The Dual-Active Histamine H3 Receptor Antagonist and Acetylcholine Esterase Inhibitor E100 Alleviates Autistic-Like Behaviors and Oxidative Stress in Valproic Acid Induced Autism in Mice

 , , , , , , and
, , , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of E100 on Sociability Impairments in Three-Chamber Behaviour (TCB) Test and Stereotyped Repetitive Behavior in Marble-Burying Behaviour (MBB)
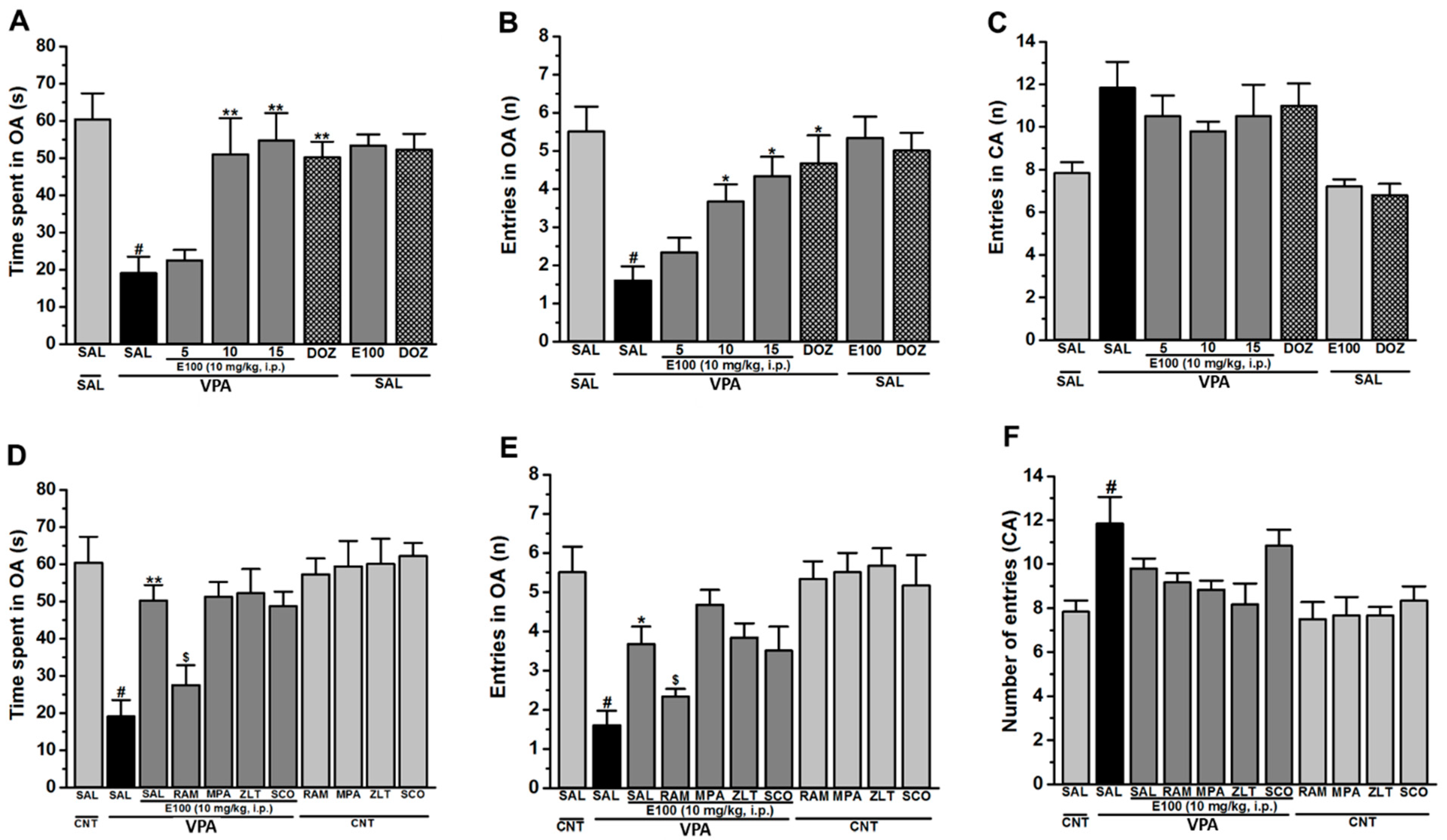
2.2. Effects of E100 on Anxiety Levels and Locomotor Activity of Valproic Acid (VPA)-Exposed Mice in Elevated Plus Maze (EPM) Test
2.3. Effect of E100 on Oxidative Stress Levels in Cerebellar Tissues of VPA-Exposed Mice
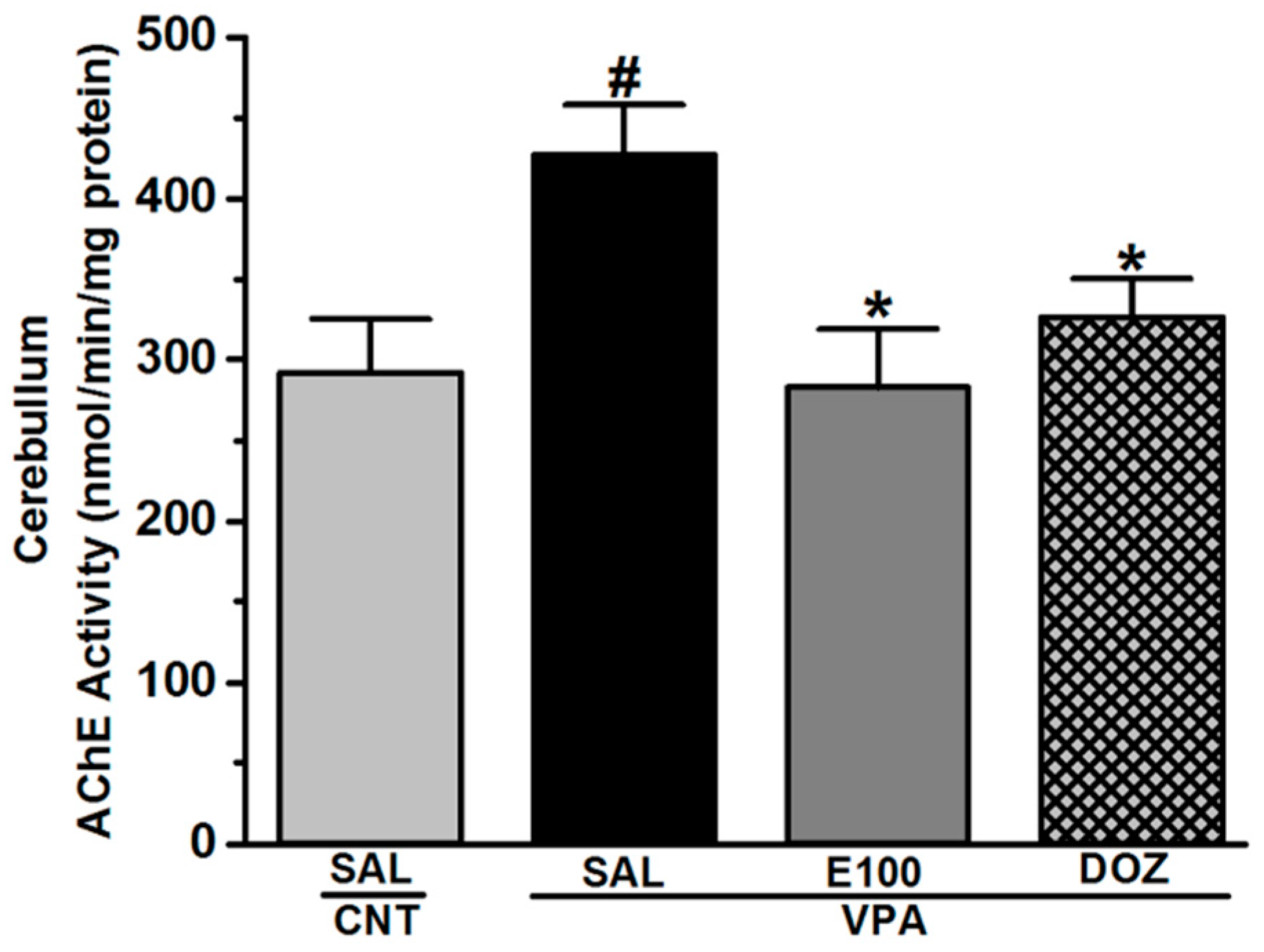
2.4. Effect of E100 on Acetylcholine Esterase Activity in Cerebellum Tissues of VPA-Exposed Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs
4.3. Study Design and Treatments
4.3.1. Prenatal Treatment
4.3.2. Postnatal Treatments
4.4. Behavioral Tests
4.4.1. Three-Chamber Behavior (TCB)
4.4.2. Marble Burying Behavior (MBB)
4.4.3. Elevated Plus Maze (EPM) Test
4.5. Biochemical Assessments
4.5.1. Brain Collection and Tissue Preparation for Biochemical Studies
4.5.2. Oxidative Stress Marker Estimations
Lipid Peroxidation Estimation
Glutathione (GSH) Estimation
Estimation of Antioxidant Enzymes Activity
Determination of Acetylcholinesterase (AChE) Activity in VPA-Exposed Mice Cerebellum
4.6. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Baronio, D.; Castro, K.; Gonchoroski, T.; de Melo, G.M.; Nunes, G.D.; Bambini-Junior, V.; Gottfried, C.; Riesgo, R. Effects of an H3R antagonist on the animal model of autism induced by prenatal exposure to valproic acid. PLoS ONE 2015, 10, e0116363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arvidsson, O.; Gillberg, C.; Lichtenstein, P.; Lundstrom, S. Secular changes in the symptom level of clinically diagnosed autism. J. Child. Psychol. Psychiatry 2018, 59, 744–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nestler, E.J.; Hyman, S.E. Animal models of neuropsychiatric disorders. Nat. Neurosci. 2010, 13, 1161–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissa, N.; Al-Houqani, M.; Sadeq, A.; Ojha, S.K.; Sasse, A.; Sadek, B. Current Enlightenment about etiology and pharmacological treatment of autism spectrum disorder. Front. Neurosci. 2018, 12, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldrick, R.C.; Carter, A.S. State-level trends in the prevalence of Autism Spectrum Disorder (ASD) from 2000 to 2012: A reanalysis of findings from the autism and developmental disabilities network. J. Autism Dev. Disord. 2018, 48, 3086–3092. [Google Scholar] [CrossRef]
- Xu, G.; Strathearn, L.; Liu, B.; Bao, W. Prevalence of Autism Spectrum Disorder among US children and adolescents, 2014–2016. JAMA 2018, 319, 81–82. [Google Scholar] [CrossRef] [Green Version]
- Cavalli, A.; Bolognesi, M.L.; Minarini, A.; Rosini, M.; Tumiatti, V.; Recanatini, M.; Melchiorre, C. Multi-target-directed ligands to combat neurodegenerative diseases. J. Med. Chem. 2008, 51, 347–372. [Google Scholar] [CrossRef]
- Decker, M. Recent advances in the development of hybrid molecules/designed multiple compounds with antiamnesic properties. Mini Rev. Med. Chem. 2007, 7, 221–229. [Google Scholar] [CrossRef]
- Karimi, P.; Kamali, E.; Mousavi, S.M.; Karahmadi, M. Environmental factors influencing the risk of autism. Off. J. Isfahan Univ. Med. Sci. 2017, 22, 27. [Google Scholar] [CrossRef]
- Ellenbroek, B.A.; Ghiabi, B. Do Histamine receptor 3 antagonists have a place in the therapy for schizophrenia? Curr. Pharm. Des. 2015, 21, 3760–3770. [Google Scholar] [CrossRef]
- Bacchelli, E.; Battaglia, A.; Cameli, C.; Lomartire, S.; Tancredi, R.; Thomson, S.; Sutcliffe, J.S.; Maestrini, E. Analysis of CHRNA7 rare variants in autism spectrum disorder susceptibility. Am. J. Med. Genet. A 2015, 167A, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Hellings, J.A.; Arnold, L.E.; Han, J.C. Dopamine antagonists for treatment resistance in autism spectrum disorders: Review and focus on BDNF stimulators loxapine and amitriptyline. Expert Opin. Pharmacother. 2017, 18, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Naaijen, J.; Bralten, J.; Poelmans, G.; Glennon, J.C.; Franke, B.; Buitelaar, J.K. Glutamatergic and GABAergic gene sets in attention-deficit/hyperactivity disorder: Association to overlapping traits in ADHD and autism. Transl. Psychiatry 2017, 7, e999. [Google Scholar] [CrossRef] [PubMed]
- Nakai, N.; Nagano, M.; Saitow, F.; Watanabe, Y.; Kawamura, Y.; Kawamoto, A.; Tamada, K.; Mizuma, H.; Onoe, H.; Watanabe, Y.; et al. Serotonin rebalances cortical tuning and behavior linked to autism symptoms in 15q11-13 CNV mice. Sci. Adv. 2017, 3, e1603001. [Google Scholar] [CrossRef] [Green Version]
- Pavăl, D. Dopamine hypothesis of autism spectrum disorder. Dev. Neurosci. 2017, 39, 355–360. [Google Scholar] [CrossRef]
- Pavăl, D.; Rad, F.; Rusu, R.; Niculae, A.S.; Colosi, H.A.; Dobrescu, I.; Dronca, E. Low retinal dehydrogenase 1 (RALDH1) Level in prepubertal boys with autism spectrum disorder: A Possible link to dopamine dysfunction? Clin. Psychopharmacol. Neurosci. 2017, 15, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.; Wing, L. Psychological approaches to chronic catatonia-like deterioration in autism spectrum disorders. Int. Rev. Neurobiol. 2006, 72, 245–264. [Google Scholar] [CrossRef]
- Wang, L.; Almeida, L.E.; Spornick, N.A.; Kenyon, N.; Kamimura, S.; Khaibullina, A.; Nouraie, M.; Quezado, Z.M. Modulation of social deficits and repetitive behaviors in a mouse model of autism: The role of the nicotinic cholinergic system. Psychopharmacology 2015, 232, 4303–4316. [Google Scholar] [CrossRef]
- Chen, R.; Davis, L.K.; Guter, S.; Wei, Q.; Jacob, S.; Potter, M.H.; Cox, N.J.; Cook, E.H.; Sutcliffe, J.S.; Li, B. Leveraging blood serotonin as an endophenotype to identify de novo and rare variants involved in autism. Mol. Autism 2017, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Hellmer, K.; Nystrom, P. Infant acetylcholine, dopamine, and melatonin dysregulation: Neonatal biomarkers and causal factors for ASD and ADHD phenotypes. Med. Hypotheses 2017, 100, 64–66. [Google Scholar] [CrossRef]
- Arnold, H.M.; Burk, J.A.; Hodgson, E.M.; Sarter, M.; Bruno, J.P. Differential cortical acetylcholine release in rats performing a sustained attention task versus behavioral control tasks that do not explicitly tax attention. Neuroscience 2002, 114, 451–460. [Google Scholar] [CrossRef]
- Ragozzino, M.E.; Pal, S.N.; Unick, K.; Stefani, M.R.; Gold, P.E. Modulation of hippocampal acetylcholine release and spontaneous alternation scores by intrahippocampal glucose injections. J. Neurosci. 1998, 18, 1595–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avale, M.E.; Chabout, J.; Pons, S.; Serreau, P.; De Chaumont, F.; Olivo-Marin, J.C.; Bourgeois, J.P.; Maskos, U.; Changeux, J.P.; Granon, S. Prefrontal nicotinic receptors control novel social interaction between mice. FASEB J. 2011, 25, 2145–2155. [Google Scholar] [CrossRef] [PubMed]
- McConville, B.J.; Sanberg, P.R.; Fogelson, M.H.; King, J.; Cirino, P.; Parker, K.W.; Norman, A.B. The effects of nicotine plus haloperidol compared to nicotine only and placebo nicotine only in reducing tic severity and frequency in Tourette’s disorder. Biol. Psychiatry 1992, 31, 832–840. [Google Scholar] [CrossRef]
- Karvat, G.; Kimchi, T. Acetylcholine elevation relieves cognitive rigidity and social deficiency in a mouse model of autism. Neuropsychopharmacology 2014, 39, 831–840. [Google Scholar] [CrossRef]
- Kemper, T.L.; Bauman, M. Neuropathology of infantile autism. J. Neuropathol. Exp. Neurol. 1998, 57, 645–652. [Google Scholar] [CrossRef] [Green Version]
- Friedman, S.D.; Shaw, D.W.; Artru, A.A.; Dawson, G.; Petropoulos, H.; Dager, S.R. Gray and white matter brain chemistry in young children with autism. Arch. Gen. Psychiatry 2006, 63, 786–794. [Google Scholar] [CrossRef]
- Mukaetova-Ladinska, E.B. Silent lives: Why do we fail community-dwelling people with dementia? Age Ageing 2017, 46, 341–343. [Google Scholar] [CrossRef] [Green Version]
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.; Stark, H.; Thurmond, R.L.; Haas, H.L. International union of basic and clinical pharmacology. XCVIII. Histamine receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panula, P.; Rinne, J.; Kuokkanen, K.; Eriksson, K.S.; Sallmen, T.; Kalimo, H.; Relja, M. Neuronal histamine deficit in Alzheimer’s disease. Neuroscience 1998, 82, 993–997. [Google Scholar] [CrossRef]
- Sadek, B.; Stark, H. Cherry-picked ligands at histamine receptor subtypes. Neuropharmacology 2016, 106, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Sadek, B.; Saad, A.; Sadeq, A.; Jalal, F.; Stark, H. Histamine H3 receptor as a potential target for cognitive symptoms in neuropsychiatric diseases. Behav. Brain Res. 2016, 312, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Arrang, J.M.; Garbarg, M.; Schwartz, J.C. Auto-inhibition of brain histamine release mediated by a novel class (H3) of histamine receptor. Nature 1983, 302, 832–837. [Google Scholar] [CrossRef]
- Lovenberg, T.W.; Roland, B.L.; Wilson, S.J.; Jiang, X.; Pyati, J.; Huvar, A.; Jackson, M.R.; Erlander, M.G. Cloning and functional expression of the human histamine H3 receptor. Mol. Pharmacol. 1999, 55, 1101–1107. [Google Scholar] [CrossRef]
- Berlin, M.; Boyce, C.W.; Ruiz Mde, L. Histamine H3 receptor as a drug discovery target. J. Med. Chem. 2011, 54, 26–53. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, R.; Anaclet, C.; Guhennec, C.; Brousseau, E.; Bricout, D.; Giboulot, T.; Bozyczko-Coyne, D.; Spiegel, K.; Ohtsu, H.; Williams, M.; et al. The brain H3-receptor as a novel therapeutic target for vigilance and sleep-wake disorders. Biochem. Pharmacol. 2007, 73, 1157–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alachkar, A.; Khan, N.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. Histamine H3 receptor antagonist E177 attenuates amnesia induced by dizocilpine without modulation of anxiety-like behaviors in rats. Neuropsychiatr. Dis. Treat. 2019, 15, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Alachkar, A.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. The histamine H3 Receptor antagonist E159 Reverses memory deficits induced by dizocilpine in passive avoidance and novel object recognition paradigm in rats. Front. Pharmacol. 2017, 8, 709. [Google Scholar] [CrossRef] [Green Version]
- Alachkar, A.; Azimullah, S.; Ojha, S.K.; Beiram, R.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. The neuroprotective effects of histamine H3 Receptor antagonist E177 on pilocarpine-induced status epilepticus in rats. Molecules 2019, 24. [Google Scholar] [CrossRef] [Green Version]
- Rossi, P.G.; Posar, A.; Parmeggiani, A.; Pipitone, E.; D’Agata, M. Niaprazine in the treatment of autistic disorder. J. Child Neurol. 1999, 14, 547–550. [Google Scholar] [CrossRef]
- Adams-Chapman, I.; Stoll, B.J. Neonatal infection and long-term neurodevelopmental outcome in the preterm infant. Curr. Opin. Infect. Dis. 2006, 19, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Lucchina, L.; Depino, A.M. Altered peripheral and central inflammatory responses in a mouse model of autism. Autism Res. 2014, 7, 273–289. [Google Scholar] [CrossRef] [PubMed]
- DeLorey, T.M.; Sahbaie, P.; Hashemi, E.; Homanics, G.E.; Clark, J.D. Gabrb3 gene deficient mice exhibit impaired social and exploratory behaviors, deficits in non-selective attention and hypoplasia of cerebellar vermal lobules: A potential model of autism spectrum disorder. Behav. Brain Res. 2008, 187, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.A.; Goldowitz, D.; Mittleman, G. Repetitive behavior and increased activity in mice with Purkinje cell loss: A model for understanding the role of cerebellar pathology in autism. Eur. J. Neurosci. 2010, 31, 544–555. [Google Scholar] [CrossRef] [Green Version]
- Koziol, L.F.; Budding, D.; Andreasen, N.; D’arrigo, S.; Bulgheroni, S.; Imamizu, H.; Ito, M.; Manto, M.; Marvel, C.; Parker, K.; et al. Consensus paper: The cerebellum’s role in movement and cognition. Cerebellum 2014, 13, 151–177. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.H.; Kloth, A.D.; Badura, A. The cerebellum, sensitive periods, and autism. Neuron 2014, 83, 518–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Smith, S.E.; Malkova, N.; Tse, D.; Su, Y.; Patterson, P.H. Activation of the maternal immune system alters cerebellar development in the offspring. Brain Behav. Immun. 2009, 23, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Miller, H.L.; Ragozzino, M.E.; Cook, E.H.; Sweeney, J.A.; Mosconi, M.W. Cognitive set shifting deficits and their relationship to repetitive behaviors in autism spectrum disorder. J. Autism Dev. Disord. 2015, 45, 805–815. [Google Scholar] [CrossRef]
- Gadad, B.S.; Hewitson, L.; Young, K.A.; German, D.C. Neuropathology and animal models of autism: Genetic and environmental factors. Autism Res. Treat. 2013, 2013, 731935. [Google Scholar] [CrossRef]
- Fernández, M.; Sierra-Arregui, T.; Peñagarikano, O. The Cerebellum and autism: More than motor control. Behav. Neurosci. 2019. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhu, J.N.; Wang, J.J. Histaminergic afferent system in the cerebellum: Structure and function. Cerebellum Ataxias 2014, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Depino, A.M.; Lucchina, L.; Pitossi, F. Early and adult hippocampal TGF-β1 overexpression have opposite effects on behavior. Brain Behav. Immun. 2011, 25, 1582–1591. [Google Scholar] [CrossRef] [PubMed]
- Witkin, J.M.; Nelson, D.L. Selective histamine H3 receptor antagonists for treatment of cognitive deficiencies and other disorders of the central nervous system. Pharmacol. Ther. 2004, 103, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, H.; Onodera, K.; Iinuma, K.; Watanabe, T. Effect of thioperamide, a histamine H3 receptor antagonist, on electrically induced convulsions in mice. Eur. J. Pharmacol. 1993, 234, 129–133. [Google Scholar] [CrossRef]
- Bhowmik, M.; Khanam, R.; Vohora, D. Histamine H3 receptor antagonists in relation to epilepsy and neurodegeneration: A systemic consideration of recent progress and perspectives. Br. J. Pharmacol. 2012, 167, 1398–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, N.; Saad, A.; Nurulain, S.M.; Darras, F.H.; Decker, M.; Sadek, B. The dual-acting H3 receptor antagonist and AChE inhibitor UW-MD-71 dose-dependently enhances memory retrieval and reverses dizocilpine-induced memory impairment in rats. Behav. Brain Res. 2016, 297, 155–164. [Google Scholar] [CrossRef]
- Sadek, B.; Khan, N.; Darras, F.H.; Pockes, S.; Decker, M. The dual-acting AChE inhibitor and H3 receptor antagonist UW-MD-72 reverses amnesia induced by scopolamine or dizocilpine in passive avoidance paradigm in rats. Physiol. Behav. 2016, 165, 383–391. [Google Scholar] [CrossRef]
- Morini, G.; Comini, M.; Rivara, M.; Rivara, S.; Bordi, F.; Plazzi, P.V.; Flammini, L.; Saccani, F.; Bertoni, S.; Ballabeni, V.; et al. Synthesis and structure-activity relationships for biphenyl H3 receptor antagonists with moderate anti-cholinesterase activity. Bioorganic Med. Chem. 2008, 16, 9911–9924. [Google Scholar] [CrossRef]
- Nikolic, K.; Filipic, S.; Agbaba, D.; Stark, H. Procognitive properties of drugs with single and multitargeting H3 receptor antagonist activities. CNS Neurosci. Ther. 2014, 20, 613–623. [Google Scholar] [CrossRef]
- Petroianu, G.; Arafat, K.; Sasse, B.C.; Stark, H. Multiple enzyme inhibitions by histamine H3 receptor antagonists as potential procognitive agents. Pharmazie 2006, 61, 179–182. [Google Scholar]
- Bembenek, S.D.; Keith, J.M.; Letavic, M.A.; Apodaca, R.; Barbier, A.J.; Dvorak, L.; Aluisio, L.; Miller, K.L.; Lovenberg, T.W.; Carruthers, N.I. Lead identification of acetylcholinesterase inhibitors-histamine H3 receptor antagonists from molecular modeling. Bioorganic Med. Chem. 2008, 16, 2968–2973. [Google Scholar] [CrossRef]
- Sadek, B.; Saad, A.; Subramanian, D.; Shafiullah, M.; Lazewska, D.; Kiec-Kononowiczc, K. Anticonvulsant and procognitive properties of the non-imidazole histamine H3 receptor antagonist DL77 in male adult rats. Neuropharmacology 2016, 106, 46–55. [Google Scholar] [CrossRef]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Lazewska, D.; Kiec-Kononowicz, K.; et al. The dual-active histamine H3 receptor antagonist and acetylcholine esterase inhibitor E100 ameliorates stereotyped repetitive behavior and neuroinflammmation in sodium valproate induced autism in mice. Chem. Biol. Interact. 2019, 312, 108775. [Google Scholar] [CrossRef] [PubMed]
- Lazewska, D.; Jonczyk, J.; Bajda, M.; Szalaj, N.; Wieckowska, A.; Panek, D.; Moore, C.; Kuder, K.; Malawska, B.; Kiec-Kononowicz, K. Cholinesterase inhibitory activity of chlorophenoxy derivatives-Histamine H3 receptor ligands. Bioorganic Med. Chem. Lett. 2016, 26, 4140–4145. [Google Scholar] [CrossRef] [PubMed]
- Kuder, K.; Lazewska, D.; Latacz, G.; Schwed, J.S.; Karcz, T.; Stark, H.; Karolak-Wojciechowska, J.; Kiec-Kononowicz, K. Chlorophenoxy aminoalkyl derivatives as histamine H(3)R ligands and antiseizure agents. Bioorganic Med. Chem. 2016, 24, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Wright, C.; Shin, J.H.; Rajpurohit, A.; Deep-Soboslay, A.; Collado-Torres, L.; Brandon, N.J.; Hyde, T.M.; Kleinman, J.E.; Jaffe, A.E.; Cross, A.J.; et al. Altered expression of histamine signaling genes in autism spectrum disorder. Transl. Psychiatry 2017, 7, e1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, G.B.; Esbenshade, T.A.; Pan, J.B.; Radek, R.J.; Krueger, K.M.; Yao, B.B.; Browman, K.E.; Buckley, M.J.; Ballard, M.E.; Komater, V.A.; et al. Pharmacological properties of ABT-239 [4-(2-{2-[(2R)-2-Methylpyrrolidinyl]ethyl}-benzofuran-5-yl)benzonitrile]: II. Neurophysiological characterization and broad preclinical efficacy in cognition and schizophrenia of a potent and selective histamine H3 receptor antagonist. J. Pharmacol. Exp. Ther. 2005, 313, 176–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, G.B.; Pan, J.B.; Esbenshade, T.A.; Bennani, Y.L.; Black, L.A.; Faghih, R.; Hancock, A.A.; Decker, M.W. Effects of histamine H(3) receptor ligands GT-2331 and ciproxifan in a repeated acquisition avoidance response in the spontaneously hypertensive rat pup. Behav. Brain Res. 2002, 131, 151–161. [Google Scholar] [CrossRef]
- Fox, G.B.; Pan, J.B.; Esbenshade, T.A.; Bitner, R.S.; Nikkel, A.L.; Miller, T.; Kang, C.H.; Bennani, Y.L.; Black, L.A.; Faghih, R.; et al. Differential in vivo effects of H3 receptor ligands in a new mouse dipsogenia model. Pharmacol. Biochem. Behav. 2002, 72, 741–750. [Google Scholar] [CrossRef]
- Fox, G.B.; Pan, J.B.; Radek, R.J.; Lewis, A.M.; Bitner, R.S.; Esbenshade, T.A.; Faghih, R.; Bennani, Y.L.; Williams, M.; Yao, B.B.; et al. Two novel and selective nonimidazole H3 receptor antagonists A-304121 and A-317920: II. In vivo behavioral and neurophysiological characterization. J. Pharmacol. Exp. Ther. 2003, 305, 897–908. [Google Scholar] [CrossRef] [Green Version]
- Noland, J.S.; Steven Reznick, J.; Stone, W.L.; Walden, T.; Sheridan, E.H. Better working memory for non-social targets in infant siblings of children with Autism Spectrum Disorder. Dev. Sci. 2010, 13, 244–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedel, G.; Kang, S.H.; Choi, D.Y.; Platt, B. Scopolamine-induced deficits in social memory in mice: Reversal by donepezil. Behav. Brain Res. 2009, 204, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Jayaprakash, P.; Azimullah, S.; Ojha, S.K.; Al-Houqani, M.; Jalal, F.Y.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. The histamine H3R antagonist DL77 attenuates autistic behaviors in a prenatal valproic acid-induced mouse model of autism. Sci. Rep. 2018, 8, 13077. [Google Scholar] [CrossRef] [PubMed]
- Brioni, J.D.; Esbenshade, T.A.; Garrison, T.R.; Bitner, S.R.; Cowart, M.D. Discovery of histamine H3 antagonists for the treatment of cognitive disorders and Alzheimer’s disease. J. Pharmacol. Exp. Ther. 2011, 336, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Paschou, P.; Fernandez, T.V.; Sharp, F.; Heiman, G.A.; Hoekstra, P.J. Genetic susceptibility and neurotransmitters in tourette syndrome. Int. Rev. Neurobiol. 2013, 112, 155–177. [Google Scholar] [CrossRef] [Green Version]
- Gillberg, C.; Billstedt, E. Autism and Asperger syndrome: Coexistence with other clinical disorders. Acta Psychiatr. Scand. 2000, 102, 321–330. [Google Scholar] [CrossRef]
- Karagiannidis, I.; Dehning, S.; Sandor, P.; Tarnok, Z.; Rizzo, R.; Wolanczyk, T.; Madruga-Garrido, M.; Hebebrand, J.; Nothen, M.M.; Lehmkuhl, G.; et al. Support of the histaminergic hypothesis in Tourette syndrome: Association of the histamine decarboxylase gene in a large sample of families. J. Med. Genet. 2013, 50, 760–764. [Google Scholar] [CrossRef] [Green Version]
- Haas, H.L.; Sergeeva, O.A.; Selbach, O. Histamine in the nervous system. Physiol. Rev. 2008, 88, 1183–1241. [Google Scholar] [CrossRef]
- Hagenow, S.; Stasiak, A.; Ramsay, R.R.; Stark, H. Ciproxifan, a histamine H3 receptor antagonist, reversibly inhibits monoamine oxidase A and B. Sci. Rep. 2017, 7, 40541. [Google Scholar] [CrossRef] [Green Version]
- Chugani, D.C. Role of altered brain serotonin mechanisms in autism. Mol. Psychiatry 2002, 7 (Suppl. 2), S16–S17. [Google Scholar] [CrossRef] [Green Version]
- Mabunga, D.F.N.; Gonzales, E.L.T.; Kim, J.W.; Kim, K.C.; Shin, C.Y. Exploring the validity of valproic acid animal model of autism. Exp. Neurobiol. 2015, 24, 285–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orekhova, E.V.; Stroganova, T.A.; Prokofyev, A.O.; Nygren, G.; Gillberg, C.; Elam, M. Sensory gating in young children with autism: Relation to age, IQ, and EEG gamma oscillations. Neurosci. Lett. 2008, 434, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.F.; Buxhoeveden, D.; Gomez, J. Disruption in the inhibitory architecture of the cell minicolumn: Implications for autism. Neuroscientist 2003, 9, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, J.L.R.; Merzenich, M.M. Model of autism: Increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav. 2003, 2, 255–267. [Google Scholar] [CrossRef]
- Bahi, A.; Schwed, J.S.; Walter, M.; Stark, H.; Sadek, B. Anxiolytic and antidepressant-like activities of the novel and potent non-imidazole histamine H(3) receptor antagonist ST-1283. Drug Des. Dev. Ther. 2014, 8, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, D. Histamine H3 receptors and its antagonism as a novel mechanism for antipsychotic effect: A current preclinical & clinical perspective. Int. J. Health Sci. 2016, 10, 564–575. [Google Scholar]
- Mahmood, D.; Khanam, R.; Pillai, K.K.; Akhtar, M. Reversal of oxidative stress by histamine H(3) receptor-ligands in experimental models of schizophrenia. Arzneimittelforschung 2012, 62, 222–229. [Google Scholar] [CrossRef]
- Bastaki, S.M.; Abdulrazzaq, Y.M.; Shafiullah, M.; Wiecek, M.; Kiec-Kononowicz, K.; Sadek, B. Anticonvulsant and reproductive toxicological studies of the imidazole-based histamine H3R antagonist 2-18 in mice. Drug Des. Dev. Ther. 2018, 12, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, S.; Takuma, K.; Hara, Y.; Maeda, Y.; Ago, Y.; Matsuda, T. Autism-like behaviours with transient histone hyperacetylation in mice treated prenatally with valproic acid. Int. J. Neuropsychopharmacol. 2013, 16, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Takuma, K.; Hara, Y.; Kataoka, S.; Kawanai, T.; Maeda, Y.; Watanabe, R.; Takano, E.; Hayata-Takano, A.; Hashimoto, H.; Ago, Y.; et al. Chronic treatment with valproic acid or sodium butyrate attenuates novel object recognition deficits and hippocampal dendritic spine loss in a mouse model of autism. Pharmacol. Biochem. Behav. 2014, 126, 43–49. [Google Scholar] [CrossRef]
- Silverman, J.L.; Yang, M.; Lord, C.; Crawley, J.N. Behavioural phenotyping assays for mouse models of autism. Nat. Rev. Neurosci. 2010, 11, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Bambini-Junior, V.; Zanatta, G.; Della Flora Nunes, G.; Mueller de Melo, G.; Michels, M.; Fontes-Dutra, M.; Nogueira Freire, V.; Riesgo, R.; Gottfried, C. Resveratrol prevents social deficits in animal model of autism induced by valproic acid. Neurosci. Lett. 2014, 583, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.; Burant, A.; Bui, N.; Graham, D.; Yuva-Paylor, L.A.; Paylor, R. Marble burying reflects a repetitive and perseverative behavior more than novelty-induced anxiety. Psychopharmacology 2009, 204, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Theoharides, T.C.; Tsilioni, I.; Patel, A.B.; Doyle, R. Atopic diseases and inflammation of the brain in the pathogenesis of autism spectrum disorders. Transl. Psychiatry 2016, 6, e844. [Google Scholar] [CrossRef] [Green Version]
- Angoa-Perez, M.; Kane, M.J.; Briggs, D.I.; Francescutti, D.M.; Kuhn, D.M. Marble burying and nestlet shredding as tests of repetitive, compulsive-like behaviors in mice. J. Vis. Exp. 2013. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Seung, H.; Kwon, K.J.; Ko, M.J.; Lee, E.J.; Oh, H.A.; Choi, C.S.; Kim, K.C.; Gonzales, E.L.; You, J.S.; et al. Subchronic treatment of donepezil rescues impaired social, hyperactive, and stereotypic behavior in valproic acid-induced animal model of autism. PLoS ONE 2014, 9, e104927. [Google Scholar] [CrossRef] [Green Version]
- Bahi, A. Individual differences in elevated plus-maze exploration predicted higher ethanol consumption and preference in outbred mice. Pharmacol. Biochem. Behav. 2013, 105, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Bahi, A. Increased anxiety, voluntary alcohol consumption and ethanol-induced place preference in mice following chronic psychosocial stress. Stress 2013, 16, 441–451. [Google Scholar] [CrossRef]
- Bahi, A.; Dreyer, J.L. Hippocampus-specific deletion of tissue plasminogen activator “tPA” in adult mice impairs depression- and anxiety-like behaviors. Eur. Neuropsychopharmacol. 2012, 22, 672–682. [Google Scholar] [CrossRef]
- Bahi, A.; Dreyer, J.L. Chronic psychosocial stress causes delayed extinction and exacerbates reinstatement of ethanol-induced conditioned place preference in mice. Psychopharmacology 2014, 231, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Ojha, S.; Javed, H.; Azimullah, S.; Khair, S.B.A.; Haque, M.E. Neuroprotective potential of ferulic acid in the rotenone model of Parkinson’s disease. Drug Des. Dev. Ther. 2015, 9, 5499–5510. [Google Scholar] [CrossRef] [Green Version]
- Javed, H.; Azimullah, S.; Abul Khair, S.B.; Ojha, S.; Haque, M.E. Neuroprotective effect of nerolidol against neuroinflammation and oxidative stress induced by rotenone. BMC Neurosci. 2016, 17, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Łażewska, D.; Kieć-Kononowicz, K.; et al. The Dual-Active Histamine H3 Receptor Antagonist and Acetylcholine Esterase Inhibitor E100 Alleviates Autistic-Like Behaviors and Oxidative Stress in Valproic Acid Induced Autism in Mice. Int. J. Mol. Sci. 2020, 21, 3996. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113996
Eissa N, Azimullah S, Jayaprakash P, Jayaraj RL, Reiner D, Ojha SK, Beiram R, Stark H, Łażewska D, Kieć-Kononowicz K, et al. The Dual-Active Histamine H3 Receptor Antagonist and Acetylcholine Esterase Inhibitor E100 Alleviates Autistic-Like Behaviors and Oxidative Stress in Valproic Acid Induced Autism in Mice. International Journal of Molecular Sciences. 2020; 21(11):3996. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113996
Chicago/Turabian StyleEissa, Nermin, Sheikh Azimullah, Petrilla Jayaprakash, Richard L. Jayaraj, David Reiner, Shreesh K. Ojha, Rami Beiram, Holger Stark, Dorota Łażewska, Katarzyna Kieć-Kononowicz, and et al. 2020. "The Dual-Active Histamine H3 Receptor Antagonist and Acetylcholine Esterase Inhibitor E100 Alleviates Autistic-Like Behaviors and Oxidative Stress in Valproic Acid Induced Autism in Mice" International Journal of Molecular Sciences 21, no. 11: 3996. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21113996