Inflammation and Oxidative Stress in Chronic Kidney Disease—Potential Therapeutic Role of Minerals, Vitamins and Plant-Derived Metabolites

 ,
,  ,
,
Abstract
:1. Introduction
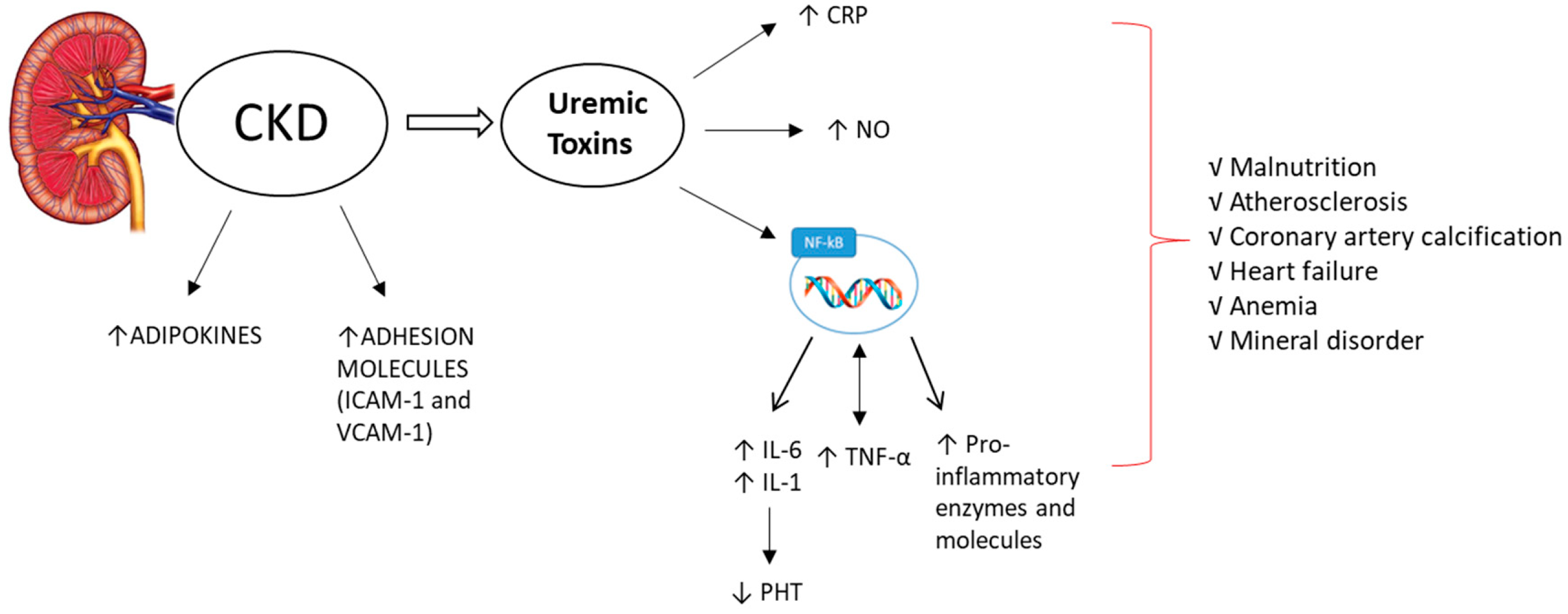
2. The Key Role of Inflammation in CKD
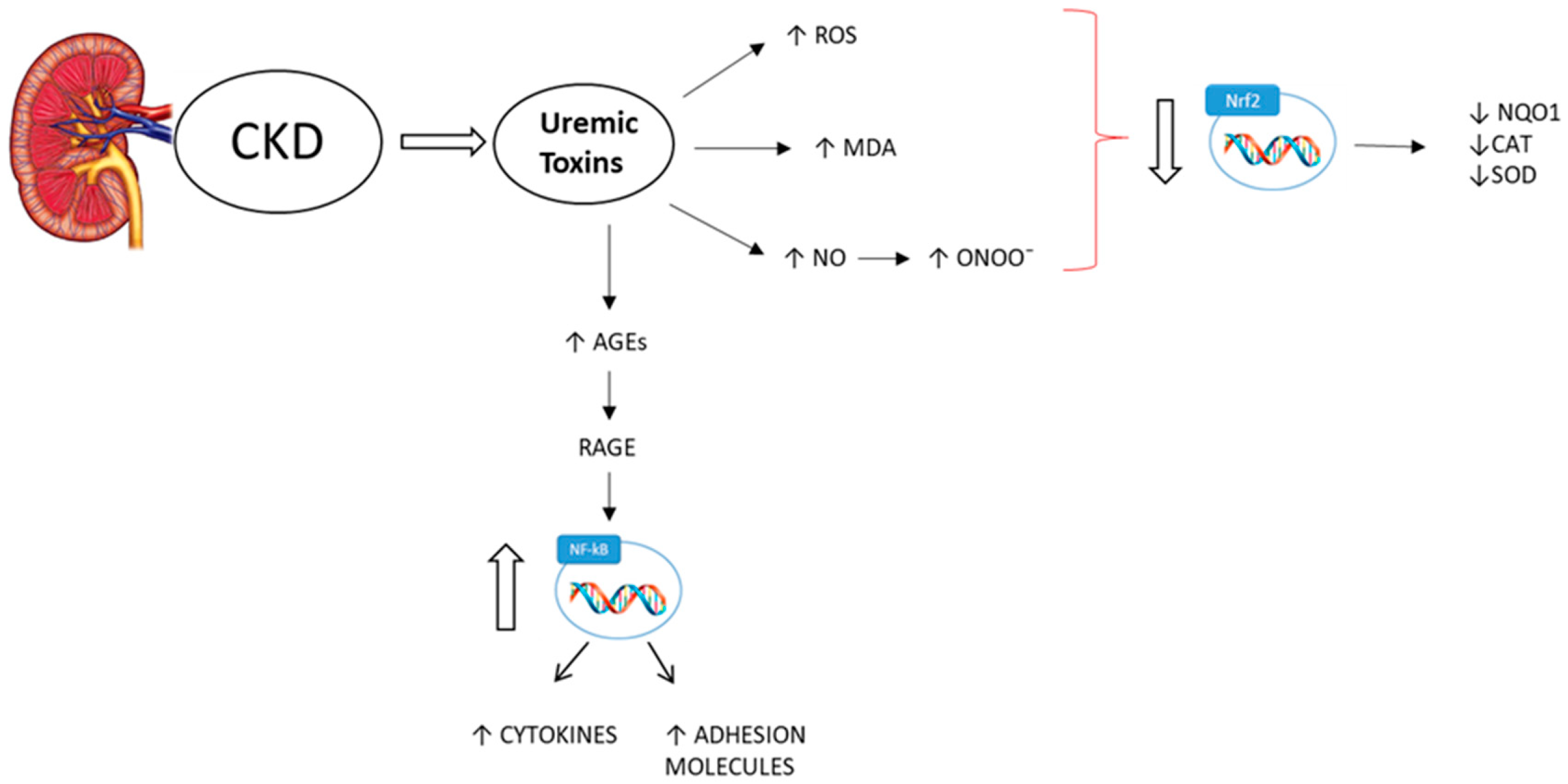
3. Oxidative Stress and CKD
4. Anti-Inflammatory and Antioxidant Compounds
4.1. Minerals and Vitamins
4.1.1. Magnesium
4.1.2. Selenium
4.1.3. Phosphate
4.1.4. Vitamin C
4.1.5. Vitamin E
4.1.6. Vitamin D
4.1.7. Vitamin A
4.1.8. Vitamin B1 (Thiamin)
4.2. Plant-Derived Metabolites
5. The Importance of Nutritional Therapy during CKD
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CKD | Chronic kidney disease |
| ESRD | End-stage renal disease |
| GFR | Glomerular filtration rate |
| Epo | Erythropoietin |
| CRP | C-reactive protein |
| IL-6 | Interleukin-6 |
| IL-1 | Interleukin-1 |
| TNF- α | Tumor necrosis factor-α |
| HD | Hemodialysis |
| MCP-1 | Monocyte chemoattractant protein-1 |
| PTH | Parathyroid hormone |
| RANKL | Receptor activator of NF-κB ligand |
| COX-2 | Cyclooxygenase-2 |
| iNOS | Inducible nitric oxide synthase |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| IS | Indoxyl sulfate |
| PC | p-cresol |
| PCS | p-cresol sulfate |
| NO | Nitrogen monoxide |
| PD | Peritoneal dialysis |
| ROS | Reactive oxygen species |
| NOX | NADPH oxidases |
| 8-OH-dG | 8-hydroxy-2′-deoxyguanosine |
| RNS | Reactive nitrogen species |
| AGEs | Advanced glycation end products |
| RAGE | Receptor system for AGEs |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| NQO1 | NAD(P)H dehydrogenase [quinone] 1 |
| MDA | Malondialdehyde |
| GSH-Px | Glutathione peroxidases |
| ATRA | All-trans retinoic acid |
| FGF23 | Fibroblast growth factor 23 |
| LDH | Lactate dehydrogenase |
| VEGF | Vascular endothelial growth factor |
| HEA | N6-(2-hydroxyethyl)-adenosine |
| UUO | Unilateral ureteral obstruction |
| EGCG | Epigallocatechin-3-gallate |
| VLPD | Very low protein diet |
References
- Jha, V.; Garcia-Garcia, G.; Iseki, K.; Li, Z.; Naicker, S.; Plattner, B.; Saran, R.; Wang, A.Y.; Yang, C.W. Chronic kidney disease: Global dimension and perspectives. Lancet 2013, 382, 260–272. [Google Scholar] [CrossRef]
- Minutolo, R.; Lapi, F.; Chiodini, P.; Simonetti, M.; Bianchini, E.; Pecchioli, S.; Cricelli, I.; Cricelli, C.; Piccinocchi, G.; Conte, G.; et al. Risk of ESRD and death in patients with CKD not referred to a nephrologist: A 7-year prospective study. Clin. J. Am. Soc. Nephrol. 2014, 9, 1586–1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Nicola, L.; Donfrancesco, C.; Minutolo, R.; Lo Noce, C.; De Curtis, A.; Palmieri, L.; Iacoviello, L.; Conte, G.; Chiodini, P.; Sorrentino, F.; et al. Epidemiology of chronic kidney disease in Italy: Current state and contribution of the CARHES study. G. Ital. Nefrol. 2011, 28, 401–407. [Google Scholar] [PubMed]
- Wen, C.P.; Cheng, T.Y.; Tsai, M.K.; Chang, Y.C.; Chan, H.T.; Tsai, S.P.; Chiang, P.H.; Hsu, C.C.; Sung, P.K.; Hsu, Y.H.; et al. All-cause mortality attributable to chronic kidney disease: A prospective cohort study based on 462 293 adults in Taiwan. Lancet 2008, 371, 2173–2182. [Google Scholar] [CrossRef]
- Black, A.P.; Cardozo, L.F.; Mafra, D. Effects of Uremic Toxins from the Gut Microbiota on Bone: A Brief Look at Chronic Kidney Disease. Ther. Apher. Dial. 2015, 19, 436–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popolo, A.; Adesso, S.; Pinto, A.; Autore, G.; Marzocco, S. L-Arginine and its metabolites in kidney and cardiovascular disease. Amino Acids 2014, 46, 2271–2286. [Google Scholar] [CrossRef]
- Marzocco, S.; Popolo, A.; Bianco, G.; Pinto, A.; Autore, G. Pro-apoptotic effect of methylguanidine on hydrogen peroxide-treated rat glioma cell line. Neurochem. Int. 2010, 57, 518–524. [Google Scholar] [CrossRef]
- Marzocco, S.; Di Paola, R.; Ribecco, M.T.; Sorrentino, R.; Domenico, B.; Genesio, M.; Pinto, A.; Autore, G.; Cuzzocrea, S. Effect of methylguanidine in a model of septic shock induced by LPS. Free Radic. Res. 2004, 38, 1143–1153. [Google Scholar] [CrossRef]
- Marzocco, S.; Di Paola, R.; Genovese, T.; Sorrentino, R.; Britti, D.; Scollo, G.; Pinto, A.; Cuzzocrea, S.; Autore, G. Methylguanidine reduces the development of non septic shock induced by zymosan in mice. Life Sci. 2004, 75, 1417–1433. [Google Scholar] [CrossRef]
- Marzocco, S.; Di Paola, R.; Serraino, I.; Sorrentino, R.; Meli, R.; Mattaceraso, G.; Cuzzocrea, S.; Pinto, A.; Autore, G. Effect of methylguanidine in carrageenan-induced acute inflammation in the rats. Eur. J. Pharmacol. 2004, 484, 341–350. [Google Scholar] [CrossRef]
- Autore, G.; Marzocco, S.; Sorrentino, R.; Mirone, V.G.; Baydoun, A.; Pinto, A. In vitro and in vivo TNFalpha synthesis modulation by methylguanidine, an uremic catabolyte. Life Sci. 1999, 65, PL121–PL127. [Google Scholar] [CrossRef] [Green Version]
- Di Iorio, B.R.; Di Micco, L.; Marzocco, S.; De Simone, E.; De Blasio, A.; Sirico, M.L.; Nardone, L. UBI Study Group. Very Low-Protein Diet (VLPD) Reduces Metabolic Acidosis in Subjects with Chronic Kidney Disease: The “Nutritional Light Signal” of the Renal Acid Load. Nutrients 2017, 9, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adesso, S.; Magnus, T.; Cuzzocrea, S.; Campolo, M.; Rissiek, B.; Paciello, O.; Autore, G.; Pinto, A.; Marzocco, S. Indoxyl Sulfate Affects Glial Function Increasing Oxidative Stress and Neuroinflammation in Chronic Kidney Disease: Interaction between Astrocytes and Microglia. Front. Pharmacol. 2017, 8, 370. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Chmielewski, M.; Honda, H.; Pecoits-Filho, R.; Matsuo, S.; Yuzawa, Y.; Tranaeus, A.; Stenvinkel, P.; Lindholm, B. Aspects of immune dysfunction in end-stage renal disease. Clin. J. Am. Soc. Nephrol. 2008, 3, 1526–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masako, K.; Kentaro, K.; Yoshiki, S.; Kunitoshi, I.; Yusuke, O. Chronic kidney disease, inflammation and cardiovascular disease risk in rheumatoid arthritis. J. Cardiol. 2018, 71, 277–283. [Google Scholar]
- Popolo, A.; Autore, G.; Pinto, A.; Marzocco, S. Oxidative stress in patients with cardiovascular disease and chronic renal failure. Free Radic. Res. 2013, 47, 346–356. [Google Scholar] [CrossRef]
- Qian, Q. Inflammation: A Key Contributor to the Genesis and Progression of Chronic Kidney Disease. Contrib. Nephrol. 2017, 191, 72–83. [Google Scholar]
- Morena, M.; Cristol, J.P.; Senécal, L.; Leray-Moragues, H.; Krieter, D.; Canaud, B. Oxidative stress in hemodialysis patients: Is NADPH oxidase complex the culprit? Kidney Int. 2002, 61, S109–S114. [Google Scholar] [CrossRef] [Green Version]
- Libetta, C.; Sepe, V.; Esposito, P.; Galli, F.; Dal Canton, A. Oxidative stress and inflammation: Implications in uremia and hemodialysis. Clin. Biochem. 2011, 44, 1189–1198. [Google Scholar] [CrossRef]
- Mihai, S.; Codrici, E.; Popescu, I.D.; Enciu, A.M.; Albulescu, L.; Necula, L.G.; Mambet, C.; Anton, G.; Tanase, C. Inflammation-Related Mechanisms in Chronic Kidney Disease Prediction, Progression and Outcome. J. Immunol. Res. 2018, 2018, 2180373. [Google Scholar] [CrossRef]
- Gupta, J.; Mitra, N.; Kanetsky, P.A.; Devaney, J.; Wing, M.R.; Reilly, M.; Shah, V.O.; Balakrishnan, V.S.; Guzman, N.J.; Girndt, M.; et al. Association between albuminuria, kidney function and inflammatory biomarker profile in CKD in CRIC. Clin. J. Am. Soc. Nephrol. 2012, 7, 1938–1946. [Google Scholar] [CrossRef] [Green Version]
- Shlipak, M.G.; Fried, L.F.; Crump, C.; Bleyer, A.J.; Manolio, T.A.; Tracy, R.P.; Furberg, C.D.; Psaty, B.M. Elevations of inflammatory and procoagulant biomarkers in elderly persons with renal insufficiency. Circulation 2002, 107, 87–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenvinkel, P.; Heimburger, O.; Paultre, F.; Diczfalusy, U.; Wang, T.; Berglund, L.; Jogestrand, T. Strong association between malnutrition, inflammation and atherosclerosis in chronic renal failure. Kidney Int. 1999, 55, 1899–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro-Gonzalez, J.F.; Mora-Fernandez, C.; Muros de Fuentes, M.; García-Pérez, J. Inflammatory molecules and pathways in the pathogenesis of diabetic nephropathy. Nat. Rev. Nephrol. 2011, 7, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Clapp, B.R.; Hingorani, A.D.; Kharbanda, R.K.; Mohamed-Ali, V.; Stephens, J.W.; Vallance, P.; MacAllister, R.J. Inflammation-induced endothelial dysfunction involves reduced nitric oxide bioavailability and increased oxidant stress. Cardiovasc. Res. 2004, 64, 172–178. [Google Scholar] [CrossRef]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef]
- Akchurin, O.M.; Kaskel, F. Update on inflammation in chronic kidney disease. Blood Purif. 2015, 39, 84–92. [Google Scholar] [CrossRef]
- De Oliveira Júnior, W.V.; Sabino Ade, P.; Figueiredo, R.C.; Rios, D.R. Inflammation and poor response to treatment with erythropoietin in chronic kidney disease. J. Bras. Nefrol. 2015, 37, 255–256. [Google Scholar] [CrossRef]
- Wagner, M.; Alam, A.; Zimmermann, J.; Rauh, K.; Koljaja-Batzner, A.; Raff, U.; Wanner, C.; Schramm, L. Endogenous erythropoietin and the association with inflammation and mortality in diabetic chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2011, 6, 1573–1579. [Google Scholar] [CrossRef]
- Goldstein, S.L.; Leung, J.C.; Silverstein, D.M. Pro- and anti-inflammatory cytokines in chronic pediatric dialysis patients: Effect of aspirin. Clin. J. Am. Soc. Nephrol. 2006, 1, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Liuzzo, G.; Biasucci, L.M.; Gallimore, J.R.; Grillo, R.L.; Rebuzzi, A.G.; Pepys, M.B.; Maseri, A. The prognostic value of C-reactive protein and serum amyloid a protein in severe unstable angina. N. Eng. J. Med. 1994, 331, 417–424. [Google Scholar] [CrossRef]
- Bazeley, J.; Bieber, B.; Li, Y.; Morgenstern, H.; de Sequera, P.; Combe, C.; Yamamoto, H.; Gallagher, M.; Port, F.K.; Robinson, B.M. C-reactive protein and prediction of 1-year mortality in prevalent hemodialysis patients. Clin. J. Am. Soc. Nephrol. 2011, 6, 2452–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roubicek, T.; Bartlova, M.; Krajickova, J.; Haluzikova, D.; Mraz, M.; Lacinova, Z.; Kudla, M.; Teplan, V.; Haluzik, M. Increased production of proinflammatory cytokines in adipose tissue of patients with end-stage renal disease. Nutrition 2009, 25, 762–768. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, B.; Alexander, D.; Janessa, A.; Häring, H.U.; Lang, F.; Risler, T. Acute effects of hemodialysis on cytokine transcription profiles: Evidence for C-reactive protein-dependency of mediator induction. Kidney Int. 2006, 70, 2124–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taraz, M.; Taraz, S.; Dashti-Khavidaki, S. Association between depression and inflammatory/anti-inflammatory cytokines in chronic kidney disease and end-stage renal disease patients: A review of literature. Hemodial. Int. 2014, 19, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Feroze, U.; Molnar, M.Z.; Dukkipati, R.; Kovesdy, C.P.; Kalantar-Zadeh, K. Insights into nutritional and inflammatory aspects of low parathyroid hormone in dialysis patients. J. Ren. Nutr. 2011, 21, 100–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.C.; Lin, Y.C.; Hsu, C.J.; Kao, C.C.; Chang, F.C.; Chen, T.E.; Chen, H.H.; Hsu, C.C.; Wu, M.S. Effect Modifying Role of Serum Calcium on Mortality-Predictability of PTH and Alkaline Phosphatase in Hemodialysis Patients: An Investigation Using Data from the Taiwan Renal Registry Data System from 2005 to 2012. PLoS ONE 2015, 10, e0129737. [Google Scholar] [CrossRef] [Green Version]
- Neirynck, N.; Glorieux, G.; Schepers, E.; Verbeke, F.; Vanholder, R. Soluble tumor necrosis factor receptor 1 and 2 predict outcomes in advanced chronic kidney disease: A prospective cohort study. PLoS ONE 2015, 10, e0122073. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, G.; Nakano, K.; Sawada, M.; Uno, K.; Shibayama, Y.; Ienaga, K.; Kondo, M. Possible role of tumor necrosis factor and interleukin-1 in the development of diabetic nephropathy. Kidney Int. 1991, 40, 1007–1012. [Google Scholar] [CrossRef] [Green Version]
- Panuccio, V.; Enia, G.; Tripepi, R.; Aliotta, R.; Mallamaci, F.; Tripepi, G.; Zoccali, C. Pro-inflammatory cytokines and bone fractures in CKD patients. An exploratory single centre study. BMC Nephrol. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Karin, M.; Greten, F.R. NF-kappaB: Linking inflammation and immunity to cancer development and progression. Nat. Rev. Immunol. 2005, 5, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Chen, Y.A.; Chen, S.S.; Chen, Y.J.; Lin, Y.K. Uremic Toxins—Novel Arrhythmogenic Factor in Chronic Kidney Disease – Related Atrial Fibrillation. Acta Cardiol. Sin. 2016, 32, 259–264. [Google Scholar] [PubMed]
- Adesso, S.; Popolo, A.; Bianco, G.; Sorrentino, R.; Pinto, A.; Autore, G.; Marzocco, S. The uremic toxin indoxyl sulphate enhances macrophage response to LPS. PLoS ONE 2013, 8, e76778. [Google Scholar] [CrossRef] [Green Version]
- Stockler-Pinto, M.B.; Saldanha, J.F.; Yi, D.; Mafra, D.; Fouque, D.; Soulage, C.O. The uremic toxin indoxyl sulfate exacerbates reactive oxygen species production and inflammation in 3T3-L1 adipose cells. Free Radic. Res. 2016, 50, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adesso, S.; Ruocco, M.; Rapa, S.F.; Dal Piaz, F.; Di Iorio, B.R.; Popolo, A.; Autore, G.; Nishijima, F.; Pinto, A.; Marzocco, S. Effect of Indoxyl Sulfate on the Repair and Intactness of Intestinal Epithelial Cells: Role of Reactive Oxygen Species’ Release. Int. J. Mol. Sci. 2019, 20, 2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Iorio, B.R.; Rocchetti, M.T.; De Angelis, M.; Cosola, C.; Marzocco, S.; Di Micco, L.; di Bari, I.; Accetturo, M.; Vacca, M.; Gobbetti, M.; et al. Nutritional Therapy Modulates Intestinal Microbiota and Reduces Serum Levels of Total and Free Indoxyl Sulfate and P-Cresyl Sulfate in Chronic Kidney Disease (Medika Study). J. Clin. Med. 2019, 8, 1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stockler-Pinto, M.B.; Soulage, C.O.; Borges, N.A.; Cardozo, L.F.M.F.; Dolenga, C.J.; Nakao, L.S.; Pecoits-Filho, R.; Fouque, D.; Mafra, D. From bench to the hemodialysis clinic: Protein-bound uremic toxins modulate NF-κB/Nrf2 expression. Int. Urol. Nephrol. 2018, 50, 347–354. [Google Scholar] [CrossRef]
- Adesso, S.; Paterniti, I.; Cuzzocrea, S.; Fujioka, M.; Autore, G.; Magnus, T.; Pinto, A.; Marzocco, S. AST-120 Reduces Neuroinflammation Induced by Indoxyl Sulfate in Glial Cells. J. Clin. Med. 2018, 7, 365. [Google Scholar] [CrossRef] [Green Version]
- Shankar, A.; Syamala, S.; Xiao, J.; Muntner, P. Relationship between plasma leptin level and chronic kidney disease. Int. J. Nephrol. 2012, 2012, 269532. [Google Scholar] [CrossRef]
- Adamczak, M.; Chudek, J.; Wiecek, A. Adiponectin in patients with chronic kidney disease. Semin. Dial. 2009, 22, 391–395. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lai, Y.J.; Lin, Y.C.; Peng, C.C.; Chen, K.C.; Chuang, M.T.; Wu, M.S.; Chang, T.H. Effect of weight loss on the estimated glomerular filtration rates of obese patients at risk of chronic kidney disease: The RIGOR-TMU study. J. Cachexia Sarcopenia Muscle 2019, 10, 756–766. [Google Scholar] [CrossRef] [Green Version]
- Bonventre, J.V.; Colvin, R.B. Adhesion molecules in renal disease. Current Opinion in Nephrology and Hypertension 1996, 5, 254–261. [Google Scholar] [CrossRef]
- Locatelli, F.; Canaud, B.; Eckardt, K.U.; Stenvinkel, P.; Wanner, C.; Zoccali, C. Oxidative stress in end-stage renal disease: An emerging threat to patient outcome. Nephrol. Dial. Transplant. 2003, 18, 1272–1280. [Google Scholar] [CrossRef] [PubMed]
- Hasselwander, O.; Young, I.S. Oxidative stress in chronic renal failure. Free Radic. Res. 1998, 29, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Annuk, M.; Zilmer, M.; Lind, L.; Linde, T.; Fellstrom, B. Oxidative stress and endothelial function in chronic renal failure. J. Am. Soc. Nephrol. 2001, 12, 2747–2752. [Google Scholar] [PubMed]
- Dounousi, E.; Papavasiliou, E.; Makedou, A.; Ioannou, K.; Katopodis, K.P.; Tselepis, A.; Siamopoulos, K.C.; Tsakiris, D. Oxidative stress is progressively enhanced with advancing stages of CKD. Am. J. Kidney Dis. 2006, 48, 752–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liakopoulos, V.; Roumeliotis, S.; Gorny, X.; Dounousi, E.; Mertens, P.R. Oxidative stress in hemodialysis patients: A review of the literature. Oxid. Med. Cell. Longev. 2017, 2017, 1–22. [Google Scholar] [CrossRef]
- Ferraro, B.; Galli, F.; Frei, B.; Kingdon, E.; Canestrari, F.; Rice-Evans, C.; Buoncristiani, U.; Davenport, A.; Moore, K.P. Peroxynitrite-induced oxidation of plasma lipids is enhanced in stable hemodialysis patients. Kidney Int. 2003, 63, 2207–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krata, N.; Zagozdzon, R.; Foroncewicz, B.; Mucha, K. Oxidative Stress in Kidney Diseases: The Cause or the Consequence? Arch. Immunol. Ther. Exp. 2017, 66, 211–220. [Google Scholar] [CrossRef] [Green Version]
- You, Y.H.; Okada, S.; Ly, S.; Jandeleit-Dahm, K.; Barit, D.; Namikoshi, T.; Sharma, K. Role of Nox2 in diabetic kidney disease. Am. J. Physiol. Ren. Physiol. 2013, 304, F840–F848. [Google Scholar] [CrossRef]
- Simone, S.; Rascio, F.; Castellano, G.; Divella, C.; Chieti, A.; Ditonno, P.; Battaglia, M.; Crovace, A.; Staffieri, F.; Oortwijn, B.; et al. Complement-dependent NADPH oxidase enzyme activation in renal ischemia/reperfusion injury. Free Radic. Biol. Med. 2014, 74, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Holterman, C.E.; Thibodeau, J.F.; Towaij, C.; Gutsol, A.; Montezano, A.C.; Parks, R.J.; Cooper, M.E.; Touyz, R.M.; Kennedy, C.R. Nephropathy and elevated BP in mice with podocyte-specific NADPH oxidase 5 expression. J. Am. Soc. Nephrol. 2014, 25, 784–797. [Google Scholar] [CrossRef] [Green Version]
- Beckman, K.B.; Ames, B.N. Oxidative decay of DNA. J. Biol. Chem. 1997, 272, 19633–19636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Descamps-Latscha, B.; Drüeke, T.; Witko-Sarsat, V. Dialysis-induced oxidative stress: Biological aspects, clinical consequences and therapy. Semin. Dial. 2001, 14, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Kinugasa, E. Markers and possible uremic toxins: Japanese Experiences. Contrib. Nephrol. 2011, 168, 134–138. [Google Scholar] [PubMed]
- Zill, H.; Gunther, R.; Erbersdobler, H.F.; Folsch, U.R.; Faist, V. RAGE expression and AGE-induced MAP kinase activation in Caco-2 cells. Biochem. Biophys. Res. Commun. 2001, 288, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Boulanger, E.; Wautier, M.P.; Wautier, J.L.; Boval, B.; Panis, Y.; Wernert, N.; Danze, P.-M.; Dequiedt, P. AGEs bind to mesothelial cells via RAGE and stimulate VCAM-1 expression. Kidney Int. 2002, 61, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, S.; Pergola, P.E.; Zager, R.A.; Vaziri, N.D. Targeting the Transcription Factor Nrf2 to Ameliorate Oxidative Stress and Inflammation in Chronic Kidney Disease. Kidney Int. 2013, 83, 1029–1041. [Google Scholar] [CrossRef] [Green Version]
- Modlinger, P.S.; Wilcox, C.S.; Aslam, S. Nitric oxide, oxidative stress and progression of chronic renal failure. Semin. Nephrol. 2004, 24, 354–365. [Google Scholar] [CrossRef]
- Modaresi, A.; Nafar, M.; Sahraei, Z. Oxidative Stress in Chronic Kidney Disease. Iran J. Kidney Dis. 2015, 9, 165–179. [Google Scholar]
- Xu, G.; Luo, K.; Liu, H.; Huang, T.; Fang, X.; Tu, W. The progress of inflammation and oxidative stress in patients with chronic kidney disease. Ren. Fail. 2015, 37, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Batista Junior, M.L.; Lopes, R.D.; Seelaender, M.C.; Lopes, A.C. Anti-inflammatory effect of physical training in heart failure: Role of TNF-alpha and IL-10. Arq. Bras. Cardiol. 2009, 93, 643–651. [Google Scholar]
- Mena, M.P.; Sacanella, E.; Vazquez-Agell, M.; Morales, M.; Fito, M.; Escoda, R.; Serrano-Martínez, M.; Salas-Salvadó, J.; Benages, N.; Casas, R.; et al. Inhibition of circulating immune cell activation: A molecular antiinflammatory effect of the mediterranean diet. Am. J. Clin. Nutr. 2009, 89, 248–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goicoechea, M.; de Vinuesa, S.G.; Lahera, V.; Cachofeiro, V.; Gomez-Campdera, F.; Vega, A.; Abad, S.; Luño, J. Effects of atorvastatin on inflammatory and fibrinolytic parameters in patients with chronic kidney disease. J. Am. Soc. Nephrol. 2006, 17 (Suppl. S3), S231–S235. [Google Scholar] [CrossRef] [Green Version]
- Stenvinkel, P.; Andersson, P.; Wang, T.; Lindholm, B.; Bergstrom, J.; Palmblad, J.; Heimbürger, O.; Cederholm, T. Do ACE-inhibitors suppress tumour necrosis factor-alpha production in advanced chronic renal failure? J. Intern. Med. 1999, 246, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.M.; Powell-Griner, E.; McFann, K.; Nahin, R.L. Complementary and alternative medicine use among adults: United States, 2002. Adv. Data 2004, 343, 1–19. [Google Scholar] [CrossRef]
- Hatcher, H.; Planalp, R.; Cho, J.; Torti, F.M.; Torti, S.V. Curcumin: From ancient medicine to current clinical trials. Cell. Mol. Life Sci. 2008, 65, 1631–1652. [Google Scholar] [CrossRef]
- Fu, Y.; Zheng, S.; Lin, J.; Ryerse, J.; Chen, A. Curcumin protects the rat liver from CCl4-caused injury and fibrogenesis by attenuating oxidative stress and suppressing inflammation. Mol. Pharm. 2008, 73, 399–409. [Google Scholar] [CrossRef] [Green Version]
- Vera, M.; Torramade-Moix, S.; Martin-Rodriguez, S.; Cases, A.; Cruzado, J.M.; Rivera, J.; Escolar, G.; Palomo, M.; Diaz-Ricart, M. Antioxidant and Anti-Inflammatory Strategies Based on the Potentiation of Glutathione Peroxidase Activity Prevent Endothelial Dysfunction in Chronic Kidney Disease. Cell. Physiol. Biochem. 2018, 51, 1287–1300. [Google Scholar] [CrossRef]
- Quamme, G.A. Control of magnesium transport in the thick ascending limb. Am. J. Physiol. 1989, 256 Pt 2, F197–F210. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, J.; Rodríguez, M.; Messa, P. Magnesium in chronic kidney disease Stages 3 and 4 and in dialysis patients. Clin. Kidney J. 2012, 5 (Suppl. S1), i39–i51. [Google Scholar] [CrossRef] [PubMed]
- Van de Wal-Visscher, E.R.; Kooman, J.P.; van der Sande, F.M. Magnesium in Chronic Kidney Disease: Should We Care? Blood Purif. 2018, 45, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; He, T.; Wang, M.; Nie, L.; Zhang, Y.; Wang, Y.; Huang, Y.; Feng, B.; Zhang, J.; Zhao, J. Serum magnesium, mortality and cardiovascular disease in chronic kidney disease and end-stage renal disease patients: A systematic review and meta-analysis. J. Nephrol. 2019, 32, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, X.; Qi, H.; Wang, J.; Wang, M.; Zhang, Y.; Yan, H.; Zhuang, S. Correlation of serum magnesium with cardiovascular risk factors in maintenance hemodialysis patients—A cross-sectional study. Magnes. Res. 2013, 26, 100–108. [Google Scholar] [CrossRef]
- Spiegel, D.M. Magnesium in chronic kidney disease: Unanswered questions. Blood Purif. 2011, 31, 172–176. [Google Scholar] [CrossRef]
- Jankowska, M.; Rutkowski, B.; Dębska-Ślizień, A. Vitamins and Microelement Bioavailability in Different Stages of Chronic Kidney Disease. Nutrients 2017, 9, 282. [Google Scholar] [CrossRef] [Green Version]
- Zachara, B.A. Selenium and selenium-dependent antioxidants in chronic kidney disease. Adv. Clin. Chem. 2015, 68, 131–151. [Google Scholar]
- Lemoine, S.; Fournier, T.; Kocevar, G.; Belloi, A.; Normand, G.; Ibarrola, D.; Sappey-Marinier, D.; Juillard, L. Intracellular Phosphate Dynamics in Muscle Measured by Magnetic Resonance Spectroscopy during Hemodialysis. J. Am. Soc. Nephrol. 2016, 27, 2062–2068. [Google Scholar] [CrossRef] [Green Version]
- O’Seaghdha, C.M.; Hwang, S.J.; Muntner, P.; Melamed, M.L.; Fox, C.S. Serum phosphorus predicts incident chronic kidney disease and end-stage renal disease. Nephrol. Dial. Transpl. 2011, 26, 2885–2890. [Google Scholar] [CrossRef] [Green Version]
- Levin, A.; Bakris, G.L.; Molitch, M.; Smulders, M.; Tian, J.; Williams, L.A.; Andress, D.L. Prevalence of abnormal serum vitamin D, PTH, calcium and phosphorus in patients with chronic kidney disease: Results of the study to evaluate early kidney disease. Kidney Int. 2007, 71, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Kidney Disease: Improving Global Outcomes (KDIGO) CKD-MBD Work Group. KDIGO clinical practice guideline for the diagnosis, evaluation, prevention and treatment of Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD). Kidney Int. Suppl. 2009, 113, S1–S130. [Google Scholar]
- Di Micco, L.; Di Lullo, L.; Bellasi, A.; Di Iorio, B.R. Very Low Protein Diet for Patients with Chronic Kidney Disease: Recent Insights. J. Clin. Med. 2019, 8, 718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Iorio, B.R.; Marzocco, S.; Bellasi, A.; De Simone, E.; Dal Piaz, F.; Rocchetti, M.T.; Cosola, C.; Di Micco, L.; Gesualdo, L. Nutritional therapy reduces protein carbamylation through urea lowering in chronic kidney disease. Nephrol. Dial. Transpl. 2018, 33, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Marzocco, S.; Dal Piaz, F.; Di Micco, L.; Torraca, S.; Sirico, M.L.; Tartaglia, D.; Autore, G.; Di Iorio, B. Very low protein diet reduces indoxyl sulfate levels in chronic kidney disease. Blood Purif. 2013, 35, 196–201. [Google Scholar] [CrossRef]
- Wang, S.; Eide, T.C.; Sogn, E.M.; Berg, K.J.; Sund, R.B. Plasma ascorbic acid in patients undergoing chronic hemodialysis. Eur. J. Clin. Pharmacol. 1999, 55, 527–532. [Google Scholar] [CrossRef]
- Deicher, R.; Ziai, F.; Bieglmayer, C.; Schillinger, M.; Horl, W.H. Low total vitamin C plasma level is a risk factor for cardiovascular morbidity and mortality in hemodialysis patients. J. Am. Soc. Nephrol. 2005, 16, 1811–1818. [Google Scholar] [CrossRef]
- Oboh, H.A.; Idogun, E.S. The assessment of plasma ascorbic acid, alpha tocopherol and albumin creatinine ratio in patients with chronic renal failure. Nig. Q. J. Hosp. Med. 2011, 21, 294–298. [Google Scholar]
- Takahashi, N.; Morimoto, S.; Okigaki, M.; Seo, M.; Someya, K.; Morita, T.; Matsubara, H.; Sugiura, T.; Iwasaka, T. Decreased plasma level of vitamin C in chronic kidney disease: Comparison between diabetic and non-diabetic patients. Nephrol. Dial. Transpl. 2011, 26, 1252–1257. [Google Scholar] [CrossRef] [Green Version]
- Koyner, J.L.; Sher Ali, R.; Murray, P.T. Antioxidants. Do they have a place in the prevention or therapy of acute kidney injury? Nephron. Exp. Nephrol. 2008, 109, e109–e117. [Google Scholar] [CrossRef]
- Chazot, C.; Jean, G.; Kopple, J.D. Can Outcomes be Improved in Dialysis Patients by Optimizing Trace Mineral, Micronutrient and Antioxidant Status? The Impact of Vitamins and their Supplementation. Semin. Dial. 2016, 29, 39–48. [Google Scholar] [CrossRef]
- Singh, U.; Devaraj, S.; Jialal, I. Vitamin E, oxidative stress and inflammation. Annu. Rev. Nutr. 2005, 25, 151–174. [Google Scholar] [CrossRef] [PubMed]
- Keegan, R.; Webb, C.B. Oxidative stress and neutrophil function in cats with chronic renal failure. J. Vet. Intern. Med. 2010, 24, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Yusuf, S.; Sleight, P.; Pogue, J.; Bosch, J.; Davies, R.; Dagenais, G. Effects of an angiotensin-converting enzyme inhibitor ramipril on cardiovascular events in high-risk patients. The Heart Outcomes Prevention Evaluation Study. N. Engl. J. Med. 2000, 342, 145–153. [Google Scholar] [PubMed]
- Mann, J.F.E.; Yi, Q.L.; Gerstein, H.C. Albuminuria as a predictor of cardiovascular and renal outcomes in people with known atherosclerotic cardiovascular disease. Kidney Int. 2004, 66, S59–S62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonn, E.; Bosch, J.; Yusuf, S.; Sheridan, P.; Pogue, J.; Arnold, J.M.; Ross, C.; Arnold, A.; Sleight, P.; Probstfield, J.; et al. HOPE and HOPE-TOO Trial Investigators. Effects of long-term vitamin E supplementation on cardiovascular events and cancer: A randomized controlled trial. JAMA 2005, 293, 1338–1347. [Google Scholar]
- Boaz, M.; Smetana, S.; Weinstein, T.; Matas, Z.; Gafter, U.; Iaina, A.; Knecht, A.; Weissgarten, Y.; Brunner, D.; Fainaru, M.; et al. Secondary prevention with antioxidants of cardiovascular disease in end stage renal disease (SPACE): Randomised placebo-controlled trial. Lancet 2000, 356, 1213–1218. [Google Scholar] [CrossRef]
- Cristol, J.P.; Bosch, J.Y.; Badiou, S.; Leblanc, M.; Lorrho, R.; Descomps, B.; Canaud, B. Erythropoietin and oxidative stress in hemodialysis: Beneficial effects of vitamin E supplementation. Nephrol. Dial. Transpl. 1997, 12, 2312–2317. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Yi, B.; Li, A.M.; Zhang, H. Effects of vitamin E-coated dialysis membranes on anemia, nutrition and dyslipidemia status in hemodialysis patients: A meta-analysis. Ren. Fail. 2015, 37, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef]
- Krassilnikova, M.; Ostrow, K.; Bader, A.; Heeger, P.; Mehrotra, A. Low Dietary Intake of Vitamin D and Vitamin D Deficiency in Hemodialysis Patients. J. Nephrol. 2014, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.S.; Kim, S.W. Vitamin D and chronic kidney disease. Korean J. Intern. Med. 2014, 29, 416–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchi, W.M.; Shimizu, M.H.; Canale, D.; Gois, P.H.; de Bragança, A.C.; Volpini, R.A.; Girardi, A.C.; Seguro, A.C. Vitamin D deficiency is a potential risk factor for contrast-induced nephropathy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 309, R215–R222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedraza-Chaverri, J.; Sánchez-Lozada, L.G.; Osorio-Alonso, H.; Tapia, E.; Scholze, A. New Pathogenic Concepts and Therapeutic Approaches to Oxidative Stress in Chronic Kidney Disease. Oxid. Med. Cell. Longev. 2016, 2016, 6043601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freundlich, M.; Quiroz, Y.; Zhang, Z.; Zhang, Y.; Bravo, Y.; Weisinger, J.R.; Li, Y.C.; Rodriguez-Iturbe, B. Suppression of renin-angiotensin gene expression in the kidney by paricalcitol. Kidney Int. 2008, 74, 1394–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Niño, M.D.; Bozic, M.; Córdoba-Lanús, E.; Valcheva, P.; Gracia, O.; Ibarz, M.; Fernandez, E.; Navarro-Gonzalez, J.F.; Ortiz, A.; Valdivielso, J.M. Beyond proteinuria: VDR activation reduces renal inflammation in experimental diabetic nephropathy. Am. J. Physiol. Ren. Physiol. 2012, 302, F647–F657. [Google Scholar] [CrossRef]
- Deb, D.K.; Sun, T.; Wong, K.E.; Zhang, Z.; Ning, G.; Zhang, Y.; Kong, J.; Shi, H.; Chang, A.; Li, Y.C. Combined vitamin D analog and AT1 receptor antagonist synergistically block the development of kidney disease in a model of type 2 diabetes. Kidney Int. 2010, 77, 1000–1009. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Kang, Y.S.; Dai, C.; Liu, Y. Blockade of Wnt/β-catenin signaling by paricalcitol ameliorates proteinuria and kidney injury. J. Am. Soc. Nephrol. 2011, 22, 90–103. [Google Scholar] [CrossRef]
- Edith, S.; Mondragona, E.; Molina, J.C.; Namorado, T.; Rafael, R.; Muñoza, J.P.; Chaverric, J.; Reyesa, L. All-trans retinoic acid ameliorates inflammatory response mediated by TLR4/NF-κB during initiation of diabetic nephropathy. J. Nutr. Biochem. 2018, 60, 47–60. [Google Scholar]
- Hung, S.C.; Hung, S.H.; Tarng, D.C.; Yang, W.C.; Chen, T.W.; Huang, T.P. Thiamine deficiency and unexplained encephalopathy in hemodialysis and peritoneal dialysis patients. Am. J. Kidney Dis. 2001, 38, 941–947. [Google Scholar] [CrossRef]
- Moradi, M.; Said, H.M. Functional thiamine deficiency in end-stage renal disease: Malnutrition despite ample nutrients. Kidney Int. 2016, 90, 252–254. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Masania, J.; Anwar, A.; Xue, M.; Zehnder, D.; Kanji, H.; Rabbani, N.; Thornalley, P.J. The uremic toxin oxythiamine causes functional thiamine deficiency in end-stage renal disease by inhibiting transketolase activity. Kidney Int. 2016, 90, 396–403. [Google Scholar] [CrossRef]
- Schupp, N. Vitamin supplementation in hemodialysis patients—Compensation for the lack of water-soluble vitamins. Dial. Aktuell. 2009, 13, 322–324. [Google Scholar]
- Cupisti, A.; Brunori, G.; Di Iorio, B.R.; D’Alessandro, C.; Pasticci, F.; Cosola, C.; Bellizzi, V.; Bolasco, P.; Capitanini, A.; Fantuzzi, A.L.; et al. Nutritional treatment of advanced CKD: Twenty consensus statements. J. Nephrol. 2018, 31, 457–473. [Google Scholar] [CrossRef] [Green Version]
- Russo, D.; Bellasi, A.; Pota, A.; Russo, L.; Di Iorio, B. Effects of phosphorus-restricted diet and phosphate-binding therapy on outcomes in patients with chronic kidney disease. J. Nephrol. 2015, 28, 73–80. [Google Scholar] [CrossRef]
- Marzocco, S.; Fazeli, G.; Di Micco, L.; Autore, G.; Adesso, S.; Dal Piaz, F.; Heidland, A.; Di Iorio, B. Supplementation of Short-Chain Fatty Acid, Sodium Propionate, in Patients on Maintenance Hemodialysis: Beneficial Effects on Inflammatory Parameters and Gut-Derived Uremic Toxins, A Pilot Study (PLAN Study). J. Clin. Med. 2018, 7, 315. [Google Scholar] [CrossRef] [Green Version]
- Di Iorio, B.R.; Marzocco, S.; Nardone, L.; Sirico, M.; De Simone, E.; Di Natale, G.; Di Micco, L. Urea and impairment of the Gut-Kidney axis in Chronic Kidney Disease. G. Ital. Nefrol. 2017, 34, 6. [Google Scholar]
- Abarikwu, S.O. Protective effect of quercetin on atrazine-induced oxidative stress in the liver, kidney, brain and heart of adult wistar rats. Toxicol. Int. 2014, 21, 148–155. [Google Scholar] [CrossRef]
- Shaik, Y.B.; Castellani, M.L.; Perrella, A. Role of quercetin (a natural herbal compound) in allergy and inflammation. J. Biol. Regul. Homeost. Agents 2006, 20, 47–52. [Google Scholar]
- Trujillo, J.; Chirino, Y.H.; Molina-Jijón, E.; Andérica-Romero, A.C.; Tapia, E.; Pedraza-Chaverr, J. Renoprotective effect of the antioxidant curcumin: Recent findings. Redox Biol. 2013, 1, 448–456. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Song, Y.; Liang, Y.; Li, R. Quercetin Treatment Improves Renal Function and Protects the Kidney in a Rat Model of Adenine-Induced Chronic Kidney Disease. Med. Sci. Monit. 2018, 24, 4760–4766. [Google Scholar] [CrossRef]
- Pan, Y.; Zhu, G.; Wang, Y.; Cai, L.; Cai, Y.; Hu, J.; Li, Y.; Yan, Y.; Wang, Z.; Li, X.; et al. Inhibition of high glucose-induced inflammatory response and macrophage infiltration by a novel curcumin derivative prevents renal injury in diabetic rats. Br. J. Pharm. 2012, 166, 1169–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soetikno, V.; Watanabe, K.; Sari, F.R.; Harima, M.; Thandavarayan, R.A.; Veeraveedu, P.T.; Arozal, W.; Sukumaran, V.; Lakshmanan, A.P.; Arumugam, S.; et al. Curcumin attenuates diabetic nephropathy by inhibiting PKC-α and PKC-β1 activity in streptozotocin-induced type I diabetic rats. Mol. Nutr. Food Res. 2011, 55, 1655–1665. [Google Scholar] [CrossRef] [PubMed]
- Manabe, I. Chronic inflammation links cardiovascular, metabolic and renal diseases. Circ. J. 2011, 75, 2739–2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barzegar, A.; Moosavi-Movahedi, A.A. Intracellular ROS protection efficiency and free radical-scavenging activity of curcumin. PLoS ONE 2011, 6, e26012. [Google Scholar] [CrossRef] [Green Version]
- Jeong, G.S.; Oh, G.S.; Pae, H.O.; Jeong, S.O.; Kim, Y.C.; Shin, M.K.; Seo, B.Y.; Han, S.Y.; Lee, H.S.; Jeong, J.G.; et al. Comparative effects of curcuminoids on endothelial heme oxygenase-1 expression: Ortho-methoxy groups are essential to enhance heme oxygenase activity and protection. Exp. Mol. Med. 2006, 38, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Ye, S.F.; Hou, Z.Q.; Zhong, L.M.; Zhang, Q.Q. Effect of curcumin on the induction of glutathione S-transferases and NADP(H): Quinone oxidoreductase and its possible mechanism of action. Yao Xue Xue Bao 2007, 42, 376–380. [Google Scholar]
- Rushworth, S.A.; Ogborne, R.M.; Charalambos, C.A.; O’Connell, M.A. Role of protein kinase C delta in curcumin-induced antioxidant response element-mediated gene expression in human monocytes. Biochem. Biophys. Res. Commun. 2006, 341, 1007–1016. [Google Scholar] [CrossRef]
- Tapia, E.; Zatarain-Barrón, Z.L.; Hernández-Pando, R.; Zarco-Márquez, G.; Molina-Jijón, E.; Cristóbal-García, M.; Santamaría, J.; Pedraza-Chaverri, J. Curcumin reverses glomerular hemodynamic alterations and oxidant stress in 5/6 nephrectomized rats. Phytomedicine 2013, 20, 359–366. [Google Scholar] [CrossRef]
- Tapia, E.; Soto, V.; Ortiz-Vega, K.M.; Zarco-Márquez, G.; Molina-Jijón, E.; Cristóbal-García, M.; Santamaría, J.; García-Niño, W.R.; Correa, F.; Zazueta, C.; et al. Curcumin induces Nrf2 nuclear translocation and prevents glomerular hypertension, hyperfiltration, oxidant stress and the decrease in antioxidant enzymes in 5/6 nephrectomized rats. Oxid. Med. Cell. Longev 2012, 2012, 269039. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Trejo, O.E.; Tapia, E.; Molina-Jijón, E.; Medina-Campos, O.N.; Macías-Ruvalcaba, N.A.; León-Contreras, J.C.; Hernández-Pando, R.; García-Arroyo, F.E.; Cristóbal, M.; Sánchez-Lozada, L.G.; et al. Curcumin prevents mitochondrial dynamics disturbances in early 5/6 nephrectomy: Relation to oxidative stress and mitochondrial bioenergetics. Biofactors 2017, 43, 293–310. [Google Scholar] [CrossRef]
- Bugyei-Twum, A.; Abadeh, A.; Thai, K.; Zhang, Y.; Mitchell, M.; Kabir, G.; Connelly, K.A. Suppression of NLRP3 Inflammasome Activation Ameliorates Chronic Kidney Disease-Induced Cardiac Fibrosis and Diastolic Dysfunction. Sci. Rep. 2016, 6, 39551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.S.; Gehr, T.W.B.; Ghosh, S. Curcumin and Chronic Kidney Disease (CKD): Major Mode of Action through Stimulating Endogenous Intestinal Alkaline Phosphatase. Molecules 2014, 19, 20139–20156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldanha, J.F.; Leal, V.O.; Rizzetto, F.; Grimmer, G.H.; Ribeiro-Alves, M.; Daleprane, J.B.; Carraro-Eduardo, J.C.; Mafra, D. Effects of Resveratrol Supplementation in Nrf2 and NF-κB Expressions in Nondialyzed Chronic Kidney Disease Patients: A Randomized, Double-Blind, Placebo-Controlled, Crossover Clinical Trial. J. Ren. Nutr. 2016, 26, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Cha, Y.N.; Surh, Y.J. A protective role of nuclear factor-erythroid 2-related factor-2 (Nrf2) in inflammatory disorders. Mutat. Res. 2010, 690, 12–23. [Google Scholar] [CrossRef]
- Castilla, P.; Dávalos, A.; Teruel, J.L.; Cerrato, F.; Fernández-Lucas, M.; Merino, J.L.; Sánchez-Martín, C.C.; Ortuño, J.; Lasunción, M.A. Comparative effects of dietary supplementation with red grape juice and vitamin E on production of superoxide by circulating neutrophil NADPH oxidase in hemodialysis patients. Am. J. Clin. Nutr. 2008, 87, 1053–1061. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Tian, S.; Han, J.; Xiong, P. Resveratrol as a therapeutic agent for renal fibrosis induced by unilateral ureteral obstruction. Ren. Fail. 2014, 36, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Cheng, K.; Song, Z.; Chen, Y.; Li, S.; Zhang, Y.; Zhang, H.; Zhang, L.; Wang, C.; Wang, T. Resveratrol Protects Against Renal Damage via Attenuation of Inflammation and Oxidative Stress in High-Fat-Diet-Induced Obese Mice. Inflammation 2019, 42, 937–945. [Google Scholar] [CrossRef]
- Palsamy, P.; Subramanian, S. Resveratrol protects diabetic kidney by attenuating hyperglycemia-mediated oxidative stress and renal inflammatory cytokines via Nrf2-Keap1 signaling. Biochim. Biophys. Acta 2011, 1812, 719–731. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.J.; Sun, Y.N.; Chen, S.J.; Liu, S.; Jiang, G.R. Resveratrol attenuates skeletal muscle atrophy induced by chronic kidney disease via MuRF1 signaling pathway. Biochem. Biophys. Res. Commun. 2017, 487, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Zhu, R.; Chen, Y.P.; Deng, Y.Y.; Zheng, R.; Zhong, Y.F.; Wang, L. Cordyceps cicadae extracts ameliorate renal malfunction in a remnant kidney model. J. Zhejiang Univ. Sci. 2011, 12, 1024–1033. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Weng, S.C.; Chou, C.J.; Chang, T.T.; Tsai, W.J. Activation and proliferation signals in primary human T lymphocytes inhibited by ergosterol peroxide isolated from Cordyceps cicadae. Br. J. Pharmacol. 2003, 140, 895–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, M.Y.; Chen, C.C.; Lee, L.Y.; Lin, T.W.; Kuo, C.F. N(6)-(2-Hydroxyethyl)adenosine in the Medicinal Mushroom Cordyceps cicadae Attenuates Lipopolysaccharide-Stimulated Pro-inflammatory Responses by Suppressing TLR4-Mediated NF-κB Signaling Pathways. J. Nat. Prod. 2015, 78, 2452–2460. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.H.; Chen, Y.P.; Deng, Y.Y. The mechanism study of Cordyceps sobolifera mycelium preventing the progression of glomerulosclerosis. Chin. J. Integr. Tradit. West. Nephrol. 2005, 6, 132–136. [Google Scholar]
- Wang, L.; Chen, Y. Effect of artificial Cordyceps cicadae on proliferation and mesangial matrix production in human glomerulus mesangial cell. Tradit. Chin. Med. Res. 2006, 19, 9–11. [Google Scholar]
- Sun, T.; Dong, W.; Jiang, G.; Yang, J.; Liu, J.; Zhao, L.; Ma, P. Cordyceps militaris Improves Chronic Kidney Disease by Affecting TLR4/NF-κB Redox Signaling Pathway. Oxid. Med. Cell. Longev 2019, 2019, 7850863. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.X.; Hu, D.; Lam, S.C.; Ge, L.; Wu, D.; Zhao, J. Comparison of antioxidant activities of different parts from snow chrysanthemum (Coreopsis tinctoria Nutt.) and identification of their natural antioxidants using high performance liquid chromatography coupled with diode array detection and mass spectrometry and 2,2′-azinobis (3-ethylbenzthiazoline-sulfonic acid) diammonium salt-based assay. J. Chromatogr. A 2016, 1428, 134–142. [Google Scholar]
- Li, Y.; Chen, X.; Xue, J.; Liu, J.; Chen, X.; Wulasihan, M. Flavonoids from Coreopsis tinctoria adjust lipid metabolism in hyperlipidemia animals by down-regulating adipose differentiation-related protein. Lipids Health Dis. 2014, 13, 193–211. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Shi, S.; Zhao, M.; Chai, X.; Tu, P. Coreosides A-D, C14-polyacetylene glycosides from the capitula of Coreopsis tinctoria and its anti-inflammatory activity against Cox-2. Fitoterapia 2013, 87, 93–97. [Google Scholar] [CrossRef]
- Lin, Y.C.; Wu, M.S.; Lin, Y.F.; Chen, C.R.; Chen, C.Y.; Chen, C.J.; Shen, C.C.; Chen, K.C.; Peng, C.C. Nifedipine Modulates Renal Lipogenesis via the AMPK-SREBP Transcriptional Pathway. Int. J. Mol Sci. 2019, 20, 1570. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Li, L.; Li, X.; Li, H.; Zhang, Y.; Zhang, R. The anti-inflammatory and anti-fibrotic effects of Coreopsis tinctoria Nutt on high-glucose-fat diet and streptozotocin-induced diabetic renal damage in rat. BMC Complement Altern. Med. 2015, 15, 314. [Google Scholar] [CrossRef] [Green Version]
- Lan, Y.; Jie, L.; Linlin, L.; Xinxia, L.; Rui, Z.; Yujie, Z.; Xinmin, M. Coreopsis tinctoria Nutt ameliorates high glucose-induced renal fibrosis and inflammation via the TGF-β1/SMADS/AMPK/NF-κB pathways. BMC Complement Altern. Med. 2019, 19, 14. [Google Scholar]
- Karuna, R.; Reddy, S.; Baskar, R.; Saralakumari, D. Antioxidant potential of aqueous extract of Phyllanthus amarus in rats. Indian J. Pharmacol. 2009, 41, 64–67. [Google Scholar]
- Giribabu, N.; Visweswara Rao, P.; Kumar, K.P.; Muniandy, S.; Swapna Rekha, S.; Salleh, N. Aqueous Extract of Phyllanthus niruri Leaves Displays In Vitro Antioxidant Activity and Prevents the Elevation of Oxidative Stress in the Kidney of Streptozotocin-Induced Diabetic Male Rats. Evid. Based Complement. Altern. Med. 2014, 2014, 834815. [Google Scholar] [CrossRef] [Green Version]
- Adejuwon, A.A.; Adokiye, S.B. Protective effect of the aqueous leaf and seed extract of Phyllanthus amarus on gentamicin and acetaminophen-induced nephrotoxic rats. J. Ethnopharmacol. 2008, 118, 318–323. [Google Scholar]
- Elkayam, A.; Peleg, E.; Grossman, E.; Shabtay, Z.; Sharabi, Y. Effects of allicin on cardiovascular risk factors in spontaneously hypertensive rats. Isr. Med. Assoc. J. 2013, 15, 170–173. [Google Scholar] [PubMed]
- Sun, X.; Ku, D.D. Allicin in garlic protects against coronary endothelial dysfunction and right heart hypertrophy in pulmonary hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H2431–H2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Trejo, E.M.; Arellano-Buendía, A.S.; Argüello-García, R.; Loredo-Mendoza, M.L.; García-Arroyo, F.E.; Arellano-Mendoza, M.G.; Castillo-Hernández, M.C.; Guevara-Balcázar, G.; Tapia, E.; Sánchez-Lozada, L.G.; et al. Effects of Allicin on Hypertension and Cardiac Function in Chronic Kidney Disease. Oxid. Med. Cell. Longev. 2016, 2016, 3850402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García Trejo, E.M.Á.; Arellano Buendía, A.S.; Sánchez Reyes, O.; García Arroyo, F.E.; Arguello García, R.; Loredo Mendoza, M.L.; Tapia, E.; Sánchez Lozada, L.G.; Osorio Alonso, H. The Beneficial Effects of Allicin in Chronic Kidney Disease Are Comparable to Losartan. Int. J. Mol. Sci. 2017, 18, 1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J. Pharmacology of oleanolic acid and ursolic acid. J. Ethnopharmacol. 1995, 49, 57–68. [Google Scholar] [CrossRef]
- Kunkel, S.D.; Suneja, M.; Ebert, S.M.; Bongers, K.S.; Fox, D.K.; Malmberg, S.E. mRNA expression signatures of human skeletal muscle atrophy identify a natural compound that increases muscle mass. Cell Metab. 2011, 13, 627–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenvinkel, P.; Ketteler, M.; Johnson, R.J.; Lindholm, B.; Pecoits-Filho, R.; Riella, M. IL-10, IL-6 and TNF-alpha: Central factors in the altered cytokine network of uremia–the good, the bad and the ugly. Kidney Int. 2005, 67, 1216–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Rajan, V.; Lin, E.; Hu, Z.; Han, H.Q.; Zhou, X. Pharmacological inhibition of myostatin suppresses systemic inflammation and muscle atrophy in mice with chronic kidney disease. FASEB J. 2011, 25, 1653–1663. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Chen, J.; Xu, J.; Cao, J.; Wang, Y.; Thomas, S.S.; Hu, Z. Suppression of muscle wasting by the plant-derived compound ursolic acid in a model of chronic kidney disease. J. Cachexia Sarcopenia Muscle 2017, 8, 327–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, X.; Chen, W.; Cui, J.; Yi, S.; Zhang, Z.; Li, K. Effects of ursolic acid on liver-protection and bile secretion. J. Chin. Med. Mater. 2003, 26, 578–581. [Google Scholar]
- Su, H.H.; Kao, C.M.; Lin, Y.C.; Lin, Y.C.; Kao, C.C.; Chen, H.H.; Hsu, C.C.; Chen, K.C.; Peng, C.C.; Wu, M.S. Relationship between serum total bilirubin levels and mortality in uremia patients undergoing long-term hemodialysis: A nationwide cohort study. Atherosclerosis 2017, 265, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Higdon, J.V.; Frei, B. Tea catechins and polyphenols: Health effects, metabolism and antioxidant functions. Crit. Rev. Food Sci. Nutr. 2003, 43, 89–143. [Google Scholar] [CrossRef]
- Bao, H.; Peng, A. The Green Tea Polyphenol(-)-epigallocatechin-3-gallate and its beneficial roles in chronic kidney disease. J. Transl. Int. Med. 2016, 4, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Tsai, P.Y.; Ka, S.M.; Chang, J.M.; Chen, H.C.; Shui, H.A.; Li, C.Y.; Hua, K.F.; Chang, W.L.; Huang, J.J.; Yang, S.S.; et al. Epigallocatechin-3-gallate prevents lupus nephritis development in mice via enhancing the Nrf2 antioxidant pathway and inhibiting NLRP3 inflammasome activation. Free Radic. Biol. Med. 2011, 51, 744–754. [Google Scholar] [CrossRef]
- Ye, T.; Zhen, J.; Du, Y.; Zhou, J.K.; Peng, A.; Vaziri, N.D.; Mohan, C.; Xu, Y.; Zhou, X.J. Green tea polyphenol (−)-epigallocatechin-3-gallate restores Nrf2 activity and ameliorates crescentic glomerulonephritis. PLoS ONE 2015, 10, e0119543. [Google Scholar] [CrossRef] [Green Version]
- Yamabe, N.; Yokozawa, T.; Oya, T.; Kim, M. Therapeutic potential of (-)-epigallocatechin 3-O-gallate on renal damage in diabetic nephropathy model rats. J. Pharmacol. Exp. Ther. 2006, 319, 228–236. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, T.; Yokozawa, T.; Sano, M.; Takeuchi, S.; Kim, M.; Minamoto, S. Activity of (-)-epigallocatechin 3-O-gallate against oxidative stress in rats with adenine-induced renal failure. J. Agric. Food Chem. 2004, 52, 2103–2107. [Google Scholar] [CrossRef] [PubMed]
- Kakuta, Y.; Okumi, M.; Isaka, Y.; Tsutahara, K.; Abe, T.; Yazawa, K.; Ichimaru, N.; Matsumura, K.; Hyon, S.H.; Takahara, S.; et al. Epigallocatechin-3-gallate protects kidneys from ischemia reperfusion injury by HO-1 upregulation and inhibition of macrophage infiltration. Transpl. Int. 2011, 24, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Satirapoj, B.; Vongwattana, P.; Supasyndh, O. Very low protein diet plus ketoacid analogs of essential amino acids supplement to retard chronic kidney disease progression. Kidney Res. Clin. Pr. 2018, 37, 384–392. [Google Scholar] [CrossRef] [Green Version]
- Koppe, L.; Fouque, D.; Soulage, C.O. The Role of Gut Microbiota and Diet on Uremic Retention Solutes Production in the Context of Chronic Kidney Disease. Toxins (Basel) 2018, 10, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Anti-Inflammatory Activity | Antioxidant Activity | Experimental Model and References | |
|---|---|---|---|
| ↑Magnesium | ↓CRP | Human study; [84] | |
| ↑Selenium | ↑GSH-Px ↓ROS | Human study; [87] | |
| ↓Phosphate | ↓FGF23 ↓PTH | Human study; [90,92,123,124] | |
| ↑Vitamin C | ↑hydroxylase/monooxygenase co-factor | ↓MDA ↓tissue lipid oxidation ↑tissue GSH ↑Co-antioxidant vit.E ↓ROS | Human study; [98] Animal model; [99] |
| ↑Vitamin E | ↓8-OHdG | ↓lipid peroxidation ↓NADPH activity ↓inflammatory mediators | Cellular and animal model; [101] Animal model; [102] Human study; [108] |
| ↑Vitamin D | ↓NF-kB signling pathway ↓RAAS ↓TGF-β/Smad ↓Wnt/β-catenin | Animal model; [114] Cellular and animal model; [115] Animal model; [116] Cellular and animal model; [117] | |
| ↑Vitamin A | ↓TLR4/NF-kB signling pathway | Animal model; [118] | |
| ↑Vitamin B1 | ↓plasma lactate levels | Human study; [119] |
| Anti-Inflammatory Activity | Antioxidant Activity | Experimental Model and References | |
|---|---|---|---|
| Quercetin | ↓FGF23 ↓PTH | ↓LDH ↓SOD | Animal model; [130] |
| Curcumin | ↓VEGF ↓TGF-β ↓CTGF ↓fibronectin and collagen IV ↓iNOS and ↓COX-2 ↓TNF-α ↓MCP-1 ↓JNK/NF-κB ↓NLRP3 | ↓NO and ↓ONOO− ↓O2− and ↓H2O2 ↑SOD ↑CAT ↑GSR ↑HO-1 ↑GST ↑NQO1 ↑GCL | Animal model; [131,132] Cellular model; [133,134,135,136,137] Animal model; [138,139,140,141] |
| Resveratrol | ↓NF-κB ↓pro-inflammatory cytokines and enzymes | ↑Nrf2 ↑antioxidant enzymes | Human study; [143,145] Animal model; [146,147,148] |
| Cordycepin | ↓NF-κB ↓TNF-α ↓IL-6 ↓IL-1β ↓TGF-β1/Smad | Animal model; [153] Cellular model; [154] Human study and cellular model; [155] | |
| Flavonoids of C. tinctoria | ↓NF-κB signling pathway ↓COX-2 ↓MCP-1 and ↓collagen IV ↓AMPK ↓TGF-β/Smad | Animal model; [158,160] Cellular model; [161] | |
| Flavonoids and polyphenols of P. niruri | ↑SOD ↑CAT ↑GPx | Animal model; [164] | |
| Allicin | ↓AT1R ↓Keap1 ↓eNOS ↑Nrf2 ↑SOD ↑CAT ↑GPx | Animal model; [167,168] | |
| Ursolic acid | ↓IL-6 ↓NF-κB ↓p-STAT3 ↓C/EBP-δ | Human study and animal model; [170] Animal model; [172] Cellular and animal model; [173] | |
| Epigallocatechin-3-gallate | ↓NF-κB ↓NLRP3 ↓caspase-1 ↓IL-1β and IL-18 ↓p-Akt ↓p-JNK ↓p-ERK1/2 ↓p-P38 ↑PPARγ and ↑SIRT1 ↓MCP-1 ↓TGF-β | ↑Nrf2 ↑GPx ↑HO-1 ↓AGE and lipid peroxidation ↓ROS | Cellular model; [176] Animal model; [177,178,179,180,181,182] |
| Reduction of Progression of CKD | CKD-Complications Management | References | |
|---|---|---|---|
| Magnesium | ✓ | ✓ | [82,83,84,85] |
| Selenium | ✓ | [87] | |
| Phosphate | ✓ | [89] | |
| Vitamin C | ✓ | [96,97,98] | |
| Vitamin E | ✓ | [102,103,104,105] | |
| Vitamin D | ✓ | ✓ | [111,112,113,114,115,116,117] |
| Vitamin A | ✓ | [118] | |
| Vitamin B1 | ✓ | ✓ | [122] |
| Quercetin | ✓ | [129,130] | |
| Curcumin | ✓ | ✓ | [131,132,133,134,135,136,137,138,139,140,141,142] |
| Resveratrol | ✓ | ✓ | [146,147,148,149] |
| Cordycepin | ✓ | [153,154,155] | |
| C. tinctoria | ✓ | [160,161] | |
| P. niruri | ✓ | [164] | |
| Allicin | ✓ | ✓ | [166,167,168] |
| Ursolic acid | ✓ | [170,171,172,173,174,175] | |
| Epigallocatechin-3-gallate | ✓ | ✓ | [177,178,179,180,181,182] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rapa, S.F.; Di Iorio, B.R.; Campiglia, P.; Heidland, A.; Marzocco, S. Inflammation and Oxidative Stress in Chronic Kidney Disease—Potential Therapeutic Role of Minerals, Vitamins and Plant-Derived Metabolites. Int. J. Mol. Sci. 2020, 21, 263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010263
Rapa SF, Di Iorio BR, Campiglia P, Heidland A, Marzocco S. Inflammation and Oxidative Stress in Chronic Kidney Disease—Potential Therapeutic Role of Minerals, Vitamins and Plant-Derived Metabolites. International Journal of Molecular Sciences. 2020; 21(1):263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010263
Chicago/Turabian StyleRapa, Shara Francesca, Biagio Raffaele Di Iorio, Pietro Campiglia, August Heidland, and Stefania Marzocco. 2020. "Inflammation and Oxidative Stress in Chronic Kidney Disease—Potential Therapeutic Role of Minerals, Vitamins and Plant-Derived Metabolites" International Journal of Molecular Sciences 21, no. 1: 263. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21010263