Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Plant Growth and Biomass Yield
2.2. Mineral Uptake, Pigment Contents, Leaf Relative Water Content (LRWC), and Antioxidant Capacity
2.3. Levels of Proline, Soluble Sugars, Soluble Proteins, Total Free Amino Acids, Phenols, and Flavonoids
2.4. Estimation of Oxidative Stress Markers
2.5. Measurement of Gas-Exchange Parameters
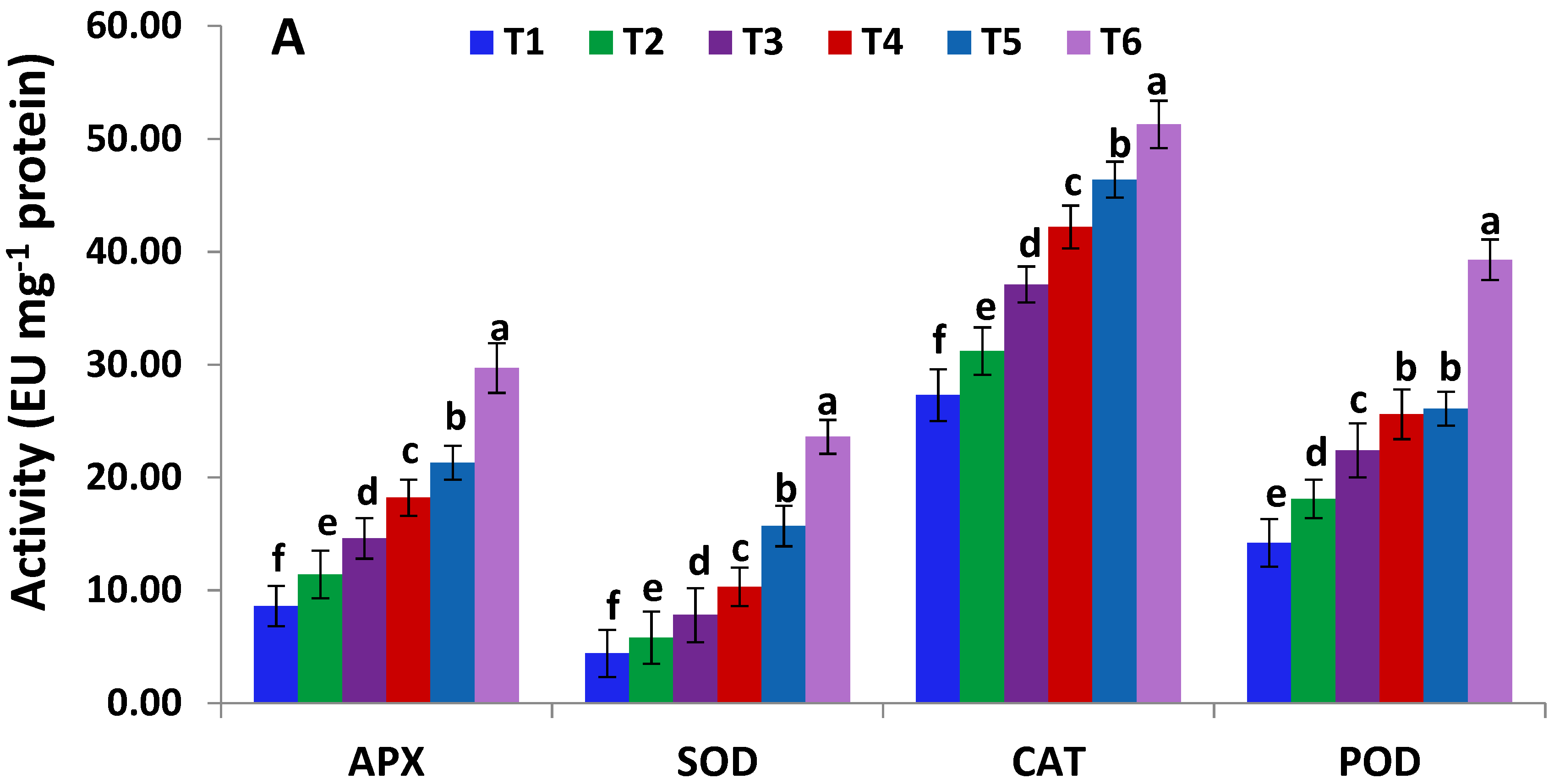
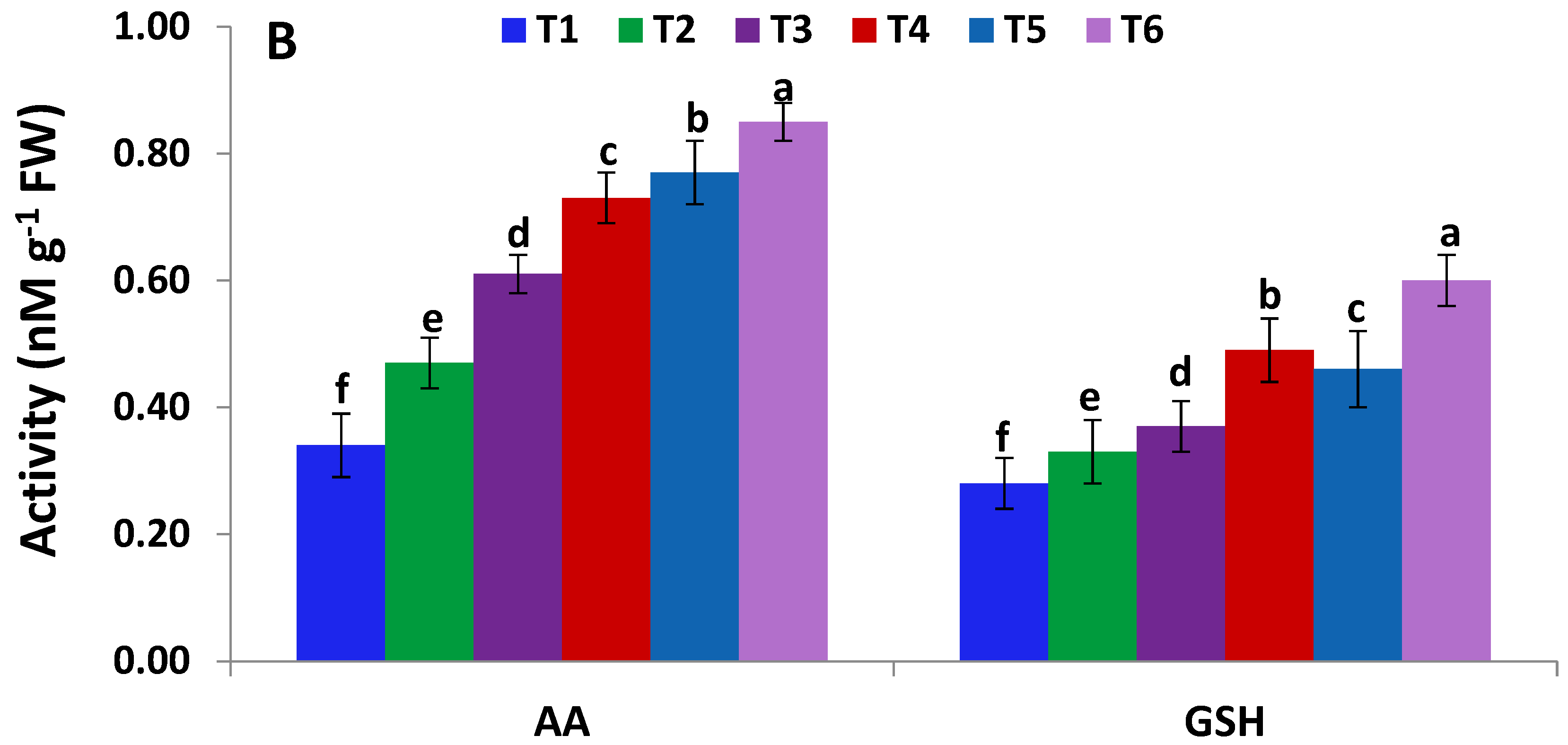
2.6. Activities of Antioxidant Enzymes and Levels of Non-Enzymatic Antioxidants
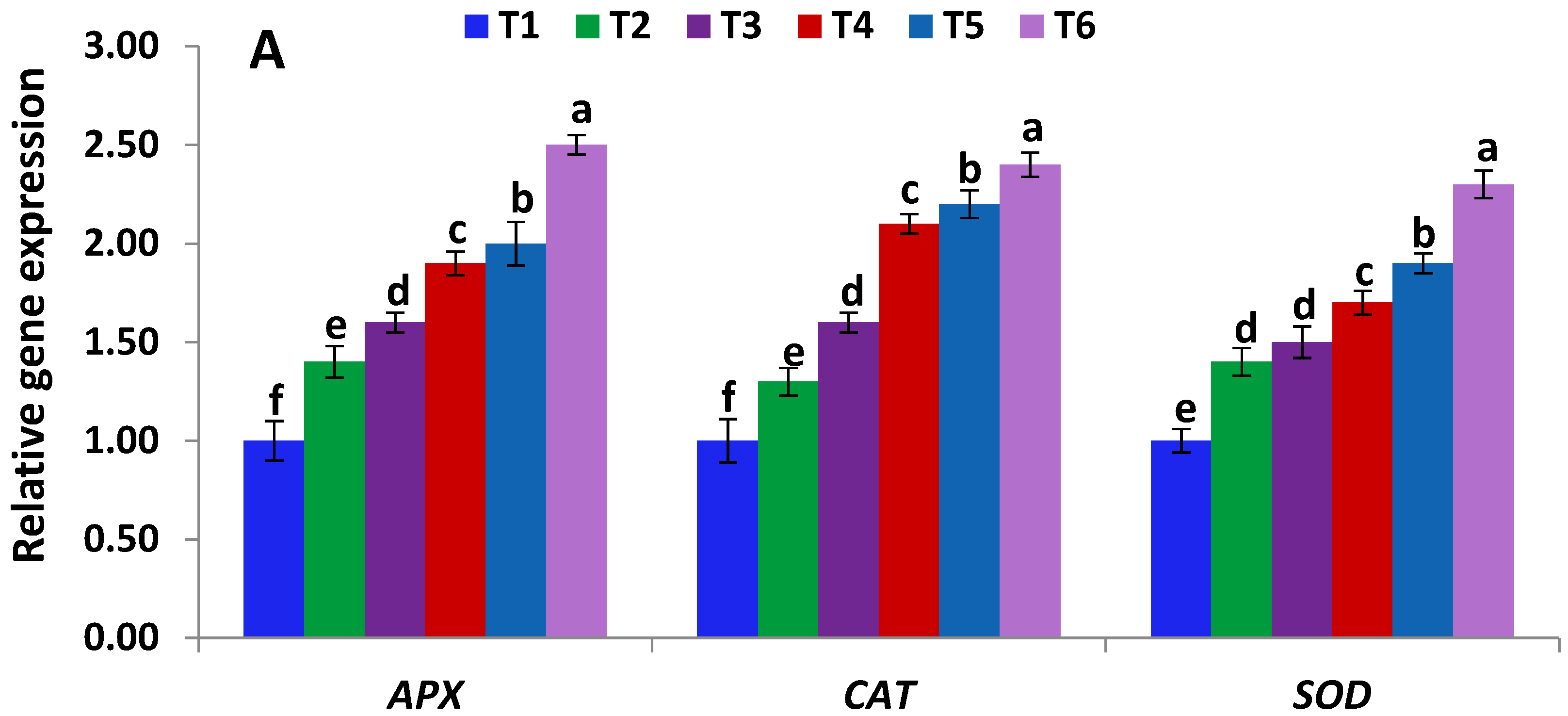
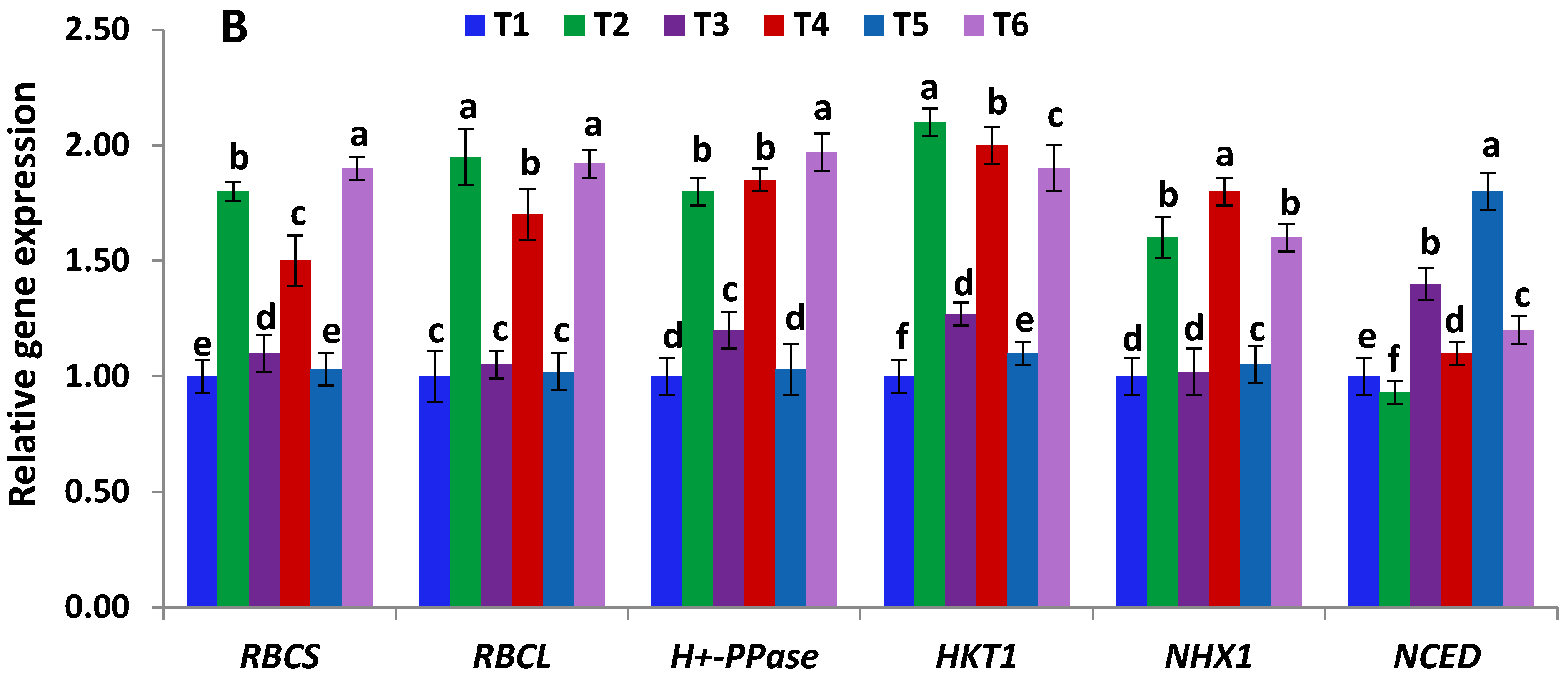
2.7. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Test of Salt Tolerance of Serratia Liquefaciens KM4
4.2. Bacterial Inoculation and Plant Growth
4.3. Estimation of Growth and Biomass Yield
4.4. Estimation of Na+, Cl−, Ca2+, and K+ Content
4.5. Determination of Pigment Contents
4.6. Estimation of Osmoprotectant Contents
4.7. Determination of Phenloic and Flavonoid Contents
4.8. Estimation of Oxidative Stress Markers
4.9. Determination of Antioxidant Capacity
4.10. Determination of LRWC and Gas-Exchange Parameters
4.11. Assays of Antioxidant Enzyme Activities
4.12. Levels of Non-Enzymatic Antioxidants
4.13. Gene Expression Analysis
4.14. Statistical Analysis
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Allakhverdiev, S.I.; Sakamoto, A.; Nishiyama, Y.; Inaba, M.; Murata, N. Ionic and osmotic effects of NaCl induced inactivation of photosystems I and II in Synechococcus sp. Plant Physiol. 2000, 123, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Jha, P.N. Alleviation of salinity-induced damage on wheat plant by an ACC deaminase-producing halophilic bacterium Serratia sp. SL- 12 isolated from a salt lake. Symbiosis 2016, 69, 101–111. [Google Scholar] [CrossRef]
- Li, H.Q.; Jiang, X.W. Inoculation with Plant Growth-Promoting Bacteria (PGPB) Improves Salt Tolerance of Maize Seedling. Russ. J. Plant Physiol. 2017, 64, 235–241. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Pottosin, I.I.; Cuin, T.A.; Fuglsang, A.T.; Tester, M.; Jha, D.; Zepeda-Jazo, I.; Zhou, M.; Palmgren, M.G.; Newman, I.A.; et al. Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley. Plant Physiol. 2007, 145, 1714–1725. [Google Scholar] [CrossRef] [PubMed]
- Islam, F.; Yasmeen, T.; Arif, M.S.; Ali, S.; Ali, B.; Hameed, S.; Zhou, W. Plant growth promoting bacteria confer salt tolerance in Vigna radiata by up-regulating antioxidant defense and biological soil fertility. Plant Growth Regul. 2016, 80, 23–36. [Google Scholar] [CrossRef]
- Shibli, R.A.; Kushad, M.; Yousef, G.G.; Lila, M.A. Physiological and biochemical responses of tomato microshoots to induced salinity stress with associated ethylene accumulation. Plant Growth Regul. 2007, 51, 159–169. [Google Scholar] [CrossRef]
- Zerrouk, I.Z.; Benchabane, M.; Khelifi, L.; Yokawa, K.; Ludwig-Muller, J.; Baluska, F. A Pseudomonas strain isolated from date-palm rhizospheres improves root growth and promotes root formation in maize exposed to salt and aluminum stress. J. Plant Physiol. 2016, 191, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.; Jabborova, D.; Räsänen, L.A.; Liao, H. Coordination between Bradyrhizobium and Pseudomonas alleviates salt stress in soybean through altering root system architecture. J. Plant Interact. 2017, 12, 100–107. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, H.J.; Pak, J.H.; Cho, H.S.; Choi, H.K.; Jung, H.W.; Lee, D.H.; Chung, Y.S. Overexpression of AtSZF2 from Arabidopsis showed enhanced tolerance to salt stress in soybean. Plant Breed. Biotechnol. 2017, 5, 1–15. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Ali, H.M.; Alayafi, A.A.; Witczak, J.; Ahmad, M. Genetic Variation and Alleviation of Salinity Stress in Barley. Molecules 2018, 23, 2488. [Google Scholar] [CrossRef] [PubMed]
- Vwioko, E.; Adinkwu, O.; El-Esawi, M.A. Comparative Physiological, Biochemical and Genetic Responses to Prolonged Waterlogging Stress in Okra and Maize Given Exogenous Ethylene Priming. Front. Physiol. 2017, 8, 632. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A.; Witczak, J.; Ahmad, M. Analysis of Genetic Variation and Enhancement of Salt Tolerance in French Pea. Int. J. Mol. Sci. 2018, 19, 2433. [Google Scholar] [CrossRef] [PubMed]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed Extracts Enhance Salam Turfgrass Performance during Prolonged Irrigation Intervals and Saline Shock. Front. Plant. Sci. 2017, 8, 830. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Sammour, R. Karyological and phylogenetic studies in the genus Lactuca L. Cytologia 2014, 79, 269–275. [Google Scholar] [CrossRef]
- El-Esawi, M.A. Genetic diversity and evolution of Brassica genetic resources: From morphology to novel genomic technologies—A review. Plant Genet. Resour. 2017, 15, 388–399. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Germaine, K.; Bourke, P.; Malone, R. Genetic diversity and population structure of Brassica oleracea germplasm in Ireland using SSR markers. C. R. Biol. 2016, 339, 133–140. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Germaine, K.; Bourke, P.; Malone, R. AFLP analysis of genetic diversity and phylogenetic relationships of Brassica oleracea in Ireland. C. R. Biol. 2016, 339, 163–170. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Mustafa, A.; Badr, S.; Sammour, R. Isozyme analysis of genetic variability and population structure of Lactuca L. germplasm. Biochem. Syst. Ecol. 2017, 70, 73–79. [Google Scholar] [CrossRef]
- El-Esawi, M.A. SSR analysis of genetic diversity and structure of the germplasm of faba bean (Vicia faba L.). C. R. Biol. 2017, 340, 474–480. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Witczak, J.; Abomohra, A.; Ali, H.M.; Elshikh, M.S.; Ahmad, M. Analysis of the Genetic Diversity and Population Structure of Austrian and Belgian Wheat Germplasm Within a Regional Context Based on DArT Markers. Genes 2018, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Elkelish, A.; Elansary, H.O.; Ali, H.M.; Elshikh, M.; Witczak, I.; Ahmad, M. Genetic Transformation and Hairy Root Induction Enhance the Antioxidant Potential of Lactuca serriola L. Oxid. Med. Cell. Long. 2017, 2017, 5604746. [Google Scholar] [CrossRef] [PubMed]
- Sallam, A.; Arbaoui, M.; El-Esawi, M.; Abshire, N.; Martsch, R. Identification and Verification of QTL Associated with Frost Tolerance Using Linkage Mapping and GWAS in Winter Faba Bean. Front. Plant Sci. 2016, 7, 1098. [Google Scholar] [CrossRef] [PubMed]
- Consentino, L.; Lambert, S.; Martino, C.; Jourdan, N.; Bouchet, P.-E.; Witczak, J.; Castello, P.; El-Esawi, M.; Corbineau, F.; d’Harlingue, A.; et al. Blue-light dependent reactive oxygen species formation by Arabidopsis cryptochrome may define a novel evolutionarily conserved signalling mechanism. New Phytol. 2015, 206, 1450–1462. [Google Scholar] [CrossRef] [PubMed]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; Ali, H.M.; Elshikh, M.S.; Abdel-Salam, E.M.; El-Esawi, M.; El-Ansary, D.O. Bioactivities of traditional medicinal plants in Alexandria. Evid. Based Complement. Altern. Med. 2018, 2018, 1463579. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A. Micropropagation technology and its applications for crop improvement. In Plant Tissue Culture: Propagation, Conservation and Crop Improvement; Anis, M., Ahmad, N., Eds.; Springer: Singapore, 2016; pp. 523–545. [Google Scholar]
- El-Esawi, M.A. Nonzygotic embryogenesis for plant development. In Plant Tissue Culture: Propagation, Conservation and Crop Improvement; Anis, M., Ahmad, N., Eds.; Springer: Singapore, 2016; pp. 583–598. [Google Scholar]
- El-Esawi, M.A. Somatic hybridization and microspore culture in Brassica improvement. In Plant Tissue Culture: Propagation, Conservation and Crop Improvement; Anis, M., Ahmad, N., Eds.; Springer: Singapore, 2016; pp. 599–609. [Google Scholar]
- El-Esawi, M.; Arthaut, L.; Jourdan, N.; d’Harlingue, A.; Martino, C.; Ahmad, M. Blue-light induced biosynthesis of ROS contributes to the signaling mechanism of Arabidopsis cryptochrome. Sci. Rep. 2017, 7, 13875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Esawi, M.; Glascoe, A.; Engle, D.; Ritz, T.; Link, J.; Ahmad, M. Cellular metabolites modulate in vivo signaling of Arabidopsis cryptochrome-1. Plant Signal. Behav. 2015, 10, e1063758. [Google Scholar] [CrossRef] [PubMed]
- Abomohra, A.E.F.; Eladel, H.; El-Esawi, M.; Wang, S.; Wang, Q.; He, Z.; Feng, Y.; Shang, H.; Hanelt, D. Effect of lipid-free microalgal biomass and waste glycerol on growth and lipid production of Scenedesmus obliquus: innovative waste recycling for extraordinary lipid production. Bioresour. Technol. 2018, 249, 992–999. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tolerance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems and stress-responsive genes expression. Plant Physiol. Biochem. 2018, 132, 375–384. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.A.; Elansary, H.O.; El-Shanhorey, N.A.; Abdel-Hamid, A.M.E.; Ali, H.M.; Elshikh, M.S. Salicylic Acid-Regulated Antioxidant Mechanisms and Gene Expression Enhance Rosemary Performance under Saline Conditions. Front. Physiol. 2017, 8, 716. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Vardharajula, S.; Zulfikar, A.S.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp., effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Selvakumar, G.; Mohan, M.; Kundu, S.; Gupta, A.D.; Joshi, P.; Nazim, S.; Gupta, H.S. Cold tolerance and plant growth promotion potential of Serratia marcescens strain SRM (MTCC 8708) isolated from flowers of summer squash (Cucurbita pepo). Lett. Appl. Microbiol. 2008, 46, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, U.; Chakraborty, B.N.; Chakraborty, A.P. Influence of Serratia marcescens TRS-1 on growth promotion and induction of resistance in Camellia sinensis against Fomes lamaoensis. J. Plant Interact. 2010, 5, 261–272. [Google Scholar] [CrossRef]
- Devi, K.A.; Pandey, P.; Sharma, G.D. Plant Growth-Promoting Endophyte Serratia marcescens AL2-16 Enhances the Growth of Achyranthes aspera L., a Medicinal Plant. HAYATI J. Biosci. 2016, 23, 173–180. [Google Scholar] [CrossRef]
- Zaheer, A.; Mirza, B.S.; Mclean, J.E.; Yasmin, S.; Shah, T.M.; Malik, K.A.; Mirza, M.S. Association of plant growth-promoting Serratia spp. with the root nodules of chickpea. Res. Microbiol. 2016, 167, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.; Wu, G.; Njeri, K.V.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 2016, 158, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Farinati, S.; DalCrso, G.; Panigati, M.; Furini, A. Interaction between selected bacterial strains and Arabidopsis halleri modulates shoot proteome and cadmium and zinc accumulation. J. Exp. Bot. 2011, 62, 3433–3447. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Zhu, J.; Zhu, J.-K. Salt stress signaling and mechanisms of plant salt tolerance. Genet. Eng. 2006, 27, 141–177. [Google Scholar]
- Zhang, H.; Kim, M.; Sun, Y.; Dowd, S.E.; Shi, H.; Paré, P.W. Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol. Plant Microbe Interact. 2008, 21, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Schilling, R.K.; Marschner, P.; Shavrukov, Y.; Berger, B.; Tester, M.; Roy, S.J.; Plett, D.C. Expression of the Arabidopsis vacuolar H+-pyrophosphatase gene (AVP1) improves the shoot biomass of transgenic barley and increases grain yield in a saline field. Plant Biotechnol. J. 2014, 12, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Plant salt tolerance. Trends Plant Sci. 2001, 6, 66–71. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 969. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kloepper, J.W.; Ryu, C. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Murzello, C.; Sun, Y.; Kim, M.; Xie, X.; Jeter, R.M.; Zak, J.C.; Dowd, S.E.; Paré, P.W. Choline and osmotic stress tolerance induced in Arabidopsis by the soil microbe Bacillus subtilis (GB03). Mol. Plant Microbe Interact. 2010, 23, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Huang, Y.; Zhu, C.; Zeng, H.; Shen, C.; Liu, C.; Zhao, Y.M. Rhizobia inoculation enhances the soybean’s tolerance to salt stress. Plant Soil 2016, 400, 209–222. [Google Scholar] [CrossRef]
- Doganlar, Z.B.; Demir, K.; Basak, H.; Gul, I. Effects of salt stress on pigment and total soluble protein contents of three different tomato cultivars. Afr. J. Agric. Res. 2010, 5, 2056–2065. [Google Scholar]
- Gupta, A.K.; Kaur, N. Sugar signalling and gene expression in relation to carbohydrate metabolism under abiotic stresses in plants. J. Biosci. 2005, 30, 761–776. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Naeem, M. Interactive role of nitric oxide and calcium chloride in enhancing tolerance to salt stress. Nitric Oxide 2012, 27, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Jogaiah, S.; Govind, S.R.; Tran, L.S.P. Systems biology-based approaches toward understanding drought tolerance in food crops. Crit. Rev. Biotechnol. 2013, 33, 23–39. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Singh, J.S.; Saxena, A.K.; Singh, D.P. Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biol. 2012, 14, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Bahadur, A.; Singh, U.P.; Sarma, B.K.; Singh, D.P.; Singh, K.P.; Singh, A. Foliar application of plant growth-promoting rhizobacteria increases antifungal compounds in pea (Pisum sativum) against Erysiphe pisi. Mycobiology 2007, 35, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.Q.; Lu, X.P.; Bai, J.P.; Qiao, Y.; Pare, P.W.; Wang, S.M.; Zhang, J.L.; Wu, Y.N.; Pang, X.P.; Xu, W.P.; Wang, Z.L. Beneficial soil bacterium Bacillus subtilis (GB03) augments salt tolerance of white clover. Front. Plant Sci. 2014, 5, 1928. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Wirth, S.; Egamberdieva, D. The interaction between arbuscular mycorrhizal fungi and endophytic bacteria enhances plant growth of Acacia gerrardii under salt stress. Front. Microbiol. 2016, 7, 1167. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Simultaneous measurement of foliar glutathione, γ-glutamylcysteine, and amino acids by high-performance liquid chromatography: Comparison with two other assay methods for glutathione. Anal. Biochem. 1998, 264, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Azeem, S.A.M. Studies on Plant Growth Promoting Rhizosphere Microorganisms. Ph.D. Thesis, Faculty of Agriculture Suez Canal University, Ismailia, Egypt, 2006. [Google Scholar]
- Abd El-Azeem, S.A.M.; Mehana, T.A.; Shabayek, A.A. Some plant-growth-promoting traits of rhizobacteria isolated from Suez Canal region, Egypt. Afr. Crop Sci. Conf. Proc. 2007, 8, 1517–1525. [Google Scholar]
- Williams, V.; Twine, S. Flame photometric method for sodium, potassium and calcium. In Modern Methods of Plant Analysis; Peach, K., Tracey, M.V., Eds.; Springer: Berlin, Germany, 1960; pp. 3–5. [Google Scholar]
- Wolf, B. A comprehensive system of leaf analysis and its use for diagnosing crop nutrient status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, R.R. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Bates, L.S.; Wladren, P.R.; Tear, D.T. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Emerich, D.W.; Sanchez-DIaz, M. Water stress induced changes in concentrations of proline and total soluble sugar in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities f protein utilizing the principle of protein binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lee, Y.P.; Takanashi, T. An improved colorimetric determination of amino acids with the use of ninhydrin. Anal. Biochem. 1966, 14, 71–77. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, J.; Shen, J.; Silva, A.; Dennis, D.; Barrow, C. A simple 96-well microplate method for estimation of total polyphenol content in seaweeds. J. Appl. Phycol. 2006, 18, 445–450. [Google Scholar] [CrossRef]
- Chang, C.; Yang, M.; Wen, H.; Chern, J. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 178–182. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Rao, K.V.M.; Sresty, T.V.S. Antioxidative parameters in the seedlings of pigeon pea (Cajanus cajan L. Millspaugh) in response to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Pyrzynska, K.; Pękal, A. Application of free radical diphenylpicrylhydrazyl (DPPH) to estimate the antioxidant capacity of food samples. Anal. Methods 2013, 5, 4288–4295. [Google Scholar] [CrossRef]
- Garíca-Mata, C.; Lamattina, L. Nitric oxide induces stomatal closure and enhances the adaptive plant responses against drought stress. Plant Physiol. 2001, 126, 1196–1204. [Google Scholar] [CrossRef]
- Holá, D.; Benešová, M.; Honnerová, J.; Hnilička, F.; Rothová, O.; Kočová, M.; Hniličková, H. The evaluation of photosynthetic parameters in maize inbred lines subjected to water deficiency: Can these parameters be used for the prediction of performance of hybrid progeny? Photosynthetica 2010, 48, 545–558. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, M.B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol. 1996, 132, 361–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide-induces chilling tolerance in mung beans mediated through ABA-independent glutathione accumulation. Funct. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NaCl (mM) | Serratia liquefaciens KM4 | Shoot Length (cm) | Shoot Fresh Weight (g·plant−1) | Shoot Dry Weight (g·plant−1) | Root Length (cm) | Root Fresh Weight (g·plant−1) | Root Dry Weight (g·plant−1) |
|---|---|---|---|---|---|---|---|
| 0 | −KM4 | 32.4 ± 1.88 b | 5.9 ± 0.41 b | 0.58 ± 0.12 b | 20.5 ± 1.21 b | 2.71 ± 0.19 b | 0.38 ± 0.16 b |
| +KM4 | 35.7 ± 1.45 a | 6.4 ± 0.35 a | 0.63 ± 0.14 a | 22.9 ± 1.32 a | 2.98 ± 0.18 a | 0.43 ± 0.15 a | |
| 80 | −KM4 | 26.2 ± 1.29 d | 5.1 ± 0.38 c | 0.52 ±0.23 d | 17.6 ± 1.21 d | 2.32 ± 0.22 d | 0.32 ±0.17 d |
| +KM4 | 30.1 ± 1.17 c | 5.4 ± 0.41 b | 0.55 ± 0.16 c | 19.2 ± 1.17 c | 2.59 ± 0.18 c | 0.36 ± 0.15 c | |
| 160 | −KM4 | 22.4 ± 1.11 f | 3.9 ± 0.27 e | 0.42 ± 0.18 f | 14.3 ± 1.15 f | 1.98 ± 0.15 f | 0.23 ± 0.12 f |
| +KM4 | 24.3 ± 1.25 e | 4.3 ± 0.31 d | 0.46 ± 0.15 e | 16.6 ± 1.24 e | 2.22 ± 0.17 e | 0.27 ± 0.11 e |
| NaCl (mM) | S. liquefaciens KM4 | Na+ (mg g−1 DW) | Cl− (mg g−1 DW) | Ca2+ (mg g−1 DW) | K+ (mg g−1 DW) |
|---|---|---|---|---|---|
| 0 | −KM4 | 5.3 ± 0.11 e | 8.5 ± 0.15 e | 17.3 ± 0.13 b | 29.4 ± 0.21 b |
| +KM4 | 4.2 ± 0.14 f | 7.1 ± 0.12 f | 19.4 ± 0.12 a | 31.6 ± 0.19 a | |
| 80 | −KM4 | 8.9 ± 0.16 c | 11.4 ± 0.17 c | 12.8± 0.14 d | 22.8 ± 0.11 d |
| +KM4 | 6.5 ± 0.12 d | 10.0 ± 0.14 d | 15.2 ± 0.19 c | 26.7 ± 0.18 c | |
| 160 | −KM4 | 16.6 ± 0.11 a | 14.6 ± 0.12 a | 10.1 ± 0.11 e | 16.2 ± 0.22 f |
| +KM4 | 12.8 ± 0.14 b | 12.1 ± 0.15 b | 13.6 ± 0.16 d | 18.3 ± 0.24 e |
| NaCl (mM) | S. liquefaciens KM4 | LRWC (%) | DPPH (IC50) | Chl a (mg g−1 FW) | Chl b (mg g−1 FW) | Total Chl (mg g−1 FW) | Carotenoid (mg g−1 FW) |
|---|---|---|---|---|---|---|---|
| 0 | −KM4 | 86.1 ± 1.23 b | 0.62 ± 0.07 a | 3.31 ± 0.12 b | 1.72 ± 0.11 b | 5.03 ± 0.11 b | 0.29 ± 0.05 e |
| +KM4 | 87.9 ± 2.12 a | 0.56 ± 0.05 b | 3.74 ± 0.14 a | 1.97 ± 0.09 a | 5.71 ± 0.17 a | 0.34 ± 0.04 a | |
| 80 | −KM4 | 78.4 ± 1.87 d | 0.51 ± 0.08 c | 2.52 ± 0.18 d | 1.36 ± 0.08 d | 3.88 ± 0.09 e | 0.24 ± 0.08 d |
| +KM4 | 82.6 ± 2.41 c | 0.44 ± 0.09 d | 3.18 ± 0.11 c | 1.61 ± 0.06 c | 4.79 ± 0.12 c | 0.28 ± 0.04 b | |
| 160 | −KM4 | 72.3 ± 2.11 f | 0.43 ± 0.05 d | 2.11 ± 0.17 e | 1.11 ± 0.09 e | 3.22 ± 0.16 f | 0.20 ± 0.06 f |
| +KM4 | 77.9 ± 1.37 e | 0.39 ± 0.03 e | 2.53 ± 0.14 d | 1.37 ± 0.08 d | 3.90 ± 0.13 d | 0.25 ± 0.03 c |
| NaCl (mM) | Serratia liquefaciens KM4 | Proline (mg g−1 DW) | Soluble Sugars (mg g−1 DW) | Proteins (mg g−1 DW) | Total Free Amino Acids (mg g−1 DW) | Phenols (µmol g−1 FW) | Flavonoid (µmol g−1 FW) |
|---|---|---|---|---|---|---|---|
| 0 | −KM4 | 1.42 ± 0.17 f | 27.54 ± 1.13 d | 23.04 ±1.51 e | 10.16 ± 0.13 f | 7.51 ± 0.41 b | 1.24 ± 0.14 b |
| +KM4 | 1.77 ± 0.12 e | 29.13 ± 1.47 a | 23.93 ±1.25 d | 11.03 ± 0.17 e | 8.26 ± 0.33 a | 1.51 ± 0.11 a | |
| 80 | −KM4 | 2.25 ± 0.14 d | 25.88 ± 1.38 e | 24.81 ±1.28 c | 12.14 ± 1.14 d | 6.03 ± 0.27 e | 1.05 ±0.16 d |
| +KM4 | 2.91 ± 0.18 c | 28.04 ± 1.29 c | 26.42 ±1.13 b | 15.26 ± 1.32 b | 7.22 ± 0.35 c | 1.17 ± 0.21 c | |
| 160 | −KM4 | 3.41 ± 0.21 b | 22.14 ± 1.37 f | 25.77 ±1.18 b | 14.17 ± 2.13 c | 5.04 ± 0.29 f | 0.98 ± 0.10 e |
| +KM4 | 3.98 ± 0.19 a | 28.89 ± 1.27 b | 33.27 ±1.24 a | 17.11 ± 2.24 a | 6.11 ±0.42 d | 1.07 ±0.12 d |
| NaCl (mM) | KM4 | H2O2 | MDA | EL (%) | Pn (μmol m2 s−1) | E (mmol m2 s−1) | gs (mol m2 s−1) |
|---|---|---|---|---|---|---|---|
| 0 | −KM4 | 17.8 ± 1.11 e | 6.9 ± 1.16 e | 48.3 ± 2.15 e | 13.24 ± 0.08 b | 1.88 ± 0.05 b | 0.09 ± 0.07 b |
| +KM4 | 15.1 ± 1.34 f | 5.1 ± 1.21 f | 42.5 ± 2.03 f | 14.91 ± 0.07 a | 2.06 ± 0.11 a | 0.12 ± 0.09 a | |
| 80 | −KM4 | 19.8 ± 1.47 c | 11.3 ± 1.24 c | 69.2 ± 2.89 b | 9.54 ± 0.05 d | 1.54 ± 0.05 d | 0.06 ±0.04 d |
| +KM4 | 18.1 ±1.32 d | 9.4 ± 1.72 d | 55.7 ±2.13 d | 10.98 ± 0.15 c | 1.72 ± 0.08 c | 0.07 ± 0.08 c | |
| 160 | −KM4 | 22.4 ± 1.22 a | 24.8 ± 1.27 a | 77.4 ± 2.26 a | 7.11 ± 0.13 f | 1.17 ± 0.09 f | 0.03 ± 0.05 f |
| +KM4 | 20.1 ± 1.14 b | 15.7 ± 1.82 b | 66.8 ± 2.91 c | 8.87 ± 0.15 e | 1.33 ± 0.07 e | 0.05 ± 0.01 e |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113310
El-Esawi MA, Alaraidh IA, Alsahli AA, Alzahrani SM, Ali HM, Alayafi AA, Ahmad M. Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. International Journal of Molecular Sciences. 2018; 19(11):3310. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113310
Chicago/Turabian StyleEl-Esawi, Mohamed A., Ibrahim A. Alaraidh, Abdulaziz A. Alsahli, Saud M. Alzahrani, Hayssam M. Ali, Aisha A. Alayafi, and Margaret Ahmad. 2018. "Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression" International Journal of Molecular Sciences 19, no. 11: 3310. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19113310