Proteinaceous Transformers: Structural and Functional Variability of Human sHsps

{kind=link}
{kind=link}
{kind=link}
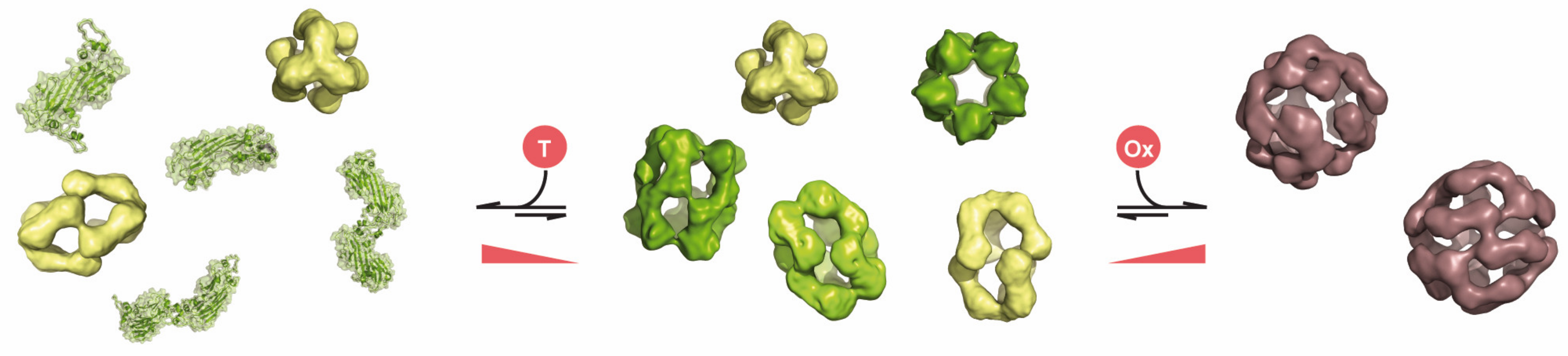{kind=link}
Abstract
:1. Introduction
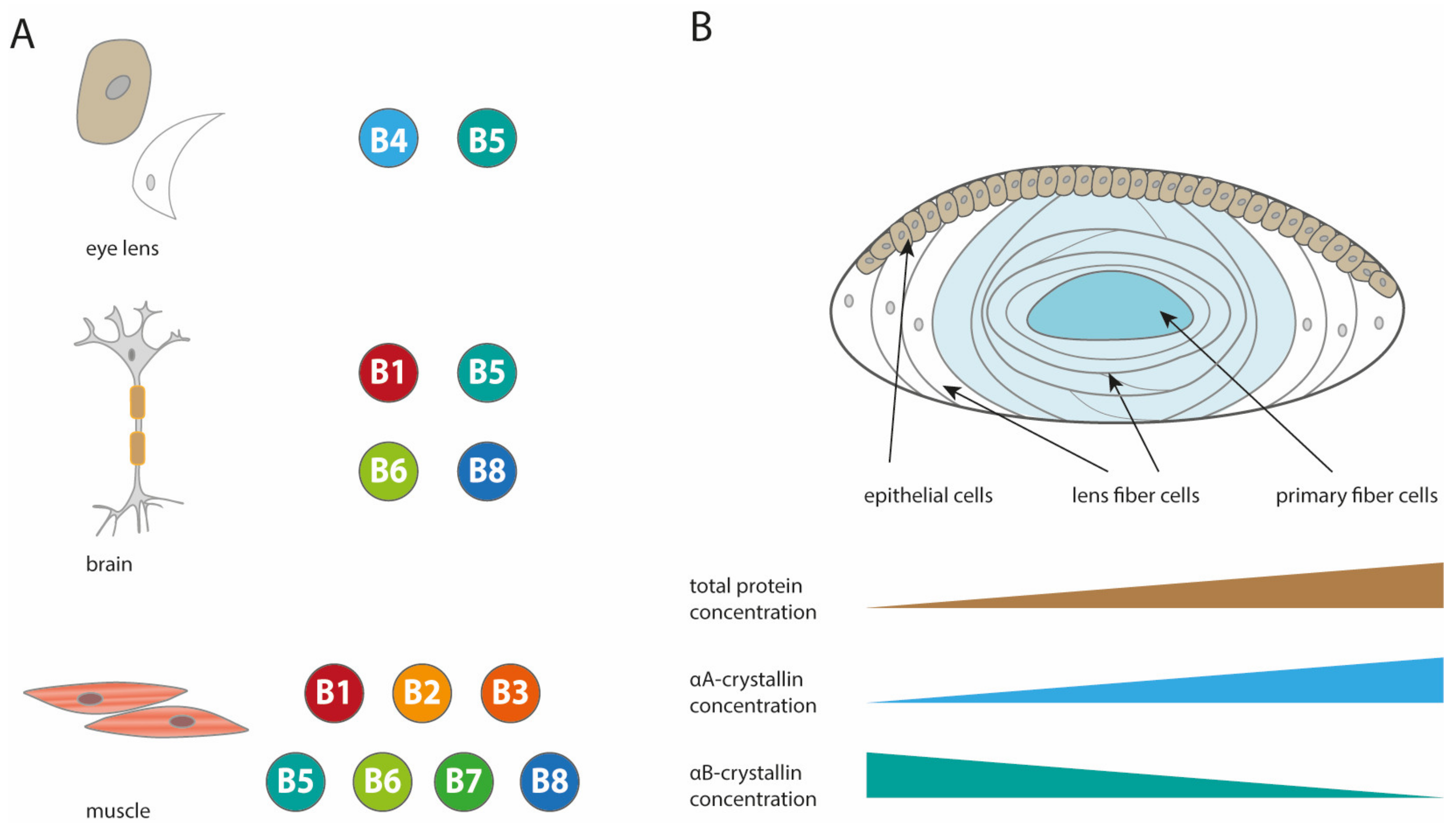
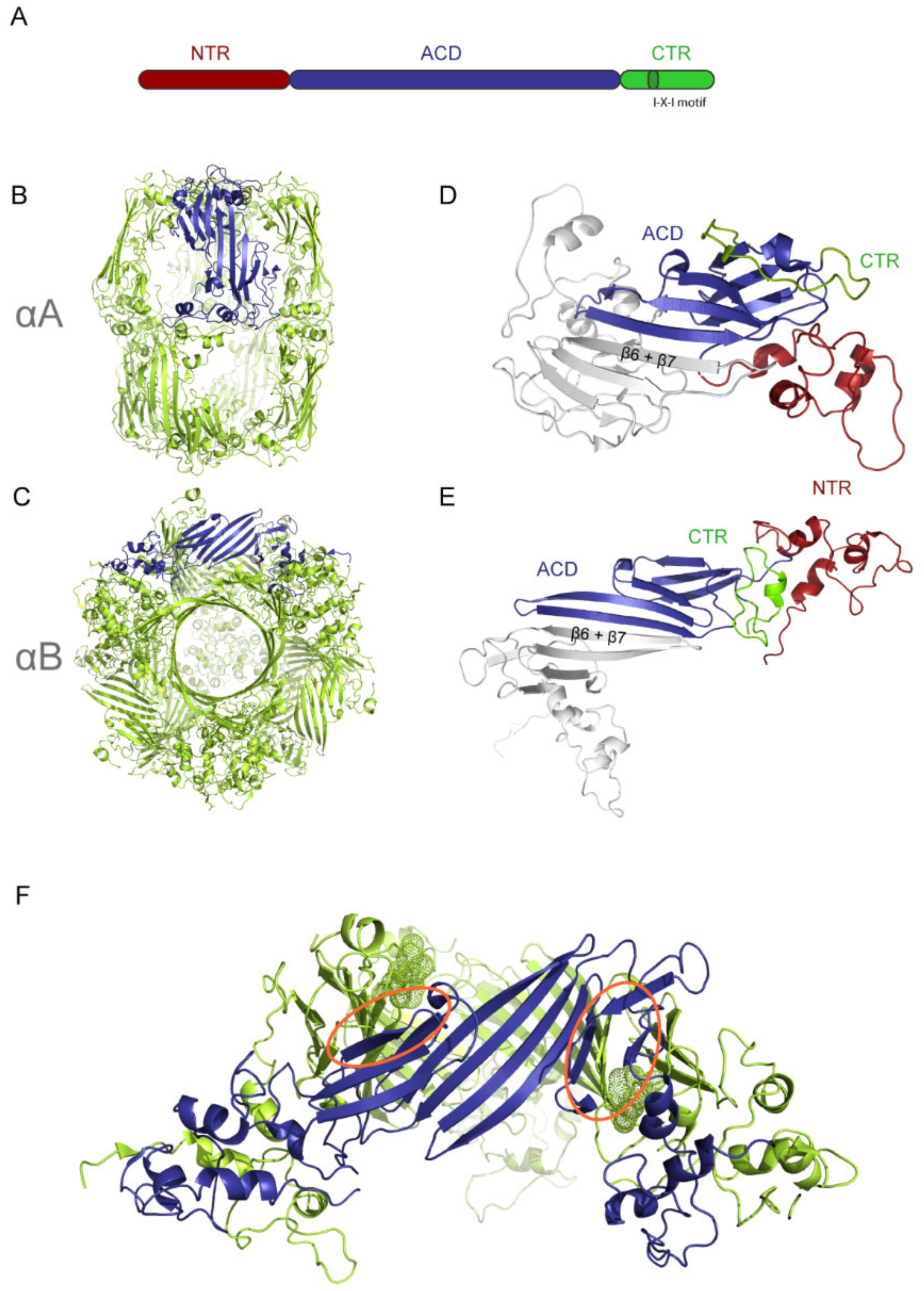
2. The Structure of Human sHsps
2.1. Structural Comparison of αA and αB
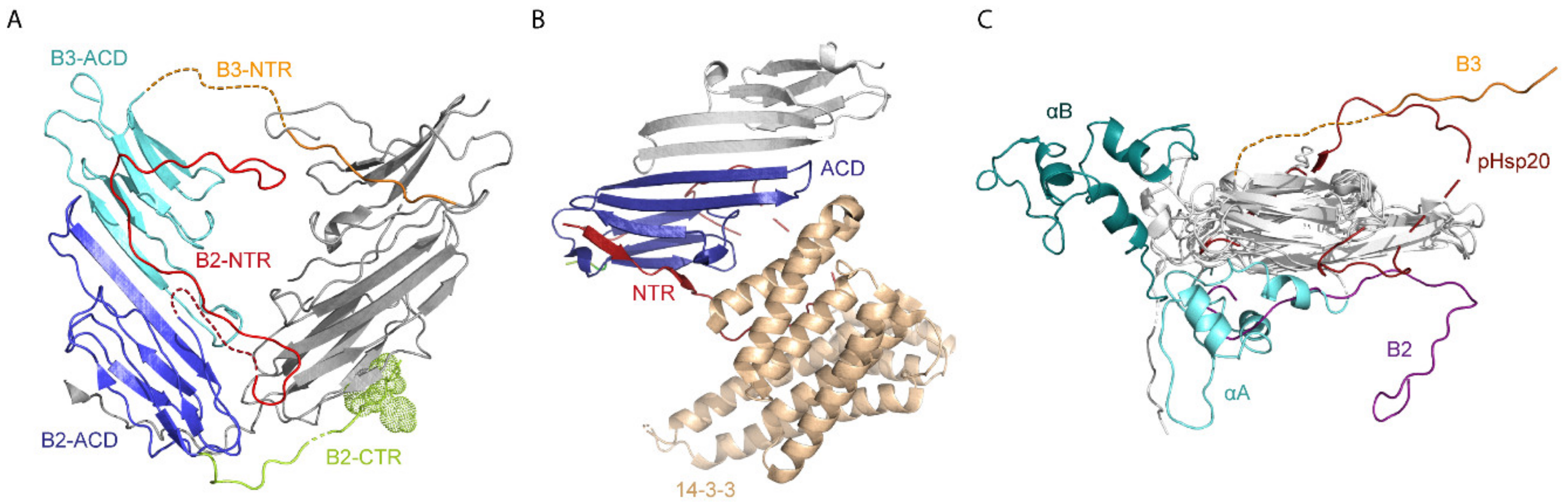
2.2. Structures of Other Human sHsps
3. Chaperone Function of Human sHsps
4. Conclusions
- sHsps are an integral component of the chaperone network and help to sustain the solubility and functionality of the proteome upon stress
- Under physiological conditions they are involved in a number of regulatory processes within the cell
- They are implicated in a variety of diseases but are usually not the causative component
- Their function is closely correlated to their structural variability
- They are commonly organized in three sequence parts, a conserved ACD flanked by divergent NTRs and CTRs
- Most sHsps form dynamic ensembles of oligomers with a variable number of subunits; some sHsps even form hetero-oligomeric species
- The assembly processes are controlled by the three sequence parts which form contacts in a hierarchical manner
- The conformational flexibility of the NTR, together with a set of binding groves on the ACD, are most likely the key to the variable oligomeric assemblies
- Changes in the composition of the dynamic ensembles of oligomers are linked to their chaperone activity and the recognition of substrates
- Changes in dynamics and composition of the ensembles lead also to modulated substrate specificities
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vabulas, R.M.; Raychaudhuri, S.; Hayer-Hartl, M.; Hartl, F.U. Protein folding in the cytoplasm and the heat shock response. Cold Spring Harb. Perspect. Biol. 2010, 2, a004390. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Vierling, E. A first line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogk, A.; Ruger-Herreros, C.; Bukau, B. Cellular Functions and Mechanisms of Action of Small Heat Shock Proteins. Annu. Rev. Microbiol. 2019, 73, 89–110. [Google Scholar] [CrossRef]
- Waters, E.R.; Vierling, E. Plant small heat shock proteins—Evolutionary and functional diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, J.H.; Linke, K.; Graf, P.C.; Lilie, H.; Jakob, U. Identification of a redox-regulated chaperone network. EMBO J. 2004, 23, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Mattoo, R.U.; Goloubinoff, P. Molecular chaperones are nanomachines that catalytically unfold misfolded and alternatively folded proteins. Cell. Mol. Life Sci. 2014, 71, 3311–3325. [Google Scholar] [CrossRef] [Green Version]
- Zwirowski, S.; Klosowska, A.; Obuchowski, I.; Nillegoda, N.B.; Pirog, A.; Zietkiewicz, S.; Bukau, B.; Mogk, A.; Liberek, K. Hsp70 displaces small heat shock proteins from aggregates to initiate protein refolding. EMBO J. 2017, 36, 783–796. [Google Scholar] [CrossRef]
- Allen, S.P.; Polazzi, J.O.; Gierse, J.K.; Easton, A.M. Two novel heat shock genes encoding proteins produced in response to heterologous protein expression in Escherichia coli. J. Bacteriol. 1992, 174, 6938–6947. [Google Scholar] [CrossRef] [Green Version]
- Bakthisaran, R.; Tangirala, R.; Rao, C.M. Small heat shock proteins: Role in cellular functions and pathology. Biochim. Biophys. Acta 2014, 1854, 291–319. [Google Scholar] [CrossRef] [Green Version]
- Mogk, A.; Bukau, B.; Kampinga, H.H. Cellular Handling of Protein Aggregates by Disaggregation Machines. Mol. Cell 2018, 69, 214–226. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Miess, A.; Stromer, T.; Walter, S.; Buchner, J. Disassembling protein aggregates in the yeast cytosol. The cooperation of Hsp26 with Ssa1 and Hsp104. J. Biol. Chem. 2005, 280, 23861–23868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogk, A.; Deuerling, E.; Vorderwulbecke, S.; Vierling, E.; Bukau, B. Small heat shock proteins, ClpB and the DnaK system form a functional triade in reversing protein aggregation. Mol. Microbiol. 2003, 50, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, E.; Zietkiewicz, S.; Liberek, K. Distinct activities of Escherichia coli small heat shock proteins IbpA and IbpB promote efficient protein disaggregation. J. Mol. Biol. 2009, 386, 178–189. [Google Scholar] [CrossRef] [PubMed]
- De Jong, W.W.; Caspers, G.J.; Leunissen, J.A. Genealogy of the alpha-crystallin--small heat-shock protein superfamily. Int. J. Biol. Macromol. 1998, 22, 151–162. [Google Scholar] [CrossRef]
- Treweek, T.M.; Meehan, S.; Ecroyd, H.; Carver, J.A. Small heat-shock proteins: Important players in regulating cellular proteostasis. Cell. Mol. Life Sci. 2015, 72, 429–451. [Google Scholar] [CrossRef]
- Haslbeck, M.; Weinkauf, S.; Buchner, J. Small heat shock proteins: Simplicity meets complexity. J. Biol. Chem. 2019, 294, 2121–2132. [Google Scholar] [CrossRef] [Green Version]
- De Jong, W.W.; Leunissen, J.A.; Voorter, C.E. Evolution of the alpha-crystallin/small heat-shock protein family. Mol. Biol. Evol. 1993, 10, 103–126. [Google Scholar]
- Kriehuber, T.; Rattei, T.; Weinmaier, T.; Bepperling, A.; Haslbeck, M.; Buchner, J. Independent evolution of the core domain and its flanking sequences in small heat shock proteins. FASEB J. 2010, 24, 3633–3642. [Google Scholar] [CrossRef]
- Kappe, G.; Franck, E.; Verschuure, P.; Boelens, W.C.; Leunissen, J.A.M.; De Jong, W.W. The human genome encodes 10 alpha-crystallin-related small heat shock proteins: HspB1-10. Cell Stress Chaperones 2003, 8, 53–61. [Google Scholar] [CrossRef]
- Kappe, G.; Verschuure, P.; Philipsen, R.L.A.; Staalduinen, A.A.; Van de Boogaart, P.; Boelens, W.C.; De Jong, W.W. Characterization of two novel human small heat shock proteins: Protein kinase-related HspB8 and testis-specific HspB9. Biochim. Biophys. Acta 2001, 1520, 1–6. [Google Scholar] [CrossRef]
- Horwitz, J. Alpha-crystallin. Exp. Eye Res. 2003, 76, 145–153. [Google Scholar] [CrossRef]
- Slingsby, C.; Wistow, G.J.; Clark, A.R. Evolution of crystallins for a role in the vertebrate eye lens. Protein Sci. 2013, 22, 367–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mymrikov, E.V.; Seit-Nebi, A.S.; Gusev, N.B. Large potentials of small heat shock proteins. Physiol. Rev. 2011, 91, 1123–1159. [Google Scholar] [CrossRef] [Green Version]
- Kampinga, H.H.; Garrido, C. HSPBs: Small proteins with big implications in human disease. Int. J. Biochem. Cell Biol. 2012, 44, 1706–1710. [Google Scholar] [CrossRef]
- Mymrikov, E.V.; Riedl, M.; Peters, C.; Weinkauf, S.; Haslbeck, M.; Buchner, J. Regulation of small heat-shock proteins by hetero-oligomer formation. J. Biol. Chem. 2020, 295, 158–169. [Google Scholar] [CrossRef]
- Bukach, O.V.; Glukhova, A.E.; Seit-Nebi, A.S.; Gusev, N.B. Heterooligomeric complexes formed by human small heat shock proteins HspB1 (Hsp27) and HspB6 (Hsp20). Biochim. Biophys. Acta 2009, 1794, 486–495. [Google Scholar] [CrossRef]
- Mymrikov, E.V.; Seit-Nebi, A.S.; Gusev, N.B. Heterooligomeric complexes of human small heat shock proteins. Cell Stress Chaperones 2012, 17, 157–169. [Google Scholar] [CrossRef] [Green Version]
- Kato, K.; Hasegawa, K.; Goto, S.; Inaguma, Y. Dissociation as a result of phosphorylation of an aggregated form of the small stress protein, hsp27. J. Biol. Chem. 1994, 269, 11274–11278. [Google Scholar]
- Siezen, R.J.; Berger, H. The quaternary structure of bovine alpha-crystallin. Size and shape studies by sedimentation, small-angle X-ray scattering and quasi-elastic light scattering. Eur. J. Biochem. 1978, 91, 397–405. [Google Scholar] [CrossRef]
- Hanson, S.R.; Smith, D.L.; Smith, J.B. Deamidation and disulfide bonding in human lens gamma-crystallins. Exp. Eye Res. 1998, 67, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Peschek, J.; Buchner, J.; Weinkauf, S. Structure and function of alpha-crystallins: Traversing from in vitro to in vivo. Biochim. Biophys. Acta 2016, 1860, 149–166. [Google Scholar] [CrossRef] [PubMed]
- Graw, J. Genetics of crystallins: Cataract and beyond. Exp. Eye Res. 2009, 88, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Duncan, M.K.; Xie, L.; David, L.L.; Robinson, M.L.; Taube, J.R.; Cui, W.; Reneker, L.W. Ectopic Pax6 expression disturbs lens fiber cell differentiation. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3589–3598. [Google Scholar] [CrossRef] [Green Version]
- Bloemendal, H.; De Jong, W.; Jaenicke, R.; Lubsen, N.H.; Slingsby, C.; Tardieu, A. Ageing and vision: Structure, stability and function of lens crystallins. Prog. Biophys. Mol. Biol. 2004, 86, 407–485. [Google Scholar] [CrossRef]
- Sharma, K.K.; Santhoshkumar, P. Lens aging: Effects of crystallins. Biochim. Biophys. Acta 2009, 1790, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Clark, A.R.; Lubsen, N.H.; Slingsby, C. sHSP in the eye lens: Crystallin mutations, cataract and proteostasis. Int. J. Biochem. Cell Biol. 2012, 44, 1687–1697. [Google Scholar] [CrossRef]
- Jaenicke, R.; Slingsby, C. Lens crystallins and their microbial homologs: Structure, stability, and function. Crit. Rev. Biochem. Mol. Biol. 2001, 36, 435–499. [Google Scholar] [CrossRef]
- Delaye, M.; Tardieu, A. Short-range order of crystallin proteins accounts for eye lens transparency. Nature 1983, 302, 415–417. [Google Scholar] [CrossRef]
- Tardieu, A. alpha-Crystallin quaternary structure and interactive properties control eye lens transparency. Int. J. Biol. Macromol. 1998, 22, 211–217. [Google Scholar] [CrossRef]
- Tardieu, A. Eye lens proteins and transparency: From light transmission theory to solution X-ray structural analysis. Annu. Rev. Biophys. Biophys. Chem. 1988, 17, 47–70. [Google Scholar] [CrossRef] [PubMed]
- Bassnett, S.; Shi, Y.; Vrensen, G.F. Biological glass: Structural determinants of eye lens transparency. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2011, 366, 1250–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nandi, S.K.; Nahomi, R.B.; Rankenberg, J.; Glomb, M.A.; Nagaraj, R.H. Glycation-mediated inter-protein cross-linking is promoted by chaperone-client complexes of alpha-crystallin: Implications for lens aging and presbyopia. J. Biol. Chem. 2020, 295, 5701–5716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicart, P.; Caron, A.; Guicheney, P.; Li, Z.; Prevost, M.C.; Faure, A.; Chateau, D.; Chapon, F.; Tome, F.; Dupret, J.M.; et al. A missense mutation in the alphaB-crystallin chaperone gene causes a desmin-related myopathy. Nat. Genet. 1998, 20, 92–95. [Google Scholar] [CrossRef]
- Litt, M.; Kramer, P.; LaMorticella, D.M.; Murphey, W.; Lovrien, E.W.; Weleber, R.G. Autosomal dominant congenital cataract associated with a missense mutation in the human alpha crystallin gene CRYAA. Hum. Mol. Genet. 1998, 7, 471–474. [Google Scholar] [CrossRef] [Green Version]
- Klemenz, R.; Frohli, E.; Steiger, R.H.; Schafer, R.; Aoyama, A. Alpha B-crystallin is a small heat shock protein. Proc. Natl. Acad. Sci. USA 1991, 88, 3652–3656. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, J. Alpha-crystallin can function as a molecular chaperone. Proc. Natl. Acad. Sci. USA 1992, 89, 10449–10453. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; MacRae, T.H. The small heat shock proteins and their role in human disease. FEBS J. 2005, 272, 2613–2627. [Google Scholar] [CrossRef]
- Webster, J.M.; Darling, A.L.; Uversky, V.N.; Blair, L.J. Small Heat Shock Proteins, Big Impact on Protein Aggregation in Neurodegenerative Disease. Front. Pharmacol. 2019, 10, 1047. [Google Scholar] [CrossRef]
- Muranova, L.K.; Ryzhavskaya, A.S.; Sudnitsyna, M.V.; Shatov, V.M.; Gusev, N.B. Small Heat Shock Proteins and Human Neurodegenerative Diseases. Biochemistry 2019, 84, 1256–1267. [Google Scholar]
- Bova, M.P.; Yaron, O.; Huang, Q.; Ding, L.; Haley, D.A.; Stewart, P.L.; Horwitz, J. Mutation R120G in alphaB-crystallin, which is linked to a desmin-related myopathy, results in an irregular structure and defective chaperone-like function. Proc. Natl. Acad. Sci. USA 1999, 96, 6137–6142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makley, L.N.; McMenimen, K.A.; DeVree, B.T.; Goldman, J.W.; McGlasson, B.N.; Rajagopal, P.; Dunyak, B.M.; McQuade, T.J.; Thompson, A.D.; Sunahara, R.; et al. Pharmacological chaperone for alpha-crystallin partially restores transparency in cataract models. Science 2015, 350, 674–677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Chen, X.J.; Zhu, J.; Xi, Y.B.; Yang, X.; Hu, L.D.; Ouyang, H.; Patel, S.H.; Jin, X.; Lin, D.; et al. Lanosterol reverses protein aggregation in cataracts. Nature 2015, 523, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Mymrikov, E.V.; Haslbeck, M. Medical implications of understanding the functions of human small heat shock proteins. Expert Rev. Proteom. 2015, 12, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Paul, C.; Seigneuric, R.; Kampinga, H.H. The small heat shock proteins family: the long forgotten chaperones. Int. J. Biochem. Cell. Biol. 2012, 44(10), 1588–1592. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Barton, K.; De Maria, A.; Petrash, J.M.; Shiels, A.; Bassnett, S. The stratified syncytium of the vertebrate lens. J. Cell Sci. 2009, 122, 1607–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delcour, J.; Papaconstantinou, J. A change in the stoichiometry of assembly of bovine lens alpha-crystallin subunits in relation to cellular differentiation. Biochem. Biophys. Res. Commun. 1974, 57, 134–141. [Google Scholar] [CrossRef]
- Delbecq, S.P.; Klevit, R.E. One size does not fit all: The oligomeric states of alphaB crystallin. FEBS Lett. 2013, 587, 1073–1080. [Google Scholar] [CrossRef] [Green Version]
- Bagneris, C.; Bateman, O.A.; Naylor, C.E.; Cronin, N.; Boelens, W.C.; Keep, N.H.; Slingsby, C. Crystal structures of alpha-crystallin domain dimers of alphaB-crystallin and Hsp20. J. Mol. Biol. 2009, 392, 1242–1252. [Google Scholar] [CrossRef]
- Laganowsky, A.; Benesch, J.L.; Landau, M.; Ding, L.; Sawaya, M.R.; Cascio, D.; Huang, Q.; Robinson, C.V.; Horwitz, J.; Eisenberg, D. Crystal structures of truncated alphaA and alphaB crystallins reveal structural mechanisms of polydispersity important for eye lens function. Protein Sci. 2010, 19, 1031–1043. [Google Scholar] [CrossRef] [Green Version]
- Baranova, E.V.; Weeks, S.D.; Beelen, S.; Bukach, O.V.; Gusev, N.B.; Strelkov, S.V. Three-Dimensional Structure of alpha-Crystallin Domain Dimers of Human Small Heat Shock Proteins HSPB1 and HSPB6. J. Mol. Biol. 2011, 411, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, T.; Kastenmuller, A.; Stein, M.L.; Peters, C.; Daake, M.; Krause, M.; Weinfurtner, D.; Haslbeck, M.; Weinkauf, S.; Groll, M.; et al. The Chaperone Activity of the Developmental Small Heat Shock Protein Sip1 Is Regulated by pH-Dependent Conformational Changes. Mol. Cell 2015, 58, 1067–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sluchanko, N.N.; Beelen, S.; Kulikova, A.A.; Weeks, S.D.; Antson, A.A.; Gusev, N.B.; Strelkov, S.V. Structural Basis for the Interaction of a Human Small Heat Shock Protein with the 14-3-3 Universal Signaling Regulator. Structure 2017, 25, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.R.; Egberts, W.V.; Kondrat, F.D.L.; Hilton, G.R.; Ray, N.J.; Cole, A.R.; Carver, J.A.; Benesch, J.L.P.; Keep, N.H.; Boelens, W.C.; et al. Terminal Regions Confer Plasticity to the Tetrameric Assembly of Human HspB2 and HspB3. J. Mol. Biol. 2018, 430, 3297–3310. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, P.; Liu, Y.; Shi, L.; Clouser, A.F.; Klevit, R.E. Structure of the alpha-crystallin domain from the redox-sensitive chaperone, HSPB1. J. Biomol. NMR 2015, 63, 223–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alderson, T.R.; Ying, J.; Bax, A.; Benesch, J.L.P.; Baldwin, A.J. Conditional Disorder in Small Heat-shock Proteins. J. Mol. Biol. 2020, 432, 3033–3049. [Google Scholar] [CrossRef]
- Hilton, G.R.; Hochberg, G.K.; Laganowsky, A.; McGinnigle, S.I.; Baldwin, A.J.; Benesch, J.L. C-terminal interactions mediate the quaternary dynamics of alphaB-crystallin. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20110405. [Google Scholar] [CrossRef] [Green Version]
- Aquilina, J.A.; Benesch, J.L.; Ding, L.L.; Yaron, O.; Horwitz, J.; Robinson, C.V. Phosphorylation of alphaB-crystallin alters chaperone function through loss of dimeric substructure. J. Biol. Chem. 2004, 279, 28675–28680. [Google Scholar] [CrossRef] [Green Version]
- Aquilina, J.A.; Benesch, J.L.; Bateman, O.A.; Slingsby, C.; Robinson, C.V. Polydispersity of a mammalian chaperone: Mass spectrometry reveals the population of oligomers in alphaB-crystallin. Proc. Natl. Acad. Sci. USA 2003, 100, 10611–10616. [Google Scholar] [CrossRef] [Green Version]
- Aquilina, J.A.; Shrestha, S.; Morris, A.M.; Ecroyd, H. Structural and functional aspects of hetero-oligomers formed by the small heat shock proteins alphaB-crystallin and HSP27. J. Biol. Chem. 2013, 288, 13602–13609. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, A.J.; Lioe, H.; Robinson, C.V.; Kay, L.E.; Benesch, J.L. alphaB-Crystallin Polydispersity Is a Consequence of Unbiased Quaternary Dynamics. J. Mol. Biol. 2011, 413, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, G.K.; Benesch, J.L. Dynamical structure of alphaB-crystallin. Prog. Biophys. Mol. Biol. 2014, 115, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Kim, R.; Kim, S.H. Crystal structure of a small heat-shock protein. Nature 1998, 394, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Van Montfort, R.L.; Basha, E.; Friedrich, K.L.; Slingsby, C.; Vierling, E. Crystal structure and assembly of a eukaryotic small heat shock protein. Nat. Struct. Biol. 2001, 8, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Hanazono, Y.; Takeda, K.; Oka, T.; Abe, T.; Tomonari, T.; Akiyama, N.; Aikawa, Y.; Yohda, M.; Miki, K. Nonequivalence observed for the 16-meric structure of a small heat shock protein, SpHsp16.0, from Schizosaccharomyces pombe. Structure 2013, 21, 220–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilario, E.; Martin, F.J.; Bertolini, M.C.; Fan, L. Crystal structures of Xanthomonas small heat shock protein provide a structural basis for an active molecular chaperone oligomer. J. Mol. Biol. 2011, 408, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Chen, J.Y.; Yang, B.; Wang, F.H.; Wang, Y.H.; Yun, C.H. Active-State Structures of a Small Heat-Shock Protein Revealed a Molecular Switch for Chaperone Function. Structure 2015, 23, 2066–2075. [Google Scholar] [CrossRef] [Green Version]
- Mani, N.; Bhandari, S.; Moreno, R.; Hu, L.; Prasad, B.V.V.; Suguna, K. Multiple oligomeric structures of a bacterial small heat shock protein. Sci. Rep. 2016, 6, 24019. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, C.J.O.; Peters, C.; Schmid, P.W.N.; Stavropoulou, M.; Zou, J.; Dahiya, V.; Mymrikov, E.V.; Rockel, B.; Asami, S.; Haslbeck, M.; et al. The structure and oxidation of the eye lens chaperone alphaA-crystallin. Nat. Struct. Mol. Biol. 2019, 26, 1141–1150. [Google Scholar] [CrossRef]
- Braun, N.; Zacharias, M.; Peschek, J.; Kastenmuller, A.; Zou, J.; Hanzlik, M.; Haslbeck, M.; Rappsilber, J.; Buchner, J.; Weinkauf, S. Multiple molecular architectures of the eye lens chaperone alphaB-crystallin elucidated by a triple hybrid approach. Proc. Natl. Acad. Sci. USA 2011, 108, 20491–20496. [Google Scholar] [CrossRef] [Green Version]
- Pasta, S.Y.; Raman, B.; Ramakrishna, T.; Rao, C. The IXI/V motif in the C-terminal extension of alpha-crystallins: Alternative interactions and oligomeric assemblies. Mol. Vis. 2004, 10, 655–662. [Google Scholar] [PubMed]
- Clouser, A.F.; Baughman, H.E.; Basanta, B.; Guttman, M.; Nath, A.; Klevit, R.E. Interplay of disordered and ordered regions of a human small heat shock protein yields an ensemble of ‘quasi-ordered’ states. eLife 2019, 8, e50259. [Google Scholar] [CrossRef] [PubMed]
- Delbecq, S.P.; Jehle, S.; Klevit, R. Binding determinants of the small heat shock protein, alphaB-crystallin: Recognition of the ‘IxI’ motif. EMBO J. 2012, 31, 4587–4594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treweek, T.M.; Rekas, A.; Walker, M.J.; Carver, J.A. A quantitative NMR spectroscopic examination of the flexibility of the C-terminal extensions of the molecular chaperones, alphaA- and alphaB-crystallin. Exp. Eye Res. 2010, 91, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, A.J.; Lioe, H.; Hilton, G.R.; Baker, L.A.; Rubinstein, J.L.; Kay, L.E.; Benesch, J.L. The polydispersity of alphaB-crystallin is rationalized by an interconverting polyhedral architecture. Structure 2011, 19, 1855–1863. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, J.; Huang, Q.L.; Ding, L.; Bova, M.P. Lens alpha-crystallin: Chaperone-like properties. Methods Enzymol. 1998, 290, 365–383. [Google Scholar]
- Kelley, P.B.; Abraham, E.C. Thermally induced disintegration of the oligomeric structure of alphaB-crystallin mutant F28S is associated with diminished chaperone activity. Mol. Cell. Biochem. 2003, 252, 273–278. [Google Scholar] [CrossRef]
- Shah, D.; Li, J.; Shaikh, A.R.; Rajagopalan, R. Arginine-aromatic interactions and their effects on arginine-induced solubilization of aromatic solutes and suppression of protein aggregation. Biotechnol. Prog. 2012, 28, 223–231. [Google Scholar] [CrossRef]
- Arakawa, T.; Ejima, D.; Tsumoto, K.; Obeyama, N.; Tanaka, Y.; Kita, Y.; Timasheff, S.N. Suppression of protein interactions by arginine: A proposed mechanism of the arginine effects. Biophys. Chem. 2007, 127, 1–8. [Google Scholar] [CrossRef]
- Shatov, V.M.; Weeks, S.D.; Strelkov, S.V.; Gusev, N.B. The Role of the Arginine in the Conserved N-Terminal Domain RLFDQxFG Motif of Human Small Heat Shock Proteins HspB1, HspB4, HspB5, HspB6, and HspB8. Int. J. Mol. Sci. 2018, 19, 2112. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Basha, E.; Wysocki, V.H.; Vierling, E. Insights into small heat shock protein and substrate structure during chaperone action derived from hydrogen/deuterium exchange and mass spectrometry. J. Biol. Chem. 2008, 283, 26634–26642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentze, N.; Narberhaus, F. Detection of oligomerisation and substrate recognition sites of small heat shock proteins by peptide arrays. Biochem. Biophys. Res. Commun. 2004, 325, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Basha, E.; Friedrich, K.L.; Vierling, E. The N-terminal arm of small heat shock proteins is important for both chaperone activity and substrate specificity. J. Biol. Chem. 2006, 281, 39943–39952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, J.G.; Shenoy, A.K., Jr.; Clark, J.I. Interactions between important regulatory proteins and human alphaB crystallin. Biochemistry 2007, 46, 6308–6317. [Google Scholar] [CrossRef]
- Ahrman, E.; Lambert, W.; Aquilina, J.A.; Robinson, C.V.; Emanuelsson, C.S. Chemical cross-linking of the chloroplast localized small heat-shock protein, Hsp21, and the model substrate citrate synthase. Protein Sci. 2007, 16, 1464–1478. [Google Scholar] [CrossRef] [Green Version]
- Jovcevski, B.; Aquilina, J.A.; Benesch, J.L.P.; Ecroyd, H. The influence of the N-terminal region proximal to the core domain on the assembly and chaperone activity of alphaB-crystallin. Cell Stress Chaperones 2018, 23, 827–836. [Google Scholar] [CrossRef]
- Cox, D.; Selig, E.; Griffin, M.D.; Carver, J.A.; Ecroyd, H. Small Heat-shock Proteins Prevent alpha-Synuclein Aggregation via Transient Interactions and Their Efficacy Is Affected by the Rate of Aggregation. J. Biol. Chem. 2016, 291, 22618–22629. [Google Scholar] [CrossRef] [Green Version]
- Mymrikov, E.V.; Daake, M.; Richter, B.; Haslbeck, M.; Buchner, J. The Chaperone Activity and Substrate Spectrum of Human Small Heat Shock Proteins. J. Biol. Chem. 2017, 292, 672–684. [Google Scholar] [CrossRef] [Green Version]
- Jiao, W.; Qian, M.; Li, P.; Zhao, L.; Chang, Z. The essential role of the flexible termini in the temperature-responsiveness of the oligomeric state and chaperone-like activity for the polydisperse small heat shock protein IbpB from Escherichia coli. J. Mol. Biol. 2005, 347, 871–884. [Google Scholar] [CrossRef]
- Peschek, J.; Braun, N.; Rohrberg, J.; Back, K.C.; Kriehuber, T.; Kastenmuller, A.; Weinkauf, S.; Buchner, J. Regulated structural transitions unleash the chaperone activity of alphaB-crystallin. Proc. Natl. Acad. Sci. USA 2013, 110, E3780–E3789. [Google Scholar] [CrossRef] [Green Version]
- Aquilina, J.A.; Watt, S.J. The N-terminal domain of alphaB-crystallin is protected from proteolysis by bound substrate. Biochem. Biophys. Res. Commun. 2007, 353, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Wintrode, P.L.; Friedrich, K.L.; Vierling, E.; Smith, J.B.; Smith, D.L. Solution structure and dynamics of a heat shock protein assembly probed by hydrogen exchange and mass spectrometry. Biochemistry 2003, 42, 10667–10673. [Google Scholar] [CrossRef] [PubMed]
- Preis, W.; Bestehorn, A.; Buchner, J.; Haslbeck, M. An alternative splice variant of human alphaA-crystallin modulates the oligomer ensemble and the chaperone activity of alpha-crystallins. Cell Stress Chaperones 2017, 22, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jehle, S.; Vollmar, B.S.; Bardiaux, B.; Dove, K.K.; Rajagopal, P.; Gonen, T.; Oschkinat, H.; Klevit, R.E. N-terminal domain of {alpha}B-crystallin provides a conformational switch for multimerization and structural heterogeneity. Proc. Natl. Acad. Sci. USA 2011, 108, 6409–6414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamler, R.; Kappe, G.; Boelens, W.; Slingsby, C. Wrapping the alpha-crystallin domain fold in a chaperone assembly. J. Mol. Biol. 2005, 353, 68–79. [Google Scholar] [CrossRef] [Green Version]
- Shatov, V.M.; Strelkov, S.V.; Gusev, N.B. The Heterooligomerization of Human Small Heat Shock Proteins Is Controlled by Conserved Motif Located in the N-Terminal Domain. Int. J. Mol. Sci. 2020, 21, 4248. [Google Scholar] [CrossRef]
- Collier, M.P.; Alderson, T.R.; De Villiers, C.P.; Nicholls, D.; Gastall, H.Y.; Allison, T.M.; Degiacomi, M.T.; Jiang, H.; Mlynek, G.; Furst, D.O.; et al. HspB1 phosphorylation regulates its intramolecular dynamics and mechanosensitive molecular chaperone interaction with filamin C. Sci. Adv. 2019, 5, eaav8421. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Kamei, K.; Iwamoto, I.; Inaguma, Y.; Nohara, D.; Kato, K. Phosphorylation-induced change of the oligomerization state of alpha B-crystallin. J. Biol. Chem. 2001, 276, 5346–5352. [Google Scholar] [CrossRef] [Green Version]
- Van den IJssel, P.R.; Overkamp, P.; Bloemendal, H.; De Jong, W.W. Phosphorylation of alphaB-crystallin and HSP27 is induced by similar stressors in HeLa cells. Biochem. Biophys. Res. Commun. 1998, 247, 518–523. [Google Scholar] [CrossRef]
- Gaestel, M. sHsp-phosphorylation: Enzymes, signaling pathways and functional implications. Prog. Mol. Subcell. Biol. 2002, 28, 151–169. [Google Scholar]
- Arrigo, A.P. Human small heat shock proteins: Protein interactomes of homo- and hetero-oligomeric complexes: An update. FEBS Lett. 2013, 587, 1959–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preville, X.; Schultz, H.; Knauf, U.; Gaestel, M.; Arrigo, A.P. Analysis of the role of Hsp25 phosphorylation reveals the importance of the oligomerization state of this small heat shock protein in its protective function against TNFalpha- and hydrogen peroxide-induced cell death. J. Cell. Biochem. 1998, 69, 436–452. [Google Scholar] [CrossRef]
- Thornell, E.; Aquilina, A. Regulation of alphaA- and alphaB-crystallins via phosphorylation in cellular homeostasis. Cell Mol. Life Sci. 2015, 72, 4127–4137. [Google Scholar] [CrossRef]
- Jovcevski, B.; Kelly, M.A.; Aquilina, J.A.; Benesch, J.L.P.; Ecroyd, H. Evaluating the Effect of Phosphorylation on the Structure and Dynamics of Hsp27 Dimers by Means of Ion Mobility Mass Spectrometry. Anal. Chem. 2017, 89, 13275–13282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peschek, J.; Braun, N.; Franzmann, T.M.; Georgalis, Y.; Haslbeck, M.; Weinkauf, S.; Buchner, J. The eye lens chaperone alpha-crystallin forms defined globular assemblies. Proc. Natl. Acad. Sci. USA 2009, 106, 13272–13277. [Google Scholar] [CrossRef] [Green Version]
- Jehle, S.; Van, R.B.; Stout, J.R.; Noguchi, S.M.; Falber, K.; Rehbein, K.; Oschkinat, H.; Klevit, R.E.; Rajagopal, P. alphaB-crystallin: A hybrid solid-state/solution-state NMR investigation reveals structural aspects of the heterogeneous oligomer. J. Mol. Biol. 2009, 385, 1481–1497. [Google Scholar] [CrossRef] [Green Version]
- Jehle, S.; Rajagopal, P.; Bardiaux, B.; Markovic, S.; Kuhne, R.; Stout, J.R.; Higman, V.A.; Klevit, R.E.; Van Rossum, B.J.; Oschkinat, H. Solid-state NMR and SAXS studies provide a structural basis for the activation of alphaB-crystallin oligomers. Nat. Struct. Mol. Biol. 2010, 17, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- McDonald, E.T.; Bortolus, M.; Koteiche, H.A.; McHaourab, H.S. Sequence, structure, and dynamic determinants of Hsp27 (HspB1) equilibrium dissociation are encoded by the N-terminal domain. Biochemistry 2012, 51, 1257–1268. [Google Scholar] [CrossRef] [Green Version]
- Rogalla, T.; Ehrnsperger, M.; Preville, X.; Kotlyarov, A.; Lutsch, G.; Ducasse, C.; Paul, C.; Wieske, M.; Arrigo, A.P.; Buchner, J.; et al. Regulation of Hsp27 oligomerization, chaperone function, and protective activity against oxidative stress/tumor necrosis factor alpha by phosphorylation. J. Biol. Chem. 1999, 274, 18947–18956. [Google Scholar] [CrossRef] [Green Version]
- Clouser, A.F.; Klevit, R.E. pH-dependent structural modulation is conserved in the human small heat shock protein HSBP1. Cell Stress Chaperones 2017, 22, 569–575. [Google Scholar] [CrossRef] [Green Version]
- Hanson, S.R.; Hasan, A.; Smith, D.L.; Smith, J.B. The major in vivo modifications of the human water-insoluble lens crystallins are disulfide bonds, deamidation, methionine oxidation and backbone cleavage. Exp. Eye Res. 2000, 71, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Chamorro, M.; Smith, D.L.; Smith, J.B. Identification of the major components of the high molecular weight crystallins from old human lenses. Curr. Eye Res. 1994, 13, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Lund, A.L.; Smith, J.B.; Smith, D.L. Modifications of the water-insoluble human lens alpha-crystallins. Exp. Eye Res. 1996, 63, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Cherian-Shaw, M.; Smith, J.B.; Jiang, X.Y.; Abraham, E.C. Intrapolypeptide disulfides in human alphaA-crystallin and their effect on chaperone-like function. Mol. Cell. Biochem. 1999, 199, 163–167. [Google Scholar] [CrossRef]
- Fan, X.; Zhou, S.; Wang, B.; Hom, G.; Guo, M.; Li, B.; Yang, J.; Vaysburg, D.; Monnier, V.M. Evidence of Highly Conserved beta-Crystallin Disulfidome that Can be Mimicked by In Vitro Oxidation in Age-Related Human Cataract and Glutathione Depleted Mouse Lens. Mol. Cell. Proteom. 2015, 14, 3211–3223. [Google Scholar] [CrossRef] [Green Version]
- Hains, P.G.; Truscott, R.J. Proteomic analysis of the oxidation of cysteine residues in human age-related nuclear cataract lenses. Biochim. Biophys. Acta 2008, 1784, 1959–1964. [Google Scholar] [CrossRef]
- Takemoto, L.J. Oxidation of cysteine residues from alpha-A crystallin during cataractogenesis of the human lens. Biochem. Biophys. Res. Commun. 1996, 223, 216–220. [Google Scholar] [CrossRef]
- Shroff, N.P.; Cherian-Shaw, M.; Bera, S.; Abraham, E.C. Mutation of R116C results in highly oligomerized alpha A-crystallin with modified structure and defective chaperone-like function. Biochemistry 2000, 39, 1420–1426. [Google Scholar] [CrossRef]
- Chiesa, R.; Spector, A. The dephosphorylation of lens alpha-crystallin A chain. Biochem. Biophys. Res. Commun. 1989, 162, 1494–1501. [Google Scholar] [CrossRef]
- Vermorken, A.J.; Bloemendal, H. alpha-Crystallin polypeptides as markers of lens cell differentiation. Nature 1978, 271, 779–781. [Google Scholar] [CrossRef]
- Spector, A.; Li, L.K.; Meretsky, D.; Augusteyn, R. What is alpha crystallin? Am. J. Ophthalmol. 1971, 71, 386–390. [Google Scholar] [CrossRef]
- Spector, A.; Li, L.-K.; Augusteyn, R.C.; Schneider, A.; Freund, T. α-Crystallin. The isolation and characterization of distinct macromolecular fractions. Biochem. J. 1971, 124, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.L.; Overbeek, P.A. Differential expression of alpha A- and alpha B-crystallin during murine ocular development. Investig. Ophthalmol. Vis. Sci. 1996, 37, 2276–2284. [Google Scholar]
- Peterson, J.J.; Young, M.M.; Takemoto, L.J. Probing alpha-crystallin structure using chemical cross-linkers and mass spectrometry. Mol. Vis. 2004, 10, 857–866. [Google Scholar]
- Santhoshkumar, P.; Murugesan, R.; Sharma, K.K. Deletion of (54)FLRAPSWF(61) residues decreases the oligomeric size and enhances the chaperone function of alphaB-crystallin. Biochemistry 2009, 48, 5066–5073. [Google Scholar] [CrossRef] [Green Version]
- Ryazantsev, S.N.; Poliansky, N.B.; Chebotareva, N.A.; Muranov, K.O. 3D structure of the native alpha-crystallin from bovine eye lens. Int. J. Biol. Macromol. 2018, 117, 1289–1298. [Google Scholar] [CrossRef]
- Li, L.K.; Spector, A. The reaggregation of purified subunits of alpha-crystallin. Exp. Eye Res. 1973, 15, 179–183. [Google Scholar] [CrossRef]
- Datta, S.A.; Rao, C.M. Packing-induced conformational and functional changes in the subunits of alpha -crystallin. J. Biol. Chem. 2000, 275, 41004–41010. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.X.; Liang, J.J. Intermolecular exchange and stabilization of recombinant human alphaA- and alphaB-crystallin. J. Biol. Chem. 1998, 273, 286–290. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, S.B.; Minton, A.P. Macromolecular crowding: Biochemical, biophysical, and physiological consequences. Annu. Rev. Biophys. Biomol. Struct. 1993, 22, 27–65. [Google Scholar] [CrossRef]
- Zimmerman, S.B.; Trach, S.O. Estimation of macromolecule concentrations and excluded volume effects for the cytoplasm of Escherichia coli. J. Mol. Biol. 1991, 222, 599–620. [Google Scholar] [CrossRef] [Green Version]
- Chebotareva, N.A.; Eronina, T.B.; Roman, S.G.; Mikhaylova, V.V.; Sluchanko, N.N.; Gusev, N.B.; Kurganov, B.I. Oligomeric state of alphaB-crystallin under crowded conditions. Biochem. Biophys. Res. Commun. 2019, 508, 1101–1105. [Google Scholar] [CrossRef] [PubMed]
- Vanhoudt, J.; Abgar, S.; Aerts, T.; Clauwaert, J. A small-angle X-ray solution scattering study of bovine alpha-crystallin. Eur. J. Biochem. 2000, 267, 3848–3858. [Google Scholar] [CrossRef] [PubMed]
- Foffi, G.; Savin, G.; Bucciarelli, S.; Dorsaz, N.; Thurston, G.M.; Stradner, A.; Schurtenberger, P. Hard sphere-like glass transition in eye lens alpha-crystallin solutions. Proc. Natl. Acad. Sci. USA 2014, 111, 16748–16753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klevit, R.E. Peeking from behind the veil of enigma: Emerging insights on small heat shock protein structure and function. Cell Stress Chaperones 2020. [Google Scholar] [CrossRef] [PubMed]
- Bepperling, A.; Alte, F.; Kriehuber, T.; Braun, N.; Weinkauf, S.; Groll, M.; Haslbeck, M.; Buchner, J. Alternative bacterial two-component small heat shock protein systems. Proc. Natl. Acad. Sci. USA 2012, 109, 20407–20412. [Google Scholar] [CrossRef] [Green Version]
- Mchaourab, H.S.; Dodson, E.K.; Koteiche, H.A. Mechanism of chaperone function in small heat shock proteins. Two-mode binding of the excited states of T4 lysozyme mutants by alphaA-crystallin. J. Biol. Chem. 2002, 277, 40557–40566. [Google Scholar] [CrossRef] [Green Version]
- Muhlhofer, M.; Berchtold, E.; Stratil, C.G.; Csaba, G.; Kunold, E.; Bach, N.C.; Sieber, S.A.; Haslbeck, M.; Zimmer, R.; Buchner, J. The Heat Shock Response in Yeast Maintains Protein Homeostasis by Chaperoning and Replenishing Proteins. Cell Rep. 2019, 29, 4593–4607. [Google Scholar] [CrossRef] [Green Version]
- Haslbeck, M. sHsps and their role in the chaperone network. Cell Mol. Life Sci. 2002, 59, 1649–1657. [Google Scholar] [CrossRef]
- Raman, B.; Rao, C.M. Chaperone-like activity and quaternary structure of alpha-crystallin. J. Biol. Chem. 1994, 269, 27264–27268. [Google Scholar]
- Jakob, U.; Gaestel, M.; Engel, K.; Buchner, J. Small heat shock proteins are molecular chaperones. J. Biol. Chem. 1993, 268, 1517–1520. [Google Scholar] [PubMed]
- Reddy, G.B.; Das, K.P.; Petrash, J.M.; Surewicz, W.K. Temperature-dependent chaperone activity and structural properties of human alphaA- and alphaB-crystallins. J. Biol. Chem. 2000, 275, 4565–4570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haslbeck, M.; Walke, S.; Stromer, T.; Ehrnsperger, M.; White, H.E.; Chen, S.; Saibil, H.R.; Buchner, J. Hsp26: A temperature-regulated chaperone. EMBO J. 1999, 18, 6744–6751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, J.; Das, K.P. Alpha-crystallin does not require temperature activation for its chaperone-like activity. Biochem. Mol. Biol. Int. 1998, 46, 249–258. [Google Scholar]
- Laskowska, E.; Wawrzynow, A.; Taylor, A. IbpA and IbpB, the new heat-shock proteins, bind to endogenous Escherichia coli proteins aggregated intracellularly by heat shock. Biochimie 1996, 78, 117–122. [Google Scholar] [CrossRef]
- Arrigo, A.P.; Suhan, J.P.; Welch, W.J. Dynamic changes in the structure and intracellular locale of the mammalian low-molecular-weight heat shock protein. Mol. Cell. Biol. 1988, 8, 5059–5071. [Google Scholar] [CrossRef] [Green Version]
- Stege, G.J.; Li, G.C.; Li, L.; Kampinga, H.H.; Konings, A.W. On the role of hsp72 in heat-induced intranuclear protein aggregation. Int. J. Hyperth. 1994, 10, 659–674. [Google Scholar] [CrossRef]
- Mainz, A.; Peschek, J.; Stavropoulou, M.; Back, K.C.; Bardiaux, B.; Asami, S.; Prade, E.; Peters, C.; Weinkauf, S.; Buchner, J.; et al. The chaperone alphaB-crystallin uses different interfaces to capture an amorphous and an amyloid client. Nat. Struct. Mol. Biol. 2015, 22, 898–905. [Google Scholar] [CrossRef]
- Mogk, A.; Bukau, B. Role of sHsps in organizing cytosolic protein aggregation and disaggregation. Cell Stress Chaperones 2017, 22, 493–502. [Google Scholar] [CrossRef]
- Specht, S.; Miller, S.B.; Mogk, A.; Bukau, B. Hsp42 is required for sequestration of protein aggregates into deposition sites in Saccharomyces cerevisiae. J. Cell Biol. 2011, 195, 617–629. [Google Scholar] [CrossRef]
- Ungelenk, S.; Moayed, F.; Ho, C.T.; Grousl, T.; Scharf, A.; Mashaghi, A.; Tans, S.; Mayer, M.P.; Mogk, A.; Bukau, B. Small heat shock proteins sequester misfolding proteins in near-native conformation for cellular protection and efficient refolding. Nat. Commun. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Grousl, T.; Ungelenk, S.; Miller, S.; Ho, C.T.; Khokhrina, M.; Mayer, M.P.; Bukau, B.; Mogk, A. A prion-like domain in Hsp42 drives chaperone-facilitated aggregation of misfolded proteins. J. Cell Biol. 2018, 217, 1269–1285. [Google Scholar] [CrossRef] [PubMed]
- Kampinga, H.H.; Brunsting, J.F.; Stege, G.J.; Konings, A.W.; Landry, J. Cells overexpressing Hsp27 show accelerated recovery from heat-induced nuclear protein aggregation. Biochem. Biophys. Res. Commun. 1994, 204, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Ehrnsperger, M.; Graber, S.; Gaestel, M.; Buchner, J. Binding of non-native protein to Hsp25 during heat shock creates a reservoir of folding intermediates for reactivation. EMBO J. 1997, 16, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Rauch, J.N.; Tse, E.; Freilich, R.; Mok, S.A.; Makley, L.N.; Southworth, D.R.; Gestwicki, J.E. BAG3 Is a Modular, Scaffolding Protein that Physically Links Heat Shock Protein 70 (Hsp70) to the Small Heat Shock Proteins. J. Mol. Biol. 2017, 429, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Hishiya, A.; Salman, M.N.; Carra, S.; Kampinga, H.H.; Takayama, S. BAG3 Directly Interacts with Mutated alphaB-Crystallin to Suppress Its Aggregation and Toxicity. PLoS ONE 2011, 6, e16828. [Google Scholar] [CrossRef] [Green Version]
- Alberti, S.; Mateju, D.; Mediani, L.; Carra, S. Granulostasis: Protein Quality Control of RNP Granules. Front. Mol. Neurosci. 2017, 10, 84. [Google Scholar] [CrossRef] [Green Version]
- Ulbricht, A.; Gehlert, S.; Leciejewski, B.; Schiffer, T.; Bloch, W.; Hohfeld, J. Induction and adaptation of chaperone-assisted selective autophagy CASA in response to resistance exercise in human skeletal muscle. Autophagy 2015, 11, 538–546. [Google Scholar] [CrossRef] [Green Version]
- Carra, S.; Brunsting, J.F.; Lambert, H.; Landry, J.; Kampinga, H.H. HspB8 participates in protein quality control by a non-chaperone-like mechanism that requires eIF2{alpha} phosphorylation. J. Biol. Chem. 2009, 284, 5523–5532. [Google Scholar] [CrossRef] [Green Version]
- Mounier, N.; Arrigo, A.P. Actin cytoskeleton and small heat shock proteins: How do they interact? Cell Stress Chaperones. 2002, 7, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Nicholl, I.D.; Quinlan, R.A. Chaperone activity of alpha-crystallins modulates intermediate filament assembly. EMBO J. 1994, 13, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.L.; Der Perng, M.; Prescott, A.R.; Jansen, K.A.; Koenderink, G.H.; Quinlan, R.A. The specificity of the interaction between alphaB-crystallin and desmin filaments and its impact on filament aggregation and cell viability. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayser, J.; Haslbeck, M.; Dempfle, L.; Krause, M.; Grashoff, C.; Buchner, J.; Herrmann, H.; Bausch, A.R. The small heat shock protein Hsp27 affects assembly dynamics and structure of keratin intermediate filament networks. Biophys. J. 2013, 105, 1778–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernik, I.S.; Seit-Nebi, A.S.; Marston, S.B.; Gusev, N.B. Small heat shock protein Hsp20 (HspB6) as a partner of 14-3-3gamma. Mol. Cell. Biochem. 2007, 295, 9–17. [Google Scholar] [CrossRef]
- Sathish, H.A.; Koteiche, H.A.; Mchaourab, H.S. Binding of destabilized betaB2-crystallin mutants to alpha-crystallin: The role of a folding intermediate. J. Biol. Chem. 2004, 279, 16425–16432. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Stein, R.A.; McHaourab, H.S. Cataract-linked gammaD-crystallin mutants have weak affinity to lens chaperones alpha-crystallins. FEBS Lett. 2012, 586, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Serebryany, E.; King, J.A. Wild-type human gammaD-crystallin promotes aggregation of its oxidation-mimicking, misfolding-prone W42Q mutant. J. Biol. Chem. 2015, 290, 11491–11503. [Google Scholar] [CrossRef] [Green Version]
- Van den IJssel, P.R.; Overkamp, P.; Knauf, U.; Gaestel, M.; De Jong, W.W. Alpha A-crystallin confers cellular thermoresistance. FEBS Lett. 1994, 355, 54–56. [Google Scholar] [CrossRef] [Green Version]
- Christopher, K.L.; Pedler, M.G.; Shieh, B.; Ammar, D.A.; Petrash, J.M.; Mueller, N.H. Alpha-crystallin-mediated protection of lens cells against heat and oxidative stress-induced cell death. Biochim. Biophys. Acta 2014, 1843, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Alderson, T.R.; Roche, J.; Gastall, H.Y.; Dias, D.M.; Pritisanac, I.; Ying, J.; Bax, A.; Benesch, J.L.P.; Baldwin, A.J. Local unfolding of the HSP27 monomer regulates chaperone activity. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Haslbeck, M.; Weinkauf, S.; Buchner, J. Regulation of the Chaperone Function of Small Hsps. In The Big Book on Small Heat Shock Proteins; Tanguay, R.M., Hightower, L.E., Eds.; Springer International Publishing AG: Cham, Switzerland, 2015; pp. 155–176. [Google Scholar]
- Wu, D.; Vonk, J.J.; Salles, F.; Vonk, D.; Haslbeck, M.; Melki, R.; Bergink, S.; Kampinga, H.H. The N terminus of the small heat shock protein HSPB7 drives its polyQ aggregation-suppressing activity. J. Biol. Chem. 2019, 294, 9985–9994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnova, E.; Chebotareva, N.; Gurvits, B. Transient transformation of oligomeric structure of alpha-crystallin during its chaperone action. Int. J. Biol. Macromol. 2013, 55, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Miesbauer, L.R.; Zhou, X.; Yang, Z.; Yang, Z.; Sun, Y.; Smith, D.L.; Smith, J.B. Post-translational modifications of water-soluble human lens crystallins from young adults. J. Biol. Chem. 1994, 269, 12494–12502. [Google Scholar] [PubMed]
- Wunderlich, M.; Glockshuber, R. Redox properties of protein disulfide isomerase (DsbA) from Escherichia coli. Protein Sci. 1993, 2, 717–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preville, X.; Salvemini, F.; Giraud, S.; Chaufour, S.; Paul, C.; Stepien, G.; Ursini, M.V.; Arrigo, A.P. Mammalian small stress proteins protect against oxidative stress through their ability to increase glucose-6-phosphate dehydrogenase activity and by maintaining optimal cellular detoxifying machinery. Exp. Cell Res. 1999, 247, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Arrigo, A.P. Hsp27: Novel regulator of intracellular redox state. IUBMB Life 2001, 52, 303–307. [Google Scholar] [CrossRef]
- Mehlen, P.; Kretz-Remy, C.; Preville, X.; Arrigo, A.P. Human hsp27, Drosophila hsp27 and human alphaB-crystallin expression-mediated increase in glutathione is essential for the protective activity of these proteins against TNFalpha-induced cell death. EMBO J. 1996, 15, 2695–2706. [Google Scholar] [CrossRef]
- Bova, M.P.; Ding, L.L.; Horwitz, J.; Fung, B.K. Subunit exchange of alphaA-crystallin. J. Biol. Chem. 1997, 272, 29511–29517. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riedl, M.; Strauch, A.; Catici, D.A.M.; Haslbeck, M. Proteinaceous Transformers: Structural and Functional Variability of Human sHsps. Int. J. Mol. Sci. 2020, 21, 5448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155448
Riedl M, Strauch A, Catici DAM, Haslbeck M. Proteinaceous Transformers: Structural and Functional Variability of Human sHsps. International Journal of Molecular Sciences. 2020; 21(15):5448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155448
Chicago/Turabian StyleRiedl, Mareike, Annika Strauch, Dragana A.M. Catici, and Martin Haslbeck. 2020. "Proteinaceous Transformers: Structural and Functional Variability of Human sHsps" International Journal of Molecular Sciences 21, no. 15: 5448. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21155448