Roles of Non-Coding RNAs in Response to Nitrogen Availability in Plants


1
Institute for Systems Genetics and Department of Biochemistry and Molecular Pharmacology, New York University Langone Health, New York, NY 10016, USA
2
Department of Applied Biological Chemistry, Graduate School of Agricultural and Life Sciences, The University of Tokyo, 1-1-1 Yayoi, Bunkyo-ku, Tokyo 113-8657, Japan
3
Department of Bioresource Science, Faculty of Agriculture, Saga University, 1 Honjo-machi, Saga 840-8502, Japan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(22), 8508; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228508
Submission received: 22 October 2020
/
Revised: 6 November 2020
/
Accepted: 10 November 2020
/
Published: 12 November 2020
(This article belongs to the Special Issue The Molecular Basis of Carbon and Nitrogen Metabolism in Plants)
{kind=link}
{kind=link}
Abstract
:Nitrogen (N) is an essential nutrient for plant growth and development; therefore, N deficiency is a major limiting factor in crop production. Plants have evolved mechanisms to cope with N deficiency, and the role of protein-coding genes in these mechanisms has been well studied. In the last decades, regulatory non-coding RNAs (ncRNAs), such as microRNAs (miRNAs), small interfering RNAs (siRNAs), and long ncRNAs (lncRNAs), have emerged as important regulators of gene expression in diverse biological processes. Recent advances in technologies for transcriptome analysis have enabled identification of N-responsive ncRNAs on a genome-wide scale. Characterization of these ncRNAs is expected to improve our understanding of the gene regulatory mechanisms of N response. In this review, we highlight recent progress in identification and characterization of N-responsive ncRNAs in Arabidopsis thaliana and several other plant species including maize, rice, and Populus.
1. Introduction
Plants require 17 essential elements (C, H, O, N, P, K, Mg, Ca, S, Cl, Fe, B, Mn, Zn, Cu, Ni, and Mo) to complete their life cycle [1]. Nitrogen (N) is an essential macronutrient, and 1–5% of total plant dry matter consists of N. The major sources of N are nitrate and ammonium, which are absorbed from the soil via nitrate transporters (NRTs) and ammonium transporters (AMTs), respectively [1]. Absorbed N is assimilated and used as a source of metabolites critical for plant survival, such as amino acids, nucleic acids, chlorophyll, coenzymes, phytohormones, and secondary metabolites [1]. It is therefore a major limiting factor for plant growth and crop production [2]. Availability of N is affected by soil properties, such as pH and moisture [3], and plants have evolved sophisticated mechanisms to adapt to variable N availability.
Plants optimize the activities of N absorption and assimilation in response to N availability in soils by regulating expression of genes involved in these processes. In Arabidopsis thaliana, expression of genes encoding high-affinity NRTs (AtNRT2.1, AtNRT2.4, and AtNRT2.5) is induced in roots under N deficiency [4]. In contrast, under N sufficiency, AtNRT2 genes are down-regulated, and nitrate absorption is switched to low-affinity mode via the AtNRT1 family [5,6]. Genes encoding nitrate reductase (NR) and nitrite reductase (NiR) are induced by nitrate supply [7,8] and facilitate its assimilation, whereas these genes are down-regulated under N deficiency, which is likely in order to limit nitrate assimilation activity under low-nitrate supply. In A. thaliana, several transcription factors are responsible for transcriptional regulation of these genes [9]. NIN-like protein (NLP) transcription factors are responsible for regulation of NR and NiR expression in response to nitrate supply [10,11,12]. NITRATE-INDUCIBLE GARP-TYPE TRANSCRIPTIONAL REPRESSOR 1 proteins (NIGT1s) directly repress AtNRT2 and suppress high-affinity nitrate transport under N sufficiency [13,14]. Expression of NIGT1s is negatively autoregulated and is also regulated by NLPs. Integration of the NLP–NIGT1 cascade and NIGT1 autoregulation seems to orchestrate the nitrate response [13].
Modification of root architecture is also important for plant adaptation to changing N availability [15,16,17,18] and has been well studied in A. thaliana. Lateral root (LR) growth is stimulated under moderate N deficiency (275 or 550 µM in media [19]), enabling plants to explore available N. Stimulation of LR growth under moderate N deficiency is thought to be caused by induction of TRYPTOPHAN AMINOTRANSFERASE-RELATED 2 (TAR2), an auxin biosynthetic gene involved in development of LR primordia [20]. By contrast, under severe N deficiency, LR growth is repressed [19,21,22], which likely restricts investment in extending roots into N-poor environments [22]. Inhibition of LR growth under severe N deficiency is mediated by CLAVATA3/ESR-related (CLE) signaling peptides and their receptor protein, CLAVATA1 (CLV1) [22,23]. CLE genes (CLE1, CLE3, CLE4, and CLE7) are induced in response to N deficiency in root cells, and accumulated CLE peptides interact with CLV1, resulting in repression of genes involved in LR development [22,23]. Repression of LR growth under severe N deficiency is also caused by NRT1.1: NRT1.1 transports not only nitrate but also auxin and facilitates shootward auxin movement in LRs, resulting in reduced auxin concentration in LR tips and repressed LR growth [21].
Despite these advances in research on plant response to N availability, we are still far from understanding the molecular mechanisms underlying complex spatiotemporal regulation of hundreds of genes in response to N availability. Over the last decade, a large number of non-coding RNAs (ncRNAs) have been discovered that are distinct from well-known housekeeping RNAs such as ribosomal RNA, transfer RNA, small nuclear RNA, and small nucleolar RNA [24]. The proportion of functional ncRNAs remains unclear, but dozens of ncRNAs regulate gene expression at transcriptional and post-transcriptional levels [25,26]. In plants, expression of many ncRNAs responds to various environmental stimuli [27,28,29,30,31,32,33,34,35,36], indicating their roles in tolerance to environmental stresses such as drought, salinity, heat, cold, and nutrient deficiency [37,38,39]. Advances in RNA sequencing (RNA-Seq) technology have enabled identification of ncRNAs with expression affected by N availability (N-responsive ncRNAs) in various plants including A. thaliana, Populus (Populus tomentosa), rice (Oryza sativa), and maize (Zea mays) [30,40,41,42,43]. Very recently, the role of N-responsive ncRNAs in plant response to N availability has been demonstrated. In this review, we summarize identification and characterization of N-responsive ncRNAs and discuss their importance in plant response to N availability.
2. Classification and Functions of Plant ncRNAs
2.1. Classification of ncRNAs
ncRNAs can be either housekeeping or regulatory. The former group has a housekeeping role in transcription and translation, whereas the latter regulates genes [44]. Regulatory ncRNAs are further classified into small ncRNAs (microRNAs, miRNAs, small interfering RNAs, and siRNAs), and long ncRNAs (lncRNAs, typically > 200 nt and lacking the potential to code proteins > 100 amino acid residues) [39].
2.2. Small Regulatory ncRNAs
The major difference between miRNAs and siRNAs is their biogenesis: miRNAs are processed from transcripts that can form local RNA hairpin precursor structures, whereas siRNAs are processed from long bimolecular RNA duplexes or extended hairpins [45]. Initially, miRNAs are transcribed from miRNA host genes (MIR genes), predominantly by RNA polymerase II, as precursors called primary miRNAs (pri-miRNAs). Mammalian miRNA host genes often encode multiple miRNAs and are frequently nested within protein-coding genes, whereas their plant counterparts are independent transcription units with a single miRNA [46]. Pri-miRNAs form intra-molecular hairpin-like structures and are processed into mature miRNAs (typically 20–22 nt) by DICER-LIKE (DCL) RNase III endonucleases, mainly DCL1 [25,47,48,49]. Mature miRNAs are loaded into ARGONAUTE 1 (AGO1) to form the RNA-induced silencing complex (RISC) and then guide the RISC to the target mRNA through sequence complementarity and trigger gene silencing [50,51,52].
Depending on the organism and the cellular context, siRNA precursors, double-stranded RNAs (dsRNAs), may arise from bidirectional transcription (sense and antisense transcripts) of a chromosomal locus or from unidirectional transcription followed by RNA-dependent RNA polymerase activity [53]. Endogenous plant siRNAs can be grouped into three classes [54]. A major class is generated from heterochromatic regions (such as centromeres, transposons, and other repetitive elements); these precursors are transcribed predominantly by the plant-specific polymerase Pol IV, followed by processing into so-called heterochromatic siRNAs (hc-siRNAs; 24 nt) [55,56]. These hc-siRNAs induce DNA methylation and histone modifications through RNA-directed DNA methylation [57,58]; their mode of action is consistent with the idea that RNA interference (RNAi) functions in silencing transposon expression and propagation [59]. Another class of siRNAs, so-called secondary siRNAs, require an initial small-RNA-directed cleavage of the primary transcript to trigger dsRNA synthesis [60,61,62]. Among siRNAs of this class, trans-acting siRNAs (tasiRNAs) are best studied and, unlike many other siRNAs, act in trans to direct post-transcriptional repression of their mRNA targets [60]. Precursors of the third siRNA class are formed from mRNAs encoded by natural cis-antisense gene pairs, which are transcribed from opposite DNA strands of the same genomic loci (cis-NATs) [63]. Similar to the case of miRNAs, siRNA precursors are processed by DCLs into small RNAs and then loaded into AGO proteins to form the RISC [64,65,66,67], which silences a target gene.
In addition to their role in transcriptional silencing, intercellular mobility of small RNAs is another important aspect to be considered. Small RNAs can move either locally (cell-to-cell) or systemically (organ-to-organ) and play a role as mobile silencing signals. Several nutrient-responsive miRNAs are known to be accumulated in the phloem during starvation, indicating that those miRNAs are systemically translocated in the plant to amplify silencing signals (reviewed in [68]). Such a non-cell autonomous effect of small RNAs likely enables rapid adaptation to changing environments.
2.3. Long Regulatory ncRNAs
LncRNAs are transcribed from intergenic, intronic, and coding regions, and regulate gene expression [26,69]. Some lncRNAs are precursors for siRNAs or miRNAs, whereas others function without being processed into small RNAs [26]. Some lncRNAs function as molecular decoys that sequester proteins or small RNAs from their target RNA as in the case of IPS1 [70] and ASCO [71]. LncRNAs can also associate with chromatin and control gene transcription. HIDDEN TREASURE 1 (HID1) interacts with the promoter region of the transcription factor gene PIF3 and represses its expression to regulate photomorphogenesis [72]. The well-known flowering-related lncRNAs COLDAIR, COLDWRAP, and COOLAIR are transcribed from FLOWERING LOCUS C (FLC) and interact with Polycomb proteins, thereby controlling epigenetic silencing of FLC mediated by histone modifications [73,74,75].
2.4. Plant ncRNA Databases
Although mammalian ncRNAs remain the best studied, with the help of recent advances in sequencing technologies, identification of plant ncRNAs has almost caught up with the mammalian field. Several databases for plant ncRNAs have been established [24]; the A. thaliana small RNA project (ASRP) database contains 218,585 unique small RNA sequences [76], and GREENC contains more than 120,000 lncRNAs from 37 plant species [77]. Many plant ncRNAs are expressed in response to abiotic stresses including drought, temperature, salinity, and nutrient deprivation [27,28,30,78]. Recently, a database of experimentally validated stress-responsive ncRNAs, named PncStress, has been established, which contains 4227 entries including lncRNAs and miRNAs from 114 plant species and covers 91 abiotic stresses and 48 biotic stresses [79].
3. N-Responsive ncRNAs in A. thaliana
3.1. miRNAs Involved in N Nutrition
Among N-responsive ncRNAs in A. thaliana, miRNAs have been relatively well studied. Several miRNAs have been characterized as associated with N stresses [80,81,82,83]. The first report on the role of a miRNA in N response was identification of miR167 and its target gene AUXIN RESPONSE FACTOR 8 (ARF8); nitrate influx suppresses expression of miR167, allowing ARF8 to accumulate in the root pericycle cells and to initiate LR outgrowth [84]. N-starvation-responsive miRNAs miR160 and miR171 [81] regulate root system architecture by suppressing expression of their target transcription factor genes ARF10/16/17 and SCARECROW-LIKE PROTEIN 6 (SCL6), respectively [85,86]. miR393 and its target AUXIN-SIGNALING F-BOX PROTEIN 3 (AFB3) also control the responses of root system architecture to both external and internal N [87,88]. Expression of AFB3 is induced by nitrate and promotes LR formation, whereas miR393 is induced by N metabolites, thus allowing plants to respond appropriately to N conditions [87]. miR169 down-regulates N uptake by inhibiting expression of its target gene, NUCLEAR FACTOR Y SUBUNIT A5 (NFYA5), a positive regulator of nitrate transporter genes AtNRT2.1 and AtNRT1.1 [89]. miR826 and miR5090 regulate N metabolism through their common target gene ALKENYL HYDROXALKYL PRODUCING 2 (AOP2), which encodes 2-oxoglutarate-dependent dioxygenase involved in synthesis of N-containing metabolites called glucosinolates [90]. Under N starvation, expression of AOP2 is post-transcriptionally repressed by miR826 and miR5090, decreasing glucosinolate synthesis and thus enabling plants to distribute N to metabolites required for survival [91]. Interestingly, both miRNAs may have evolved by inverted duplication of the genomic region containing AOP2, implying coevolution of miR826, miR5090, and AOP2 in adaptation to N deprivation [91,92].
3.2. Genome-Wide Identification of N-Responsive miRNAs
N-responsive miRNAs have been identified by isolation of small RNAs from total RNA by size fractionation followed by sequencing. Initially, a genome-wide search of nitrate-induced miRNAs using 454-sequencing technology identified miR393 [93]. Subsequently, using Illumina sequencing, Liang et al. [81] reported expression profiles of 177 miRNAs under N-sufficient and deficient conditions; some of these miRNAs may be involved in root system development in response to N [81]. The authors also identified miRNAs responsive to carbon and sulfate, which may help to explain crosstalk between different nutrient responses [94]. Hundreds of N-responsive miRNAs have been identified, but their roles remain largely uncharacterized.
3.3. T5120 and TAS3: lncRNAs Involved in N Response
Recently, T5120 and TAS3 have been reported to be involved in N response [30,40]. Expression of T5120 was significantly induced by 2h treatment of A. thaliana roots with nitrate [40]. Induction was mediated by direct binding of transcription factor NLP7 to the promoter region in a nitrate-dependent manner. NLP7 acts on nitrate-responsive genes, such as NRT2.1, NIR, NRT2.2, and NIA, and orchestrates early response to nitrate [95]. In transgenic A. thaliana, overexpression of T5120 significantly increases nitrate reductase activity, amino acid content, seedling biomass, and expression of nitrate-assimilatory genes including NIA1, NIA2, NIR, and GLN1.1, whereas nitrate uptake is not affected [40]. Interestingly, T5120 gene is positioned adjacent to NIA1 on the chromosome and might regulate its expression by affecting chromatin structure or modification [40], as in the case of lncRNAs APOLO, and COOLAIR [73,96]. Further investigation of regulatory mechanisms behind T5120-mediated control of N assimilation is required for T5120 engineering to develop crops with improved N use efficiency.
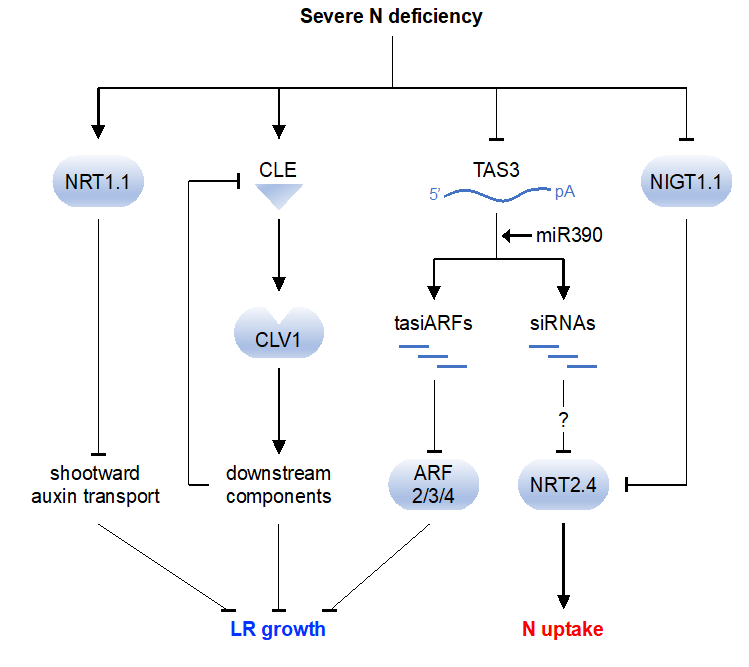
TAS3 is a precursor of tasiRNAs called tasiARFs, which target ARF2, ARF3, and ARF4 [97]. TAS3 transcripts are bound and cleaved by miR390, an evolutionarily conserved miRNA, which triggers tasiARF production [60]. The miR390/TAS3/ARF pathway is part of the auxin-mediated molecular network that orchestrates LR formation in A. thaliana [98,99]. Recently, it was found by RNA-Seq analysis that expression of TAS3 is significantly reduced under severe N deficiency [30]. Because TAS3 positively regulates LR growth [98], we hypothesized that TAS3 down-regulation in response to N deficiency suppresses LR outgrowth by increasing expression of ARFs. Consistent with this hypothesis, expression of ARF2–4, especially ARF4, is up-regulated by N deficiency, whereas TAS3 overexpression down-regulates ARF4 and significantly increases LR length, even under severe N deficiency [30]. Interestingly, we found that TAS3 decreases not only expression of ARFs but also that of NRT2.4 [100]. Our computational analysis suggests an interaction between TAS3-derived small RNA and the NRT2.4 transcript [30]. We confirmed TAS3-dependent cleavage of the NRT2.4 transcript by rapid amplification of cDNA ends [30]. These observations indicate the role of TAS3 as a source of multiple tasiRNAs that regulate multiple signaling pathways depending on N availability. Inhibition of LR growth in response to severe N deficiency has been explained by NRT1.1-mediated auxin transport [21] and CLE–CLV1 peptide-receptor signaling module [22,23]. We propose an additional regulatory mechanism involving TAS3-derived tasiARFs (Figure 1). In parallel, TAS3 might also regulate N uptake cooperative with the NIGT1.1/NRT2.4 transcriptional cascade [14,100] (Figure 1).
3.4. Genome-Wide Identification of N-Responsive lncRNAs
RNA-Seq data have provided a catalog of N-responsive lncRNAs. Liu et al. [40] analyzed the root transcriptome of 7-day-old A. thaliana roots treated with 10 mM KNO3 for 2 h and identified 7 up-regulated and 1 down-regulated lncRNAs among a total of 695 lncRNAs (|log2 fold change| ≥ 1 vs. untreated controls) [40]. We have analyzed the transcriptome of A. thaliana roots exposed to 12 different nutrient starvation conditions (-N, -K, -Ca, -Mg, -P, -S, -B, -Fe, -Mn, -Zn, -Cu, or -Mo) for 4 days and identified 60 lncRNAs differentially expressed (|log2 fold change| ≥ 1) under at least one nutrient deficiency in comparison with nutrient-sufficient controls [30,101]. Among them, 6 lncRNAs were up-regulated and 15 were down-regulated in response to N deficiency. Among these 21 lncRNAs, AT1G11185, AT1G67105, and TAS3 have been also reported to be N responsive by Liu et al. [40]. For further characterization of lncRNAs, we have developed a bioinformatics pipeline to predict molecular interactions between lncRNAs and mRNAs, and have identified hundreds of lncRNA–mRNA pairs with low interaction energy (high interaction potential) and high correlation of expression patterns. Among these pairs, AT1G67105 was predicted to bind the mRNA of an ammonium transporter gene AMT1;2 [30,102]. Under N deficiency, AT1G67105 is down-regulated, whereas AMT1;2 is up-regulated, indicating the possible role of AT1G67105 in regulation of ammonium uptake by controlling expression of AMT1;2 [30]. Our study provided tens of candidate lncRNAs that function in low-nutrient responses and proposed an approach for predicting their target genes. This approach would help to explore how those lncRNAs contribute to gene regulation under nutrient deficiencies.
4. N-Responsive ncRNAs in Other Plant Species
4.1. Maize ncRNAs
Genome-wide microarray-based analysis identified 14 differentially expressed miRNAs in maize (miR160, miR164, miR167, miR168, miR169, miR172, miR319, miR395, miR397, miR398, miR399, miR408, miR528, and miR827) under chronic (15 days) or transient (2 h) N deficiency [103]. They were categorized into miRNAs targeting (i) transcription factors (e.g., miR167–ARF8 [104]), (ii) genes involved in energy metabolism (e.g., miR398–COX [105]), and (iii) genes involved in miRNA regulation (e.g., miR168–AGO1 [106,107]). Using in situ hybridization, Trevisan et al. [108,109] characterized spatial distribution of N-responsive miRNAs miR528a/b, miR169i/j/k, miR166j/k/n, and miR408/b under different N conditions [108]. Deep sequencing technologies have accelerated identification of N-responsive miRNAs. Of a total of 85 miRNAs newly identified by small-RNA sequencing, 25 showed > two-fold relative change in response to low N [110,111]. Another study reported that 106 miRNAs were differentially expressed between the control and N-deficient groups and 103 between N-deficient and N-resupply groups [112].
Of 7245 lncRNAs identified by RNA-Seq analysis in maize leaves under N sufficiency or N deficiency, 637 were responsive to low N [113]. Coexpression analyses suggested that most of those lncRNAs are involved in energy metabolic pathways related to NADH dehydrogenase activity, oxidative phosphorylation, and the N compound metabolic process; however, the exact function of each lncRNA remains uncharacterized.
4.2. Rice ncRNAs
Expression of miRNAs in rice roots exposed to different N sources and concentrations has been comprehensively analyzed by small RNA-Seq [114]. In N-starved rice shoots and roots, a combination of different sequencing datasets (strand-specific RNA-Seq, small RNA-Seq, poly(A)-primed sequencing (2P-Seq), and degradome sequencing) has provided comparative expression profiles of protein-coding genes, lncRNAs, and miRNAs along with their putative target genes [42]. This study identified 918 differentially expressed lncRNAs and 91 target genes of 40 miRNAs [42]. The authors have proposed a new model for small RNA generation from two cis-NAT pairs, AMT1.1–cis-NATAMT1.1 and AMT1.2–cis-NATAMT1.2. In general, mRNA–cis-NAT pairs whose partial or entire sequences are complementary to other transcripts form dsRNAs that can be cleaved by a Dicer endonuclease to produce small RNAs [63,115]. However, the authors have shown that small RNAs are generated from AMT1 mRNA itself rather than from dsRNAs, indicating a different mechanism of small RNA biogenesis [42]. As cis-NATs are transcribed from the AMT gene loci of other species such as A. thaliana, barley (Hordeum vulgare), and maize, cis-NATAMT might have an evolutionally conserved role in N response.
4.3. Populus ncRNAs
Populus is a fast-growing tree of high economic and environmental value, especially in timber production and forestry. Expression profiling of ncRNAs in P. tomentosa plantlets under N deficiency has recently been performed [41,116,117]. Using small RNA-Seq and degradome sequencing, 60 N-responsive miRNAs were identified including 39 conserved, 13 non-conserved, and 8 novel miRNAs, along with their potential target genes [116,117]. Chen et al. [41] identified 126 differentially expressed lncRNAs in N-starved plantlets and classified them into pri-miRNAs, miRNA targets, and antisense lncRNAs. These studies lay the foundation for further studies on molecular mechanisms of N response in Populus and will help to engineer woody plants with improved N-use efficiency. There is no report available on genome-wide identification of N-responsive ncRNAs in other tree species at this time.
4.4. ncRNAs in Other Crops
Further to the plant species described above, N-responsive miRNAs have also been identified from dicot crop species including soybean (Glycine max) [118], potato (Indian potato variety Kufri Jyoti) [119], tea plant (Camellia sinensis) [120], and from other monocot species such as barley (Hordeum vulgare) [121] and wheat (Triticum aestivum and Triticum turgidum) [122,123,124,125], while N-responsive lncRNAs have yet to be systematically explored.
5. Evolution of ncRNAs
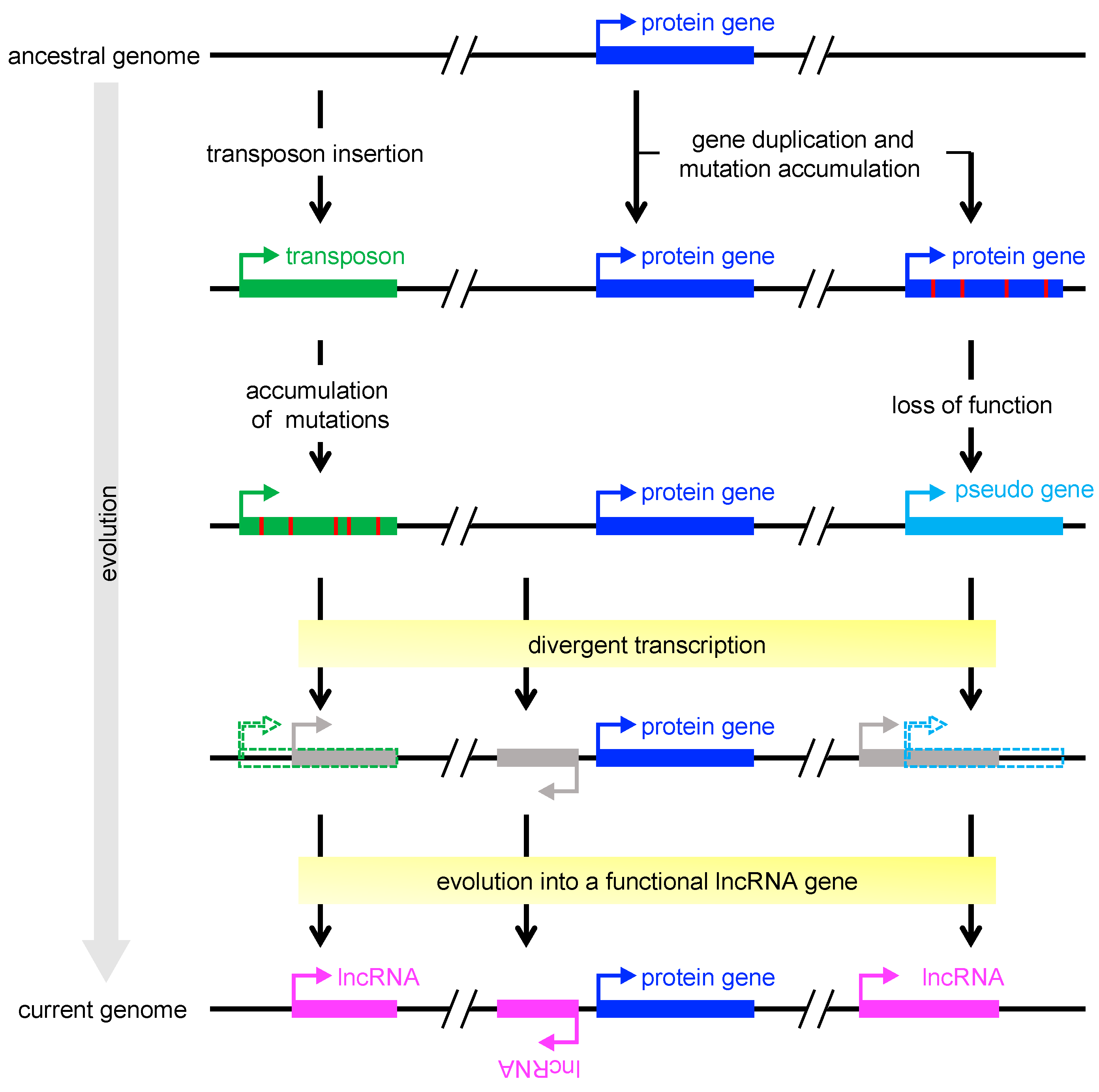
It is now obvious that the plant genome produces tens of thousands of lncRNAs [126]. The sequences of lncRNAs are less conserved than those of mRNAs [127,128,129], and lncRNAs are often transcribed in a strict stress-specific manner [37], implying that many of them play an ancillary rather than essential role. This raises the question of why the genome contains so many ncRNA genes, comparable to the number of protein-coding genes. The relatively low essentiality of ncRNAs could be explained by their origins. Investigations of the origin of lncRNAs have focused on possible contributions of protein-coding genes and transposons. Transposable elements (TEs) are a major force shaping the lncRNA repertoire through their capacity to introduce regulatory sequences essential for transcription, such as transcription initiation, splice, and polyadenylation sites, upon chromosomal insertion both in mammals and plants [130,131,132]. In plants, most of non-TE lncRNAs (ranging from 50% to 70%, depending on species) are predicted to originate from divergent transcription at promoters of active protein-coding genes [133]. Another large fraction (20.2% on average) of non-TE lncRNAs are located within the 2-kb proximal upstream regions of pseudogenes [133], indicating that pseudogenes also contribute to the makeup, evolutionary origins, and regulation of lncRNAs (Figure 2). In either case, lncRNA loci have originated through pre-existing gene regulatory sequences; therefore, they are likely to have evolved later than most protein-coding genes. lncRNAs seem to have evolved to modulate pre-existing gene networks and, for example, improve adaptation to environmental stresses for organism survival. Such adaptive evolution is advantageous, especially for plants, which are sessile. Identification of miRNAs from various plant species revealed that plant MIR genes have originated mainly by duplication of pre-existing MIR genes or protein-coding genes [134]. In addition, only a few miRNA families are conserved [135], suggesting that the majority of miRNAs have also emerged later than protein-coding genes. Plants seem to have evolved and diversified MIR genes in a lineage-specific manner, which may have allowed organisms to develop more complex gene regulatory networks to adapt to various stress conditions. From a practical perspective, ncRNAs are attractive targets of genome engineering to enhance the stress tolerance of crops without disrupting any essential genes.
6. Conclusions and Perspectives
N response in plants has been extensively studied from multiple perspectives such as morphology, development, fertility, metabolites, hormonal signaling, and gene expression. However, how the expression of N-responsive genes is differentially and appropriately regulated depending on N conditions remains to be further investigated. ncRNAs are important regulators of gene expression, and molecular and functional characterization of N-responsive ncRNAs will extend our understanding of gene regulatory mechanisms in N response. Although a large number of N-responsive ncRNAs have been identified in several different plant species over the last decades, dissecting their role and regulation remains a major challenge. Further studies on ncRNAs that provide experimental validation of their biological function are of great importance in the coming decades. Moreover, in silico approaches to predict ncRNA functions using multiple sources of different high-throughput data are necessary to accelerate ncRNA characterization [136]. Engineering of N-responsive ncRNAs could facilitate development of crops with improved N-use efficiency and/or low-N tolerance. For example, overexpression of miR169o, which is down-regulated by N deficiency, significantly improves N use efficiency in rice [137]. This approach could increase crop yields and promote sustainable agriculture while reducing use of N fertilizers and environmental pollution [138]. However, ncRNA overexpression using a constitutive promoter may be prone to cause undesirable effects, such as reduced resistance to bacterial infection as is the case with miR169o overexpression [137]. Advanced strategies for ncRNA engineering that allow more precise and specific control of expression and/or function of ncRNA will be required.
Author Contributions
M.F. and S.N. wrote the manuscript with support from T.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by JSPS KAKENHI Grant Number JP18KK0426.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Chapter 6—Functions of Macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar] [CrossRef]
- Engels, C.; Kirkby, E.; White, P. Chapter 5—Mineral Nutrition, Yield and Source–Sink Relationships. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar] [CrossRef]
- Daniels, W.L. The Nature and Properties of Soils, 15th ed.; Pearson Education: London, UK, 2016; Volume 80, p. 1428. [Google Scholar] [CrossRef]
- Orsel, M.; Krapp, A.; Daniel-Vedele, F. Analysis of the NRT2 Nitrate Transporter Family in Arabidopsis. Structure and Gene Expression. Plant Physiol. 2002, 129, 886–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, N.C.; Liu, K.H.; Lo, H.J.; Tsay, Y.F. Cloning and Functional Characterization of an Arabidopsis Nitrate Transporter Gene That Encodes a Constitutive Component of Low-Affinity Uptake. Plant Cell 1999, 11, 1381–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, N.C.; Chiang, C.S.; Crawford, N.M.; Tsay, Y.F. CHL1 encodes a component of the low-affinity nitrate uptake system in Arabidopsis and shows cell type-specific expression in roots. Plant Cell 1996, 8, 2183–2191. [Google Scholar] [CrossRef] [Green Version]
- Tang, P.S.; Wu, H.Y. Adaptive Formation of Nitrate Reductase in Rice Seedlings. Nature 1957, 179, 1355–1356. [Google Scholar] [CrossRef]
- Wang, R.; Guegler, K.; LaBrie, S.T.; Crawford, N.M. Genomic Analysis of a Nutrient Response in Arabidopsis Reveals Diverse Expression Patterns and Novel Metabolic and Potential Regulatory Genes Induced by Nitrate. Plant Cell 2000, 12, 1491–1509. [Google Scholar] [CrossRef]
- Yanagisawa, S. Transcription factors involved in controlling the expression of nitrate reductase genes in higher plants. Plant Sci. 2014, 229, 167–171. [Google Scholar] [CrossRef]
- Castaings, L.; Camargo, A.; Pocholle, D.; Gaudon, V.; Texier, Y.; Boutet-Mercey, S.; Taconnat, L.; Renou, J.-P.; Daniel-Vedele, F.; Fernandez, E.; et al. The nodule inception-like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J. 2009, 57, 426–435. [Google Scholar] [CrossRef]
- Wang, R.; Xing, X.; Wang, Y.; Tran, A.; Crawford, N.M. A Genetic Screen for Nitrate Regulatory Mutants Captures the Nitrate Transporter Gene NRT1.1. Plant Physiol. 2009, 151, 472–478. [Google Scholar] [CrossRef] [Green Version]
- Konishi, M.; Yanagisawa, S. Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nat. Commun. 2013, 4, 1617. [Google Scholar] [CrossRef] [Green Version]
- Maeda, Y.; Konishi, M.; Kiba, T.; Sakuraba, Y.; Sawaki, N.; Kurai, T.; Ueda, Y.; Sakakibara, H.; Yanagisawa, S. A NIGT1-centred transcriptional cascade regulates nitrate signalling and incorporates phosphorus starvation signals in Arabidopsis. Nat. Commun. 2018, 9, 1376. [Google Scholar] [CrossRef] [Green Version]
- Kiba, T.; Inaba, J.; Kudo, T.; Ueda, N.; Konishi, M.; Mitsuda, N.; Takiguchi, Y.; Kondou, Y.; Yoshizumi, T.; Ohme-Takagi, M.; et al. Repression of Nitrogen Starvation Responses by Members of the Arabidopsis GARP-Type Transcription Factor NIGT1/HRS1 Subfamily. Plant Cell 2018, 30, 925–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Forde, B.G. Regulation of Arabidopsis root development by nitrate availability. J. Exp. Bot. 2000, 51, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.G. Nitrogen signalling pathways shaping root system architecture: An update. Curr. Opin. Plant Biol. 2014, 21, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Kiba, T.; Krapp, A. Plant Nitrogen Acquisition Under Low Availability: Regulation of Uptake and Root Architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Chen, F.; Yuan, L.; Mi, G. The physiological mechanism underlying root elongation in response to nitrogen deficiency in crop plants. Planta 2020, 251, 84. [Google Scholar] [CrossRef] [PubMed]
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; von Wirén, N. Plasticity of the Arabidopsis Root System under Nutrient Deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [Green Version]
- Ma, W.; Li, J.; Qu, B.; He, X.; Zhao, X.; Li, B.; Fu, X.; Tong, Y. Auxin biosynthetic gene TAR2 is involved in low nitrogen-mediated reprogramming of root architecture in Arabidopsis. Plant J. 2014, 78, 70–79. [Google Scholar] [CrossRef]
- Krouk, G.; Lacombe, B.; Bielach, A.; Perrine-Walker, F.; Malinska, K.; Mounier, E.; Hoyerova, K.; Tillard, P.; Leon, S.; Ljung, K.; et al. Nitrate-Regulated Auxin Transport by NRT1.1 Defines a Mechanism for Nutrient Sensing in Plants. Dev. Cell 2010, 18, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Araya, T.; Miyamoto, M.; Wibowo, J.; Suzuki, A.; Kojima, S.; Tsuchiya, Y.N.; Sawa, S.; Fukuda, H.; von Wirén, N.; Takahashi, H. CLE-CLAVATA1 peptide-receptor signaling module regulates the expansion of plant root systems in a nitrogen-dependent manner. Proc. Natl. Acad. Sci. USA 2014, 111, 2029–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araya, T.; von Wirén, N.; Takahashi, H. CLE peptide signaling and nitrogen interactions in plant root development. Plant Mol. Biol. 2016, 91, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Liao, P.; Li, S.; Cui, X.; Zheng, Y. A comprehensive review of web-based resources of non-coding RNAs for plant science research. Int. J. Biol. Sci. 2018, 14, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell Biol. 2015, 16, 727–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.V.; Chekanova, J.A. Long Noncoding RNAs in Plants. Adv. Exp. Med. Biol. 2017, 1008, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jung, C.; Xu, J.; Wang, H.; Deng, S.; Bernad, L.; Arenas-Huertero, C.; Chua, N.H. Genome-wide analysis uncovers regulation of long intergenic noncoding RNAs in Arabidopsis. Plant Cell 2012, 24, 4333–4345. [Google Scholar] [CrossRef] [Green Version]
- Matsui, A.; Ishida, J.; Morosawa, T.; Mochizuki, Y.; Kaminuma, E.; Endo, T.A.; Okamoto, M.; Nambara, E.; Nakajima, M.; Kawashima, M.; et al. Arabidopsis transcriptome analysis under drought, cold, high-salinity and ABA treatment conditions using a tiling array. Plant Cell Physiol. 2008, 49, 1135–1149. [Google Scholar] [CrossRef] [Green Version]
- Ding, Z.; Tie, W.; Fu, L.; Yan, Y.; Liu, G.; Yan, W.; Li, Y.; Wu, C.; Zhang, J.; Hu, W. Strand-specific RNA-seq based identification and functional prediction of drought-responsive lncRNAs in cassava. BMC Genom. 2019, 20, 214. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, M.; Nishida, S.; Kakei, Y.; Shimada, Y.; Fujiwara, T. Genome-Wide Analysis of Long Intergenic Noncoding RNAs Responding to Low-Nutrient Conditions in Arabidopsis thaliana: Possible Involvement of Trans-Acting siRNA3 in Response to Low Nitrogen. Plant Cell Physiol. 2019, 60, 1961–1973. [Google Scholar] [CrossRef]
- Pang, J.; Zhang, X.; Ma, X.; Zhao, J. Spatio-Temporal Transcriptional Dynamics of Maize Long Non-Coding RNAs Responsive to Drought Stress. Genes 2019, 10, 138. [Google Scholar] [CrossRef] [Green Version]
- Qi, X.; Xie, S.; Liu, Y.; Yi, F.; Yu, J. Genome-wide annotation of genes and noncoding RNAs of foxtail millet in response to simulated drought stress by deep sequencing. Plant Mol. Biol. 2013, 83, 459–473. [Google Scholar] [CrossRef]
- Zhang, W.; Han, Z.; Guo, Q.; Liu, Y.; Zheng, Y.; Wu, F.; Jin, W. Identification of maize long non-coding RNAs responsive to drought stress. PLoS ONE 2014, 9, e98958. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Hu, J.; Gao, C.; Chen, G.; Wang, B.; Lin, C.; Song, L.; Ding, Y.; Zhou, G. Genome-wide analysis of long non-coding RNAs unveils the regulatory roles in the heat tolerance of Chinese cabbage (Brassica rapa ssp.chinensis). Sci. Rep. 2019, 9, 5002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Dai, L.; Ai, J.; Wang, Y.; Ren, F. Identification and functional prediction of cold-related long non-coding RNA (lncRNA) in grapevine. Sci. Rep. 2019, 9, 6638. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Z.; Liu, M.; Zhao, M.G.; Chen, R.; Zhang, W.H. Identification and characterization of long non-coding RNAs involved in osmotic and salt stress in Medicago truncatula using genome-wide high-throughput sequencing. BMC Plant Biol. 2015, 15, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nejat, N.; Mantri, N. Emerging roles of long non-coding RNAs in plant response to biotic and abiotic stresses. Crit. Rev. Biotechnol. 2018, 38, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Castillo-Gonzalez, C.; Yu, B.; Zhang, X. The functions of plant small RNAs in development and in stress responses. Plant J. 2017, 90, 654–670. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Zhang, Y.; Chen, X.; Chen, Y. Plant Noncoding RNAs: Hidden Players in Development and Stress Responses. Annu. Rev. Cell Dev. Biol. 2019, 35, 407–431. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Y.; Chang, K.; Li, S.; Liu, Z.; Qi, S.; Jia, J.; Zhang, M.; Crawford, N.M.; Wang, Y. The long noncoding RNA T5120 regulates nitrate response and assimilation in Arabidopsis. New Phytol. 2019, 224, 117–131. [Google Scholar] [CrossRef]
- Chen, M.; Wang, C.; Bao, H.; Chen, H.; Wang, Y. Genome-wide identification and characterization of novel lncRNAs in Populus under nitrogen deficiency. Mol. Genet. Genom. 2016, 291, 1663–1680. [Google Scholar] [CrossRef]
- Shin, S.Y.; Jeong, J.S.; Lim, J.Y.; Kim, T.; Park, J.H.; Kim, J.K.; Shin, C. Transcriptomic analyses of rice (Oryza sativa) genes and non-coding RNAs under nitrogen starvation using multiple omics technologies. BMC Genom. 2018, 19, 532. [Google Scholar] [CrossRef]
- Li, L.; Eichten, S.R.; Shimizu, R.; Petsch, K.; Yeh, C.T.; Wu, W.; Chettoor, A.M.; Givan, S.A.; Cole, R.A.; Fowler, J.E.; et al. Genome-wide discovery and characterization of maize long non-coding RNAs. Genome Biol. 2014, 15, R40. [Google Scholar] [CrossRef] [Green Version]
- Rymarquis, L.A.; Kastenmayer, J.P.; Hüttenhofer, A.G.; Green, P.J. Diamonds in the rough: mRNA-like non-coding RNAs. Trends Plant Sci. 2008, 13, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Bartel, B.; Bartel, D.P. MicroRNAs: At the Root of Plant Development? Plant Physiol. 2003, 132, 709–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. miRBase: Tools for microRNA genomics. Nucleic Acids Res 2008, 36, D154–D158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Mei, J.; Ren, G. Plant microRNAs: Biogenesis, Homeostasis, and Degradation. Front. Plant Sci. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Han, M.-H.; Fedoroff, N. The RNA-binding proteins HYL1 and SE promote accurate in vitro processing of pri-miRNA by DCL1. Proc. Natl. Acad. Sci. USA 2008, 105, 9970–9975. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Spector, D.L. Identification of Nuclear Dicing Bodies Containing Proteins for MicroRNA Biogenesis in Living Arabidopsis Plants. Curr. Biol. 2007, 17, 818–823. [Google Scholar] [CrossRef] [Green Version]
- Mi, S.; Cai, T.; Hu, Y.; Chen, Y.; Hodges, E.; Ni, F.; Wu, L.; Li, S.; Zhou, H.; Long, C.; et al. Sorting of Small RNAs into Arabidopsis Argonaute Complexes Is Directed by the 5′ Terminal Nucleotide. Cell 2008, 133, 116–127. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Zhang, Q.; Zhou, H.; Ni, F.; Wu, X.; Qi, Y. Rice MicroRNA Effector Complexes and Targets. Plant Cell 2009, 21, 3421–3435. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Xia, R.; Meyers, B.C.; Walbot, V. Evolution, functions, and mysteries of plant ARGONAUTE proteins. Curr. Opin. Plant Biol. 2015, 27, 84–90. [Google Scholar] [CrossRef] [Green Version]
- Sontheimer, E.J.; Carthew, R.W. Silence from within: Endogenous siRNAs and miRNAs. Cell 2005, 122, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Axtell, M.J. Classification and comparison of small RNAs from plants. Annu. Rev. Plant Biol. 2013, 64, 137–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosher, R.A.; Schwach, F.; Studholme, D.; Baulcombe, D.C. PolIVb influences RNA-directed DNA methylation independently of its role in siRNA biogenesis. Proc. Natl. Acad. Sci. USA 2008, 105, 3145–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Henderson, I.R.; Lu, C.; Green, P.J.; Jacobsen, S.E. Role of RNA polymerase IV in plant small RNA metabolism. Proc. Natl. Acad. Sci. USA 2007, 104, 4536–4541. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Johnson, L.M.; Jacobsen, S.E.; Patel, D.J. DNA methylation pathways and their crosstalk with histone methylation. Nat. Rev. Mol. Cell Biol. 2015, 16, 519–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzke, M.A.; Kanno, T.; Matzke, A.J.M. RNA-Directed DNA Methylation: The Evolution of a Complex Epigenetic Pathway in Flowering Plants. Annu. Rev. Plant Biol. 2015, 66, 243–267. [Google Scholar] [CrossRef] [PubMed]
- Zilberman, D.; Cao, X.; Jacobsen, S.E. ARGONAUTE4 Control of Locus-Specific siRNA Accumulation and DNA and Histone Methylation. Science 2003, 299, 716–719. [Google Scholar] [CrossRef] [Green Version]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, M.; Peragine, A.; Park, M.Y.; Poethig, R.S. A pathway for the biogenesis of trans-acting siRNAs in Arabidopsis. Genes Dev. 2005, 19, 2164–2175. [Google Scholar] [CrossRef] [Green Version]
- Axtell, M.J.; Jan, C.; Rajagopalan, R.; Bartel, D.P. A two-hit trigger for siRNA biogenesis in plants. Cell 2006, 127, 565–577. [Google Scholar] [CrossRef] [Green Version]
- Borsani, O.; Zhu, J.; Verslues, P.E.; Sunkar, R.; Zhu, J.-K. Endogenous siRNAs Derived from a Pair of Natural cis-Antisense Transcripts Regulate Salt Tolerance in Arabidopsis. Cell 2005, 123, 1279–1291. [Google Scholar] [CrossRef] [Green Version]
- Qi, Y.; Denli, A.M.; Hannon, G.J. Biochemical Specialization within Arabidopsis RNA Silencing Pathways. Mol. Cell 2005, 19, 421–428. [Google Scholar] [CrossRef]
- Baumberger, N.; Baulcombe, D.C. Arabidopsis ARGONAUTE1 is an RNA Slicer that selectively recruits microRNAs and short interfering RNAs. Proc. Natl. Acad. Sci. USA 2005, 102, 11928–11933. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Iwasaki, S.; Watanabe, T.; Utsumi, M.; Watanabe, Y. The Mechanism Selecting the Guide Strand from Small RNA Duplexes is Different Among Argonaute Proteins. Plant Cell Physiol. 2008, 49, 493–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaucheret, H. Plant ARGONAUTES. Trends Plant Sci. 2008, 13, 350–358. [Google Scholar] [CrossRef]
- Pagliarani, C.; Gambino, G. Small RNA Mobility: Spread of RNA Silencing Effectors and its Effect on Developmental Processes and Stress Adaptation in Plants. Int. J. Mol. Sci. 2019, 20, 4306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, F.; Mendell, J.T. Functional Classification and Experimental Dissection of Long Noncoding RNAs. Cell 2018, 172, 393–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franco-Zorrilla, J.M.; Valli, A.; Todesco, M.; Mateos, I.; Puga, M.I.; Rubio-Somoza, I.; Leyva, A.; Weigel, D.; Garcia, J.A.; Paz-Ares, J. Target mimicry provides a new mechanism for regulation of microRNA activity. Nat. Genet. 2007, 39, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Bardou, F.; Ariel, F.; Simpson, C.G.; Romero-Barrios, N.; Laporte, P.; Balzergue, S.; Brown, J.W.; Crespi, M. Long noncoding RNA modulates alternative splicing regulators in Arabidopsis. Dev. Cell 2014, 30, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Fan, X.; Lin, F.; He, G.; Terzaghi, W.; Zhu, D.; Deng, X.W. Arabidopsis noncoding RNA mediates control of photomorphogenesis by red light. Proc. Natl. Acad. Sci. USA 2014, 111, 10359–10364. [Google Scholar] [CrossRef] [Green Version]
- Swiezewski, S.; Liu, F.; Magusin, A.; Dean, C. Cold-induced silencing by long antisense transcripts of an Arabidopsis Polycomb target. Nature 2009, 462, 799–802. [Google Scholar] [CrossRef]
- Kim, D.H.; Sung, S. Vernalization-Triggered Intragenic Chromatin Loop Formation by Long Noncoding RNAs. Dev. Cell 2017, 40, 302–312.e304. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.B.; Sung, S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 2011, 331, 76–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backman, T.W.; Sullivan, C.M.; Cumbie, J.S.; Miller, Z.A.; Chapman, E.J.; Fahlgren, N.; Givan, S.A.; Carrington, J.C.; Kasschau, K.D. Update of ASRP: The Arabidopsis Small RNA Project database. Nucleic Acids Res 2008, 36, D982–D985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paytuvi Gallart, A.; Hermoso Pulido, A.; Anzar Martinez de Lagran, I.; Sanseverino, W.; Aiese Cigliano, R. GREENC: A Wiki-based database of plant lncRNAs. Nucleic Acids Res 2016, 44, D1161–D1166. [Google Scholar] [CrossRef]
- Di, C.; Yuan, J.; Wu, Y.; Li, J.; Lin, H.; Hu, L.; Zhang, T.; Qi, Y.; Gerstein, M.B.; Guo, Y.; et al. Characterization of stress-responsive lncRNAs in Arabidopsis thaliana by integrating expression, epigenetic and structural features. Plant J. 2014, 80, 848–861. [Google Scholar] [CrossRef]
- Wu, W.; Wu, Y.; Hu, D.; Zhou, Y.; Hu, Y.; Chen, Y.; Chen, M. PncStress: A manually curated database of experimentally validated stress-responsive non-coding RNAs in plants. Database 2020, 2020, baaa001. [Google Scholar] [CrossRef] [Green Version]
- Pant, B.D.; Musialak-Lange, M.; Nuc, P.; May, P.; Buhtz, A.; Kehr, J.; Walther, D.; Scheible, W.R. Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing. Plant Physiol. 2009, 150, 1541–1555. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; He, H.; Yu, D. Identification of nitrogen starvation-responsive microRNAs in Arabidopsis thaliana. PLoS ONE 2012, 7, e48951. [Google Scholar] [CrossRef] [Green Version]
- Nath, M.; Tuteja, N. NPKS uptake, sensing, and signaling and miRNAs in plant nutrient stress. Protoplasma 2016, 253, 767–786. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; Correa, R.; Gomes, I.A.; de Lima, J.C.; Margis, R. NPK macronutrients and microRNA homeostasis. Front. Plant Sci. 2015, 6, 451. [Google Scholar] [CrossRef] [Green Version]
- Gifford, M.L.; Dean, A.; Gutierrez, R.A.; Coruzzi, G.M.; Birnbaum, K.D. Cell-specific nitrogen responses mediate developmental plasticity. Proc. Natl. Acad. Sci. USA 2008, 105, 803–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Mai, Y.X.; Zhang, Y.C.; Luo, Q.; Yang, H.Q. MicroRNA171c-targeted SCL6-II, SCL6-III, and SCL6-IV genes regulate shoot branching in Arabidopsis. Mol. Plant 2010, 3, 794–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.W.; Wang, L.J.; Mao, Y.B.; Cai, W.J.; Xue, H.W.; Chen, X.Y. Control of root cap formation by MicroRNA-targeted auxin response factors in Arabidopsis. Plant Cell 2005, 17, 2204–2216. [Google Scholar] [CrossRef] [Green Version]
- Vidal, E.A.; Araus, V.; Lu, C.; Parry, G.; Green, P.J.; Coruzzi, G.M.; Gutierrez, R.A. Nitrate-responsive miR393/AFB3 regulatory module controls root system architecture in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 4477–4482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, E.A.; Moyano, T.C.; Riveras, E.; Contreras-López, O.; Gutiérrez, R.A. Systems approaches map regulatory networks downstream of the auxin receptor AFB3 in the nitrate response of Arabidopsis thaliana roots. Proc. Natl. Acad. Sci. USA 2013, 110, 12840–12845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Ding, H.; Zhu, J.K.; Zhang, F.; Li, W.X. Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol. 2011, 190, 906–915. [Google Scholar] [CrossRef] [Green Version]
- Kliebenstein, D.J.; Lambrix, V.M.; Reichelt, M.; Gershenzon, J.; Mitchell-Olds, T. Gene duplication in the diversification of secondary metabolism: Tandem 2-oxoglutarate-dependent dioxygenases control glucosinolate biosynthesis in Arabidopsis. Plant Cell 2001, 13, 681–693. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Liang, G.; Li, Y.; Wang, F.; Yu, D. Two young MicroRNAs originating from target duplication mediate nitrogen starvation adaptation via regulation of glucosinolate synthesis in Arabidopsis thaliana. Plant Physiol. 2014, 164, 853–865. [Google Scholar] [CrossRef] [Green Version]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Sung, G.H.; Spatafora, J.W.; Carrington, J.C. Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana. Nat. Genet. 2004, 36, 1282–1290. [Google Scholar] [CrossRef]
- Vidal, E.A.; Tamayo, K.P.; Gutierrez, R.A. Gene networks for nitrogen sensing, signaling, and response in Arabidopsis thaliana. Wiley Interdiscip Rev. Syst. Biol. Med. 2010, 2, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.; Ai, Q.; Yu, D. Uncovering miRNAs involved in crosstalk between nutrient deficiencies in Arabidopsis. Sci. Rep. 2015, 5, 11813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchive, C.; Roudier, F.; Castaings, L.; Bréhaut, V.; Blondet, E.; Colot, V.; Meyer, C.; Krapp, A. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 2013, 4, 1713. [Google Scholar] [CrossRef] [PubMed]
- Ariel, F.; Jegu, T.; Latrasse, D.; Romero-Barrios, N.; Christ, A.; Benhamed, M.; Crespi, M. Noncoding transcription by alternative RNA polymerases dynamically regulates an auxin-driven chromatin loop. Mol. Cell 2014, 55, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, L.; Carles, C.C.; Osmont, K.S.; Fletcher, J.C. A database analysis method identifies an endogenous trans-acting short-interfering RNA that targets the Arabidopsis ARF2, ARF3, and ARF4 genes. Proc. Natl. Acad. Sci. USA 2005, 102, 9703–9708. [Google Scholar] [CrossRef] [Green Version]
- Marin, E.; Jouannet, V.; Herz, A.; Lokerse, A.S.; Weijers, D.; Vaucheret, H.; Nussaume, L.; Crespi, M.D.; Maizel, A. miR390, Arabidopsis TAS3 tasiRNAs, and their AUXIN RESPONSE FACTOR targets define an autoregulatory network quantitatively regulating lateral root growth. Plant Cell 2010, 22, 1104–1117. [Google Scholar] [CrossRef] [Green Version]
- Yoon, E.K.; Yang, J.H.; Lim, J.; Kim, S.H.; Kim, S.K.; Lee, W.S. Auxin regulation of the microRNA390-dependent transacting small interfering RNA pathway in Arabidopsis lateral root development. Nucleic Acids Res. 2010, 38, 1382–1391. [Google Scholar] [CrossRef] [Green Version]
- Kiba, T.; Feria-Bourrellier, A.B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Brehaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H.; et al. The Arabidopsis nitrate transporter NRT2.4 plays a double role in roots and shoots of nitrogen-starved plants. Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef] [Green Version]
- Nishida, S.; Kakei, Y.; Shimada, Y.; Fujiwara, T. Genome-wide analysis of specific alterations in transcript structure and accumulation caused by nutrient deficiencies in Arabidopsis thaliana. Plant J. 2017, 91, 741–753. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Loque, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; von Wiren, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhong, S.; Li, X.; Li, W.; Rothstein, S.J.; Zhang, S.; Bi, Y.; Xie, C. Genome-wide identification of microRNAs in response to low nitrate availability in maize leaves and roots. PLoS ONE 2011, 6, e28009. [Google Scholar] [CrossRef] [Green Version]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, B.; Bartel, D.P. Prediction of plant microRNA targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational Identification of Plant MicroRNAs and Their Targets, Including a Stress-Induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H.; Vazquez, F.; Crété, P.; Bartel, D.P. The action of ARGONAUTE1 in the miRNA pathway and its regulation by the miRNA pathway are crucial for plant development. Genes Dev. 2004, 18, 1187–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaucheret, H.; Mallory, A.C.; Bartel, D.P. AGO1 Homeostasis Entails Coexpression of MIR168 and AGO1 and Preferential Stabilization of miR168 by AGO1. Mol. Cell 2006, 22, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevisan, S.; Nonis, A.; Begheldo, M.; Manoli, A.; Palme, K.; Caporale, G.; Ruperti, B.; Quaggiotti, S. Expression and tissue-specific localization of nitrate-responsive miRNAs in roots of maize seedlings. Plant Cell Environ. 2012, 35, 1137–1155. [Google Scholar] [CrossRef]
- Trevisan, S.; Begheldo, M.; Nonis, A.; Quaggiotti, S. The miRNA-mediated post-transcriptional regulation of maize response to nitrate. Plant Signal. Behav. 2012, 7, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Xu, Z.; Mo, Q.; Zou, C.; Li, W.; Xu, Y.; Xie, C. Combined small RNA and degradome sequencing reveals novel miRNAs and their targets in response to low nitrate availability in maize. Ann. Bot. 2013, 112, 633–642. [Google Scholar] [CrossRef]
- Zhao, M.; Tai, H.; Sun, S.; Zhang, F.; Xu, Y.; Li, W.X. Cloning and characterization of maize miRNAs involved in responses to nitrogen deficiency. PLoS ONE 2012, 7, e29669. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Wang, Z.; Yang, C.; Yang, Z.; Li, H.; Wu, Y. Physiological responses and small RNAs changes in maize under nitrogen deficiency and resupply. Genes Genom. 2019, 41, 1183–1194. [Google Scholar] [CrossRef]
- Lv, Y.; Liang, Z.; Ge, M.; Qi, W.; Zhang, T.; Lin, F.; Peng, Z.; Zhao, H. Genome-wide identification and functional prediction of nitrogen-responsive intergenic and intronic long non-coding RNAs in maize (Zea mays L.). BMC Genom. 2016, 17, 350. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Hu, B.; Wang, W.; Zhang, Z.; Liang, Y.; Gao, X.; Li, P.; Liu, Y.; Zhang, L.; Chu, C. Identification of microRNAs in rice root in response to nitrate and ammonium. J. Genet. Genom. 2016, 43, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Rosikiewicz, W.; Makalowska, I. Biological functions of natural antisense transcripts. Acta Biochim. Pol. 2016, 63, 665–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Bao, H.; Wu, Q.; Wang, Y. Transcriptome-Wide Identification of miRNA Targets under Nitrogen Deficiency in Populus tomentosa Using Degradome Sequencing. Int. J. Mol. Sci. 2015, 16, 13937–13958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Y.; Sun, F.; Hou, J.; Chen, L.; Zhang, Y.; Kang, X.; Wang, Y. Differential profiling analysis of miRNAs reveals a regulatory role in low N stress response of Populus. Funct. Integr. Genom. 2015, 15, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, C.; Hao, Q.; Sha, A.; Zhou, R.; Zhou, X.; Yuan, L. Elucidation of miRNAs-mediated responses to low nitrogen stress by deep sequencing of two soybean genotypes. PLoS ONE 2013, 8, e67423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, J.K.; Buckseth, T.; Zinta, R.; Saraswati, A.; Singh, R.K.; Rawat, S.; Chakrabarti, S.K. Genome-wide identification and characterization of microRNAs by small RNA sequencing for low nitrogen stress in potato. PLoS ONE 2020, 15, e0233076. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.W.; Li, H.; Liu, J.X.; Wang, Y.; Zhuang, J. Integrative transcriptome, proteome, and microRNA analysis reveals the effects of nitrogen sufficiency and deficiency conditions on theanine metabolism in the tea plant (Camellia sinensis). Hortic Res. 2020, 7, 65. [Google Scholar] [CrossRef]
- Grabowska, A.; Smoczynska, A.; Bielewicz, D.; Pacak, A.; Jarmolowski, A.; Szweykowska-Kulinska, Z. Barley microRNAs as metabolic sensors for soil nitrogen availability. Plant Sci. 2020, 299, 110608. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, L.; Lu, W.; Li, X.; Chen, H.; Guo, C.; Xiao, K. Expression pattern analysis of microRNAs in root tissue of wheat (Triticum aestivum L.) under normal nitrogen and low nitrogen conditions. J. Plant Biochem. Biotechnol. 2015, 24, 143–153. [Google Scholar] [CrossRef]
- Hou, G.; Du, C.; Gao, H.; Liu, S.; Sun, W.; Lu, H.; Kang, J.; Xie, Y.; Ma, D.; Wang, C. Identification of microRNAs in developing wheat grain that are potentially involved in regulating grain characteristics and the response to nitrogen levels. BMC Plant Biol. 2020, 20, 87. [Google Scholar] [CrossRef]
- Zuluaga, D.L.; De Paola, D.; Janni, M.; Curci, P.L.; Sonnante, G. Durum wheat miRNAs in response to nitrogen starvation at the grain filling stage. PLoS ONE 2017, 12, e0183253. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, D.L.; Liuzzi, V.; Curci, P.L.; Sonnante, G. MicroRNAs in durum wheat seedlings under chronic and short-term nitrogen stress. Funct. Integr. Genom. 2018, 18, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, C.M.A.; Weretilnyk, E.A.; Golding, G.B. Molecular Traits of Long Non-protein Coding RNAs from Diverse Plant Species Show Little Evidence of Phylogenetic Relationships. G3 Genes Genomes Genet. 2019, 9, 2511–2520. [Google Scholar] [CrossRef] [Green Version]
- Ponjavic, J.; Ponting, C.P.; Lunter, G. Functionality or transcriptional noise? Evidence for selection within long noncoding RNAs. Genome Res. 2007, 17, 556–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diederichs, S. The four dimensions of noncoding RNA conservation. Trends Genet. 2014, 30, 121–123. [Google Scholar] [CrossRef]
- Vandivier, L.E.; Anderson, S.J.; Foley, S.W.; Gregory, B.D. The Conservation and Function of RNA Secondary Structure in Plants. Annu. Rev. Plant Biol. 2016, 67, 463–488. [Google Scholar] [CrossRef] [Green Version]
- Kapusta, A.; Kronenberg, Z.; Lynch, V.J.; Zhuo, X.; Ramsay, L.; Bourque, G.; Yandell, M.; Feschotte, C. Transposable Elements Are Major Contributors to the Origin, Diversification, and Regulation of Vertebrate Long Noncoding RNAs. PLoS Genet. 2013, 9, e1003470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J. Transposon-Derived Non-coding RNAs and Their Function in Plants. Front. Plant Sci. 2018, 9, 600. [Google Scholar] [CrossRef]
- Sigova, A.A.; Mullen, A.C.; Molinie, B.; Gupta, S.; Orlando, D.A.; Guenther, M.G.; Almada, A.E.; Lin, C.; Sharp, P.A.; Giallourakis, C.C.; et al. Divergent transcription of long noncoding RNA/mRNA gene pairs in embryonic stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2876–2881. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Li, Y.; Liu, X.; Zhao, Y.; Li, B.; Ingvarsson, P.K.; Zhang, D. Evolutionary Origins of Pseudogenes and Their Association with Regulatory Sequences in Plants. Plant Cell 2019, 31, 563–578. [Google Scholar] [CrossRef] [Green Version]
- Nozawa, M.; Miura, S.; Nei, M. Origins and evolution of microRNA genes in plant species. Genome Biol. Evol. 2012, 4, 230–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, J.; You, C.; Chen, X. The evolution of microRNAs in plants. Curr. Opin. Plant Biol. 2017, 35, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budak, H.; Kaya, S.B.; Cagirici, H.B. Long Non-coding RNA in Plants in the Era of Reference Sequences. Front. Plant Sci. 2020, 11, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.; Chen, Y.; Cao, Y.; Chen, H.; Wang, J.; Bi, Y.M.; Tian, F.; Yang, F.; Rothstein, S.J.; Zhou, X.; et al. Overexpression of miR169o, an Overlapping MicroRNA in Response to Both Nitrogen Limitation and Bacterial Infection, Promotes Nitrogen Use Efficiency and Susceptibility to Bacterial Blight in Rice. Plant Cell Physiol. 2018, 59, 1234–1247. [Google Scholar] [CrossRef]
- Kanter, D.R.; Bartolini, F.; Kugelberg, S.; Leip, A.; Oenema, O.; Uwizeye, A. Nitrogen pollution policy beyond the farm. Nat. Food 2020, 1, 27–32. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Signaling pathways modulating lateral root (LR) growth and N uptake under severe N deficiency in A. thaliana. Under low-N conditions, expression of TAS3 is suppressed; therefore, production of tasiARFs is reduced [30]. As a result, expression of ARF2/3/4 is derepressed and LR growth is inhibited [98]. Inhibition of LR growth is also controlled by NRT1.1-mediated auxin transport [21] and the CLE–CLAVATA1 (CLV1) peptide–receptor signaling module [22]. On the other hand, TAS3 down-regulates expression of NRT2.4 by inducing cleavage of NRT2.4 mRNA [30]. Considering that NRT2.4 is transcriptionally suppressed by NIGT1.1 [14], TAS3 might act to enhance suppression. pA, poly A tail.
Figure 1.
Signaling pathways modulating lateral root (LR) growth and N uptake under severe N deficiency in A. thaliana. Under low-N conditions, expression of TAS3 is suppressed; therefore, production of tasiARFs is reduced [30]. As a result, expression of ARF2/3/4 is derepressed and LR growth is inhibited [98]. Inhibition of LR growth is also controlled by NRT1.1-mediated auxin transport [21] and the CLE–CLAVATA1 (CLV1) peptide–receptor signaling module [22]. On the other hand, TAS3 down-regulates expression of NRT2.4 by inducing cleavage of NRT2.4 mRNA [30]. Considering that NRT2.4 is transcriptionally suppressed by NIGT1.1 [14], TAS3 might act to enhance suppression. pA, poly A tail.

Figure 2.
Schematic model of lncRNA emergence. A transcription unit of long non-coding RNA (lncRNA) could originate from pre-existing transcription regulatory sequences of transposable elements (left), protein-coding genes (middle), or pseudogenes (right). Transposable elements could be a source of sequences and signals essential for transcription (e.g., transcription start sites) and processing (e.g., splice and polyadenylation sites) [130], whereas protein-coding genes and pseudogenes could provide transcription factor–binding sites that serve as promoters and enhancers [133]. Gray boxes indicate transcribed loci of unknown function. Red vertical bars indicate mutations.
Figure 2.
Schematic model of lncRNA emergence. A transcription unit of long non-coding RNA (lncRNA) could originate from pre-existing transcription regulatory sequences of transposable elements (left), protein-coding genes (middle), or pseudogenes (right). Transposable elements could be a source of sequences and signals essential for transcription (e.g., transcription start sites) and processing (e.g., splice and polyadenylation sites) [130], whereas protein-coding genes and pseudogenes could provide transcription factor–binding sites that serve as promoters and enhancers [133]. Gray boxes indicate transcribed loci of unknown function. Red vertical bars indicate mutations.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fukuda, M.; Fujiwara, T.; Nishida, S. Roles of Non-Coding RNAs in Response to Nitrogen Availability in Plants. Int. J. Mol. Sci. 2020, 21, 8508. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228508
AMA Style
Fukuda M, Fujiwara T, Nishida S. Roles of Non-Coding RNAs in Response to Nitrogen Availability in Plants. International Journal of Molecular Sciences. 2020; 21(22):8508. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228508
Chicago/Turabian StyleFukuda, Makiha, Toru Fujiwara, and Sho Nishida. 2020. "Roles of Non-Coding RNAs in Response to Nitrogen Availability in Plants" International Journal of Molecular Sciences 21, no. 22: 8508. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228508
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.