
Novel QTL Associated with Shoot Branching Identified in Doubled Haploid Rice (Oryza sativa L.) under Low Nitrogen Cultivation


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mapping Population, Growth Conditions, and Phenotypic Measurements
2.2. Frequency Distribution, Quantile–Quantile Plots, Principal Component Analysis, and Correlation Analysis
2.3. Genotyping and Molecular Markers Analysis
2.4. Construction of Linkage Maps and QTL Analysis
3. Results
3.1. Differential Growth Patterns between Doubled Haploid Lines under Normal and Low Nitrogen Conditions
3.2. Relatedness, Principal Component Analysis, and Correlation between Traits
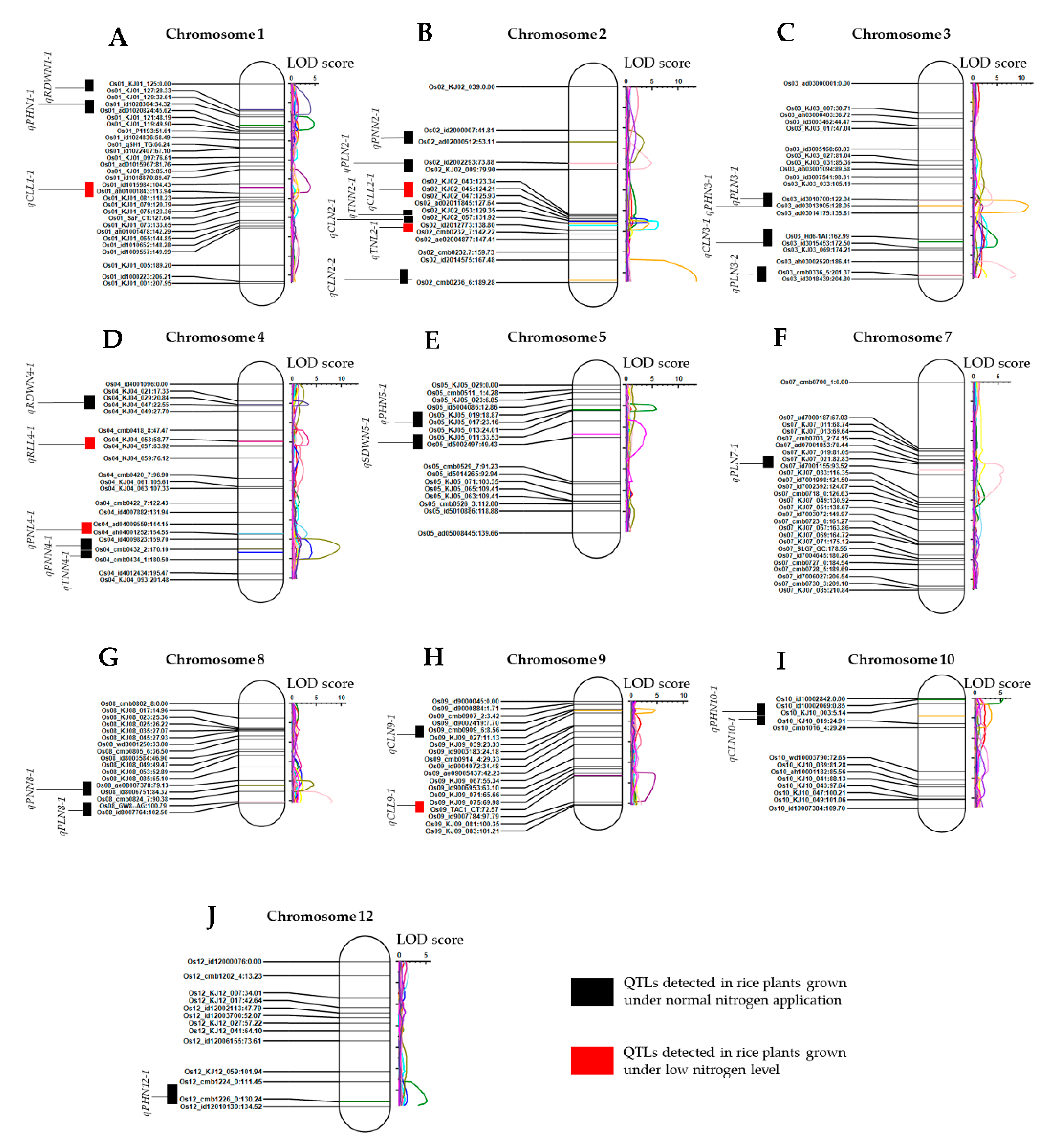
3.3. Quantitative Trait Loci (QTLs) Associated with Plant Growth and Shoot Branching under Normal and Low Nitrogen Cultivation Were Mapped to Different Chromosomes
4. Discussion
4.1. Tiller Number Positively Correlated with Shoot Dry Weight, While Shoot and Root Dry Weights Showed Positive Relationship
4.2. Quantiative Trait Loci (QTLs) Associated with Shoot Branching under Low Nitrogen Cultivation Conditions Are Identified
4.3. QTLs qTNL2-1 and qTN4-1 Harbor Candidate Genes Proposed to Be Involved in the Control of Shoot Branching in Rice under Low and Regular Nitrogen Cultivation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DESA UN. World Population Projected to Reach 9.8 Billion in 2050, and 11.2 Billion in 2100. 2019, Volume 2. Available online: https://www.un.org/development/desa/en/news/population/world-population-prospects-2017.html (accessed on 8 March 2021).
- Olson, R.; Kurtz, L. Crop nitrogen requirements, utilization, and fertilization. Nitrogen Agric. Soils 1982, 22, 567–604. [Google Scholar] [CrossRef]
- Leghari, S.J.; Wahocho, N.A.; Laghari, G.M.; Hafeez Laghari, A.; Mustafa Bhabhan, G.; Hussain Talpur, K.; Bhutto, T.A.; Wahocho, S.A.; Lashari, A.A. Role of nitrogen for plant growth and development: A review. Adv. Environ. Biol. 2016, 10, 209–219. [Google Scholar]
- Causin, H.F. The central role of amino acids on nitrogen utilization and plant growth. J. Plant Physiol. 1996, 149, 358–362. [Google Scholar] [CrossRef]
- Novoa, R.; Loomis, R.S. Nitrogen and plant production. Plant Soil 1981, 58, 177–204. [Google Scholar] [CrossRef]
- Kirkby, E. Plant growth in relation to nitrogen supply. Ecol. Bull. 1981, 33, 249–267. [Google Scholar]
- Van Keulen, H. Nitrogen Requirements of Rice with Special Reference to Java; CRIA: Bogor, Indonesia, 1977; Volume 30. [Google Scholar]
- Fageria, N.; Baligar, V.C. Lowland rice response to nitrogen fertilization. Commun. Soil Sci. Plant Anal. 2001, 32, 1405–1429. [Google Scholar] [CrossRef]
- Mohanty, S.R.; Bodelier, P.L.; Floris, V.; Conrad, R.J.A. Differential effects of nitrogenous fertilizers on methane-consuming microbes in rice field and forest soils. Appl. Environ. Microbiol. 2006, 72, 1346–1354. [Google Scholar] [CrossRef] [Green Version]
- Dubey, S.K. Microbial ecology of methane emission in rice agroecosystem: A review. Appl. Ecol. Environ. Res. 2005, 3, 1–27. [Google Scholar] [CrossRef]
- Minamikawa, K.; Sakai, N.; Yagi, K.J.M. Methane emission from paddy fields and its mitigation options on a field scale. Microbes Environ. 2006, 21, 135–147. [Google Scholar] [CrossRef] [Green Version]
- Schimel, J.J.N. Rice, microbes and methane. Nature 2000, 403, 375–377. [Google Scholar] [CrossRef]
- Namai, S.; Toriyama, K.; Fukuta, Y. Genetic variations in dry matter production and physiological nitrogen use efficiency in rice (Oryza sativa L.) varieties. Breed. Sci. 2009, 59, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Fei, W.; Peng, S.-B. Yield potential and nitrogen use efficiency of China’s super rice. J. Integr. Agric. 2017, 16, 1000–1008. [Google Scholar] [CrossRef] [Green Version]
- Raun, W.R.; Johnson, G.V. Improving nitrogen use efficiency for cereal production. Agron. J. 1999, 91, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Chen, Y.; Zhao, G.; Cheng, W.; Guo, S.; Zhang, H.; Shi, W. Do high nitrogen use efficiency rice cultivars reduce nitrogen losses from paddy fields? Agric. Ecosyst. Environ. 2015, 209, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Pires, M.V.; da Cunha, D.A.; de Matos Carlos, S.; Costa, M.H. Nitrogen-use efficiency, nitrous oxide emissions, and cereal production in Brazil: Current trends and forecasts. PLoS ONE 2015, 10, e0135234. [Google Scholar] [CrossRef] [Green Version]
- Neue, H.-U.; Sass, R.L. Trace gas emissions from rice fields. In Global Atmospheric-Biospheric Chemistry; Springer: Berlin/Heidelberg, Germany, 1994; pp. 119–147. [Google Scholar] [CrossRef]
- Yasuno, N.; Takamure, I.; Kidou, S.; Tokuji, Y.; Ureshi, A.; Funabiki, A.; Ashikaga, K.; Yamanouchi, U.; Yano, M.; Kato, K. Rice shoot branching requires an ATP-binding cassette subfamily G protein. New Phytol. 2009, 182, 91–101. [Google Scholar] [CrossRef]
- Ariyaratne, M.; Takamure, I.; Kato, K. Shoot branching control by reduced culm number 4 in rice (Oryza sativa L.). New Phytol. 2009, 176, 744–748. [Google Scholar] [CrossRef]
- Leyser, O. The control of shoot branching: An example of plant information processing. Plant Cell Environ. 2009, 32, 694–703. [Google Scholar] [CrossRef]
- Yaish, M.; Guevara, D.; El-Kereamy, A.; Rothstein, S. Axillary shoot branching in plants. In Plant Developmental Biology-Biotechnological Perspectives; Springer: Berlin/Heidelberg, Germany, 2010; pp. 37–52. [Google Scholar]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.J.N. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pagès, V.; Dun, E.A.; Pillot, J.-P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.-C.J.N. Strigolactone inhibition of shoot branching. Nature 2008, 455, 189–194. [Google Scholar] [CrossRef]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An update on the signals controlling shoot branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef]
- Peleg, Z.; Blumwald, E. Hormone balance and abiotic stress tolerance in crop plants. Curr. Opin. Plant Biol. 2011, 14, 290–295. [Google Scholar] [CrossRef]
- Liu, G.; Li, M.; Wen, J.; Du, Y.; Zhang, Y.-M. Functional mapping of quantitative trait loci associated with rice tillering. Mol. Genet. Genom. 2010, 284, 263–271. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, T.; Wang, M.; Liu, Y.; Yuan, S.; Gao, Y.; Yin, L.; Sun, W.; Peng, L.; Zhang, W.J.P.; et al. DWARF3 participates in an SCF complex and associates with DWARF14 to suppress rice shoot branching. Plant Cell Physiol. 2014, 55, 1096–1109. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Shang, L.; Yu, H.; Zeng, L.; Hu, J.; Ni, S.; Rao, Y.; Li, S.; Chu, J.; Meng, X.J.; et al. A strigolactone biosynthesis gene contributed to the green revolution in rice. Mol. Plant 2020, 13, 923–932. [Google Scholar] [CrossRef]
- Gao, Z.; Qian, Q.; Liu, X.; Yan, M.; Feng, Q.; Dong, G.; Liu, J.; Han, B. Dwarf 88, a novel putative esterase gene affecting architecture of rice plant. Plant Mol. Biol. 2009, 71, 265–276. [Google Scholar] [CrossRef]
- Liu, W.; Wu, C.; Fu, Y.; Hu, G.; Si, H.; Zhu, L.; Luan, W.; He, Z.; Sun, Z.J.P. Identification and characterization of HTD2: A novel gene negatively regulating tiller bud outgrowth in rice. Planta 2009, 230, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, K.P.; Vishwakarma, C.; Sahoo, S.P.; Lima, J.M.; Nath, M.; Dokku, P.; Gacche, R.N.; Mohapatra, T.; Robin, S.; Sarla, N.; et al. A substitution mutation in OsCCD7 cosegregates with dwarf and increased tillering phenotype in rice. J. Genet. 2014, 93, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Lo, S.-F.; Yang, S.-Y.; Chen, K.-T.; Hsing, Y.-I.; Zeevaart, J.A.; Chen, L.-J.; Yu, S.-M. A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.Q.; Wang, J.G.; Wang, L.Y.; Wang, J.F.; Wang, Q.; Yu, P.; Bai, M.Y.; Fan, M. Gibberellin repression of axillary bud formation in Arabidopsis by modulation of DELLA-SPL9 complex activity. J. Integr. Plant Biol. 2020, 62, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Wang, Q.; Chen, P.; Yin, C.; Lin, Y. Strigolactones regulate shoot elongation by mediating gibberellin metabolism and signaling in rice (Oryza sativa L.). J. Plant Physiol. 2019, 237, 72–79. [Google Scholar] [CrossRef]
- Wang, M.; Le Moigne, M.-A.; Bertheloot, J.; Crespel, L.; Perez-Garcia, M.-D.; Ogé, L.; Demotes-Mainard, S.; Hamama, L.; Davière, J.-M.; Sakr, S. BRANCHED1: A key hub of shoot branching. Front. Plant Sci. 2019, 10, 76. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, K.; Maekawa, M.; Ujiie, S.; Satake, Y.; Furutani, I.; Okamoto, H.; Shimamoto, K.; Kyozuka, J. LAX and SPA: Major regulators of shoot branching in rice. Proc. Natl. Acad. Sci. USA 2003, 100, 11765–11770. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Ikegami, M.; Kuze, J.; Sawada, K.; Hashimoto, Z.; Ishii, T.; Nakamura, C.; Kamijima, O. QTL analysis for plant and grain characters of sake-brewing rice using a doubled haploid population. Breed. Sci. 2002, 52, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Zhu, J.; He, C.; Benmoussa, M.; Wu, P. Quantitative trait loci analysis for the developmental behavior of tiller number in rice (Oryza sativa L.). Theor. Appl. Genet. 1998, 97, 267–274. [Google Scholar] [CrossRef]
- Onishi, K.; Horiuchi, Y.; Ishigoh-Oka, N.; Takagi, K.; Ichikawa, N.; Maruoka, M.; Sano, Y. A QTL cluster for plant architecture and its ecological significance in Asian wild rice. Breed. Sci. 2007, 57, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Khush, G.S. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol. 1997, 35, 25–34. [Google Scholar] [CrossRef]
- Zhu, Q.; Ge, S. Phylogenetic relationships among A-genome species of the genus Oryza revealed by intron sequences of four nuclear genes. New Phytol. 2005, 167, 249–265. [Google Scholar] [CrossRef]
- Kato, S. On the affinity of rice varieties as shown by fertility of hybrid plants. Bull. Sci. Fac. Agric. Kyushu Univ. 1928, 3, 132–147. [Google Scholar]
- Park, H.J.; Kim, S.U.; Jung, K.Y.; Lee, S.; Choi, Y.D.; Owens, V.N.; Kumar, S.; Yun, S.W.; Hong, C.O. Cadmium phytoavailability from 1976 through 2016: Changes in soil amended with phosphate fertilizer and compost. Sci. Total Environ. 2021, 762, 143132. [Google Scholar] [CrossRef]
- Thorpe, R.S. Multiple group principal component analysis and population differentiation. J. Zool. 1988, 216, 37–40. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Kang, J.-W.; Kabange, N.R.; Phyo, Z.; Park, S.-Y.; Lee, S.-M.; Lee, J.-Y.; Shin, D.; Cho, J.H.; Park, D.-S.; Ko, J.-M. Combined Linkage Mapping and Genome-Wide Association Study Identified QTLs Associated with Grain Shape and Weight in Rice (Oryza sativa L.). Agronomy 2020, 10, 1532. [Google Scholar] [CrossRef]
- Cheon, K.-S.; Baek, J.; Cho, Y.-I.; Jeong, Y.-M.; Lee, Y.-Y.; Oh, J.; Won, Y.J.; Kang, D.-Y.; Oh, H.; Kim, S.L.; et al. Single nucleotide polymorphism (SNP) discovery and kompetitive allele-specific PCR (KASP) marker development with Korean japonica rice varieties. Plant Breed. Biotechnol. 2018, 6, 391–403. [Google Scholar] [CrossRef]
- Seo, J.; Lee, G.; Jin, Z.; Kim, B.; Chin, J.H.; Koh, H.-J. Development and application of indica–japonica SNP assays using the Fluidigm platform for rice genetic analysis and molecular breeding. Mol. Breed. 2020, 40, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop. J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Nada, R.M.; Abogadallah, G.M. Restricting the above ground sink corrects the root/shoot ratio and substantially boosts the yield potential per panicle in field-grown rice (Oryza sativa L.). Physiol. Plant. 2016, 156, 371–386. [Google Scholar] [CrossRef]
- Andrews, M.; Sprent, J.; Raven, J.; Eady, P.E. Relationships between shoot to root ratio, growth and leaf soluble protein concentration of Pisum sativum, Phaseolus vulgaris and Triticum aestivum under different nutrient deficiencies. Plant Cell Environ. 1999, 22, 949–958. [Google Scholar] [CrossRef]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity in rice seedlings. Acta Physiol. Plant. 2015, 37, 9. [Google Scholar] [CrossRef]
- Pysh, L.D.; Wysocka-Diller, J.W.; Camilleri, C.; Bouchez, D.; Benfey, P.N. The GRAS gene family in Arabidopsis: Sequence characterization and basic expression analysis of the SCARECROW-LIKE genes. Plant J. 1999, 18, 111–119. [Google Scholar] [CrossRef]
- Passaia, G.; Fonini, L.S.; Caverzan, A.; Jardim-Messeder, D.; Christoff, A.P.; Gaeta, M.L.; de Araujo Mariath, J.E.; Margis, R.; Margis-Pinheiro, M. The mitochondrial glutathione peroxidase GPX3 is essential for H2O2 homeostasis and root and shoot development in rice. Plant Sci. 2013, 208, 93–101. [Google Scholar] [CrossRef]
- Paiva, A.L.S.; Passaia, G.; Jardim-Messeder, D.; Nogueira, F.C.; Domont, G.B.; Margis-Pinheiro, M. The mitochondrial isoform glutathione peroxidase 3 (OsGPX3) is involved in ABA responses in rice plants. J. Proteom. 2021, 232, 104029. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-C.; Kim, S.-H.; Kim, S.-R. Drought inducible OsDhn1 promoter is activated by OsDREB1A and OsDREB1D. J. Plant Biol. 2013, 56, 115–121. [Google Scholar] [CrossRef]
- Lee, S.-C.; Lee, M.-Y.; Kim, S.-J.; Jun, S.-H.; An, G.; Kim, S.-R. Characterization of an abiotic stress-inducible dehydrin gene, OsDhn1, in rice (Oryza sativa L.). Mol. Cells 2005, 19, 212–218. [Google Scholar]
- Sharma, A.; Hussain, A.; Mun, B.-G.; Imran, Q.M.; Falak, N.; Lee, S.-U.; Kim, J.Y.; Hong, J.K.; Loake, G.J.; Ali, A.; et al. Comprehensive analysis of plant rapid alkalization factor (RALF) genes. Plant Physiol. Biochem. 2016, 106, 82–90. [Google Scholar] [CrossRef]
- Nietzsche, M.; Schießl, I.; Börnke, F. The complex becomes more complex: Protein-protein interactions of SnRK1 with DUF581 family proteins provide a framework for cell-and stimulus type-specific SnRK1 signaling in plants. Front. Plant Sci. 2014, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Van Dijk, A.D.; Scaffidi, A.; Flematti, G.R.; Hofmann, M.; Charnikhova, T.; Verstappen, F.; Hepworth, J.; Van Der Krol, S.; Leyser, O.; et al. Rice cytochrome P450 MAX1 homologs catalyze distinct steps in strigolactone biosynthesis. Nat. Chem. Biol. 2014, 10, 1028–1033. [Google Scholar] [CrossRef]
- Luo, A.; Qian, Q.; Yin, H.; Liu, X.; Yin, C.; Lan, Y.; Tang, J.; Tang, Z.; Cao, S.; Wang, X.J.P.; et al. EUI1, encoding a putative cytochrome P450 monooxygenase, regulates internode elongation by modulating gibberellin responses in rice. Plant Cell Physiol. 2006, 47, 181–191. [Google Scholar] [CrossRef]
- Deng, L.; Li, L.; Zhang, S.; Shen, J.; Li, S.; Hu, S.; Peng, Q.; Xiao, J.; Wu, C. Suppressor of rid1 (SID1) shares common targets with RID1 on florigen genes to initiate floral transition in rice. PLoS Genet. 2017, 13, e1006642. [Google Scholar] [CrossRef]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. Ann. Rev. Plant Biol. 2010, 61, 421–442. [Google Scholar] [CrossRef]
- Zhen, X.; Li, X.; Yu, J.; Xu, F. OsATG8c-mediated increased autophagy regulates the yield and nitrogen use efficiency in rice. Int. J. Mol. Sci. 2019, 20, 4956. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, P.; Ramakrishnan, B.; Reddy, K.R.; Reddy, V. High-temperature effects on rice growth, yield, and grain quality. Adv. Agron. 2011, 111, 87–206. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, J.; Ren, T.; Hussain, S.; Guo, C.; Wang, S.; Cong, R.; Li, X. Effects of nitrogen and tiller type on grain yield and physiological responses in rice. AoB Plants 2017, 9. [Google Scholar] [CrossRef]
- Zhang, J.; Tong, T.; Potcho, P.M.; Huang, S.; Ma, L.; Tang, X.J.A. Nitrogen Effects on Yield, Quality and Physiological Characteristics of Giant Rice. Agronomy 2020, 10, 1816. [Google Scholar] [CrossRef]
- Tanaka, A.; Garcia, C. Studies of the relationship between tillering and nitrogen uptake of the rice plant: 2. Relation between tillering and nitrogen metabolism of the plant. Soil Sci. Plant Nutr. 1965, 11, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Tian, G.; Gao, L.; Kong, Y.; Hu, X.; Xie, K.; Zhang, R.; Ling, N.; Shen, Q.; Guo, S. Improving rice population productivity by reducing nitrogen rate and increasing plant density. PLoS ONE 2017, 12, e0182310. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Shan, S.; Cao, J.; Fang, S.; Tian, A.; Liu, Y.; Cao, F.; Yin, X.; Zou, Y. Primary-tiller panicle number is critical to achieving high grain yields in machine-transplanted hybrid rice. Sci. Rep. 2020, 10, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Badshah, M.; Naimei, T.; Zou, Y.; Ibrahim, M.; Wang, K. Yield and tillering response of super hybrid rice Liangyoupeijiu to tillage and establishment methods. Crop. J. 2014, 2, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lu, K.; Nie, H.; Zeng, Q.; Wu, B.; Qian, J.; Fang, Z.J.R. Rice nitrate transporter OsNPF7.2 positively regulates tiller number and grain yield. Rice 2018, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, H.; Jiang, Z.; Wang, W.; Xu, R.; Wang, Q.; Zhang, Z.; Li, A.; Liang, Y.; Ou, S.J.N. Genomic basis of geographical adaptation to soil nitrogen in rice. Nature 2021, 1–6. [Google Scholar] [CrossRef]
- Liao, Z.; Yu, H.; Duan, J.; Yuan, K.; Yu, C.; Meng, X.; Kou, L.; Chen, M.; Jing, Y.; Liu, G.; et al. SLR1 inhibits MOC1 degradation to coordinate tiller number and plant height in rice. Nat. Commun. 2019, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lacchini, E.; Kiegle, E.; Castellani, M.; Adam, H.; Jouannic, S.; Gregis, V.; Kater, M.M. CRISPR-mediated accelerated domestication of African rice landraces. PLoS ONE 2020, 15, e0229782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.; Sun, L.; Chen, Y.; Xue, P.; Yang, Q.; Wang, B.; Yu, N.; Cao, Y.; Zhang, Y.; Gong, K.; et al. Rice dwarf and low tillering 10 (OsDLT10) regulates tiller number by monitoring auxin homeostasis. Plant Sci. 2020, 297, 110502. [Google Scholar] [CrossRef] [PubMed]
- Groff, P.A.; Kaplan, D.R. The relation of root systems to shoot systems in vascular plants. Bot. Rev. 1988, 54, 387–422. [Google Scholar] [CrossRef]
- Qi, Y.; Wei, W.; Chen, C.; Chen, L. Plant root-shoot biomass allocation over diverse biomes: A global synthesis. Glob. Ecol. Conserv. 2019, 18, e00606. [Google Scholar] [CrossRef]
- Yano, M.; Sasaki, T. Genetic and molecular dissection of quantitative traits in rice. In Oryza: From Molecule to Plant; Springer: Berlin/Heidelberg, Germany, 1997; pp. 145–153. [Google Scholar] [CrossRef]
- Yamamoto, T.; Yonemaru, J.; Yano, M. Towards the understanding of complex traits in rice: Substantially or superficially? DNA Res. 2009, 16, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Zhu, H.; Zhang, G.; Li, L.; Ye, G. Dynamic analysis of QTLs on tiller number in rice (Oryza sativa L.) with single segment substitution lines. Theor. Appl. Genet. 2012, 125, 143–153. [Google Scholar] [CrossRef]
- Zhu, J.; Zhou, Y.; Liu, Y.; Wang, Z.; Tang, Z.; Yi, C.; Tang, S.; Gu, M.; Liang, G. Fine mapping of a major QTL controlling panicle number in rice. Mol. Breed. 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Bolle, C. The role of GRAS proteins in plant signal transduction and development. Planta 2004, 218, 683–692. [Google Scholar] [CrossRef]
- Xu, K.; Chen, S.; Li, T.; Ma, X.; Liang, X.; Ding, X.; Liu, H.; Luo, L. OsGRAS23, a rice GRAS transcription factor gene, is involved in drought stress response through regulating expression of stress-responsive genes. BMC Plant Biol. 2015, 15, 141. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yang, Z.; Wu, D.; Yu, F. RALF-FERONIA signaling: Linking plant immune response with cell growth. Plant Commun. 2020, 100084. [Google Scholar] [CrossRef]
- Chaban, C.; Waller, F.; Furuya, M.; Nick, P. Auxin responsiveness of a novel cytochrome P450 in rice coleoptiles. Plant Physiol. 2003, 133, 2000–2009. [Google Scholar] [CrossRef] [Green Version]
- Marzec, M.; Situmorang, A.; Brewer, P.B.; Brąszewska, A.J.G. Diverse roles of MAX1 homologues in rice. Genes 2020, 11, 1348. [Google Scholar] [CrossRef]
- Itoh, H.; Shimada, A.; Ueguchi-Tanaka, M.; Kamiya, N.; Hasegawa, Y.; Ashikari, M.; Matsuoka, M. Overexpression of a GRAS protein lacking the DELLA domain confers altered gibberellin responses in rice. Plant J. 2005, 44, 669–679. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, S.; Zhang, W.; Li, G.; Chen, Z.; Zhai, W.; Zhao, X.; Pan, X.; Xie, Q.; Zhu, L. The rice HIGH-TILLERING DWARF1 encoding an ortholog of Arabidopsis MAX3 is required for negative regulation of the outgrowth of axillary buds. Plant J. 2006, 48, 687–698. [Google Scholar] [CrossRef]
- Zou, J.; Chen, Z.; Zhang, S.; Zhang, W.; Jiang, G.; Zhao, X.; Zhai, W.; Pan, X.; Zhu, L. Characterizations and fine mapping of a mutant gene for high tillering and dwarf in rice (Oryza sativa L.). Planta 2005, 222, 604–612. [Google Scholar] [CrossRef]
- Rolly, N.K.; Mun, B.-G.; Yun, B.-W. Insights into the Transcriptional Regulation of Branching Hormonal Signaling Pathways Genes under Drought Stress in Arabidopsis. Genes 2021, 12, 298. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field | pH | TN (%) | EC | OM (%) | P2O5 (mg kg−1) | K+ (cmolc kg−1) | Ca2+ (cmolc kg−1) | Mg2+ (cmolc kg−1) | Na+ (cmolc kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| Low N | 5.5 | 0.27 | 0.75 | 2.74 | 44.77 | 0.84 | 4.76 | 1.01 | 0.18 |
| Normal N | 5.7 | 1.38 | 1.38 | 3.79 | 96.87 | 0.27 | 1.17 | 7.82 | 1.27 |
| Traits (a) | QTL (b) | Chr.(c) | Position (cM) (d) | Left Marker (e) | Right Marker (f) | LOD (g) | PVE (%) (h) | Add (i) |
|---|---|---|---|---|---|---|---|---|
| PHN | qPHN1-1 | 1 | 43 | Os01_id1028304 | Os01_ad01020824 | 4.703 | 8.745 | 2.08 |
| qPHN3-1 | 3 | 166 | Os03_Hd6-1AT | Os03_id3015453 | 4.883 | 8.798 | −2.1 | |
| qPHN5-1 | 5 | 23 | Os05_KJ05_019 | Os05_KJ05_017 | 5.694 | 9.915 | −2.2 | |
| qPHN10-1 | 10 | 1 | Os10_id10002069 | Os10_KJ10_003 | 5.305 | 9.147 | 2.14 | |
| qPHN12-1 | 12 | 130 | Os12_cmb1224_0 | Os12_cmb1226_0 | 5.134 | 8.836 | −2 | |
| TNL | qTNL2-1 | 2 | 134 | Os02_KJ02_057 | Os02_id2012773 | 6.052 | 20.45 | 1.81 |
| TNN | qTNN2-1 | 2 | 130 | Os02_KJ02_053 | Os02_KJ02_057 | 4.141 | 14.51 | 1.25 |
| qTNN4-1 | 4 | 173 | Os04_cmb0432_2 | Os04_cmb0434_1 | 3.922 | 14.49 | 1.33 | |
| CLL | qCLL1-1 | 1 | 108 | Os01_id1015984 | Os01_ah01001843 | 3.947 | 10.57 | 3.3 |
| qCLL9-1 | 9 | 72 | Os09_KJ09_075 | Os09_TAC1_CT | 4.632 | 11.59 | −2.8 | |
| CLN | qCLN2-1 | 2 | 132 | Os02_KJ02_057 | Os02_id2012773 | 4.42 | 6.509 | −1.9 |
| qCLN2-2 | 2 | 187 | Os02_id2014575 | Os02_cmb0236_6 | 13.45 | 24.81 | 3.77 | |
| qCLN3-1 | 3 | 128 | Os03_id3010700 | Os03_ad03013905 | 11.45 | 19.51 | 3.37 | |
| qCLN9-1 | 9 | 9 | Os09_cmb0909_6 | Os09_KJ09_027 | 4.556 | 6.805 | −2.2 | |
| qCLN10-1 | 10 | 17 | Os10_KJ10_003 | Os10_KJ10_019 | 3.178 | 5.401 | 1.89 | |
| PLN | qPLN2-1 | 2 | 74 | Os02_id2002293 | Os02_KJ02_009 | 4.811 | 7.985 | −0.7 |
| qPLN3-1 | 3 | 123 | Os03_id3010700 | Os03_ad03013905 | 3.917 | 6.768 | 0.61 | |
| qPLN3-2 | 3 | 201 | Os03_ah03002520 | Os03_cmb0336_5 | 8.844 | 16.06 | 0.93 | |
| qPLN7-1 | 7 | 89 | Os07_KJ07_021 | Os07_id7001155 | 5.938 | 11.14 | 0.78 | |
| qPLN8-1 | 8 | 101 | Os08_GW8-AG | Os08_id8007764 | 8.059 | 14.35 | −0.9 | |
| PNL | qPNL4-1 | 4 | 154 | Os04_ad04009559 | Os04_ah04001252 | 2.954 | 11.99 | 1.18 |
| PNN | qPNN2-1 | 2 | 53 | Os02_id2000007 | Os02_ad02000512 | 3.487 | 7.269 | 0.63 |
| qPNN4-1 | 4 | 169 | Os04_id4009823 | Os04_cmb0432_2 | 9.633 | 23.76 | 0.94 | |
| qPNN8-1 | 8 | 84 | Os08_ae08007378 | Os08_id8006751 | 4.34 | 9.338 | 0.55 | |
| RLL | qRLL4-1 | 4 | 59 | Os04_KJ04_053 | Os04_KJ04_057 | 3.241 | 11.99 | −1.1 |
| SDWN | qSDWN5-1 | 5 | 46 | Os05_KJ05_011 | Os05_id5002497 | 3.748 | 12.88 | −0.6 |
| RDWN | qRDWN1-1 | 1 | 27 | Os01_KJ01_125 | Os01_KJ01_127 | 4.069 | 11.74 | −0.4 |
| qRDWN4-1 | 4 | 21 | Os04_KJ04_029 | Os04_KJ04_047 | 3.218 | 8.654 | −0.3 |
| No. | Gene Name | Locus | Annotation | Molecular Function | Cellular Component |
|---|---|---|---|---|---|
| 1 | OsPBF19 | Os02g44220 | Peroxisomal biogenesis factor 19, putative, expressed | Protein binding | Cytosol; peroxisome |
| 2 | OsSc11/ | Os02g44360 | Scarecrow transcription factor family protein, putative, expressed; GRAS: GIBBERELLIN-INSENSITIVE (GAI), REPRESSOR of GA1-3 (RGA), and SCARECROW (SCR) | Sequence-specific DNA binding transcription factor activity | Plastid, nucleus |
| 3 | OsGRAS | Os02g44370 | Scarecrow, putative, expressed; GRAS transcription factor domain containing protein NSENSITIVE (GAI), REPRESSOR of GA1-3 (RGA), and SCARECROW (SCR) | Sequence-specific DNA binding transcription factor activity | Plastid, nucleus |
| 4 | OsGpx3 | Os02g44500 | Glutathione peroxidase, putative, expressed; OsGpx3|GPX3|OsGPx03 | Glutathione peroxidase activity; peroxidase activity; oxidoreductase activity | Mitochondrion; plasma membrane; plastid; cytosol; chloroplast; |
| 5 | OsHUGT1 | Os02g44510 | UDP-glucose glycoprotein glucosyltransferase 1 precursor, putative, expressed | UDP-glucose:glycoprotein glucosyltransferase activity | Endoplasmic reticulum |
| 6 | OsSub19 | Os02g44520 | Putative Subtilisin homologue, expressed; Peptidase S8 and S53, subtilisin, kexin, sedolisin domain containing protein | Hydrolase activity; serine-type endopeptidase activity | Vacuole; cytoplasm; ribosome; cytosol; plastid; vacuolar membrane; chloroplast |
| 7 | OsMalic | Os02g44550 | NADP-dependent malic enzyme, putative, expressed | Catalytic activity; malic enzyme activity; oxidoreductase activity, acting on the CH-OH group of donors | Cytosol |
| 8 | Os02g44570 | Mitochondrial carrier protein, putative, expressed; Mitochondrial substrate carrier family protein | binding | Mitochondrial inner membrane | |
| 9 | OsPIP1;1|RWC1 | Os02g44630 | Aquaporin. Plasma membrane intrinsic proteins subfamily | Transporter activity | Plasma membrane; mitochondrion; chloroplast envelop |
| 10 | MAP3K.10—STE | Os02g44642 | STE_MEKK_ste11_MAP3K.10—STE kinases include homologs to sterile 7, sterile 11 and sterile 20 from yeast, expressed; Similar to MAP3Ka | Transferring phosphorus-containing groups; ATP binding; protein kinase activity | |
| 11 | CYP450 | Os02g44654 | Cytochrome P450, putative, expressed; Similar to Cytochrome P450 | Heme binding; oxidoreductase activity | |
| 12 | Os02g44700 | Zinc finger, C3HC4 type domain containing protein, expressed; Zinc finger, RING-type domain containing protein | Binding; catalytic activity; zinc ion binding; ubiquitin–protein transferase activity; metal ion binding; protein binding | ||
| 13 | OsDhn1/OsLip9 | Os02g44870 | Dehydrin, putative, expressed; Dehydrin family protein | Binding | Cytosol; plasma membrane; plastid |
| 14 | OsHMG1 | Os02g44930 | HMG1/2, putative, expressed; Similar to HMGc1 protein | Sequence-specific DNA binding transcription factor activity; structural molecule activity | Intracellular |
| 15 | OsRALFL8 | Os02g44940 | Rapid Alkalinization Factor RALF family protein precursor, expressed; Rapid Alkalinization Factor family protein | Signal transducer activity | Extracellular region |
| 15 | Os02g44980 | Transmembrane amino acid transporter protein, putative, expressed; Similar to amino acid transport protein | Transporter activity | Membrane; integral component of membrane | |
| 16 | OsFBDUF1/OsMAIF13 | Os02g44990 | F-box and DUF domain containing protein, expressed; Cyclin-like F-box domain containing protein. | Protein binding | |
| 17 | bHLH | Os02g45010 | Ethylene-responsive protein related, putative, expressed; Basic helix-loop-helix dimerization region bHLH domain containing protein | Sequence-specific DNA binding transcription factor activity; protein dimerization activity | Nucleus |
| 18 | SID1|OsIDD4 | Os02g45054 | ZOS2-15—C2H2-type domain containing protein, expressed; Zinc finger | Nucleic acid and protein binding; sequence-specific DNA binding transcription factor activity; metal ion binding | Nucleus |
| 19 | MYB | Os02g45080 | MYB family transcription factor, putative, expressed, containing a myb_SHAQKYF class DNA-binding domain at the C-terminal end of the motif | DNA binding; sequence-specific DNA binding transcription factor activity | |
| 20 | Os02g45130 | Protein kinase, putative, expressed | protein serine/threonine kinase activity; ATP binding | Nucleus | |
| 21 | OsALMT7 | Os02g45160 | Aluminum-activated malate transporter, putative, expressed; Similar to ALMT3 | Malate transport |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, Y.-H.; Kabange, N.-R.; Lee, J.-Y.; Lee, S.-M.; Cha, J.-K.; Shin, D.-J.; Cho, J.-H.; Kang, J.-W.; Ko, J.-M.; Lee, J.-H. Novel QTL Associated with Shoot Branching Identified in Doubled Haploid Rice (Oryza sativa L.) under Low Nitrogen Cultivation. Genes 2021, 12, 745. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050745
Kwon Y-H, Kabange N-R, Lee J-Y, Lee S-M, Cha J-K, Shin D-J, Cho J-H, Kang J-W, Ko J-M, Lee J-H. Novel QTL Associated with Shoot Branching Identified in Doubled Haploid Rice (Oryza sativa L.) under Low Nitrogen Cultivation. Genes. 2021; 12(5):745. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050745
Chicago/Turabian StyleKwon, Young-Ho, Nkulu-Rolly Kabange, Ji-Yun Lee, So-Myeong Lee, Jin-Kyung Cha, Dong-Jin Shin, Jun-Hyeon Cho, Ju-Won Kang, Jong-Min Ko, and Jong-Hee Lee. 2021. "Novel QTL Associated with Shoot Branching Identified in Doubled Haploid Rice (Oryza sativa L.) under Low Nitrogen Cultivation" Genes 12, no. 5: 745. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050745