15, 2115–2123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterlin, B.M.; Brogie, J.E.; Price, D.H. 7SK snRNA: A noncoding RNA that plays a major role in regulating eukaryotic tran-scription. Wiley Interdiscip. Rev. RNA 2012, 3, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Barboric, M.; Kohoutek, J.; Price, J.P.; Blazek, D.; Price, D.H.; Peterlin, B.M. Interplay between 7SK snRNA and oppositely charged regions in HEXIM1 direct the inhibition of P-TEFb. EMBO J. 2005, 24, 4291–4303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhalluin, C.; Carlson, J.E.; Zeng, L.; He, C.; Aggarwal, A.K.; Zhou, M.-M. Structure and ligand of a histone acetyltransferase bromodomain. Nat. Cell Biol. 1999, 399, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Ma, Q.; Wong, K.; Li, W.; Ohgi, K.; Zhang, J.; Aggarwal, A.K.; Rosenfeld, M.G. Brd4 and JMJD6-Associated Anti-Pause Enhancers in Regulation of Transcriptional Pause Release. Cell 2013, 155, 1581–1595. [Google Scholar] [CrossRef] [Green Version]
- Dow, E.C.; Liu, H.; Rice, A.P. T-loop phosphorylated Cdk9 localizes to nuclear speckle domains which may serve as sites of active P-TEFb function and exchange between the Brd4 and 7SK/HEXIM1 regulatory complexes. J. Cell. Physiol. 2010, 224, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Yik, J.H.N.; Chen, R.; He, N.; Jang, M.K.; Ozato, K.; Zhou, Q. Recruitment of P-TEFb for Stimulation of Transcriptional Elongation by the Bromodomain Protein Brd4. Mol. Cell 2005, 19, 535–545. [Google Scholar] [CrossRef]
- Jang, M.K.; Mochizuki, K.; Zhou, M.; Jeong, H.-S.; Brady, J.N.; Ozato, K. The Bromodomain Protein Brd4 Is a Positive Regulatory Component of P-TEFb and Stimulates RNA Polymerase II-Dependent Transcription. Mol. Cell 2005, 19, 523–534. [Google Scholar] [CrossRef]
- Devaiah, B.N.; Lewis, B.A.; Cherman, N.; Hewitt, M.C.; Albrecht, B.K.; Robey, P.G.; Ozato, K.; Sims, R.J.; Singer, D.S. BRD4 is an atypical kinase that phosphorylates Serine2 of the RNA Polymerase II carboxy-terminal domain. Proc. Natl. Acad. Sci. USA 2012, 109, 6927–6932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.X.; Chen, L.; Li, Y.; Cloe, A.; Yue, M.; Wei, J.; Watanabe, K.A.; Shammo, J.M.; Anastasi, J.; Shen, Q.J.; et al. RNA cytosine methylation and methyltransferases mediate chromatin organization and 5-azacytidine response and resistance in leukaemia. Nat. Commun. 2018, 9, 1163. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.C.; Kong, Y.W.; Huang, Q.; Han, T.-L.V.; Maffa, A.D.; Kasper, E.M.; Yaffe, M.B. BRD4 prevents the accumulation of R-loops and protects against transcription–replication collision events and DNA damage. Nat. Commun. 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Licklider, L.J.; Gygi, S.P.; Reed, R. Comprehensive proteomic analysis of the human spliceosome. Nat. Cell Biol. 2002, 419, 182–185. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, M.E.; Charenton, C.; Nagai, K. RNA Splicing by the Spliceosome. Annu. Rev. Biochem. 2020, 89, 359–388. [Google Scholar] [CrossRef]
- Naftelberg, S.; Schor, I.E.; Ast, G.; Kornblihtt, A.R. Regulation of Alternative Splicing Through Coupling with Transcription and Chromatin Structure. Annu. Rev. Biochem. 2015, 84, 165–198. [Google Scholar] [CrossRef] [PubMed]
- Baralle, F.E.; Giudice, J. Alternative splicing as a regulator of development and tissue identity. Nat. Rev. Mol. Cell Biol. 2017, 18, 437–451. [Google Scholar] [CrossRef]
- Hsin, J.-P.; Manley, J.L. The RNA polymerase II CTD coordinates transcription and RNA processing. Genes Dev. 2012, 26, 2119–2137. [Google Scholar] [CrossRef] [Green Version]
- Herzel, L.; Ottoz, D.S.M.; Alpert, T.; Neugebauer, K.M. Splicing and transcription touch base: Co-transcriptional spliceosome assembly and function. Nat. Rev. Mol. Cell Biol. 2017, 18, 637–650. [Google Scholar] [CrossRef]
- Lee, Y.; Rio, D.C. Mechanisms and Regulation of Alternative Pre-mRNA Splicing. Annu. Rev. Biochem. 2015, 84, 291–323. [Google Scholar] [CrossRef] [Green Version]
- Kondo, Y.; Oubridge, C.; van Roon, A.-M.M.; Nagai, K. Crystal structure of human U1 snRNP, a small nuclear ribonucleoprotein particle, reveals the mechanism of 5′ splice site recognition. Elife 2015, 4, e04986. [Google Scholar] [CrossRef]
- Dybkov, O.; Will, C.L.; Deckert, J.; Behzadnia, N.; Hartmuth, K.; Lührmann, R. U2 snRNA-protein contacts in purified human 17S U2 snRNPs and in spliceosomal A and B complexes. Mol. Cell. Biol. 2006, 26, 2803–2816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenasi, T.; Barboric, M. P-TEFb stimulates transcription elongation and pre-mRNA splicing through multilateral mechanisms. RNA Biol. 2010, 7, 145–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matera, A.G.; Wang, Z. A day in the life of the spliceosome. Nat. Rev. Mol. Cell Biol. 2014, 15, 108–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyer, A.L.; Osheim, Y.N. Splice site selection, rate of splicing, and alternative splicing on nascent transcripts. Genes Dev. 1988, 2, 754–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, X.; Yan, C.; Zhang, X.; Lei, J.; Shi, Y. Structure of a human catalytic step I spliceosome. Science 2018, 359, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldi, T.; Cortazar, M.A.; Sheridan, R.M.; Bentley, D.L. Coupling of RNA Polymerase II Transcription Elongation with Pre-mRNA Splicing. J. Mol. Biol. 2016, 428, 2623–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matlin, A.J.; Clark, F.; Smith, C.W.J. Understanding alternative splicing: Towards a cellular code. Nat. Rev. Mol. Cell Biol. 2005, 6, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, S.; Zhang, L.; Issaian, A.; Hill, R.C.; Espinosa, S.; Shi, S.; Cui, Y.; Kappel, K.; Das, R.; et al. A unified mechanism for intron and exon definition and back-splicing. Nat. Cell Biol. 2019, 573, 375–380. [Google Scholar] [CrossRef]
- Yan, C.; Wan, R.; Shi, Y. Molecular mechanisms of pre-mRNA splicing through structural biology of the spliceosome. Cold Spring Harb. Perspect. Biol. 2019, 11, a032409. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Burge, C.B. Splicing regulation: From a parts list of regulatory elements to an integrated splicing code. RNA 2008, 14, 802–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S. SR Proteins: Binders, Regulators, and Connectors of RNA. Mol. Cells 2017, 40, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Manley, J.L. Mechanisms of alternative splicing regulation: Insights from molecular and genomics approaches. Nat. Rev. Mol. Cell Biol. 2009, 10, 741–754. [Google Scholar] [CrossRef]
- Akhtar, J.; Kreim, N.; Marini, F.; Mohana, G.; Brüne, D.; Binder, H.; Roignant, J.-Y. Promoter-proximal pausing mediated by the exon junction complex regulates splicing. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.-D.; Ares, M. Context-dependent control of alternative splicing by RNA-binding proteins. Nat. Rev. Genet. 2014, 15, 689–701. [Google Scholar] [CrossRef]
- Dujardin, G.; Lafaille, C.; Petrillo, E.; Buggiano, V.; Acuña, L.I.G.; Fiszbein, A.; Herz, M.A.G.; Moreno, N.N.; Muñoz, M.J.; Alló, M.; et al. Transcriptional elongation and alternative splicing. Biochim. Biophys. Acta (BBA) Bioenergy 2013, 1829, 134–140. [Google Scholar] [CrossRef]
- Koh, C.M.; Bezzi, M.; Low, D.H.; Ang, W.X.; Teo, S.X.; Gay, F.P.; Al-Haddawi, M.; Tan, S.Y.; Osato, M.; Sabò, A.; et al. MYC regulates the core pre-mRNA splicing machinery as an essential step in lymphomagenesis. Nature 2015, 523, 96–100. [Google Scholar] [CrossRef]
- Phillips, J.W.; Pan, Y.; Tsai, B.L.; Xie, Z.; Demirdjian, L.; Xiao, W.; Yang, H.T.; Zhang, Y.; Lin, C.H.; Cheng, D.; et al. Pathway-guided analysis identifies Myc-dependent alternative pre-mRNA splicing in aggressive prostate cancers. Proc. Natl. Acad. Sci. USA 2020, 117, 5269–5279. [Google Scholar] [CrossRef] [Green Version]
- Rauch, J.; Moran-Jones, K.; Albrecht, V.; Schwarzl, T.; Hunter, K.; Gires, O.; Kolch, W. c-Myc Regulates RNA Splicing of the A-Raf Kinase and Its Activation of the ERK Pathway. Cancer Res. 2011, 71, 4664–4674. [Google Scholar] [CrossRef] [Green Version]
- Uppal, S.; Gegonne, A.; Chen, Q.; Thompson, P.S.; Cheng, D.; Mu, J.; Meerzaman, D.; Misra, H.S.; Singer, D.S. The Bromodomain Protein 4 Contributes to the Regulation of Alternative Splicing. Cell Rep. 2019, 29, 2450–2460.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussong, M.; Kaehler, C.; Kerick, M.; Grimm, C.; Franz, A.; Timmermann, B.; Welzel, F.; Isensee, J.; Hucho, T.; Krobitsch, S.; et al. The bromodomain protein BRD4 regulates splicing during heat shock. Nucleic Acids Res. 2017, 45, 382–394. [Google Scholar] [CrossRef] [Green Version]
- Black, D.L. Mechanisms of Alternative Pre-Messenger RNA Splicing. Annu. Rev. Biochem. 2003, 72, 291–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesselberth, J.R. Lives that introns lead after splicing. Wiley Interdiscip. Rev. RNA 2013, 4, 677–691. [Google Scholar] [CrossRef]
- Millevoi, S.; Loulergue, C.; Dettwiler, S.; Karaa, S.Z.; Keller, W.; Antoniou, M.; Vagner, S. An interaction between U2AF 65 and CF I(m) links the splicing and 3’ end processing machineries. EMBO J. 2006, 25, 4854–4864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balbo, P.B.; Bohm, A. Mechanism of poly(A) polymerase: Structure of the enzyme-MgATP-RNA ternary complex and kinetic analysis. Structure 2007, 15, 1117–1131. [Google Scholar] [CrossRef] [Green Version]
- Nilsen, T.W.; Graveley, B.R. Expansion of the eukaryotic proteome by alternative splicing. Nat. Cell Biol. 2010, 463, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Tilgner, H.; Nikolaou, C.; Althammer, S.; Sammeth, M.; Beato, M.; Valcárcel, J.; Guigó, R. Nucleosome positioning as a determinant of exon recognition. Nat. Struct. Mol. Biol. 2009, 16, 996–1001. [Google Scholar] [CrossRef]
- Schwartz, S.; Meshorer, E.; Ast, G. Chromatin organization marks exon-intron structure. Nat. Struct. Mol. Biol. 2009, 16, 990–995. [Google Scholar] [CrossRef]
- Acuña, L.I.G.; Fiszbein, A.; Allo, M.; Schor, I.E.; Kornblihtt, A.R. Connections between chromatin signatures and splicing. Wiley Interdiscip. Rev. RNA 2012, 4, 77–91. [Google Scholar] [CrossRef]
- Yuan, H.; Li, N.; Fu, D.; Ren, J.; Hui, J.; Peng, J.; Liu, Y.; Qiu, T.; Jiang, M.; Pan, Q.; et al. Histone methyltransferase SETD2 modulates alternative splicing to inhibit intestinal tumorigenesis. J. Clin. Investig. 2017, 127, 3375–3391. [Google Scholar] [CrossRef]
- Maor, G.L.; Yearim, A.; Ast, G. The alternative role of DNA methylation in splicing regulation. Trends Genet. 2015, 31, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Khan, D.H.; Gonzalez, C.; Cooper, C.; Sun, J.-M.; Chen, H.Y.; Healy, S.; Xu, W.; Smith, K.T.; Workman, J.L.; Leygue, E.; et al. RNA-dependent dynamic histone acetylation regulates MCL1 alternative splicing. Nucleic Acids Res. 2013, 42, 1656–1670. [Google Scholar] [CrossRef] [Green Version]
- Khan, D.H.; Gonzalez, C.; Tailor, N.; Hamedani, M.K.; Leygue, E.; Davie, J.R. Dynamic Histone Acetylation of H3K4me3 Nucleosome Regulates MCL1 Pre-mRNA Splicing. J. Cell. Physiol. 2016, 231, 2196–2204. [Google Scholar] [CrossRef] [PubMed]
- Luco, R.F.; Pan, Q.; Tominaga, K.; Blencowe, B.J.; Pereira-Smith, O.M.; Misteli, T. Regulation of Alternative Splicing by Histone Modifications. Science 2010, 327, 996–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, J.M.; Hacker, K.E.; Singh, D.; Brannon, A.R.; Parker, J.S.; Weiser, M.; Ho, T.H.; Kuan, P.-F.; Jonasch, E.; Furey, T.S.; et al. Variation in chromatin accessibility in human kidney cancer links H3K36 methyltransferase loss with widespread RNA processing defects. Genome Res. 2014, 24, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Zheng, L.; Park, J.W.; Lv, R.; Chen, H.; Jiao, F.; Xu, W.; Mu, S.; Wen, H.; Qiu, J.; et al. BS69/ZMYND11 reads and connects histone H3.3 lysine 36 trimethylation-decorated chromatin to regu-lated pre-mRNA processing. Mol. Cell 2014, 56, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.; Duerksen, J.D. Chromatin-associated RNA content of heterochromatin and euchromatin. Mol. Cell. Biochem. 1975, 9, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Gelfman, S.; Cohen, N.; Yearim, A.; Ast, G. DNA-methylation effect on cotranscriptional splicing is dependent on GC architecture of the ex-on-intron structure. Genome Res. 2013, 23, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Marina, R.J.; Oberdoerffer, S. Epigenomics meets splicing through the TETs and CTCF. Cell Cycle 2016, 15, 1397–1399. [Google Scholar] [CrossRef] [Green Version]
- Young, J.I.; Hong, E.P.; Castle, J.C.; Crespo-Barreto, J.; Bowman, A.B.; Rose, M.F.; Kang, D.; Richman, R.; Johnson, J.M.; Berget, S.; et al. Regulation of RNA splicing by the methylation-dependent transcriptional repressor methyl-CpG binding protein 2. Proc. Natl. Acad. Sci. USA 2005, 102, 17551–17558. [Google Scholar] [CrossRef] [Green Version]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Legarde, J.; Lin, W.; Schelsinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, E.P. An integrated encyclopedia of DNA elements in the human genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef]
- Dana, H.; Chalbatani, G.M.; Mahmoodzadeh, H.; Karimloo, R.; Rezaiean, O.; Moradzadeh, A.; Mehmandoost, N.; Moazzen, F.; Mazraeh, A.; Marmari, V.; et al. Molecular Mechanisms and Biological Functions of siRNA. Int. J. Biomed. Sci. IJBS 2017, 13, 48–57. [Google Scholar] [PubMed]
- Schroeder, A.; Levins, C.G.; Cortez, C.; Langer, R.; Anderson, D.G. Lipid-based nanotherapeutics for siRNA delivery. J. Intern. Med. 2009, 267, 9–21. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Wen, J.; Huang, Z.; Chen, X.-P.; Zhang, B.-X.; Chu, L. Small Nucleolar RNAs: Insight Into Their Function in Cancer. Front. Oncol. 2019, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Cao, T.; Rajasingh, S.; Samanta, S.; Dawn, B.; Bittel, D.C.; Rajasingh, J. Biology and clinical relevance of noncoding sno/scaRNAs. Trends Cardiovasc. Med. 2018, 28, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Fullwood, M.J. Roles, Functions, and Mechanisms of Long Non-coding RNAs in Cancer. Genom. Proteom. Bioinform. 2016, 14, 42–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skalska, L.; Beltran-Nebot, M.; Ule, J.; Jenner, R.G. Regulatory feedback from nascent RNA to chromatin and transcription. Nat. Rev. Mol. Cell Biol. 2017, 18, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-K.; Shiekhattar, R. Architectural and Functional Commonalities between Enhancers and Promoters. Cell 2015, 162, 948–959. [Google Scholar] [CrossRef] [Green Version]
- Scruggs, B.S.; Adelman, K. The Importance of Controlling Transcription Elongation at Coding and Noncoding RNA Loci. Cold Spring Harb. Symp. Quant. Biol. 2015, 80, 33–44. [Google Scholar] [CrossRef]
- Preker, P.; Nielsen, J.; Kammler, S.; Lykke-Andersen, S.; Christensen, M.S.; Mapendano, C.K.; Schierup, M.H.; Jensen, T.H. RNA Exosome Depletion Reveals Transcription Upstream of Active Human Promoters. Science 2008, 322, 1851–1854. [Google Scholar] [CrossRef] [Green Version]
- Seila, A.C.; Core, L.J.; Lis, J.T.; Sharp, P.A. Divergent transcription: A new feature of active promoters. Cell Cycle 2009, 8, 2557–2564. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Notani, D.; Rosenfeld, W.L.D.N.M.G. Enhancers as non-coding RNA transcription units: Recent insights and future perspectives. Nat. Rev. Genet. 2016, 17, 207–223. [Google Scholar] [CrossRef]
- Li, Y.; Syed, J.; Sugiyama, H. RNA-DNA Triplex Formation by Long Noncoding RNAs. Cell Chem. Biol. 2016, 23, 1325–1333. [Google Scholar] [CrossRef] [Green Version]
- Frank-Kamenetskii, M.D.; Mirkin, S.M. Triplex DNA Structures. Annu. Rev. Biochem. 1995, 64, 65–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Vasquez, K.M. Impact of alternative DNA structures on DNA damage, DNA repair, and genetic instability. DNA Repair 2014, 19, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soibam, B. Super-lncRNAs: Identification of lncRNAs that target super-enhancers via RNA:DNA:DNA triplex for-mation. RNA 2017, 23, 1729–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Giles, K.E.; Felsenfeld, G. DNA.RNA triple helix formation can function as a cis-acting regulatory mecha-nism at the human beta-globin locus. Proc. Natl. Acad. Sci. USA 2019, 116, 6130–6139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlundt, A.; Tants, J.-N.; Sattler, M. Integrated structural biology to unravel molecular mechanisms of protein-RNA recognition. Methods 2017, 118-119, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Campagne, S.; Krepl, M.; Sponer, J.; Allain, F.H.-T. Combining NMR Spectroscopy and Molecular Dynamic Simulations to Solve and Analyze the Structure of Protein–RNA Complexes. Methods Enzymol. 2019, 614, 393–422. [Google Scholar] [CrossRef]
- Michel, S.L.J.; Guerrerio, A.A.L.; Berg, J.M. Selective RNA Binding by a Single CCCH Zinc-Binding Domain from Nup475 (Tristetraprolin). Biochemistry 2003, 42, 4626–4630. [Google Scholar] [CrossRef]
- Brown, R.S. Zinc finger proteins: Getting a grip on RNA. Curr. Opin. Struct. Biol. 2005, 15, 94–98. [Google Scholar] [CrossRef]
- Maris, C.; Dominguez, C.; Allain, F.H.-T. The RNA recognition motif, a plastic RNA-binding platform to regulate post-transcriptional gene expression. FEBS J. 2005, 272, 2118–2131. [Google Scholar] [CrossRef]
- Masliah, G.; Barraud, P.; Allain, F.H. RNA recognition by double-stranded RNA binding domains: A matter of shape and se-quence. Cell. Mol. Life Sci. 2013, 70, 1875–1895. [Google Scholar] [CrossRef] [Green Version]
- Thisted, T.; Lyakhov, D.L.; Liebhaber, S.A. Optimized RNA Targets of Two Closely Related Triple KH Domain Proteins, Heterogeneous Nuclear Ribonucleoprotein K and αCP-2KL, Suggest Distinct Modes of RNA Recognition. J. Biol. Chem. 2001, 276, 17484–17496. [Google Scholar] [CrossRef] [Green Version]
- Nicastro, G.; Taylor, I.A.; Ramos, A. KH–RNA interactions: Back in the groove. Curr. Opin. Struct. Biol. 2015, 30, 63–70. [Google Scholar] [CrossRef]
- Bomsztyk, K.; Denisenko, O.; Ostrowski, J. hnRNP K: One protein multiple processes. BioEssays 2004, 26, 629–638. [Google Scholar] [CrossRef]
- Geuens, T.; Bouhy, D.; Timmerman, V. The hnRNP family: Insights into their role in health and disease. Qual. Life Res. 2016, 135, 851–867. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Qiu, H.; He, J.; Liu, L.; Xue, W.; Fox, A.; Tickner, J.; Xu, J. The emerging roles of hnRNPK. J. Cell. Physiol. 2020, 235, 1995–2008. [Google Scholar] [CrossRef]
- Moumen, A.; Masterson, P.; O’Connor, M.J.; Jackson, S.P. hnRNP K: An HDM2 Target and Transcriptional Coactivator of p53 in Response to DNA Damage. Cell 2005, 123, 1065–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffery, L.; Nakielny, S. Components of the DNA methylation system of chromatin control are RNA-binding proteins. J. Biol. Chem. 2004, 279, 49479–49487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Zhang, H.; Cardoso, M.C. MeCP2 and Chromatin Compartmentalization. Cells 2020, 9, 878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hentze, M.W.; Castello, A.; Schwarzl, T.; Preiss, T. A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell Biol. 2018, 19, 327–341. [Google Scholar] [CrossRef]
- Melikishvili, M.; Chariker, J.H.; Rouchka, E.C.; Fondufe-Mittendorf, Y.N. Transcriptome-wide identification of the RNA-binding landscape of the chromatin-associated protein PARP1 reveals functions in RNA biogenesis. Cell Discov. 2017, 3, 17043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrickson, D.G.; Kelley, D.R.; Tenen, D.; Bernstein, B.; Rinn, J.L. Widespread RNA binding by chromatin-associated proteins. Genome Biol. 2016, 17, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khelifi, G.; Hussein, S.M.I. A New View of Genome Organization through RNA Directed Interactions. Front. Cell Dev. Biol. 2020, 8, 517. [Google Scholar] [CrossRef]
- Xiao, R.; Chen, J.-Y.; Liang, Z.; Luo, D.; Chen, G.; Lu, Z.J.; Chen, Y.; Zhou, B.; Li, H.; Du, X.; et al. Pervasive Chromatin-RNA Binding Protein Interactions Enable RNA-Based Regulation of Transcription. Cell 2019, 178, 107–121.e18. [Google Scholar] [CrossRef] [PubMed]
- Brannan, K.W.; Jin, W.; Huelga, S.C.; Banks, C.A.; Gilmore, J.M.; Florens, L.; Washburn, M.P.; Van Nostrand, E.L.; Pratt, G.A.; Schwinn, M.K.; et al. SONAR Discovers RNA-Binding Proteins from Analysis of Large-Scale Protein-Protein Interactomes. Mol. Cell 2016, 64, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Alarcón, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m6A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Cao, S.; Huang, Q.; Xia, L.; Deng, M.; Yang, M.; Jia, G.; Liu, X.; Shi, J.; Wang, W.; et al. The RNA N6-methyladenosine modification landscape of human fetal tissues. Nat. Cell Biol. 2019, 21, 651–661. [Google Scholar] [CrossRef]
- Tzelepis, K.; Rausch, O.; Kouzarides, T. RNA-modifying enzymes and their function in a chromatin context. Nat. Struct. Mol. Biol. 2019, 26, 858–862. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, C.M.; Batista, P.J. It’s complicated. m(6)A-dependent regulation of gene expression in cancer. Biochim. Biophys. Acta Gene. Regul. Mech. 2019, 1862, 382–393. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Zhou, K.; Wu, T.; Zhao, B.S.; Sun, M.; Chen, Z.; Deng, X.; Xiao, G.; Auer, F.; et al. Histone H3 trimethylation at lysine 36 guides m6A RNA modification co-transcriptionally. Nat. Cell Biol. 2019, 567, 414–419. [Google Scholar] [CrossRef]
- Lee, D.-F.; Walsh, M.J.; Aguiló, F. ZNF217/ZFP217 Meets Chromatin and RNA. Trends Biochem. Sci. 2016, 41, 986–988. [Google Scholar] [CrossRef] [Green Version]
- Aguilo, F.; Zhang, F.; Sancho, A.; Fidalgo, M.; Di Cecilia, S.; Vashisht, A.; Lee, D.-F.; Chen, C.-H.; Rengasamy, M.; Andino, B.; et al. Coordination of m 6 A mRNA Methylation and Gene Transcription by ZFP217 Regulates Pluripotency and Reprogramming. Cell Stem Cell 2015, 17, 689–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xia, L.; Tan, K.; Ye, X.; Zuo, Z.; Li, M.; Xiao, R.; Wang, Z.; Liu, X.; Deng, M.; et al. N6-Methyladenosine co-transcriptionally directs the demethylation of histone H3K9me2. Nat. Genet. 2020, 52, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dou, X.; Chen, C.; Chen, C.; Liu, C.; Xu, M.M.; Zhao, S.; Shen, B.; Gao, Y.; Han, D.; et al. N(6)-methyladenosine of chromosome-associated regulatory RNA regulates chromatin state and transcription. Science 2020, 367, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Kuppers, D.A.; Arora, S.; Lim, Y.; Lim, A.R.; Carter, L.M.; Corrin, P.D.; Plaisier, C.L.; Basom, R.; Delrow, J.J.; Wang, S.; et al. N6-methyladenosine mRNA marking promotes selective translation of regulons required for human erythropoiesis. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertero, A.; Brown, S.; Madrigal, P.; Osnato, A.; Ortmann, D.; Yiangou, L.; Kadiwala, J.; Hubner, N.C.; de Los Mozos, I.R.; Sadée, C.; et al. The SMAD2/3 interactome reveals that TGFbeta controls m(6)A mRNA methylation in pluripotency. Nature 2018, 555, 256–259. [Google Scholar] [CrossRef]
- Xie, Y.; Castro-Hernández, R.; Sokpor, G.; Pham, L.; Narayanan, R.; Rosenbusch, J.; Staiger, J.F.; Tuoc, T. RBM15 Modulates the Function of Chromatin Remodeling Factor BAF155 Through RNA Methylation in Developing Cortex. Mol. Neurobiol. 2019, 56, 7305–7320. [Google Scholar] [CrossRef]
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piątkowski, P.; Bagiński, B.; Wirecki, T.K.; De Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018, 46, D303–D307. [Google Scholar] [CrossRef]
- Schumann, U.; Zhang, H.-N.; Sibbritt, T.; Pan, A.; Horvath, A.; Gross, S.; Clark, S.J.; Yang, L.; Preiss, T. Multiple links between 5-methylcytosine content of mRNA and translation. BMC Biol. 2020, 18, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Aguilo, F.; Li, S.; Balasubramaniyan, N.; Sancho, A.; Benko, S.; Zhang, F.; Vashisht, A.; Rengasamy, M.; Andino, B.; Chen, C.-H.; et al. Deposition of 5-Methylcytosine on Enhancer RNAs Enables the Coactivator Function of PGC-1α. Cell Rep. 2016, 14, 479–492. [Google Scholar] [CrossRef] [Green Version]
- Tillotson, R.; Selfridge, J.; Koerner, M.V.; Gadalla, K.K.E.; Guy, J.; De Sousa, D.; Hector, R.D.; Cobb, S.R.; Bird, A. Radically truncated MeCP2 rescues Rett syndrome-like neurological defects. Nat. Cell Biol. 2017, 550, 398–401. [Google Scholar] [CrossRef]
- Maxwell, S.S.; Pelka, G.J.; Tam, P.P.; El-Osta, A. Chromatin context and ncRNA highlight targets of MeCP2 in brain. RNA Biol. 2013, 10, 1741–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Lee, H.; Han, N.; Kwak, S.; Lee, H.-T.; Kim, J.-H.; Kang, K.; Youn, B.H.; Yang, J.-H.; Jeong, H.-J.; et al. Long non-coding RNA ChRO1 facilitates ATRX/DAXX-dependent H3.3 deposition for transcription-associated heterochromatin reorganization. Nucleic Acids Res. 2018, 46, 11759–11775. [Google Scholar] [CrossRef] [Green Version]
- Fioriniello, S.; Csukony, E.; Marano, D.; Brancaccio, A.; Madonna, M.; Zarrillo, C.; Romano, A.; Marracino, F.; Matarazzo, M.R. MeCP2 and Major Satellite Forward RNA Cooperate for Pericentric Heterochromatin Organization. Stem Cell Rep. 2020, 15, 1317–1332. [Google Scholar] [CrossRef] [PubMed]
- Marnef, A.; Legube, G. m6A RNA modification as a new player in R-loop regulation. Nat. Genet. 2019, 52, 27–28. [Google Scholar] [CrossRef]
- Yang, X.; Liu, Q.-L.; Xu, W.; Zhang, Y.-C.; Yang, Y.; Ju, L.-F.; Chen, J.; Chen, Y.-S.; Li, K.; Ren, J.; et al. m6A promotes R-loop formation to facilitate transcription termination. Cell Res. 2019, 29, 1035–1038. [Google Scholar] [CrossRef]
- Lee, J.-H.; Paull, T.T. Activation and regulation of ATM kinase activity in response to DNA double-strand breaks. Oncogene 2007, 26, 7741–7748. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Chen, L.; Peng, D.; Jiang, A.; He, Y.; Zeng, Y.; Xie, C.; Zhou, H.; Luo, X.; Liu, H.; et al. METTL3 and N6-Methyladenosine Promote Homologous Recombination-Mediated Repair of DSBs by Modulating DNA-RNA Hybrid Accumulation. Mol. Cell 2020, 79, 425–442.e7. [Google Scholar] [CrossRef] [PubMed]
- Abakir, A.; Giles, T.C.; Cristini, A.; Foster, J.M.; Dai, N.; Starczak, M.; Rubio-Roldan, A.; Li, M.; Eleftheriou, M.; Crutchley, J.; et al. N6-methyladenosine regulates the stability of RNA:DNA hybrids in human cells. Nat. Genet. 2019, 52, 48–55. [Google Scholar] [CrossRef]
- Crossley, M.P.; Bocek, M.; Cimprich, K.A. R-Loops as Cellular Regulators and Genomic Threats. Mol. Cell 2019, 73, 398–411. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Wei, J.; He, C. Where, When, and How: Context-Dependent Functions of RNA Methylation Writers, Readers, and Erasers. Mol. Cell 2019, 74, 640–650. [Google Scholar] [CrossRef]
- Teng, Y.; Yadav, T.; Duan, M.; Tan, J.; Xiang, Y.; Gao, B.; Xu, J.; Liang, Z.; Liu, Y.; Nakajima, S.; et al. ROS-induced R loops trigger a transcription-coupled but BRCA1/2-independent homologous recombina-tion pathway through CSB. Nat. Commun. 2018, 9, 4115. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.-T.; Hawley, B.R.; Skalka, G.L.; Baldock, R.A.; Smith, E.M.; Bader, A.S.; Malewicz, M.; Watts, F.Z.; Wilczynska, A.; Bushell, M. Drosha drives the formation of DNA:RNA hybrids around DNA break sites to facilitate DNA repair. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Moon, H.J.; Redman, K.L. Trm4 and Nsun2 RNA:m5C Methyltransferases Form Metabolite-Dependent, Covalent Adducts with Previously Methylated RNA. Biochemistry 2014, 53, 7132–7144. [Google Scholar] [CrossRef] [PubMed]
- Rai, K.; Chidester, S.; Zavala, C.V.; Manos, E.J.; James, S.R.; Karpf, A.R.; Jones, D.A.; Cairns, B.R. Dnmt2 functions in the cytoplasm to promote liver, brain, and retina development in zebrafish. Genes Dev. 2007, 21, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Jurkowski, T.P.; Meusburger, M.; Phalke, S.; Helm, M.; Nellen, W.; Reuter, G.; Jeltsch, A. Human DNMT2 methylates tRNA(Asp) molecules using a DNA methyltransferase-like catalytic mechanism. RNA 2008, 14, 1663–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Yang, H.; Zhu, X.; Yadav, T.; Ouyang, J.; Truesdell, S.S.; Tan, J.; Wang, Y.; Duan, M.; Wei, L.; et al. m5C modification of mRNA serves a DNA damage code to promote homologous recombination. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Xu, L.; Liu, X.; Sheng, N.; Oo, K.S.; Liang, J.; Chionh, Y.H.; Xu, J.; Ye, F.; Gao, Y.-G.; Dedon, P.C.; et al. Three distinct 3-methylcytidine (m3C) methyltransferases modify tRNA and mRNA in mice and humans. J. Biol. Chem. 2017, 292, 14695–14703. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.-H.; Zhang, X.-Y.; Hu, T.; Chen, X.-Y.; Li, J.-J.; Raida, M.; Sun, N.; Luo, Y.; Gao, X. The SUMOylated METTL8 Induces R-loop and Tumorigenesis via m3C. iScience 2020, 23, 100968. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Klungland, A. Modifications and interactions at the R-loop. DNA Repair 2020, 96, 102958. [Google Scholar] [CrossRef] [PubMed]
- Levanon, K.; Eisenberg, E.; Rechavi, G.; Levanon, E.Y.; Rose, S.; Flipse, S.M.; Van Der Sanden, M.C.A.; Radstake, M.; De Winde, J.H.; Osseweijer, P.; et al. Letter from the editor: Adenosine-to-inosine RNA editing in Alu repeats in the human genome. EMBO Rep. 2005, 6, 831–835. [Google Scholar] [CrossRef] [Green Version]
- Nigita, G.; Marceca, G.P.; Tomasello, L.; Distefano, R.; Calore, F.; Veneziano, D.; Romano, G.; Nana-Sinkam, S.P.; Acunzo, M.; Croce, C.M. ncRNA Editing: Functional Characterization and Computational Resources. Breast Cancer 2019, 1912, 133–174. [Google Scholar] [CrossRef]
- Savva, Y.A.; Jepson, J.E.C.; Chang, Y.-J.; Whitaker, R.N.; Jones, B.C.; Laurent, G.S.; Tackett, M.R.; Kapranov, P.; Jiang, N.; Du, G.; et al. RNA editing regulates transposon-mediated heterochromatic gene silencing. Nat. Commun. 2013, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shevchenko, G.; Morris, K.V. All I’s on the RADAR: Role of ADAR in gene regulation. FEBS Lett. 2018, 592, 2860–2873. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.H.; Jansen, R.-P. A jack of all trades: The RNA-binding protein vigilin. Wiley Interdiscip. Rev. RNA 2017, 8, e1448. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Z.; Blackwell, K.; Carmichael, G.G. Vigilins bind to promiscuously A-to-I-edited RNAs and are involved in the formation of heterochromatin. Curr. Biol. 2005, 15, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Rea, S.; Eisenhaber, F.; O’Carroll, D.; Strahl, B.D.; Sun, Z.-W.; Schmid, M.; Opravil, S.; Mechtler, K.; Ponting, C.P.; Allis, C.D.; et al. Regulation of chromatin structure by site-specific histone H3 methyltransferases. Nat. Cell Biol. 2000, 406, 593–599. [Google Scholar] [CrossRef]
- Soldi, M.; Bonaldi, T. The Proteomic Investigation of Chromatin Functional Domains Reveals Novel Synergisms among Distinct Heterochromatin Components. Mol. Cell. Proteom. 2013, 12, 764–780. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, Q.; Chen, L.-L.; Carmichael, G.G. On the mechanism of induction of heterochromatin by the RNA-binding protein vigilin. RNA 2008, 14, 1773–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldstein, B.; Agranat-Tamir, L.; Light, D.; Zgayer, O.B.-N.; Fishman, A.; Lamm, A.T. A-to-I RNA editing promotes developmental stage–specific gene and lncRNA expression. Genome Res. 2017, 27, 462–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, V.; Davidson, N.O. C-to-U RNA Editing: Mechanisms Leading to Genetic Diversity. J. Biol. Chem. 2003, 278, 1395–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazanov, M.D.; Roberts, S.A.; Polak, P.; Stamatoyannopoulos, J.; Klimczak, L.J.; Gordenin, D.A.; Sunyaev, S.R. APOBEC-Induced Cancer Mutations Are Uniquely Enriched in Early-Replicating, Gene-Dense, and Active Chromatin Regions. Cell Rep. 2015, 13, 1103–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periyasamy, M.; Patel, H.; Lai, C.-F.; Nguyen, V.T.; Nevedomskaya, E.; Harrod, A.; Russell, R.; Remenyi, J.; Ochocka, A.M.; Thomas, R.S.; et al. APOBEC3B-Mediated Cytidine Deamination Is Required for Estrogen Receptor Action in Breast Cancer. Cell Rep. 2015, 13, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The m6A epitranscriptome: Transcriptome plasticity in brain development and function. Nat. Rev. Neurosci. 2020, 21, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Widagdo, J.; Anggono, V. The m6A-epitranscriptomic signature in neurobiology: From neurodevelopment to brain plasticity. J. Neurochem. 2018, 147, 137–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.W.; Lee, S.M.; Yoon, K.J. Epitranscriptomic regulation of transcriptome plasticity in development and dis-eases of the brain. BMB Rep. 2020, 53, 551–564. [Google Scholar] [CrossRef]
- Vissers, C.; Sinha, A.; Ming, G.-L.; Song, H. The epitranscriptome in stem cell biology and neural development. Neurobiol. Dis. 2020, 146, 105139. [Google Scholar] [CrossRef]
- Yoon, K.-J.; Ringeling, F.R.; Vissers, C.; Jacob, F.; Pokrass, M.; Jimenez-Cyrus, D.; Su, Y.; Kim, N.-S.; Zhu, Y.; Zheng, L.; et al. Temporal Control of Mammalian Cortical Neurogenesis by m6A Methylation. Cell 2017, 171, 877–889.e17. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-X.; Cui, G.-S.; Liu, X.; Xu, K.; Wang, M.; Zhang, X.-X.; Jiang, L.-Y.; Li, A.; Yang, Y.; Lai, W.-Y.; et al. METTL3-mediated m6A modification is required for cerebellar development. PLoS Biol. 2018, 16, e2004880. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Yue, M.; Wang, J.; Kumar, S.; Wechsler-Reya, R.J.; Zhang, Z.; Ogawa, Y.; Kellis, M.; Duester, G.; et al. N6-methyladenosine RNA modification regulates embryonic neural stem cell self-renewal through histone modifications. Nat. Neurosci. 2018, 21, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, Y.-C.; Huang, C.; Shen, H.; Sun, B.; Cheng, X.; Zhang, Y.-J.; Yang, Y.-G.; Shu, Q.; Yang, Y.; et al. m6A Regulates Neurogenesis and Neuronal Development by Modulating Histone Methyltransferase Ezh2. Genom. Proteom. Bioinform. 2019, 17, 154–168. [Google Scholar] [CrossRef]
- Weng, Y.-L.; Wang, X.; An, R.; Cassin, J.; Vissers, C.; Liu, Y.; Liu, Y.; Xu, T.; Wang, X.; Wong, S.Z.H.; et al. Epitranscriptomic m6A Regulation of Axon Regeneration in the Adult Mammalian Nervous System. Neuron 2018, 97, 313–325.e6. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Zhuang, Y.; Chen, J.; Xu, W.; Shou, Y.; Huang, X.; Shu, Q.; Li, X. Dynamic effects of Fto in regulating the proliferation and differentiation of adult neural stem cells of mice. Hum. Mol. Genet. 2020, 29, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zang, L.; Zhang, F.; Chen, J.; Shen, H.; Shu, L.; Liang, F.; Feng, C.; Chen, D.; Tao, H.; et al. Fat mass and obesity-associated (FTO) protein regulates adult neurogenesis. Hum. Mol. Genet. 2017, 26, 2398–2411. [Google Scholar] [CrossRef]
- Engel, M.; Eggert, C.; Kaplick, P.M.; Eder, M.; Röh, S.; Tietze, L.; Namendorf, C.; Arloth, J.; Weber, P.; Rex-Haffner, M.; et al. The Role of m6A/m-RNA Methylation in Stress Response Regulation. Neuron 2018, 99, 389–403.e9. [Google Scholar] [CrossRef] [PubMed]
- Walters, B.J.; Mercaldo, V.; Gillon, C.J.; Yip, M.; Neve, R.L.; Boyce, F.M.; Frankland, P.W.; Josselyn, S.A. The Role of the RNA Demethylase FTO (Fat Mass and Obesity-Associated) and mRNA Methylation in Hippocampal Memory Formation. Neuropsychopharmacology 2017, 42, 1502–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Zhang, X.; Weng, Y.-L.; Lu, Z.; Liu, Y.; Lu, Z.; Li, J.; Hao, P.; Zhang, Y.; Zhang, F.; et al. m6A facilitates hippocampus-dependent learning and memory through YTHDF1. Nat. Cell Biol. 2018, 563, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ren, Y.; Mao, K.; Hua, F.; Yang, Y.; Wei, N.; Yue, C.; Li, D.; Zhang, H. FTO is involved in Alzheimer’s disease by targeting TSC1-mTOR-Tau signaling. Biochem. Biophys. Res. Commun. 2018, 498, 234–239. [Google Scholar] [CrossRef]
- Chen, X.; Yu, C.; Guo, M.; Zheng, X.; Ali, S.; Huang, H.; Zhang, L.; Wang, S.; Huang, Y.; Qie, S.; et al. Down-Regulation of m6A mRNA Methylation Is Involved in Dopaminergic Neuronal Death. ACS Chem. Neurosci. 2019, 10, 2355–2363. [Google Scholar] [CrossRef]
- Annapoorna, P.; Iyer, H.; Parnaik, T.; Narasimhan, H.; Bhattacharya, A.; Kumar, A. FTO: An Emerging Molecular Player in Neuropsychiatric Diseases. Neuroscience 2019, 418, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Maas, S.; Single, F.N.; Hartner, J.; Rozov, A.; Burnashev, N.; Feldmeyer, D.; Sprengel, R.; Seeburg, P.H. Point mutation in an AMPA receptor gene rescues lethality in mice deficient in the RNA-editing enzyme ADAR2. Nature 2000, 406, 78–81. [Google Scholar] [CrossRef]
- Rosenthal, J.J.; Seeburg, P.H. A-to-I RNA Editing: Effects on Proteins Key to Neural Excitability. Neuron 2012, 74, 432–439. [Google Scholar] [CrossRef] [Green Version]
- Behm, M.; Öhman, M. RNA Editing: A Contributor to Neuronal Dynamics in the Mammalian Brain. Trends Genet. 2016, 32, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Hwang, T.; Park, C.-K.; Leung, A.K.L.; Gao, Y.; Hyde, T.M.; Kleinman, J.E.; Rajpurohit, A.; Tao, R.; Shin, J.H.; Weinberger, D.R. Dynamic regulation of RNA editing in human brain development and disease. Nat. Neurosci. 2016, 19, 1093–1099. [Google Scholar] [CrossRef] [PubMed]
- Kankowski, S.; Förstera, B.; Winkelmann, A.; Knauff, P.; Wanker, E.E.; You, X.A.; Semtner, M.; Hetsch, F.; Meier, J.C. A Novel RNA Editing Sensor Tool and a Specific Agonist Determine Neuronal Protein Expression of RNA-Edited Glycine Receptors and Identify a Genomic APOBEC1 Dimorphism as a New Genetic Risk Factor of Epilepsy. Front. Mol. Neurosci. 2017, 10, 439. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.N.; Xu, B.S.; Liu, Y.P.; Zhou, M.; Long, T.; Li, H.; Dong, H.; Nie, Y.; Chen, P.R.; et al. Intellectual disability-associated gene ftsj1 is responsible for 2’-O-methylation of specific tRNAs. EMBO Rep. 2020, 21, e50095. [Google Scholar] [CrossRef]
- Jensen, L.R.; Garrett, L.; Hölter, S.M.; Rathkolb, B.; Rácz, I.; Adler, T.; Prehn, C.; Hans, W.; Rozman, J.; Becker, L.; et al. A mouse model for intellectual disability caused by mutations in the X-linked 2′-O-methyltransferase Ftsj1 gene. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2019, 1865, 2083–2093. [Google Scholar] [CrossRef] [PubMed]
- Freude, K.; Hoffmann, K.; Jensen, L.-R.; Delatycki, M.B.; Des Portes, V.; Moser, B.; Hamel, B.; Van Bokhoven, H.; Moraine, C.; Fryns, J.-P.; et al. Mutations in the FTSJ1 gene coding for a novel S-adenosylmethionine-binding protein cause nonsyn-dromic X-linked mental retardation. Am. J. Hum. Genet. 2004, 75, 305–309. [Google Scholar] [CrossRef]
- Lee, Y.-L.; Kung, F.-C.; Lin, C.-H.; Huang, Y.-S. CMTR1-Catalyzed 2′-O-Ribose Methylation Controls Neuronal Development by Regulating Camk2α Expression Independent of RIG-I Signaling. Cell Rep. 2020, 33, 108269. [Google Scholar] [CrossRef]
- Chi, L.; Delgado-Olguin, P. Expression of NOL1/NOP2/sun domain (Nsun) RNA methyltransferase family genes in early mouse embryogenesis. Gene. Expr. Patterns 2013, 13, 319–327. [Google Scholar] [CrossRef]
- Flores, J.V.; Cordero-Espinoza, L.; Oeztuerk-Winder, F.; Andersson-Rolf, A.; Selmi, T.; Blanco, S.; Tailor, J.; Dietmann, S.; Frye, M. Cytosine-5 RNA Methylation Regulates Neural Stem Cell Differentiation and Motility. Stem Cell Rep. 2017, 8, 112–124. [Google Scholar] [CrossRef] [Green Version]
- Blanco, S.; Dietmann, S.; Flores, J.V.; Hussain, S.; Kutter, C.; Humphreys, P.; Lukk, M.; Lombard, P.; Treps, L.; Popis, M.; et al. Aberrant methylation of tRNAs links cellular stress to neuro-developmental disorders. EMBO J. 2014, 33, 2020–2039. [Google Scholar] [CrossRef]
- Abbasi-Moheb, L.; Mertel, S.; Gonsior, M.; Nouri-Vahid, L.; Kahrizi, K.; Cirak, S.; Wieczorek, D.; Motazacker, M.M.; Esmaeeli-Nieh, S.; Cremer, K.; et al. Mutations in NSUN2 Cause Autosomal- Recessive Intellectual Disability. Am. J. Hum. Genet. 2012, 90, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.A.; Rafiq, M.A.; Noor, A.; Hussain, S.; Flores, J.V.; Rupp, V.; Vincent, A.K.; Malli, R.; Ali, G.; Khan, F.S.; et al. Mutation in NSUN2, which encodes an RNA methyltransferase, causes autosomal-recessive intellectual disability. Am. J. Hum. Genet. 2012, 90, 856–863. [Google Scholar] [CrossRef] [Green Version]
- Doan, R.N.; Lim, E.T.; De Rubeis, S.; Betancur, C.; Cutler, D.J.; Chiocchetti, A.G.; Overman, L.M.; Soucy, A.; Goetze, S.; Freitag, C.M.; et al. Recessive gene disruptions in autism spectrum disorder. Nat. Genet. 2019, 51, 1092–1098. [Google Scholar] [CrossRef]
- Innes, A.M.; McInnes, B.L.; Dyment, D.A. Clinical and genetic heterogeneity in Dubowitz syndrome: Implications for diagnosis, management and further research. Am. J. Med. Genet. Part C Semin. Med. Genet. 2018, 178, 387–397. [Google Scholar] [CrossRef] [PubMed]
- De Jesus, D.F.; Zhang, Z.; Kahraman, S.; Brown, N.K.; Chen, M.; Hu, J.; Gupta, M.K.; He, C.; Kulkarni, R.N. m(6)A mRNA Methylation Regulates Human beta-Cell Biology in Physiological States and in Type 2 Diabetes. Nat. Metab. 2019, 1, 765–774. [Google Scholar] [CrossRef]
- Liu, J.; Luo, G.; Sun, J.; Men, L.; Ye, H.; He, C.; Ren, D. METTL14 is essential for beta-cell survival and insulin secretion. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 2138–2148. [Google Scholar] [CrossRef] [PubMed]
- Men, L.; Sun, J.; Luo, G.; Ren, D. Acute Deletion of METTL14 in beta-Cells of Adult Mice Results in Glucose Intolerance. Endocrinology 2019, 160, 2388–2394. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Ohsugi, M.; Sasako, T.; Awazawa, M.; Umehara, T.; Iwane, A.; Kobayashi, N.; Okazaki, Y.; Kubota, N.; Suzuki, R.; et al. The RNA Methyltransferase Complex of WTAP, METTL3, and METTL14 Regulates Mitotic Clonal Expansion in Adipogenesis. Mol. Cell. Biol. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Zhao, Y.; Wu, R.; Jiang, Q.; Cai, M.; Bi, Z.; Liu, Y.; Yao, Y.; Feng, J.; Wang, Y.; et al. ZFP217 regulates adipogenesis by controlling mitotic clonal expansion in a METTL3-m6A dependent manner. RNA Biol. 2019, 16, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Koch, L.; Emmerling, C.; Vierkotten, J.; Peters, T.; Brüning, J.C.; Rüther, U. Inactivation of the Fto gene protects from obesity. Nat. Cell Biol. 2009, 458, 894–898. [Google Scholar] [CrossRef]
- Smemo, S.; Tena, J.J.; Kim, K.-H.; Gamazon, E.R.; Sakabe, N.J.; Gómez-Marín, C.; Aneas, I.; Credidio, F.L.; Sobreira, D.R.; Wasserman, N.F.; et al. Obesity-associated variants within FTO form long-range functional connections with IRX3. Nat. Cell Biol. 2014, 507, 371–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merkestein, M.; Laber, S.; McMurray, F.; Andrew, D.; Sachse, G.; Sanderson, J.; Li, M.; Usher, S.; Sellayah, D.; Ashcroft, F.M.; et al. FTO influences adipogenesis by regulating mitotic clonal expansion. Nat. Commun. 2015, 6, 6792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Luo, Y.; Jia, G.; Liu, G.; Zhao, H.; Huang, Z. FTO Promotes Adipogenesis through Inhibition of the Wnt/beta-catenin Signaling Pathway in Porcine Intramuscular Preadipocytes. Anim. Biotechnol. 2017, 28, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, R.; Liu, Y.; Zhao, Y.; Bi, Z.; Yao, Y.; Liu, Q.; Shi, H.; Wang, F.; Wang, Y. m6A mRNA methylation controls autophagy and adipogenesis by targeting Atg5 and Atg7. Autophagy 2019, 16, 1221–1235. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, J.; Huang, C.; Shen, M.; Zhan, H.; Xu, K. RNA N6-methyladenosine: A promising molecular target in metabolic diseases. Cell Biosci. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shulman, Z.; Stern-Ginossar, N. The RNA modification N6-methyladenosine as a novel regulator of the immune system. Nat. Immunol. 2020, 21, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.D.; Gokhale, N.S.; Horner, S.M. Regulation of Viral Infection by the RNA ModificationN6-Methyladenosine. Annu. Rev. Virol. 2019, 6, 235–253. [Google Scholar] [CrossRef]
- Wu, J.; Chen, Z.J. Innate Immune Sensing and Signaling of Cytosolic Nucleic Acids. Annu. Rev. Immunol. 2014, 32, 461–488. [Google Scholar] [CrossRef]
- Roers, A.; Hiller, B.; Hornung, V. Recognition of Endogenous Nucleic Acids by the Innate Immune System. Immunity 2016, 44, 739–754. [Google Scholar] [CrossRef] [Green Version]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of RNA Recognition by Toll-like Receptors: The Impact of Nucleoside Modification and the Evolutionary Origin of RNA. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Hu, X.; Huang, M.; Liu, J.; Gu, Y.; Ma, L.; Zhou, Q.; Cao, X. Mettl3-mediated mRNA m6A methylation promotes dendritic cell activation. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-B.; Tong, J.; Zhu, S.; Batista, P.J.; Duffy, E.E.; Zhao, J.; Bailis, W.; Cao, G.; Kroehling, L.; Chen, Y.; et al. m(6)A mRNA methylation controls T cell homeostasis by targeting the IL-7/STAT5/SOCS pathways. Nature 2017, 548, 338–342. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Cao, G.; Zhang, T.; Sefik, E.; Vesely, M.C.A.; Broughton, J.P.; Zhu, S.; Li, H.-B.; Li, B.; Chen, L.; et al. m6A mRNA methylation sustains Treg suppressive functions. Cell Res. 2018, 28, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Winkler, R.; Gillis, E.; Lasman, L.; Safra, M.; Geula, S.; Soyris, C.; Nachshon, A.; Tai-Schmiedel, J.; Friedman, N.; Le-Trilling, V.T.K.; et al. m6A modification controls the innate immune response to infection by targeting type I interferons. Nat. Immunol. 2019, 20, 173–182. [Google Scholar] [CrossRef]
- Rubio, R.M.; Depledge, D.P.; Bianco, C.; Thompson, L.; Mohr, I. RNA m(6) A modification enzymes shape innate responses to DNA by regulating interferon beta. Genes Dev. 2018, 32, 1472–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichinchi, G.; Gao, S.; Saletore, Y.; Gonzalez, G.M.; Bansal, V.; Wang, Y.; Mason, C.E.; Rana, T.M. Dynamics of the human and viral m6A RNA methylomes during HIV-1 infection of T cells. Nat. Microbiol. 2016, 1, 1–9. [Google Scholar] [CrossRef]
- Nakahama, T.; Kawahara, Y. Adenosine-to-inosine RNA editing in the immune system: Friend or foe? Cell. Mol. Life Sci. 2020, 77, 2931–2948. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.; Calis, J.J.; Wu, X.; Sun, T.; Yu, Y.; Sarbanes, S.L.; Thi, V.L.D.; Shilvock, A.R.; Hoffmann, H.-H.; Rosenberg, B.R.; et al. Human ADAR1 Prevents Endogenous RNA from Triggering Translational Shutdown. Cell 2018, 172, 811–824.e14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, G.I.; Kasher, P.R.; Forte, G.M.A.; Mannion, N.M.; Greenwood, S.M.; Szynkiewicz, M.; Dickerson, J.E.; Bhaskar, S.S.; Zampini, M.; Briggs, T.A.; et al. Mutations in ADAR1 cause Aicardi-Goutières syndrome associated with a type I interferon signature. Nat. Genet. 2012, 44, 1243–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalinger, Z.B.; Elliott, R.; Rose, K.M.; Weiss, S.R. MDA5 Is Critical to Host Defense during Infection with Murine Coronavirus. J. Virol. 2015, 89, 12330–12340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichlmair, A.; Lassnig, C.; Eberle, C.-A.; Górna, M.W.; Baumann, C.L.; Burkard, T.R.; Bürckstümmer, T.; Stefanovic, A.; Krieger, S.; Bennett, K.L.; et al. IFIT1 is an antiviral protein that recognizes 5′-triphosphate RNA. Nat. Immunol. 2011, 12, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Sweeney, T.R.; Skabkin, M.A.; Skabkina, O.V.; Hellen, C.U.T.; Pestova, T.V. Inhibition of translation by IFIT family members is determined by their ability to interact selectively with the 5′-terminal regions of cap0-, cap1- and 5′ppp- mRNAs. Nucleic Acids Res. 2014, 42, 3228–3245. [Google Scholar] [CrossRef] [Green Version]
- Habjan, M.; Hubel, P.; Lacerda, L.; Benda, C.; Holze, C.; Eberl, C.H.; Mann, A.; Kindler, E.; Gil-Cruz, C.; Ziebuhr, J.; et al. Sequestration by IFIT1 Impairs Translation of 2′O-unmethylated Capped RNA. PLOS Pathog. 2013, 9, e1003663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, Y.M.; Laudenbach, B.T.; Martínez-Montero, S.; Cencic, R.; Habjan, M.; Pichlmair, A.; Damha, M.J.; Pelletier, J.; Nagar, B. Structure of human IFIT1 with capped RNA reveals adaptable mRNA binding and mechanisms for sensing N1 and N2 ribose 2′-O methylations. Proc. Natl. Acad. Sci. USA 2017, 114, E2106–E2115. [Google Scholar] [CrossRef] [Green Version]
- Viswanathan, T.; Arya, S.; Chan, S.-H.; Qi, S.; Dai, N.; Misra, A.; Park, J.-G.; Oladunni, F.; Kovalskyy, D.; Hromas, R.A.; et al. Structural basis of RNA cap modification by SARS-CoV-2. Nat. Commun. 2020, 11, 3718. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Hong, Y.; Yang, B.; Shrestha, P.; Sajjad, N.; Chen, J.-L. Induction of the Antiviral Immune Response and Its Circumvention by Coronaviruses. Viruses 2020, 12, 1039. [Google Scholar] [CrossRef]
- Barbieri, I.; Kouzarides, T. Role of RNA modifications in cancer. Nat. Rev. Cancer 2020, 20, 303–322. [Google Scholar] [CrossRef]
- Haruehanroengra, P.; Zheng, Y.Y.; Zhou, Y.; Huang, Y.; Sheng, J. RNA modifications and cancer. RNA Biol. 2020, 17, 1560–1575. [Google Scholar] [CrossRef]
- Wood, S.; Willbanks, A.; Cheng, J.X. The Role of RNA Modifications and RNA-modifying Proteins in Cancer Thera-py and Drug Resistance. Curr. Cancer Drug Targets 2021. [Google Scholar] [CrossRef]
- Vu, L.P.; Pickering, B.F.; Cheng, Y.; Zaccara, S.; Nguyen, D.; Minuesa, G.; Chou, T.; Chow, A.; Saletore, Y.; Mackay, M.; et al. The N6-methyladenosine (m6A)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells. Nat. Med. 2017, 23, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, I.; Tzelepis, K.; Pandolfini, L.; Namshik, H.; Millán-Zambrano, G.; Robson, S.C.; Aspris, D.; Migliori, V.; Bannister, A.J.; Hannes, P.; et al. Promoter-bound METTL3 maintains myeloid leukaemia by m6A-dependent translation control. Nat. Cell Biol. 2017, 552, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, C.R.; Lee, H.; Goodarzi, H.; Halberg, N.; Tavazoie, S.F. N6-methyladenosine marks primary microRNAs for processing. Nat. Cell Biol. 2015, 519, 482–485. [Google Scholar] [CrossRef]
- Han, J.; Wang, J.-z.; Yang, X.; Yu, H.; Zhou, R.; Lu, H.-C.; Yuan, W.-B.; Lu, J.-c.; Zhou, Z.-j.; Lu, Q.; et al. METTL3 promote tumor proliferation of bladder cancer by accelerating pri-miR221/222 maturation in m6A-dependent manner. Mol. Cancer 2019, 18, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Deng, Q.; Lv, Z.; Ling, Y.; Hou, X.; Chen, Z.; Dinglin, X.; Ma, S.; Li, D.; Wu, Y.; et al. N6-methyladenosine induced miR-143-3p promotes the brain metastasis of lung cancer via regulation of VASH1. Mol. Cancer 2019, 18, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.; Wang, X.; Cao, C.; Gao, Y.; Zhang, S.; Yang, Z.; Liu, Y.; Zhang, X.; Zhang, W.; Ye, L. HBXIP-elevated methyltransferase METTL3 promotes the progression of breast cancer via inhibiting tu-mor suppressor let-7g. Cancer Lett. 2018, 415, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Yue, B.; Song, C.; Yang, L.; Cui, R.; Cheng, X.; Zhang, Z.; Zhao, G. METTL3-mediated N6-methyladenosine modification is critical for epithelial-mesenchymal transition and metastasis of gastric cancer. Mol. Cancer 2019, 18, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Xu, M.; Xu, X.; Zeng, K.; Liu, X.; Sun, L.; Pan, B.; He, B.; Pan, Y.; Sun, H.; et al. METTL14 Suppresses CRC Progression via Regulating N6-Methyladenosine-Dependent Primary miR-375 Processing. Mol. Ther. 2020, 28, 599–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Q.; Shi, H.; Ye, P.; Li, L.; Qu, Q.; Sun, G.; Sun, G.; Lu, Z.; Huang, Y.; Yang, C.-G.; et al. m 6 A RNA Methylation Regulates the Self-Renewal and Tumorigenesis of Glioblastoma Stem Cells. Cell Rep. 2017, 18, 2622–2634. [Google Scholar] [CrossRef]
- Gu, C.; Wang, Z.; Zhou, N.; Li, G.; Kou, Y.; Luo, Y.; Wang, Y.; Yang, J.; Tian, F. Mettl14 inhibits bladder TIC self-renewal and bladder tumorigenesis through N6-methyladenosine of Notch1. Mol. Cancer 2019, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Eckert, M.A.; Harada, B.T.; Liu, S.-M.; Lu, Z.; Yu, K.; Tienda, S.M.; Chryplewicz, A.; Zhu, A.C.; Yang, Y.; et al. m(6)A mRNA methylation regulates AKT activity to promote the proliferation and tumorigenicity of en-dometrial cancer. Nat. Cell. Biol. 2018, 20, 1074–1083. [Google Scholar] [CrossRef]
- Gong, D.; Zhang, J.; Chen, Y.; Xu, Y.; Ma, J.; Hu, G.; Huang, Y.; Zheng, J.; Zhai, W.; Xue, W. The m(6)A-suppressed P2RX6 activation promotes renal cancer cells migration and invasion through ATP-induced Ca(2+) influx modulating ERK1/2 phosphorylation and MMP9 signaling pathway. J. Exp. Clin. Cancer Res. 2019, 38, 233. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, S.; He, C.; Xue, P.; Zhang, L.; He, Z.; Zang, L.; Feng, B.; Sun, J.; Zheng, M. METTL14 suppresses proliferation and metastasis of colorectal cancer by down-regulating oncogenic long non-coding RNA XIST. Mol. Cancer 2020, 19, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Yang, F.; Zhou, C.; Liu, F.; Yuan, J.; Wang, F.; Wang, T.; Xu, Q.; Zhou, W.; Sun, S. METTL14 suppresses the metastatic potential of hepatocellular carcinoma by modulating N 6-methyladenosine-dependent primary MicroRNA processing. Hepatology 2017, 65, 529–543. [Google Scholar] [CrossRef]
- Sun, C.; Chang, L.; Liu, C.; Chen, X.; Zhu, X. The study of METTL3 and METTL14 expressions in childhood ETV6/RUNX1-positive acute lymphoblastic leukemia. Mol. Genet. Genom. Med. 2019, 7, e00933. [Google Scholar] [CrossRef] [Green Version]
- Weng, H.; Huang, H.; Wu, H.; Qin, X.; Zhao, B.S.; Dong, L.; Shi, H.; Skibbe, J.; Shen, C.; Hu, C.; et al. METTL14 Inhibits Hematopoietic Stem/Progenitor Differentiation and Promotes Leukemogenesis via mRNA m6A Modification. Cell Stem Cell 2018, 22, 191–205.e9. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, Y.; Konno, M.; Asai, A.; Koseki, J.; Kawamoto, K.; Miyoshi, N.; Takahashi, H.; Nishida, N.; Haraguchi, N.; Sakai, D.; et al. Oncogene c-Myc promotes epitranscriptome m6A reader YTHDF1 expression in colorectal cancer. Oncotarget 2018, 9, 7476–7486. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Yang, C.; Wu, R.; Huang, L.; Song, S.; Li, W.; Yan, P.; Lin, C.; Li, D.; Zhang, Y. YTHDF1 Regulates Tumorigenicity and Cancer Stem Cell-Like Activity in Human Colorectal Carcinoma. Front. Oncol. 2019, 9, 332. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Wei, Q.; Jin, J.; Luo, Q.; Liu, Y.; Yang, Y.; Cheng, C.; Li, L.; Pi, J.; Si, Y.; et al. The m6A reader YTHDF1 promotes ovarian cancer progression via augmenting EIF3C translation. Nucleic Acids Res. 2020, 48, 3816–3831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.; Liu, J.; Chen, C.; Dong, L.; Liu, Y.; Chang, R.; Huang, X.; Liu, Y.; Wang, J.; Dougherty, U.; et al. Anti-tumour immunity controlled through mRNA m6A methylation and YTHDF1 in dendritic cells. Nat. Cell Biol. 2019, 566, 270–274. [Google Scholar] [CrossRef]
- Chen, J.; Sun, Y.; Xu, X.; Wang, D.; He, J.; Zhou, H.; Lu, Y.; Zeng, J.; Du, F.; Gong, A.; et al. YTH domain family 2 orchestrates epithelial-mesenchymal transition/proliferation dichotomy in pan-creatic cancer cells. Cell Cycle 2017, 16, 2259–2271. [Google Scholar] [CrossRef] [Green Version]
- Zanconato, F.; Cordenonsi, M.; Piccolo, S. YAP and TAZ: A signalling hub of the tumour microenvironment. Nat. Rev. Cancer 2019, 19, 454–464. [Google Scholar] [CrossRef]
- Yan, J.; Huang, X.; Zhang, X.; Chen, Z.; Ye, C.; Xiang, W.; Huang, Z. LncRNA LINC00470 promotes the degradation of PTEN mRNA to facilitate malignant behavior in gastric cancer cells. Biochem. Biophys. Res. Commun. 2020, 521, 887–893. [Google Scholar] [CrossRef]
- Li, F.; Yi, Y.; Miao, Y.; Long, W.; Long, T.; Chen, S.; Cheng, W.; Zou, C.; Zheng, Y.; Wu, X.; et al. N6-Methyladenosine Modulates Nonsense-Mediated mRNA Decay in Human Glioblastoma. Cancer Res. 2019, 79, 5785–5798. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, A.; Konno, J.; Tanikawa, K.; Sahara, H. Transcriptional machinery of TNF-α-inducible YTH domain containing 2 (YTHDC2) gene. Gene 2014, 535, 24–32. [Google Scholar] [CrossRef]
- Tanabe, A.; Tanikawa, K.; Tsunetomi, M.; Takai, K.; Ikeda, H.; Konno, J.; Torigoe, T.; Maeda, H.; Kutomi, G.; Okita, K.; et al. RNA helicase YTHDC2 promotes cancer metastasis via the enhancement of the efficiency by which HIF-1α mRNA is translated. Cancer Lett. 2016, 376, 34–42. [Google Scholar] [CrossRef]
- Jia, R.; Chai, P.; Wang, S.; Sun, B.; Xu, Y.; Yang, Y.; Ge, S.; Jia, R.; Yang, Y.-G.; Fan, X. m(6)A modification suppresses ocular melanoma through modulating HINT2 mRNA translation. Mol. Cancer 2019, 18, 161. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Liao, D.; Zhang, M.; Zeng, C.; Li, X.; Zhang, R.; Ma, H.; Kang, T. YTHDF2 suppresses cell proliferation and growth via destabilizing the EGFR mRNA in hepatocellular carcinoma. Cancer Lett. 2019, 442, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, R.; Zhang, L.; Li, J.; Lou, K.; Shi, B. The lipid metabolism gene FTO influences breast cancer cell energy metabolism via the PI3K/AKT signal-ing pathway. Oncol. Lett. 2017, 13, 4685–4690. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Lin, Z.; Wan, A.; Chen, H.; Liang, H.; Sun, L.; Wang, Y.; Li, X.; Xiong, X.-F.; Wei, B.; et al. RNA N6-methyladenosine demethylase FTO promotes breast tumor progression through inhibiting BNIP3. Mol. Cancer 2019, 18, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, C.; Zhuang, C.; Luo, X.; Huang, X.; Yao, L.; Li, J.; Li, Y.; Xiong, T.; Ye, J.; Zhang, F.; et al. N6-methyladenosine demethylase FTO suppresses clear cell renal cell carcinoma through a novel FTO-PGC-1alpha signalling axis. J. Cell. Mol. Med. 2019, 23, 2163–2173. [Google Scholar] [CrossRef]
- Du, X.; Shao, Y.; Gao, H.; Zhang, X.; Zhang, H.; Ban, Y.; Qin, H.; Tai, Y. CMTR1-ALK: An ALK fusion in a patient with no response to ALK inhibitor crizotinib. Cancer Biol. Ther. 2018, 19, 962–966. [Google Scholar] [CrossRef] [PubMed]
- Falaleeva, M.; Pages, A.; Matuszek, Z.; Hidmi, S.; Agranat-Tamir, L.; Korotkov, K.; Nevo, Y.; Eyras, E.; Sperling, R.; Stamm, S. Dual function of C/D box small nucleolar RNAs in rRNA modification and alternative pre-mRNA splicing. Proc. Natl. Acad. Sci. USA 2016, 113, E1625–E1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcel, V.; Ghayad, S.E.; Belin, S.; Therizols, G.; Morel, A.-P.; Solano-Gonzàlez, E.; Vendrell, J.A.; Hacot, S.; Mertani, H.C.; Albaret, M.A.; et al. p53 acts as a safeguard of translational control by regulating fibrillarin and rRNA methylation in cancer. Cancer Cell 2013, 24, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Qin, W.; Lu, S.; Wang, X.; Zhang, J.; Sun, T.; Hu, X.; Li, Y.; Chen, Q.; Wang, Y.; et al. Long noncoding RNA ZFAS1 promoting small nucleolar RNA-mediated 2’-O-methylation via NOP58 re-cruitment in colorectal cancer. Mol. Cancer 2020, 19, 95. [Google Scholar] [CrossRef] [PubMed]
- Pauli, C.; Liu, Y.; Rohde, C.; Cui, C.; Fijalkowska, D.; Gerloff, D.; Walter, C.; Krijgsveld, J.; Dugas, M.; Edemir, B.; et al. Site-specific methylation of 18S ribosomal RNA by SNORD42A is required for acute myeloid leukemia cell proliferation. Blood 2020, 135, 2059–2070. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Liu, Y.; Rohde, C.; Pauli, C.; Gerloff, D.; Köhn, M.; Misiak, D.; Bäumer, N.; Cui, C.; Göllner, S.; et al. AML1-ETO requires enhanced C/D box snoRNA/RNP formation to induce self-renewal and leukaemia. Nat. Cell Biol. 2017, 19, 844–855. [Google Scholar] [CrossRef]
- Liang, H.; Jiao, Z.; Rong, W.; Qu, S.; Liao, Z.; Sun, X.; Wei, Y.; Zhao, Q.; Wang, J.; Liu, Y.; et al. 3’-Terminal 2’-O-methylation of lung cancer miR-21-5p enhances its stability and association with Argonaute 2. Nucleic Acids Res. 2020, 48, 7027–7040. [Google Scholar] [CrossRef]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G. Adenosine-to-inosine RNA editing meets cancer. Carcinog. 2011, 32, 1569–1577. [Google Scholar] [CrossRef]
- Caponio, V.C.A.; Troiano, G.; Botti, G.; Pedicillo, M.C.; Russo, L.L.; Mastrangelo, F.; Ciavarella, D.; Losito, N.S.; Aquino, G.; Nocini, R.; et al. Overexpression of ADAR 1 into the cytoplasm correlates with a better prognosis of patients with oral squamous cells carcinoma. J. Oral Pathol. Med. 2018, 48, 108–114. [Google Scholar] [CrossRef]
- Ramírez-Moya, J.; Baker, A.R.; Slack, F.J.; Santisteban, P. ADAR1-mediated RNA editing is a novel oncogenic process in thyroid cancer and regulates miR-200 activity. Oncogene 2020, 39, 3738–3753. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Fu, Y.; Huang, J.; Wu, M.; Zhang, Z.; Xu, R.; Zhang, P.; Zhao, S.; Liu, L.; Jiang, H. ADAR1 promotes the epithelial-to-mesenchymal transition and stem-like cell phenotype of oral cancer by facilitating oncogenic microRNA maturation. J. Exp. Clin. Cancer Res. 2019, 38, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.J.; Shen, H.; An, O.; Hong, H.; Li, J.; Song, Y.; Han, J.; Tay, D.J.T.; Ng, V.H.E.; Molias, F.B.; et al. Cis- and trans-regulations of pre-mRNA splicing by RNA editing enzymes influence cancer development. Nat. Commun. 2020, 11, 799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Li, Y.; Lin, C.H.; Chan, T.H.M.; Chow, R.K.K.; Song, Y.; Liu, M.; Yuan, Y.-F.; Fu, L.; Kong, K.L.; et al. Recoding RNA editing of AZIN1 predisposes to hepatocellular carcinoma. Nat. Med. 2013, 19, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Sagredo, E.A.; Blanco, A.; Sagredo, A.I.; Pérez, P.; Sepúlveda-Hermosilla, G.; Morales, F.; Müller, B.; Verdugo, R.; Marcelain, K.; Harismendy, O.; et al. ADAR1-mediated RNA-editing of 3′UTRs in breast cancer. Biol. Res. 2018, 51, 36. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Fan, J.; Wang, B.; Meng, Z.; Ren, D.; Zhao, J.; Liu, Z.; Li, D.; Jin, X.; Wu, H. The aberrant expression of ADAR1 promotes resistance to BET inhibitors in pancreatic cancer by stabiliz-ing c-Myc. Am. J. Cancer Res. 2020, 10, 148–163. [Google Scholar]
- Ishizuka, J.J.; Manguso, R.T.; Cheruiyot, C.K.; Bi, K.; Panda, A.; Iracheta-Vellve, A.; Miller, B.C.; Du, P.P.; Yates, K.B.; Dubrot, J.; et al. Loss of ADAR1 in tumours overcomes resistance to immune checkpoint blockade. Nat. Cell Biol. 2019, 565, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Mehdipour, P.; Marhon, S.A.; Ettayebi, I.; Chakravarthy, A.; Hosseini, A.; Wang, Y.; De Castro, F.A.; Yau, H.L.; Ishak, C.; Abelson, S.; et al. Epigenetic therapy induces transcription of inverted SINEs and ADAR1 dependency. Nat. Cell Biol. 2020, 588, 169–173. [Google Scholar] [CrossRef]
- Tomaselli, S.; Galeano, F.; Alon, S.; Raho, S.; Galardi, S.; Polito, V.A.; Presutti, C.; Vincenti, S.; Eisenberg, E.; Locatelli, F.; et al. Modulation of microRNA editing, expression and processing by ADAR2 deaminase in glioblastoma. Genome Biol. 2015, 16, 5. [Google Scholar] [CrossRef]
- Cesarini, V.; Silvestris, D.A.; Tassinari, V.; Tomaselli, S.; Alon, S.; Eisenberg, E.; Locatelli, F.; Gallo, A. ADAR2/miR-589-3p axis controls glioblastoma cell migration/invasion. Nucleic Acids Res. 2018, 46, 2045–2059. [Google Scholar] [CrossRef]
- Xu, X.; Wang, Y.; Mojumdar, K.; Zhou, Z.; Jeong, K.J.; Mangala, L.S.; Yu, S.; Tsang, Y.H.; Rodriguez-Aguayo, C.; Lu, Y.; et al. A-to-I–edited miRNA-379-5p inhibits cancer cell proliferation through CD97-induced apoptosis. J. Clin. Investig. 2019, 129, 5343–5356. [Google Scholar] [CrossRef] [Green Version]
- Freeman, J.W.; Busch, R.K.; Gyorkey, F.; Gyorkey, P.; Ross, B.E.; Busch, H. Identification and characterization of a human proliferation-associated nucleolar antigen with a molecular weight of 120,000 expressed in early G1 phase. Cancer Res. 1988, 48, 1244–1251. [Google Scholar]
- Hazlewood, J.; Fonagy, A.; Henning, D.; Freeman, J.W.; Busch, R.K.; Busch, H. mRNA Levels For Human Nucleolar Protein P120 in Tumor and Nontumor Cells. Cancer Commun. 1989, 1, 29–34. [Google Scholar] [CrossRef]
- Freeman, J.; McGrath, P.; Bondada, V.; Selliah, N.; Ownby, H.; Maloney, T.; Busch, R.; Busch, H. Prognostic significance of proliferation associated nucleolar antigen P120 in human breast carcinoma. Cancer Res. 1991, 51, 1973–1978. [Google Scholar] [PubMed]
- Sun, F.; Wu, K.; Yao, Z.; Mu, X.; Zheng, Z.; Sun, M.; Wang, Y.; Liu, Z.; Zhu, Y. Long Noncoding RNA PVT1 Promotes Prostate Cancer Metastasis by Increasing NOP2 Expression via Targeting Tumor Suppressor MicroRNAs. OncoTargets Ther. 2020, 13, 6755–6765. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Wu, K.; Yao, Z.; Mu, X.; Zheng, Z.; Sun, M.; Wang, Y.; Liu, Z.; Zhu, Y. Long noncoding RNA LINC00963 induces NOP2 expression by sponging tumor suppressor miR-542-3p to promote metastasis in prostate cancer. Aging 2020, 12, 11500–11516. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, Z.; Zhu, Y.; Zhu, Q.; Yang, Y.; Jin, Y.; Zhang, F.; Jiang, L.; Ye, Y.; Li, H.; et al. NOP 2/Sun RNA methyltransferase 2 promotes tumor progression via its interacting partner RPL 6 in gallbladder carcinoma. Cancer Sci. 2019, 110, 3510–3519. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, M.; Hirata, S.; Sato, S.; Koga, S.; Fujii, M.; Qi, G.; Ogawa, I.; Takata, T.; Shimamoto, F.; Tatsuka, M. Frequent increased gene copy number and high protein expression of tRNA (cyto-sine-5-)-methyltransferase (NSUN2) in human cancers. DNA Cell. Biol. 2012, 31, 660–671. [Google Scholar] [CrossRef]
- Frye, M.; Watt, F.M. The RNA methyltransferase Misu (NSun2) mediates Myc-induced proliferation and is upregulated in tumors. Curr. Biol. 2006, 16, 971–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Li, A.; Sun, B.-F.; Yang, Y.; Han, Y.-N.; Yuan, X.; Chen, R.-X.; Wei, W.-S.; Liu, Y.; Gao, C.-C.; et al. 5-methylcytosine promotes pathogenesis of bladder cancer through stabilizing mRNAs. Nat. Cell Biol. 2019, 21, 978–990. [Google Scholar] [CrossRef]
- Xing, J.; Yi, J.; Cai, X.; Tang, H.; Liu, Z.; Zhang, X.; Martindale, J.L.; Yang, X.; Jiang, B.; Gorospe, M.; et al. NSun2 Promotes Cell Growth via Elevating Cyclin-Dependent Kinase 1 Translation. Mol. Cell. Biol. 2015, 35, 4043–4052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.; Fan, X.; Xing, J.; Liu, Z.; Jiang, B.; Dou, Y.; Gorospe, M.; Wang, W. NSun2 delays replicative senescence by repressing p27 (KIP1) translation and elevating CDK1 translation. Aging 2015, 7, 1143–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, L.; Shen, C.; Miao, R.; Wang, J.-Z.; Cao, M.-D.; Zhang, Y.-S.; Shi, L.-H.; Zhao, G.-H.; Wang, M.-H.; Wu, L.-S.; et al. RNA methyltransferase NSUN2 promotes gastric cancer cell proliferation by repressing p57Kip2 by an m5C-dependent manner. Cell Death Dis. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Subcellular Distribution of RNA Modifications and RMPs in Eukaryotes. Abbreviations: RMP, RNA modifying protein; RBP, RNA binding proteins; m7G, 7-methylguanosine; Nm, 2′-O-methylation; m6A, N6-methyladenosine; m5C, 5-methylcytosine; hnRNPs, heterogeneous nuclear ribonucleoproteins.
Figure 1.
Subcellular Distribution of RNA Modifications and RMPs in Eukaryotes. Abbreviations: RMP, RNA modifying protein; RBP, RNA binding proteins; m7G, 7-methylguanosine; Nm, 2′-O-methylation; m6A, N6-methyladenosine; m5C, 5-methylcytosine; hnRNPs, heterogeneous nuclear ribonucleoproteins.

Figure 2.
Formation, Recognition and Removal of RNA m6A and m5C in Eukaryotes. (A). Formation, Recognition and Removal of RNA:m6A. The METTL3/14 methyltransferase complex transfers methyl groups from SAM to N6-adenosines at the RRAH motifs in RNA. m6A is then recognized by m6A readers (m6A-selective binding proteins), and eventually removed by RNA m6A erasers. (B). Formation, Recognition and Removal of RNA:m5C. RNA m5C writers methylate cytosine residues, which are then recognized by m5C readers, or TETs, which oxidize m5C to hm5C, f5C and ca5C, respectively. Abbreviations: SAM, S-adenosylmethionine; SAH, S-adenosyl homocysteine; RBM15, RNA binding motif protein 15; METTL3/14, methyltransferase like 3/14; ZC3H13, zinc finger CCCH-type containing 13; WTAP, Wilms tumor suppressor gene WT1; VIRMA, Vir-like m6A methyltransferase associated; HAKAI, Cbl Proto-Oncogene Like 1; FTO, Fat mass and obesity associated; ALKBH5, AlkB homolog 5; YTHDC1/2, YTH domain containing 1/2; YTHDF2/3, YTH N6, methyladenosine RNA-binding protein 2/3; HNRNP family, heterogeneous nuclear ribonucleoproteins; IGF2BP1/2/3, insulin-like growth factor 2 mRNA binding protein 1/2/3. NSUN family, NOL1/NOP2/sun domain; DNMT2, DNA methyltransferase 2; ALYREF, Aly/REF export factor; YTHDF2, YTH N6-methyladenosine RNA binding protein 2; TETs, ten eleven translocation elements; hm5C, 5-hydroxymethylcytosine; f5C, 5-formylcytosine; ca5C, 5-carboxylcytosine.
Figure 2.
Formation, Recognition and Removal of RNA m6A and m5C in Eukaryotes. (A). Formation, Recognition and Removal of RNA:m6A. The METTL3/14 methyltransferase complex transfers methyl groups from SAM to N6-adenosines at the RRAH motifs in RNA. m6A is then recognized by m6A readers (m6A-selective binding proteins), and eventually removed by RNA m6A erasers. (B). Formation, Recognition and Removal of RNA:m5C. RNA m5C writers methylate cytosine residues, which are then recognized by m5C readers, or TETs, which oxidize m5C to hm5C, f5C and ca5C, respectively. Abbreviations: SAM, S-adenosylmethionine; SAH, S-adenosyl homocysteine; RBM15, RNA binding motif protein 15; METTL3/14, methyltransferase like 3/14; ZC3H13, zinc finger CCCH-type containing 13; WTAP, Wilms tumor suppressor gene WT1; VIRMA, Vir-like m6A methyltransferase associated; HAKAI, Cbl Proto-Oncogene Like 1; FTO, Fat mass and obesity associated; ALKBH5, AlkB homolog 5; YTHDC1/2, YTH domain containing 1/2; YTHDF2/3, YTH N6, methyladenosine RNA-binding protein 2/3; HNRNP family, heterogeneous nuclear ribonucleoproteins; IGF2BP1/2/3, insulin-like growth factor 2 mRNA binding protein 1/2/3. NSUN family, NOL1/NOP2/sun domain; DNMT2, DNA methyltransferase 2; ALYREF, Aly/REF export factor; YTHDF2, YTH N6-methyladenosine RNA binding protein 2; TETs, ten eleven translocation elements; hm5C, 5-hydroxymethylcytosine; f5C, 5-formylcytosine; ca5C, 5-carboxylcytosine.

Figure 3.
Molecular Reactions of RNA Adenosine to Inosine (A-to-I) and Cytidine to Uridine (C-to-U) Editing in Eukaryotes. (A). A-to-I RNA Editing Mechanism. ADAR1 and ADAR2 catalyze the site-specific conversion of A-to-I within imperfectly duplexed RNA. Meanwhile ADAR3 inhibits A-to-I editing. (B). C-to-U RNA Editing Mechanism. APOBEC1 and ACF bind to the RNA duplex, and RBM47 interacts with APOBEC1 and ACF, to produce C-to-U conversion via hydrolytic deamination of cytidine. Abbreviations: ADAR1/2, adenosine deaminases acting on RNA; APOBEC1, apolipoprotein B mRNA editing enzyme catalytic subunit 1; RBM47, RNA binding motif protein 47; ACF/A1CF, APOBEC1-complementation factor.
Figure 3.
Molecular Reactions of RNA Adenosine to Inosine (A-to-I) and Cytidine to Uridine (C-to-U) Editing in Eukaryotes. (A). A-to-I RNA Editing Mechanism. ADAR1 and ADAR2 catalyze the site-specific conversion of A-to-I within imperfectly duplexed RNA. Meanwhile ADAR3 inhibits A-to-I editing. (B). C-to-U RNA Editing Mechanism. APOBEC1 and ACF bind to the RNA duplex, and RBM47 interacts with APOBEC1 and ACF, to produce C-to-U conversion via hydrolytic deamination of cytidine. Abbreviations: ADAR1/2, adenosine deaminases acting on RNA; APOBEC1, apolipoprotein B mRNA editing enzyme catalytic subunit 1; RBM47, RNA binding motif protein 47; ACF/A1CF, APOBEC1-complementation factor.

Figure 4.
5′ Cap Structure of a Human Pre-mRNA and the Associated Methylations and Methyltransferase Complexes. Abbreviations: m7G, 7-methylguanosine; RGMT, RNA guanine methyltransferase; RNGTT, RNA guanylyltransferase and 5′ phosphatase; RTPase-RNA triphosphatase; SAM, S-adenosylmethionine; SAH, S-adenosyl homocysteine; PCIF1/CAPAM, PDX C-terminal inhibiting factor 1; CMTR1/2, cap methyltransferase 1/2.
Figure 4.
5′ Cap Structure of a Human Pre-mRNA and the Associated Methylations and Methyltransferase Complexes. Abbreviations: m7G, 7-methylguanosine; RGMT, RNA guanine methyltransferase; RNGTT, RNA guanylyltransferase and 5′ phosphatase; RTPase-RNA triphosphatase; SAM, S-adenosylmethionine; SAH, S-adenosyl homocysteine; PCIF1/CAPAM, PDX C-terminal inhibiting factor 1; CMTR1/2, cap methyltransferase 1/2.

Figure 5.
Transcriptionally Active Chromatin Structure and the Five Steps of Transcription Activation. Step 1. Chromatin remodeling; Step 2. Formation of transcription pre-initiation complex (PIC); Step 3. Unwinding of the DNA strands to form the open complex; Step 4. RNA polymerase II (RNAPII) pausing and escaping from the promoter; Step 5. Productive transcript elongation by RNAPII. Abbreviations: SWI-SNF, switch/Sucrose non-fermentable; TF, transcription factor; H, histone; PIC, preinitiation complex; GTFs, general transcription factors; TFIIA-F, transcription factor II A–F; TAF1, TATA-box binding protein associated factor 1; TBP, TATA-box binding protein; CDK7/TFIIH, cyclin-dependent kinase 7; TSS, transcription start Site; Spt5/4, suppressor of Ty 5 and 4 (transcription elongation factors 5 and 4); DSIF, DRB sensitivity-inducing factor; CDK9/p-TEFb, cyclin-dependent kinase 9.
Figure 5.
Transcriptionally Active Chromatin Structure and the Five Steps of Transcription Activation. Step 1. Chromatin remodeling; Step 2. Formation of transcription pre-initiation complex (PIC); Step 3. Unwinding of the DNA strands to form the open complex; Step 4. RNA polymerase II (RNAPII) pausing and escaping from the promoter; Step 5. Productive transcript elongation by RNAPII. Abbreviations: SWI-SNF, switch/Sucrose non-fermentable; TF, transcription factor; H, histone; PIC, preinitiation complex; GTFs, general transcription factors; TFIIA-F, transcription factor II A–F; TAF1, TATA-box binding protein associated factor 1; TBP, TATA-box binding protein; CDK7/TFIIH, cyclin-dependent kinase 7; TSS, transcription start Site; Spt5/4, suppressor of Ty 5 and 4 (transcription elongation factors 5 and 4); DSIF, DRB sensitivity-inducing factor; CDK9/p-TEFb, cyclin-dependent kinase 9.

Figure 6.
MYC-mediated RNA 5′ Capping, Transcription Elongation and Chromatin Structure. Abbreviations: E-box- enhancer box; MYC- MYC proto-oncogene BHLH transcription factor; RNMT, RNA methyltransferase; CMTR1, cap methyltransferase 1; K-ac, acetylated lysine; CDK9/P-TEFb, cyclin-dependent Kinase 9; CDK7/TFIIH, Cyclin-dependent Kinase 7; BRD4, Bromodomain Containing 4; 7SK snRNP, 7SK small nuclear ribonucleoprotein.
Figure 6.
MYC-mediated RNA 5′ Capping, Transcription Elongation and Chromatin Structure. Abbreviations: E-box- enhancer box; MYC- MYC proto-oncogene BHLH transcription factor; RNMT, RNA methyltransferase; CMTR1, cap methyltransferase 1; K-ac, acetylated lysine; CDK9/P-TEFb, cyclin-dependent Kinase 9; CDK7/TFIIH, Cyclin-dependent Kinase 7; BRD4, Bromodomain Containing 4; 7SK snRNP, 7SK small nuclear ribonucleoprotein.

Figure 7.
Transcription-associated Spliceosome Assembly and Alternative Splicing in Eukaryotes. (A). Determination of the first exon-intron boundary and formation of initial spliceosome on pre-mRNA. (B). Transcription-associated determination of the internal exon-intron boundaries, spliceosome assembly and alternative splicing of pre-mRNA. Abbreviations: CTCF, CCCTC binding protein; MeCP2, methyl-CpG binding protein 2; DNMT, DNA methyltransferase; TET, ten eleven translocation elements; E-box, enhancer box; CE/RNGTT, RNA guanylyltransferase and 5′-Phosphatase; HMT, histone methyltransferase; HAT, histone acetyltransferase; BRD4, bromodomain containing 4; U1-U6, small nuclear ribonucleoprotein U1-U6 subunit; CDK9/P-TEFb, cyclin-dependent kinase 9; 7SK snRNP, 7SK small nuclear ribonucleoprotein; SF3A/B, splicing factor 3a/b subunit; CDK7/TFIIH, cyclin-dependent kinase 7; TETs, ten eleven translocation elements; SRSF, serine/arginine-rich splicing factors; hnRNP, heterogeneous nuclear ribonucleoproteins; RBP, RNA binding proteins.
Figure 7.
Transcription-associated Spliceosome Assembly and Alternative Splicing in Eukaryotes. (A). Determination of the first exon-intron boundary and formation of initial spliceosome on pre-mRNA. (B). Transcription-associated determination of the internal exon-intron boundaries, spliceosome assembly and alternative splicing of pre-mRNA. Abbreviations: CTCF, CCCTC binding protein; MeCP2, methyl-CpG binding protein 2; DNMT, DNA methyltransferase; TET, ten eleven translocation elements; E-box, enhancer box; CE/RNGTT, RNA guanylyltransferase and 5′-Phosphatase; HMT, histone methyltransferase; HAT, histone acetyltransferase; BRD4, bromodomain containing 4; U1-U6, small nuclear ribonucleoprotein U1-U6 subunit; CDK9/P-TEFb, cyclin-dependent kinase 9; 7SK snRNP, 7SK small nuclear ribonucleoprotein; SF3A/B, splicing factor 3a/b subunit; CDK7/TFIIH, cyclin-dependent kinase 7; TETs, ten eleven translocation elements; SRSF, serine/arginine-rich splicing factors; hnRNP, heterogeneous nuclear ribonucleoproteins; RBP, RNA binding proteins.

Figure 8.
RNA Modification/RMP-mediated, Transcription-associated Chromatin Structural Changes in Mammalian Cells. (A). m6A and its RMPs-mediated chromatin structural changes. (B). m5C and its RMPs-mediated chromatin structural changes. Abbreviations: m6A, N6-methyladenine; RNMT, RNA Methyltransferase; CMTR1, cap methyltransferase 1; KDM3B, lysine demethylase 3B; YTHDC1, YTH Domain Containing 1; EZH2, enhancer of Zeste 2 Polycomb repressive Complex 2 Subunit; METTL3/14, methyltransferase Like 3/14; K9me, lysine 9 methyl; K27me, lysine 27 methyl; TFs, Transcription Factors; ZFP217, zinc finger Protein 217; H3K4me36, histone 3 lysine 36 tri-methylation; E-box, enhancer box; CDK9/P-TEFb, cyclin-dependent kinase 9; CDK7/TFIIH, cyclin-dependent kinase 7. m5C, 5-methylcytosine; MeCP2, methyl-CpG binding protein 2; SET7/9, SET Domain containing 7/9 histone lysine methyltransferase; SAGA, Spt-Ada-Gcn5-acetyltransferase; PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1-α; NSUN7, NOP2/Sun RNA methyltransferase member 7; H3K4me36, histone 3 lysine 36 tri-methylation; NOP2/NSUN1, NOP2 nucleolar protein/NOP2/Sun RNA methyltransferase 1; K-ac, lysine acetylation; BRD5/BET, bromodomain containing 4; E-box, enhancer box; CDK9/pTEFb, cyclin-dependent kinase 9; CDK7/TFIIH, cyclin-dependent Kinase 7; MYC, MYC proto-oncogene BHLH transcription factor.
Figure 8.
RNA Modification/RMP-mediated, Transcription-associated Chromatin Structural Changes in Mammalian Cells. (A). m6A and its RMPs-mediated chromatin structural changes. (B). m5C and its RMPs-mediated chromatin structural changes. Abbreviations: m6A, N6-methyladenine; RNMT, RNA Methyltransferase; CMTR1, cap methyltransferase 1; KDM3B, lysine demethylase 3B; YTHDC1, YTH Domain Containing 1; EZH2, enhancer of Zeste 2 Polycomb repressive Complex 2 Subunit; METTL3/14, methyltransferase Like 3/14; K9me, lysine 9 methyl; K27me, lysine 27 methyl; TFs, Transcription Factors; ZFP217, zinc finger Protein 217; H3K4me36, histone 3 lysine 36 tri-methylation; E-box, enhancer box; CDK9/P-TEFb, cyclin-dependent kinase 9; CDK7/TFIIH, cyclin-dependent kinase 7. m5C, 5-methylcytosine; MeCP2, methyl-CpG binding protein 2; SET7/9, SET Domain containing 7/9 histone lysine methyltransferase; SAGA, Spt-Ada-Gcn5-acetyltransferase; PGC-1α, peroxisome proliferator-activated receptor γ coactivator 1-α; NSUN7, NOP2/Sun RNA methyltransferase member 7; H3K4me36, histone 3 lysine 36 tri-methylation; NOP2/NSUN1, NOP2 nucleolar protein/NOP2/Sun RNA methyltransferase 1; K-ac, lysine acetylation; BRD5/BET, bromodomain containing 4; E-box, enhancer box; CDK9/pTEFb, cyclin-dependent kinase 9; CDK7/TFIIH, cyclin-dependent Kinase 7; MYC, MYC proto-oncogene BHLH transcription factor.


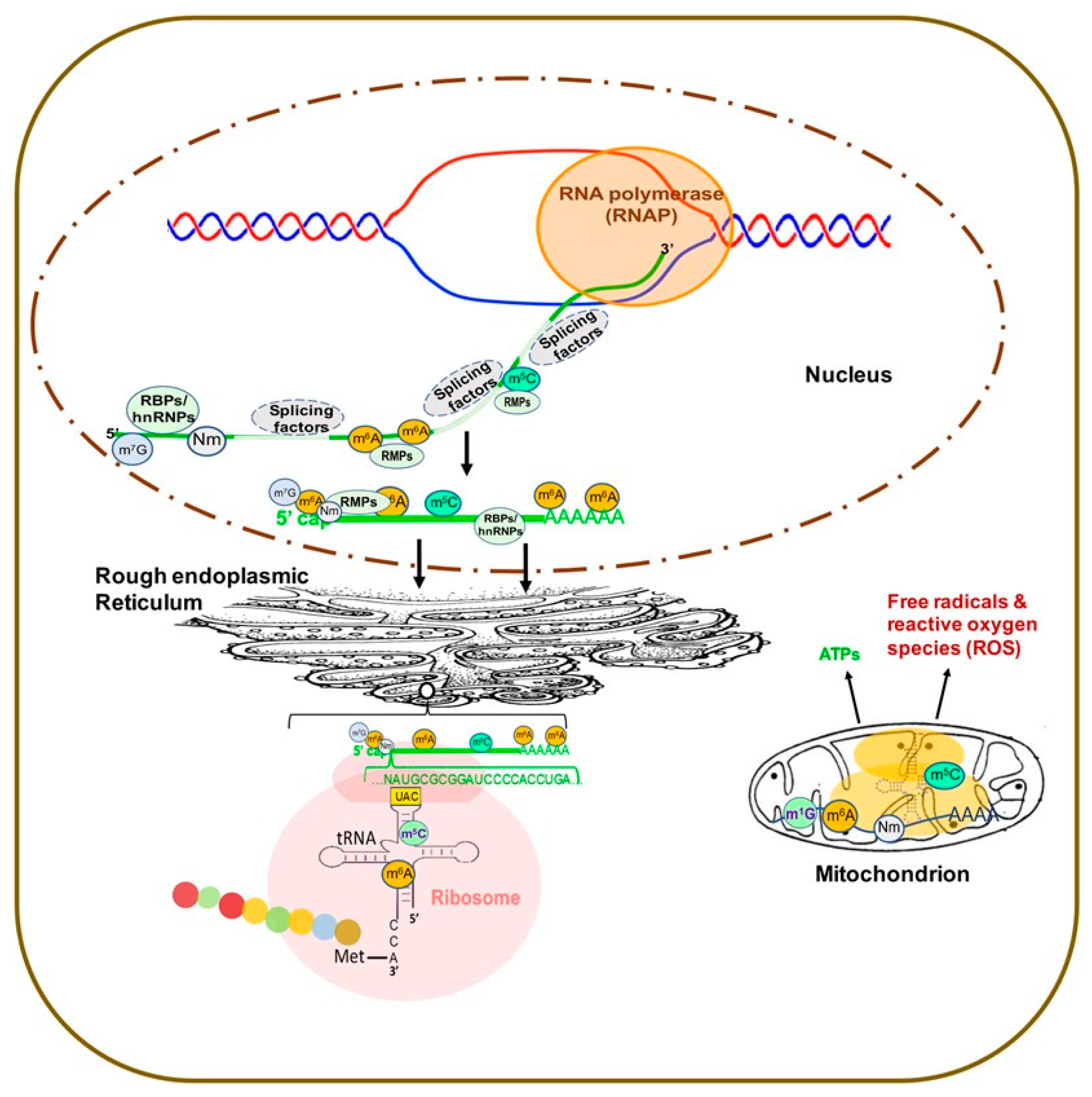{kind=link}
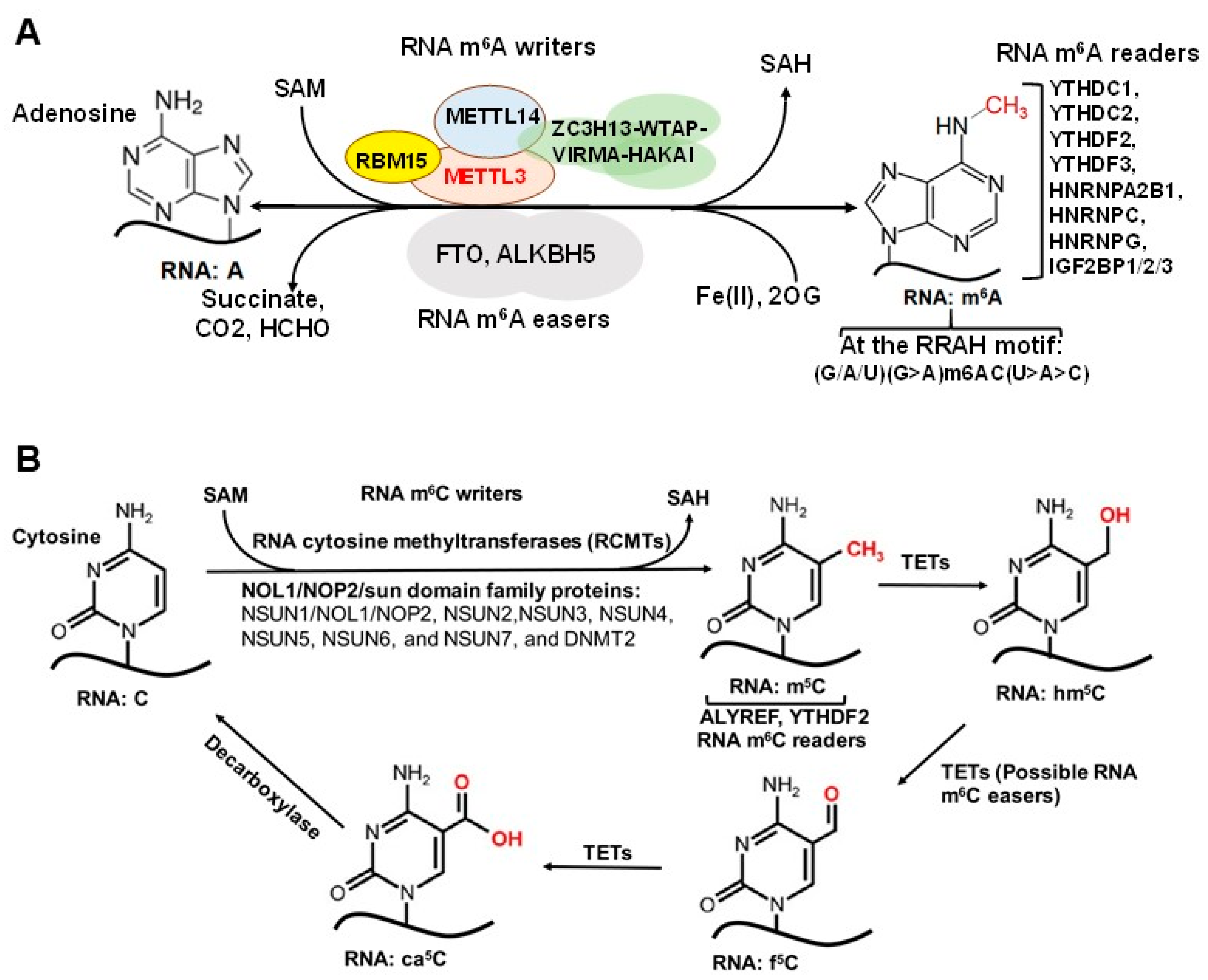{kind=link}
{kind=link}
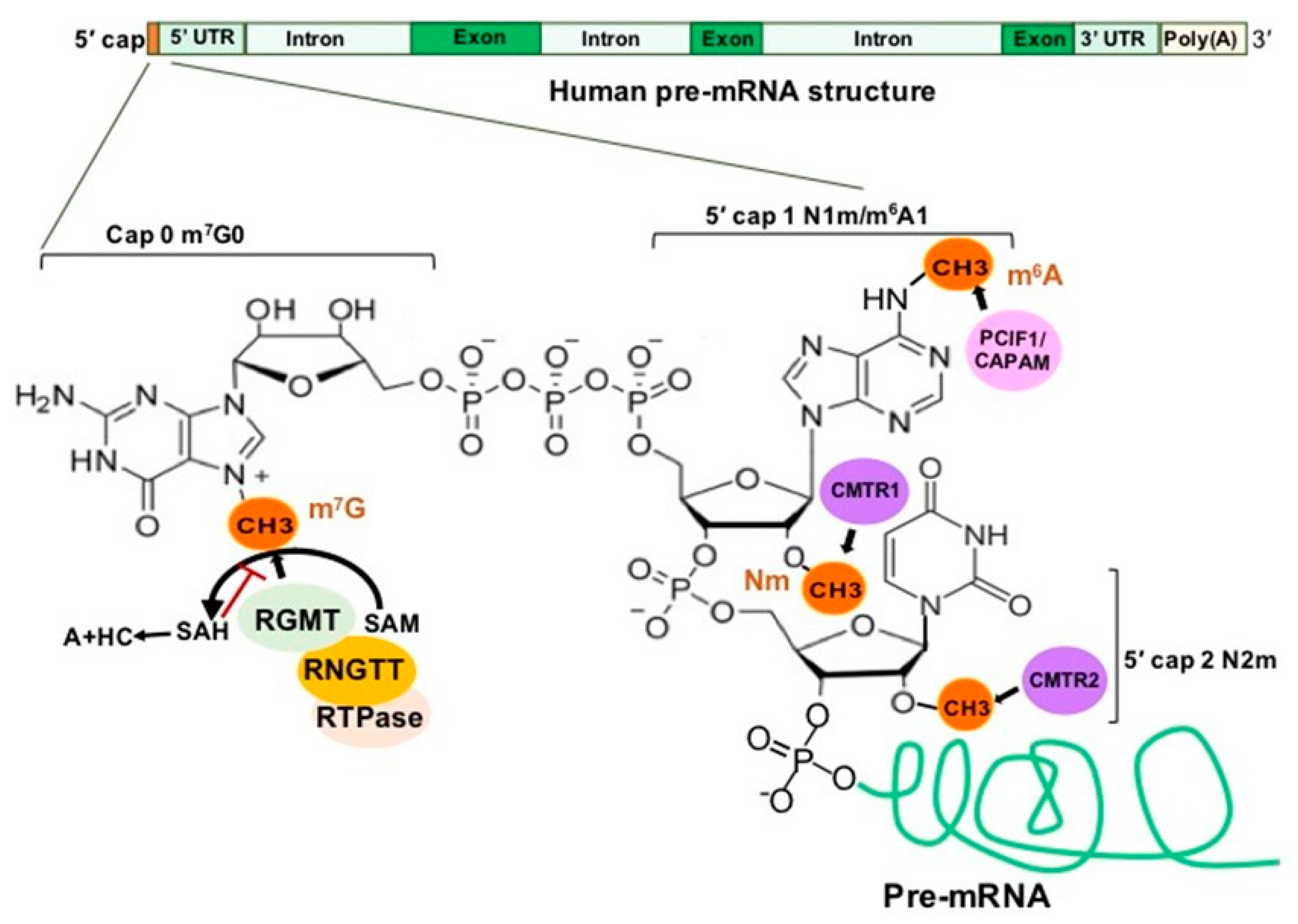{kind=link}
{kind=link}
{kind=link}
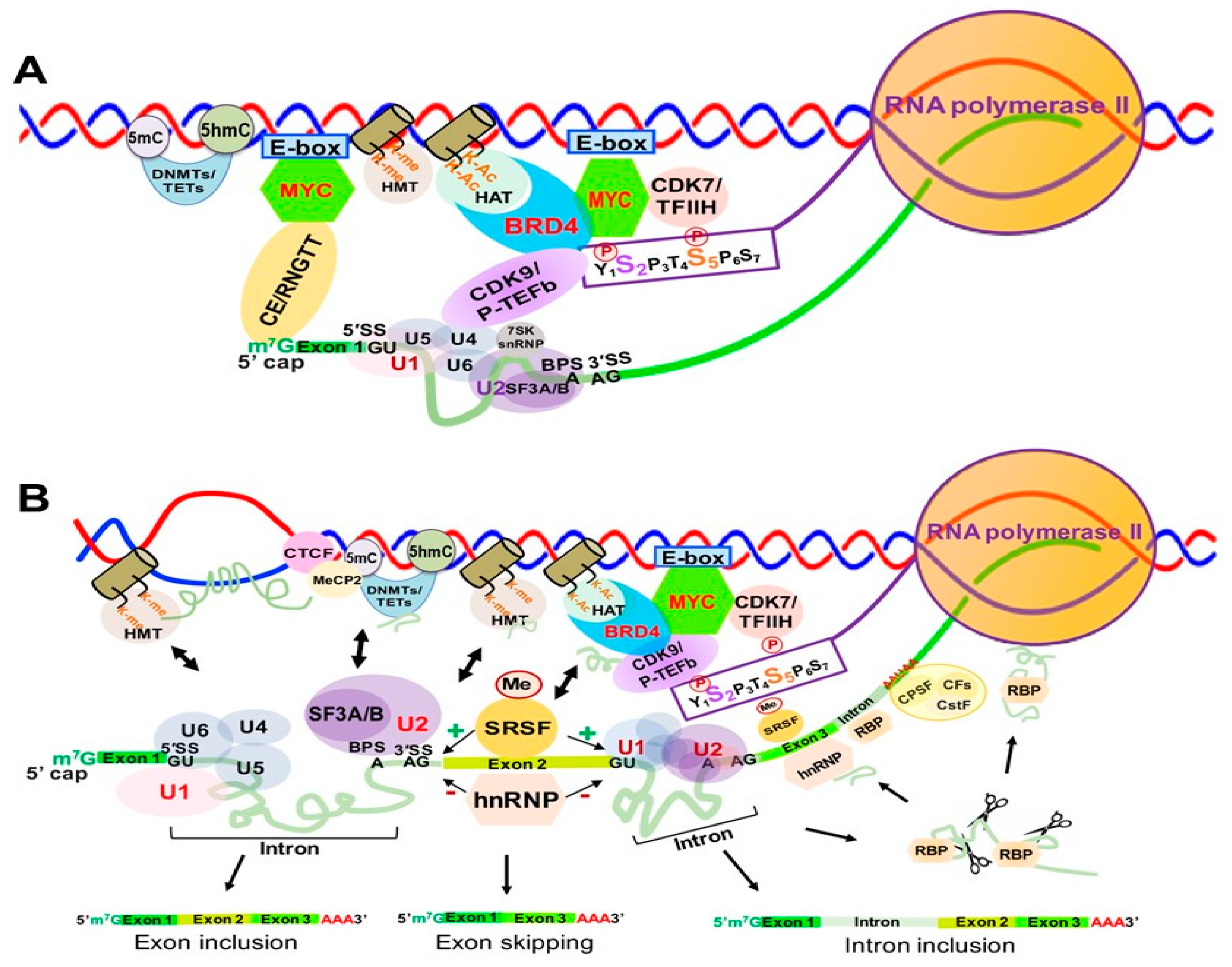{kind=link}
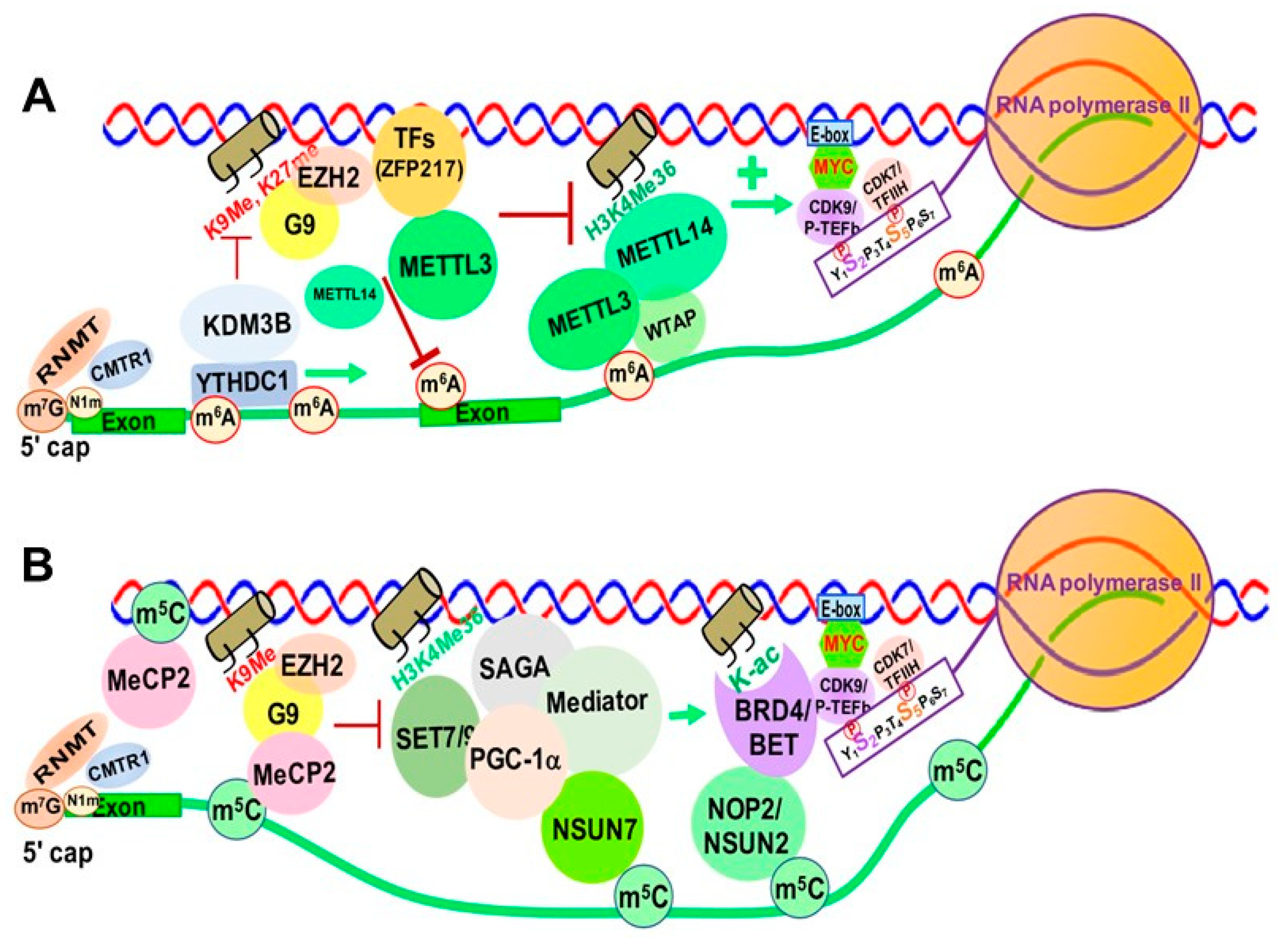{kind=link}
Table 1.
The types and functions of regulatory non-coding RNAs (ncRNAs) in mammalian cells.
| Types of ncRNAs | NcRNA Abbreviation | ncRNA Strands | Size of ncRNAs | Function | References |
|---|---|---|---|---|---|
| Small ncRNAs | <200 nucleotides | ||||
| Small interfering RNAs | siRNAs | double | 21–23 | suppress homology containing transcripts via mRNA degradation | [179,180] |
| Micro-RNAs | miRNAs | single | ~22 | repress translation; accelerate mRNA degradation | [181] |
| Piwi-interacting RNAs | piRNAs | single | 21–25 | regulate gene expression; fight viral infection | [182,183] |
| Small nucleolar RNAs | snoRNAs | single | 60–300 | rRNA, snRNA, and other RNA posttranscriptional modification | [182,183] |
| Small Cajal body RNAs (snoRNA) | scaRNAs | single | 60–300 | biochemical modification of premature spliceosomal RNAs | [182,183] |
| Long non-coding RNAs | lncRNAs | single | >200 nucleotides | Chromatin remodeling; transcriptional and post-transcriptional regulation | [184] |
Table 2.
Implications of common RNA modifications and RNA modifying proteins in human diseases.
| Disease Categories/ Subcategories | RNA Modifications & RMPs | Possible Pathogenic Mechanisms | References |
|---|---|---|---|
| Neuronal Systems: 1. Neurogenesis defects & neural degenerative disorders. 2. Memory defects 3. Psychiatric Diseases 4. Intellectual Disability | M6A: | ||
| METTL3/14 | Knockout (KO) of Mettl3/14 → ↑ cell cycle and ↓ NSC proliferation via histone modifications | [268,269,270,271] | |
| YTHDF1 | ↑ Translation of axon guidance protein Robo3.1 | [272] | |
| FTO | FTO regulates Pdfra/Socs5-Stat3; TSC1-mTOR-Tau signaling | [273,274] | |
| RNA editing: | |||
| ADARs | Affect mRNA abundance and miRNA binding→ regulating development and function of neurons and neurotransmitters | [283,284] | |
| Nm: | |||
| FTSJ1, CMTR1 | FTSJ1 affects tRNA:Nm and neurotransmitter translationCMTR1 affects 5′ cap RNA: Nm and translation | [286,287,288,289] | |
| M5C: | |||
| NSUN2 | Affects tRNA:m5C and protein translation | [291,292,293,294,295,296] | |
| Metabolic Disorders: 1 Type 2 Diabetes (T2D) 2. Obesity | M6A: | ||
| METTL3, | ↑METTL3/14/WTAP (↑ insulin-IGF1-AKT-PDX1 pathway(↑ β-cell survival(glucose homeostasis | [297,298,299] | |
| METTL14 | ↑METTL3/14/WTAP (↑MCE (↑ adipogenesis | [300] | |
| YTHDF2 | YTHDF2 →↓ ZFP217-induced MCE → ↓ adipogenesis | [301] | |
| FTO | ↑ FTO→ ↑ adipogenic RUNX1T1; ↓Wnt/β-catenin; ↑ATG5/7→ adipogenesis | [302,303,304,305,306,307] | |
| Immune System: 1. Innate immunity, autoimmune & viral infections 2. Adaptive immunity | M6A: | ||
| METTL3 | ↓METTL3/14 or YTHDF2( ↑ IFNB1 mRNA | [308,309,310,311,312,313,314,315,316,317,318] | |
| METTL14 | ↓METTL3( ↓Rev-RRE binding( ↓viral RNA export & production | ||
| YTHDF2 | ↓METTL3( ↓DC activation; ↓ IL7/2-STAT5-SOCS | ||
| A-to-I editing | |||
| ADAR1/p150 | ↓ADAR1→ ↑dsRNA sensor-triggered INF responses → autoimmune and cell death | [319,320,321] | |
| 5′ cap Nm: | |||
| CMTR1 | ↑5′cap Nm→ ↓viral mRNA binding by sensor Mda5 & IFIT1→ ↓INF | [21,322,323,324,325,326] | |
| Cancer & Leukemia 1. Tumorigenesis, cell proliferation, and metastasis 2. Immune evasion and immunotherapy 3. Drug Resistance Cancer & Leukemia1 1. Tumorigenesis, cell proliferation, and metastasis 2. Immune evasion and immunotherapy 3. Drug Resistance | M6A: METTL3 YTHDF1 YTHDF2 YTHDC1 YTHDC2 FTO | Acting as oncogene: | |
| ↑METTL3( ↑oncogenic mRNA & miRNAs; ↑MYC; ↑Wnt/β-catenin signaling | [332,333,334,335,336,337,338,339,340,341,342,343] | ||
| ↑YTHDF1( ↑EiF3C translation; ↑Wnt/β-catenin; ↓ antigen presentation by DCs( immune evasion | [348,349,350,351,352] | ||
| ↑YTHDF2( ↑oncogenic lncRNA; ↑YAP signalng; ↑YTHDC( ↑translation of c-JUN, ATF-2, HIF-1a | [353,354,355,356,357] | ||
| ↑ FTO( ↑ PI3K/AKT; ↓ BNIP3 mRNA | [360,361] | ||
| METTL14 YTHDF1 YTHDF2 FTO | Acting as tumor suppressor: | ||
| ↑METTL14 ↑ (↑tumor suppressor miR-375/Yes-associated protein 1 (YAP1) pathway; ↓lncRNA XIST; ↓ERK9/MMP9 signaling; ↓oncogenic miRNAs | [339,344,345,346,347] | ||
| ↑YTHDF1( ↑ tumor suppressor HINT2; ↑YTHDF2( Destabilizing EGFR mRNA | [358,359] | ||
| ↑ FTO(↑PGC-1α signaling | [362] | ||
| Nm: Fibrillarin HENMT1 | Acting as oncogene: | ||
| ↑Fibrillarin(↑ rRNA:Nm(↑translation of oncogenic proteins | [365] | ||
| ↑HENMT1(stablizing miR-21-5p(↓programed cell death protein 4 | [369] | ||
| RNA editing: ADAR1/p150 ADAR2 | Acting as oncogene: | ||
| ↑ADAR1(↑oncogenic miRNAs; ↑SRSF7-mediated intron inclusion; “gain-of-function” mutation in AZIN1 | [371,372,373,374,375] | ||
| ↑ADAR1(A-to-I editing in 3′UTRs of ATM, GINS4 and POLH Mrna | [376] | ||
| ↑ADAR1( ↑ resistance to BET inhibitors & checkpoint blockade | [377,378,379] | ||
| Acting as tumor suppressor: ↑ ADAR2( ↑oncogenic miRNAs; ↑ tumor suppressor miRNAs | [380,381,382] | ||
| M5C: NSUN1 NSUN2 | Acting as tumor suppressor: | ||
| ↑NSUN1(↑BRD4-mediated eRNAPII recruitment to pre-mRNA | [127] | ||
| ↑NSUN1(↑oncogenic mRNA & miRNAs | [383,384,385,386,387,388] | ||
| ↑NSUN2(↑MYC-mediated RNAPIII transcripts; ↑mRNA stability | [389,390,391,392,393,394] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Willbanks, A.; Wood, S.; Cheng, J.X. RNA Epigenetics: Fine-Tuning Chromatin Plasticity and Transcriptional Regulation, and the Implications in Human Diseases. Genes 2021, 12, 627. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050627
AMA Style
Willbanks A, Wood S, Cheng JX. RNA Epigenetics: Fine-Tuning Chromatin Plasticity and Transcriptional Regulation, and the Implications in Human Diseases. Genes. 2021; 12(5):627. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050627
Chicago/Turabian StyleWillbanks, Amber, Shaun Wood, and Jason X. Cheng. 2021. "RNA Epigenetics: Fine-Tuning Chromatin Plasticity and Transcriptional Regulation, and the Implications in Human Diseases" Genes 12, no. 5: 627. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050627
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.