
Effects of Subchronic Administrations of Vortioxetine, Lurasidone, and Escitalopram on Thalamocortical Glutamatergic Transmission Associated with Serotonin 5-HT7 Receptor


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
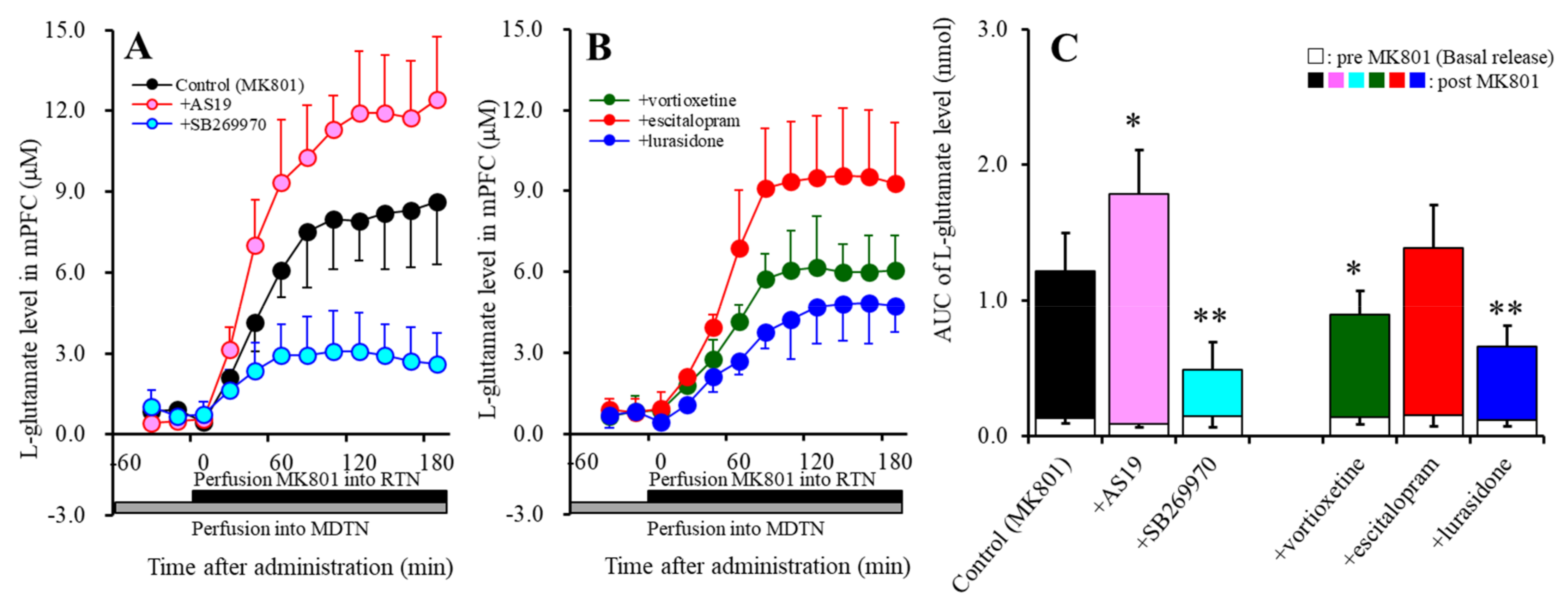
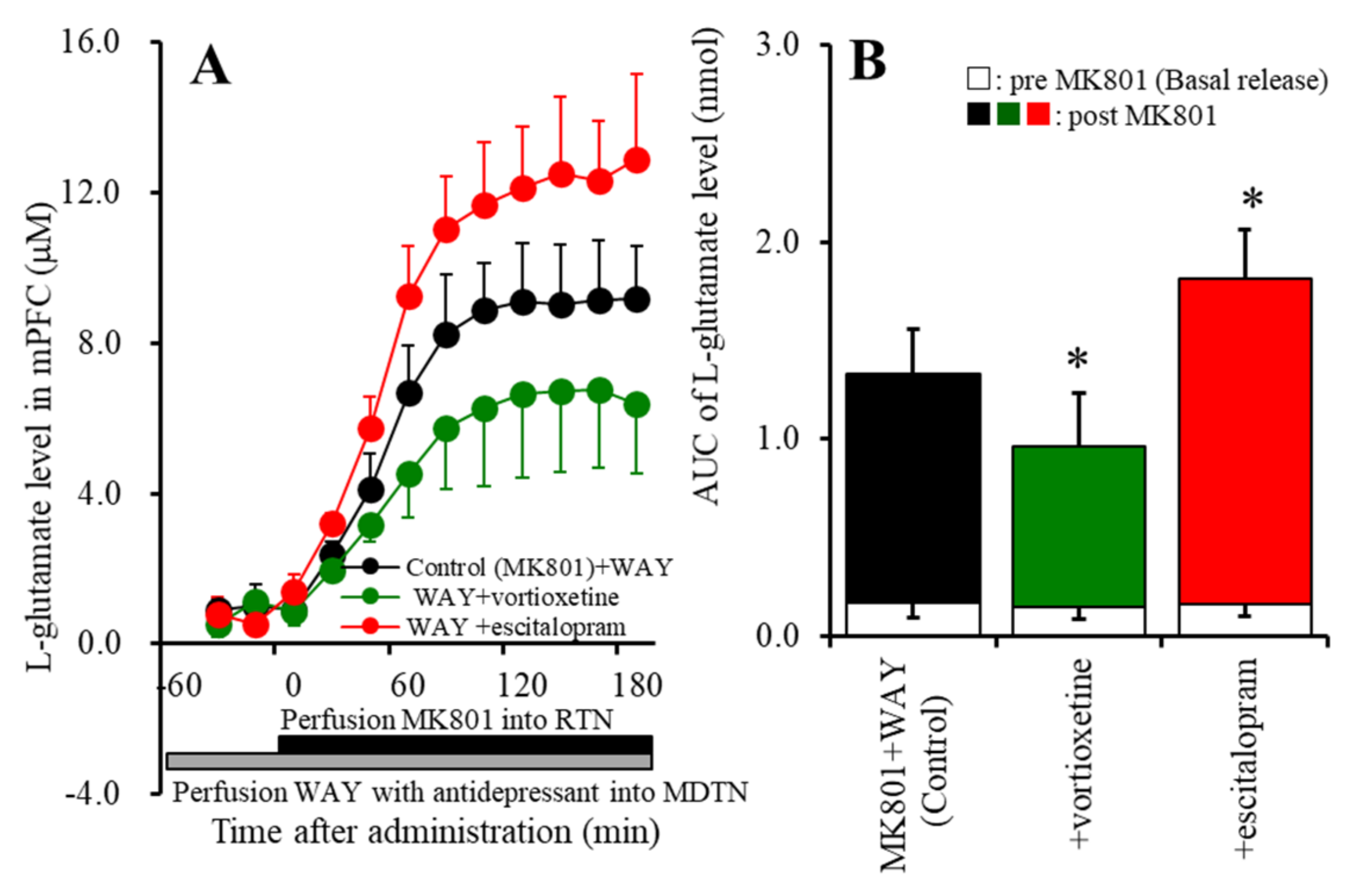
2.1. Effects of Perfusion with a 5-HT7R Agonist, Antagonist, Vortioxetine, Escitalopram, and Lurasidone into the MDTN on MK801-Induced L-Glutamate Release in the mPFC (Study 1)
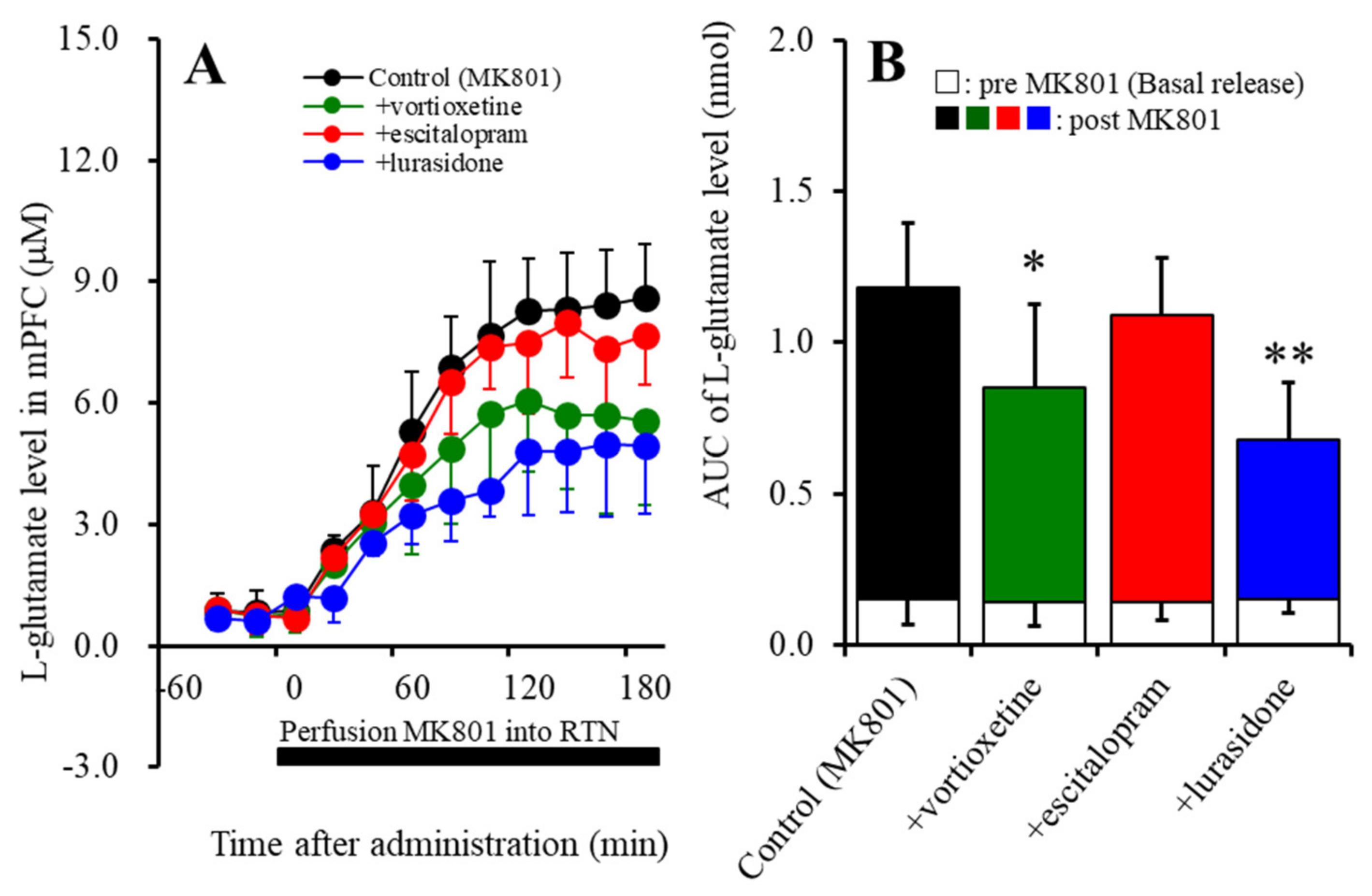
2.2. Effects of the Subchronic, Systemic Administration of Effective Doses of Vortioxetine, Escitalopram, and Lurasidone on MK801-Induced L-Glutamate Release in the mPFC (Study 2)
2.2.1. Effects of the Subchronic Administration of Vortioxetine, Escitalopram, and Lurasidone on MK801-Induced L-Glutamate Release in the mPFC
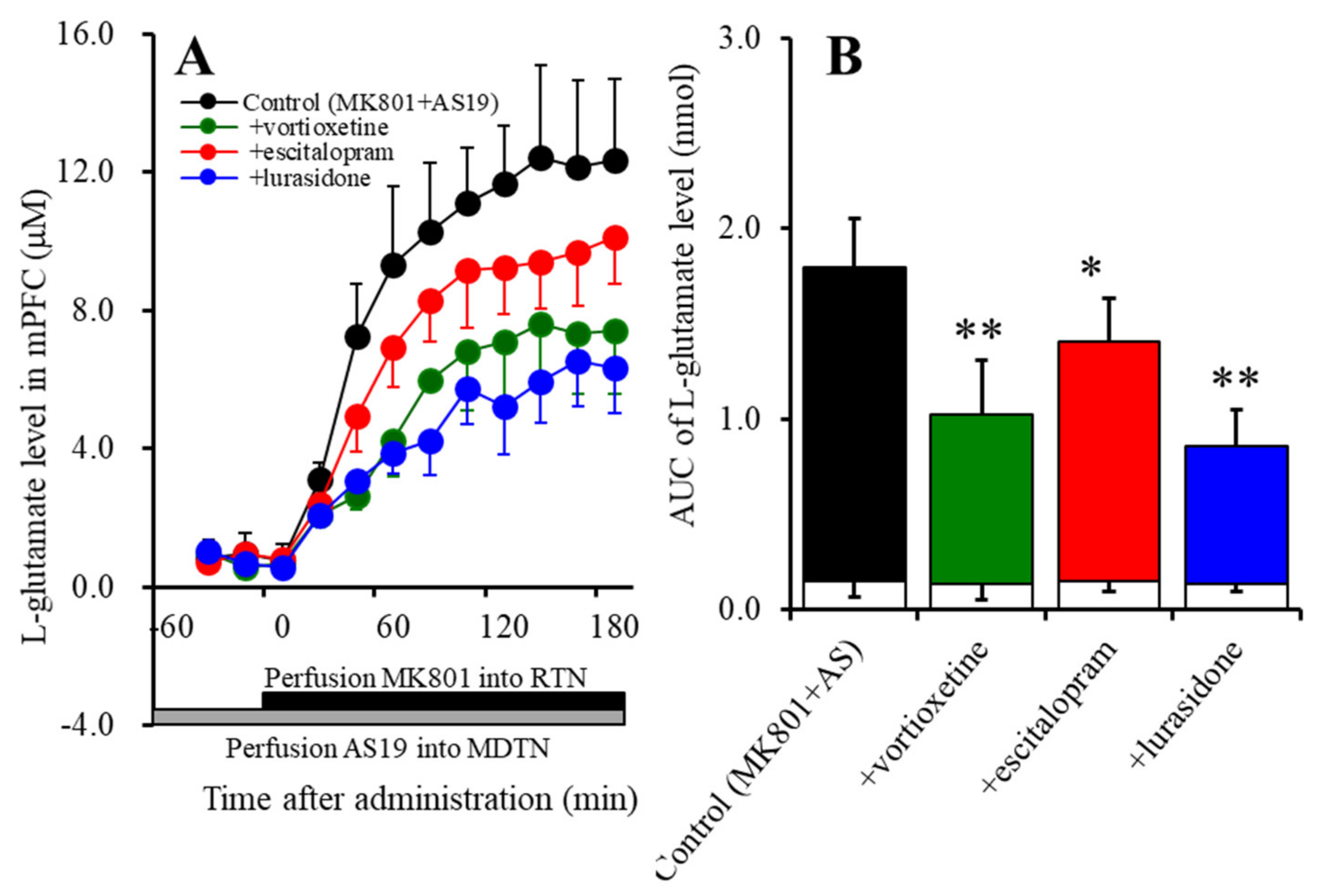
2.2.2. Effects of Subchronic, Systemic Administration of Effective Doses of Vortioxetine, Escitalopram, and Lurasidone on 5-HT7R Associated with MK801-Induced L-Glutamate Release in the mPFC
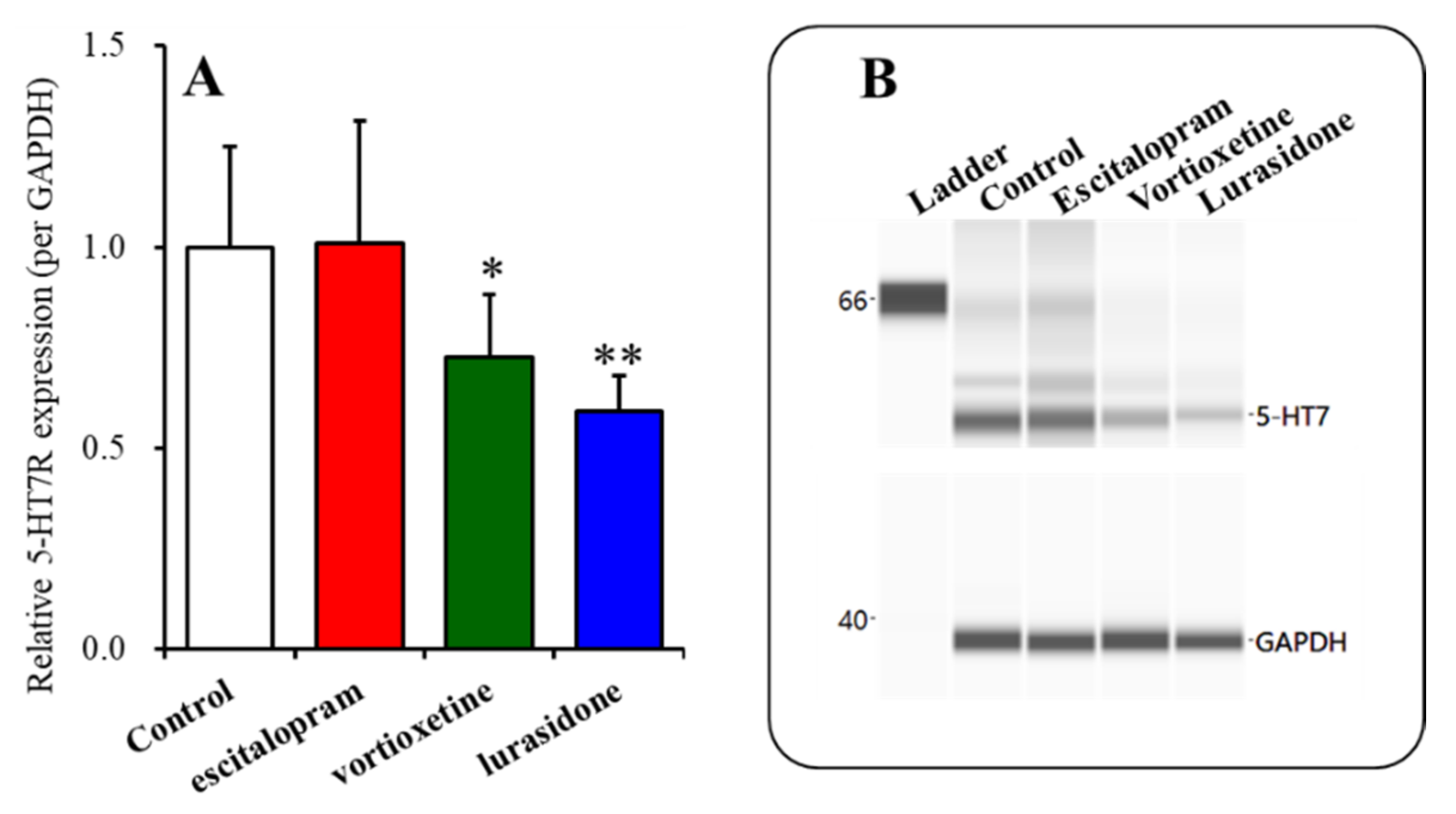
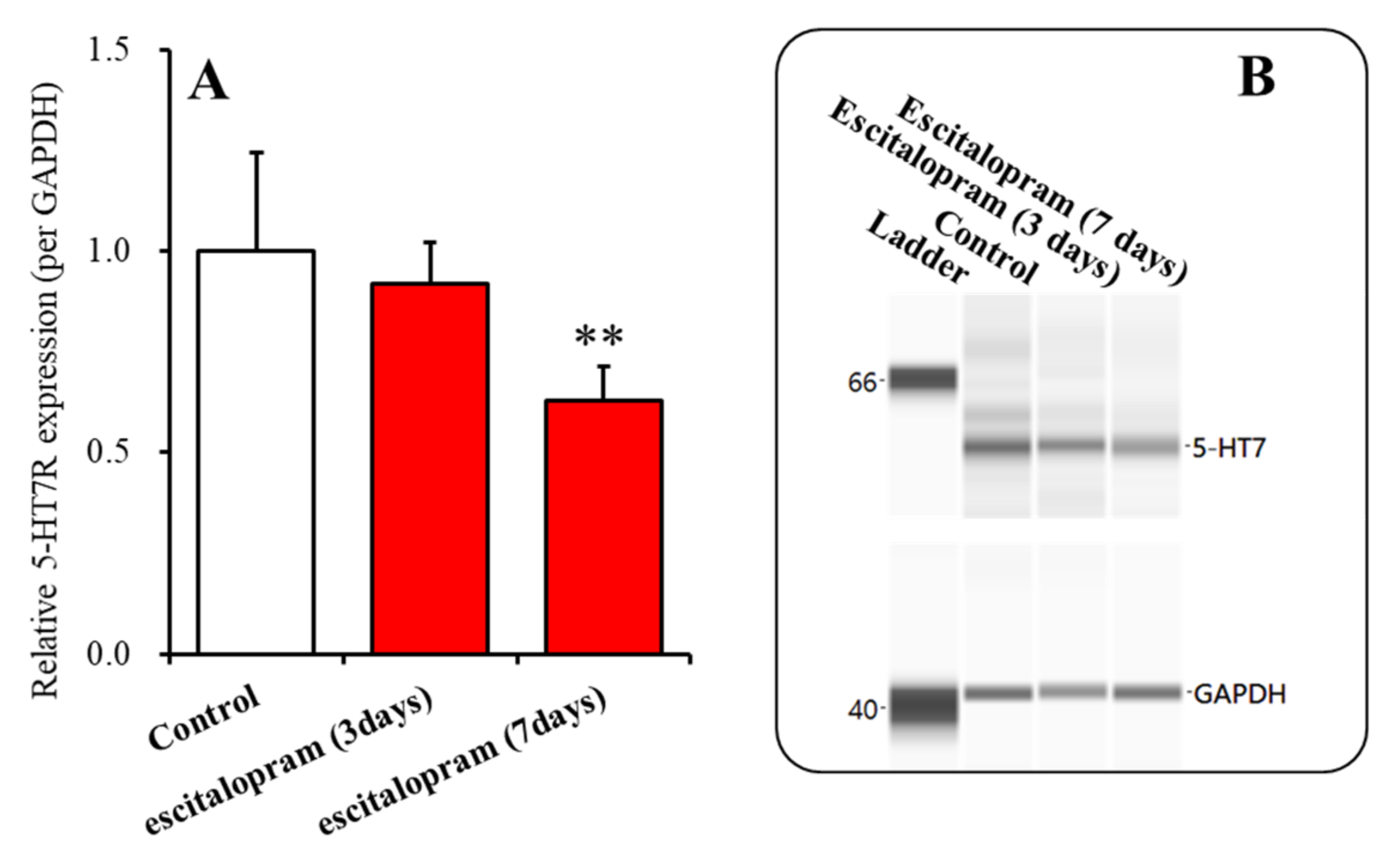
2.3. Time-Dependent Effects of Subchronic Administration of Effective Doses of Vortioxetine, Escitalopram, and Lurasidone on Expression of 5-HT7R in the Thalamic Plasma Membrane Fraction (Study 3)
3. Discussion
3.1. Effects of Acute Local Administration of Vortioxetine, Escitalopram and Lurasidone on Thalamocortical Glutamatergic Transmission
3.2. Effects of Subchronic, Systemic Administration of Vortioxetine, Escitalopram, and Lurasidone on Thalamocortical Glutamatergic Transmission Associated with 5-HT7R
3.3. Clinical Implications of Vortioxetine, Escitalopram, and Lurasidone Associated with 5-HT7R
4. Materials and Methods
4.1. Chemical Agents
4.2. Microdialysis System
4.3. Determination of Extracellular L-Glutamate Level
4.4. Capillary Immunoblotting Analysis
4.5. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Coppen, A. The biochemistry of affective disorders. Br. J. Psychiatry 1967, 113, 1237–1264. [Google Scholar] [CrossRef] [PubMed]
- Kraus, C.; Castren, E.; Kasper, S.; Lanzenberger, R. Serotonin and neuroplasticity—Links between molecular, functional and structural pathophysiology in depression. Neurosci. Biobehav. Rev. 2017, 77, 317–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, M.; Oka, T.; Nakamoto, M.; Fukuyama, K.; Shiroyama, T. Astroglial Connexin43 as a Potential Target for a Mood Stabiliser. Int. J. Mol. Sci. 2020, 22, 339. [Google Scholar] [CrossRef] [PubMed]
- Frazer, A.; Benmansour, S. Delayed pharmacological effects of antidepressants. Mol. Psychiatry 2002, 7, S23–S28. [Google Scholar] [CrossRef]
- Okada, M.; Kawata, Y.; Murakami, T.; Wada, K.; Mizuno, K.; Kondo, T.; Kaneko, S. Differential effects of adenosine receptor subtypes on release and reuptake of hippocampal serotonin. Eur. J. Neurosci. 1999, 11, 1–9. [Google Scholar] [CrossRef]
- Okada, M.; Okubo, R.; Fukuyama, K. Vortioxetine Subchronically Activates Serotonergic Transmission via Desensitization of Serotonin 5-HT1A Receptor with 5-HT3 Receptor Inhibition in Rats. Int. J. Mol. Sci. 2019, 20, 6235. [Google Scholar] [CrossRef] [Green Version]
- Okada, M.; Kawata, Y.; Kiryu, K.; Mizuno, K.; Wada, K.; Tasaki, H.; Kaneko, S. Effects of adenosine receptor subtypes on hippocampal extracellular serotonin level and serotonin reuptake activity. J. Neurochem. 1997, 69, 2581–2588. [Google Scholar]
- Fukuyama, K.; Tanahashi, S.; Hamaguchi, T.; Nakagawa, M.; Shiroyama, T.; Motomura, E.; Okada, M. Differential mechanisms underlie the regulation of serotonergic transmission in the dorsal and median raphe nuclei by mirtazapine: A dual probe microdialysis study. Psychopharmacology 2013, 229, 617–626. [Google Scholar]
- Tanahashi, S.; Ueda, Y.; Nakajima, A.; Yamamura, S.; Nagase, H.; Okada, M. Novel delta1-receptor agonist KNT-127 increases the release of dopamine and L-glutamate in the striatum, nucleus accumbens and median pre-frontal cortex. Neuropharmacology 2012, 62, 2057–2067. [Google Scholar] [CrossRef]
- Yamamura, S.; Abe, M.; Nakagawa, M.; Ochi, S.; Ueno, S.; Okada, M. Different actions for acute and chronic administration of mirtazapine on serotonergic transmission associated with raphe nuclei and their innervation cortical regions. Neuropharmacology 2011, 60, 550–560. [Google Scholar] [CrossRef]
- Bortolozzi, A.; Castane, A.; Semakova, J.; Santana, N.; Alvarado, G.; Cortes, R.; Ferres-Coy, A.; Fernandez, G.; Carmona, M.C.; Toth, M.; et al. Selective siRNA-mediated suppression of 5-HT1A autoreceptors evokes strong anti-depressant-like effects. Mol. Psychiatry 2012, 17, 612–623. [Google Scholar] [CrossRef] [Green Version]
- Popa, D.; Cerdan, J.; Reperant, C.; Guiard, B.P.; Guilloux, J.P.; David, D.J.; Gardier, A.M. A longitudinal study of 5-HT outflow during chronic fluoxetine treatment using a new technique of chronic microdialysis in a highly emotional mouse strain. Eur. J. Pharmacol. 2010, 628, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Gaynes, B.N.; Warden, D.; Trivedi, M.H.; Wisniewski, S.R.; Fava, M.; Rush, A.J. What did STAR*D teach us? Results from a large-scale, practical, clinical trial for patients with depression. Psychiatry Serv. 2009, 60, 1439–1445. [Google Scholar] [CrossRef]
- Okada, M.; Kawano, Y.; Fukuyama, K.; Motomura, E.; Shiroyama, T. Candidate Strategies for Development of a Rapid-Acting Antidepressant Class That Does Not Result in Neuropsychiatric Adverse Effects: Prevention of Ketamine-Induced Neuropsychiatric Adverse Reactions. Int. J. Mol. Sci. 2020, 21, 7951. [Google Scholar]
- Bonaventure, P.; Dugovic, C.; Kramer, M.; De Boer, P.; Singh, J.; Wilson, S.; Bertelsen, K.; Di, J.; Shelton, J.; Aluisio, L.; et al. Translational evaluation of JNJ-18038683, a 5-hydroxytryptamine type 7 receptor antagonist, on rapid eye movement sleep and in major depressive disorder. J. Pharmacol. Exp. Ther. 2012, 342, 429–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesolowska, A.; Nikiforuk, A.; Stachowicz, K.; Tatarczynska, E. Effect of the selective 5-HT7 receptor antagonist SB 269970 in animal models of anxiety and depression. Neuropharmacology 2006, 51, 578–586. [Google Scholar] [CrossRef]
- Bonaventure, P.; Kelly, L.; Aluisio, L.; Shelton, J.; Lord, B.; Galici, R.; Miller, K.; Atack, J.; Lovenberg, T.W.; Dugovic, C. Selective blockade of 5-hydroxytryptamine (5-HT)7 receptors enhances 5-HT transmission, antidepressant-like behavior, and rapid eye movement sleep suppression induced by citalopram in rodents. J. Pharmacol. Exp. Ther. 2007, 321, 690–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mnie-Filali, O.; Faure, C.; Lambas-Senas, L.; El Mansari, M.; Belblidia, H.; Gondard, E.; Etievant, A.; Scarna, H.; Didier, A.; Berod, A.; et al. Pharmacological blockade of 5-HT7 receptors as a putative fast acting antidepressant strategy. Neuropsychopharmacology 2011, 36, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Sajatovic, M.; Forester, B.P.; Tsai, J.; Kroger, H.; Pikalov, A.; Cucchiaro, J.; Loebel, A. Efficacy of Lurasidone in Adults Aged 55 Years and Older With Bipolar Depression: Post Hoc Analysis of 2 Double-Blind, Placebo-Controlled Studies. J. Clin. Psychiatry 2016, 77, e1324–e1331. [Google Scholar] [CrossRef]
- Suppes, T.; Silva, R.; Cucchiaro, J.; Mao, Y.; Targum, S.; Streicher, C.; Pikalov, A.; Loebel, A. Lurasidone for the Treatment of Major Depressive Disorder With Mixed Features: A Randomized, Double-Blind, Placebo-Controlled Study. Am. J. Psychiatry 2016, 173, 400–407. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.; Thase, M.E.; Mao, Y.; Ng-Mak, D.; Pikalov, A.; Loebel, A. Lurasidone for major depressive disorder with mixed features and anxiety: A post-hoc analysis of a randomized, placebo-controlled study. CNS Spectr. 2017, 22, 236–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostacher, M.; Ng-Mak, D.; Patel, P.; Ntais, D.; Schlueter, M.; Loebel, A. Lurasidone compared to other atypical antipsychotic monotherapies for bipolar depression: A systematic review and network meta-analysis. World J. Biol. Psychiatry 2018, 19, 586–601. [Google Scholar] [CrossRef] [PubMed]
- Vieta, E.; Florea, I.; Schmidt, S.N.; Areberg, J.; Ettrup, A. Intravenous vortioxetine to accelerate onset of effect in major depressive disorder: A 2-week, randomized, double-blind, placebo-controlled study. Int. Clin. Psychopharmacol. 2019, 34, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, L.I.; Christmas, D.M.; Hood, S.D.; Potokar, J.P.; Robertson, A.; Isaac, A.; Srivastava, S.; Nutt, D.J.; Davies, S.J. Antidepressant-induced jitteriness/anxiety syndrome: Systematic review. Br. J. Psychiatry 2009, 194, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naslund, J.; Hieronymus, F.; Emilsson, J.F.; Lisinski, A.; Nilsson, S.; Eriksson, E. Incidence of early anxiety aggravation in trials of selective serotonin reuptake inhibitors in depression. Acta Psychiatry Scand. 2017, 136, 343–351. [Google Scholar]
- Subeesh, V.; Singh, H.; Maheswari, E.; Beulah, E. Novel adverse events of vortioxetine: A disproportionality analysis in USFDA adverse event reporting system database. Asian J. Psychiatry 2017, 30, 152–156. [Google Scholar] [CrossRef]
- Kelliny, M.; Croarkin, P.E.; Moore, K.M.; Bobo, W.V. Profile of vortioxetine in the treatment of major depressive disorder: An overview of the primary and secondary literature. Ther. Clin. Risk Manag. 2015, 11, 1193–1212. [Google Scholar]
- Zmudzka, E.; Salaciak, K.; Sapa, J.; Pytka, K. Serotonin receptors in depression and anxiety: Insights from animal studies. Life Sci. 2018, 210, 106–124. [Google Scholar] [CrossRef]
- Gupta, D.; Prabhakar, V.; Radhakrishnan, M. 5HT3 receptors: Target for new antidepressant drugs. Neurosci. Biobehav. Rev. 2016, 64, 311–325. [Google Scholar] [CrossRef]
- Mestre, T.A.; Zurowski, M.; Fox, S.H. 5-Hydroxytryptamine 2A receptor antagonists as potential treatment for psychiatric disorders. Expert Opin. Investig. Drugs 2013, 22, 411–421. [Google Scholar] [CrossRef]
- Olivier, B.; van Wijngaarden, I.; Soudijn, W. 5-HT(3) receptor antagonists and anxiety; a preclinical and clinical review. Eur. Neuropsychopharmacol. 2000, 10, 77–95. [Google Scholar] [CrossRef]
- Hothersall, J.D.; Alexander, A.; Samson, A.J.; Moffat, C.; Bollan, K.A.; Connolly, C.N. 5-Hydroxytryptamine (5-HT) cellular sequestration during chronic exposure delays 5-HT3 receptor resensitization due to its subsequent release. J. Biol. Chem. 2014, 289, 32020–32029. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, J.; Gleason, S.D.; Falcone, J.; Svensson, K.; Balcer, O.M.; Li, X.; Witkin, J.M. Effects of 5-HT7 receptor antagonists on behaviors of mice that detect drugs used in the treatment of anxiety, depression, or schizophrenia. Behav. Brain Res. 2019, 359, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Balcer, O.M.; Seager, M.A.; Gleason, S.D.; Li, X.; Rasmussen, K.; Maxwell, J.K.; Nomikos, G.; Degroot, A.; Witkin, J.M. Evaluation of 5-HT7 receptor antagonism for the treatment of anxiety, depression, and schizophrenia through the use of receptor-deficient mice. Behav. Brain Res. 2019, 360, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Fukuyama, K.; Ueda, Y. Lurasidone inhibits NMDA/glutamate antagonist-induced functional abnormality of thalamocortical glutamatergic transmission via 5-HT7 receptor blockade. Br. J. Pharmacol. 2019, 176, 4002–4018. [Google Scholar] [CrossRef]
- Okada, M.; Fukuyama, K.; Okubo, R.; Shiroyama, T.; Ueda, Y. Lurasidone Sub-Chronically Activates Serotonergic Transmission via Desensitization of 5-HT1A and 5-HT7 Receptors in Dorsal Raphe Nucleus. Pharmaceuticals 2019, 12, 149. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, T.; Horisawa, T.; Tokuda, K.; Ishiyama, T.; Ogasa, M.; Tagashira, R.; Matsumoto, K.; Nishikawa, H.; Ueda, Y.; Toma, S.; et al. Pharmacological profile of lurasidone, a novel antipsychotic agent with potent 5-hydroxytryptamine 7 (5-HT7) and 5-HT1A receptor activity. J. Pharmacol. Exp. Ther. 2010, 334, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Andressen, K.W.; Manfra, O.; Brevik, C.H.; Ulsund, A.H.; Vanhoenacker, P.; Levy, F.O.; Krobert, K.A. The atypical antipsychotics clozapine and olanzapine promote down-regulation and display functional selectivity at human 5-HT7 receptors. Br. J. Pharmacol. 2015, 172, 3846–3860. [Google Scholar]
- Bang-Andersen, B.; Ruhland, T.; Jorgensen, M.; Smith, G.; Frederiksen, K.; Jensen, K.G.; Zhong, H.; Nielsen, S.M.; Hogg, S.; Mork, A.; et al. Discovery of 1-[2-(2,4-dimethylphenylsulfanyl)phenyl]piperazine (Lu AA21004): A novel multimodal compound for the treatment of major depressive disorder. J. Med. Chem. 2011, 54, 3206–3221. [Google Scholar] [CrossRef]
- Mork, A.; Montezinho, L.P.; Miller, S.; Trippodi-Murphy, C.; Plath, N.; Li, Y.; Gulinello, M.; Sanchez, C. Vortioxetine (Lu AA21004), a novel multimodal antidepressant, enhances memory in rats. Pharmacol. Biochem. Behav. 2013, 105, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Takekita, Y.; Fabbri, C.; Kato, M.; Koshikawa, Y.; Tajika, A.; Kinoshita, T.; Serretti, A. HTR1A Polymorphisms and Clinical Efficacy of Antipsychotic Drug Treatment in Schizophrenia: A Meta-Analysis. Int. J. Neuropsychopharmacol. 2016, 19, pyv125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takekita, Y.; Fabbri, C.; Kato, M.; Nonen, S.; Sakai, S.; Sunada, N.; Koshikawa, Y.; Wakeno, M.; Okugawa, G.; Kinoshita, T.; et al. Serotonin 7 Receptor Variants Are Not Associated with Response to Second-Generation Antipsychotics in Japanese Schizophrenia Patients. Neuropsychobiology 2015, 72, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, A.; Li, J.; Meltzer, H.Y. A functional HTR1A polymorphism, rs6295, predicts short-term response to lurasidone: Confirmation with meta-analysis of other antipsychotic drugs. Pharm. J. 2020, 20, 260–270. [Google Scholar]
- Wei, Y.B.; McCarthy, M.; Ren, H.; Carrillo-Roa, T.; Shekhtman, T.; DeModena, A.; Liu, J.J.; Leckband, S.G.; Mors, O.; Rietschel, M.; et al. A functional variant in the serotonin receptor 7 gene (HTR7), rs7905446, is associated with good response to SSRIs in bipolar and unipolar depression. Mol. Psychiatry 2020, 25, 1312–1322. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Fukuyama, K. Interaction between Mesocortical and Mesothalamic Catecholaminergic Transmissions Associated with NMDA Receptor in the Locus Coeruleus. Biomolecules 2020, 10, 990. [Google Scholar] [CrossRef] [PubMed]
- L’Estrade, E.T.; Erlandsson, M.; Edgar, F.G.; Ohlsson, T.; Knudsen, G.M.; Herth, M.M. Towards selective CNS PET imaging of the 5-HT7 receptor system: Past, present and future. Neuropharmacology 2020, 172, 107830. [Google Scholar] [CrossRef]
- Gellynck, E.; Heyninck, K.; Andressen, K.W.; Haegeman, G.; Levy, F.O.; Vanhoenacker, P.; Van Craenenbroeck, K. The serotonin 5-HT 7 receptors: Two decades of research. Exp. Brain Res. 2013, 230, 555–568. [Google Scholar] [CrossRef]
- Matthys, A.; Haegeman, G.; Van Craenenbroeck, K.; Vanhoenacker, P. Role of the 5-HT7 receptor in the central nervous system: From current status to future perspectives. Mol. Neurobiol. 2011, 43, 228–253. [Google Scholar]
- Costa, L.; Spatuzza, M.; D’Antoni, S.; Bonaccorso, C.M.; Trovato, C.; Musumeci, S.A.; Leopoldo, M.; Lacivita, E.; Catania, M.V.; Ciranna, L. Activation of 5-HT7 serotonin receptors reverses metabotropic glutamate receptor-mediated synaptic plasticity in wild-type and Fmr1 knockout mice, a model of Fragile X syndrome. Biol. Psychiatry 2012, 72, 924–933. [Google Scholar] [CrossRef]
- Zareifopoulos, N.; Papatheodoropoulos, C. Effects of 5-HT-7 receptor ligands on memory and cognition. Neurobiol. Learn Mem. 2016, 136, 204–209. [Google Scholar] [CrossRef]
- Lippiello, P.; Hoxha, E.; Speranza, L.; Volpicelli, F.; Ferraro, A.; Leopoldo, M.; Lacivita, E.; Perrone-Capano, C.; Tempia, F.; Miniaci, M.C. The 5-HT7 receptor triggers cerebellar long-term synaptic depression via PKC-MAPK. Neuropharmacology 2016, 101, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Haddjeri, N.; Sanchez, C. Escitalopram, an antidepressant with an allosteric effect at the serotonin transporter—A review of current understanding of its mechanism of action. Psychopharmacology 2012, 219, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Fukuyama, K.; Shiroyama, T.; Murata, M. A Working Hypothesis Regarding Identical Pathomechanisms between Clinical Efficacy and Adverse Reaction of Clozapine via the Activation of Connexin43. Int. J. Mol. Sci. 2020, 21, 7019. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Fukuyama, K.; Nakano, T.; Ueda, Y. Pharmacological Discrimination of Effects of MK801 on Thalamocortical, Mesothalamic, and Mesocortical Transmissions. Biomolecules 2019, 9, 746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, M.; Fukuyama, K.; Kawano, Y.; Shiroyama, T.; Ueda, Y. Memantine protects thalamocortical hyper-glutamatergic transmission induced by NMDA receptor antagonism via activation of system xc−. Pharmacol. Res. Perspect. 2019, 7, e00457. [Google Scholar] [CrossRef] [Green Version]
- Fukuyama, K.; Kato, R.; Murata, M.; Shiroyama, T.; Okada, M. Clozapine Normalizes a Glutamatergic Transmission Abnormality Induced by an Impaired NMDA Receptor in the Thalamocortical Pathway via the Activation of a Group III Metabotropic Glutamate Receptor. Biomolecules 2019, 9, 234. [Google Scholar] [CrossRef] [Green Version]
- Alexander, S.P.H.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Mathie, A.; Peters, J.A.; Veale, E.L.; Armstrong, J.F.; Faccenda, E.; Harding, S.D.; et al. THE CONCISE GUIDE TO PHARMACOLOGY 2019/20: G protein-coupled receptors. Br. J. Pharmacol. 2019, 176 (Suppl. 1), S21–S141. [Google Scholar] [CrossRef] [Green Version]
- Carter, F.; Chapman, C.A. Serotonin 5-HT1A Receptor-Mediated Reduction of Excitatory Synaptic Transmission in Layers II/III of the Parasubiculum. Neuroscience 2019, 406, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Dale, E.; Grunnet, M.; Pehrson, A.L.; Frederiksen, K.; Larsen, P.H.; Nielsen, J.; Stensbol, T.B.; Ebert, B.; Yin, H.; Lu, D.; et al. The multimodal antidepressant vortioxetine may facilitate pyramidal cell firing by inhibition of 5-HT3 receptor expressing interneurons: An in vitro study in rat hippocampus slices. Brain Res. 2018, 1689, 1–11. [Google Scholar] [CrossRef]
- Tanahashi, S.; Yamamura, S.; Nakagawa, M.; Motomura, E.; Okada, M. Dopamine D2 and serotonin 5-HT1A receptors mediate the actions of aripiprazole in mesocortical and mesoaccumbens transmission. Neuropharmacology 2012, 62, 765–774. [Google Scholar] [CrossRef]
- Koo, T.S.; Kim, S.J.; Lee, J.; Ha, D.J.; Baek, M.; Moon, H. Quantification of lurasidone, an atypical antipsychotic drug, in rat plasma with high-performance liquid chromatography with tandem mass spectrometry. Biomed. Chromatogr. 2011, 25, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Pastor, B.; Ortega, J.E.; Grandoso, L.; Castro, E.; Ugedo, L.; Pazos, A.; Meana, J.J. Chronic citalopram administration desensitizes prefrontal cortex but not somatodendritic alpha2-adrenoceptors in rat brain. Neuropharmacology 2017, 114, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.; Asin, K.E.; Artigas, F. Vortioxetine, a novel antidepressant with multimodal activity: Review of preclinical and clinical data. Pharmacol Ther 2015, 145, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Krobert, K.A.; Levy, F.O. The human 5-HT7 serotonin receptor splice variants: Constitutive activity and inverse agonist effects. Br. J. Pharmacol. 2002, 135, 1563–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glowinski, J. Regional studies of catecholamines in the rat brain. J. Neurochem. 1966, 13, 655–669. [Google Scholar] [PubMed]
- Koyama, Y.; Kondo, M.; Shimada, S. Building a 5-HT3A Receptor Expression Map in the Mouse Brain. Sci. Rep. 2017, 7, 42884. [Google Scholar] [CrossRef] [PubMed]
- Sleight, A.J.; Carolo, C.; Petit, N.; Zwingelstein, C.; Bourson, A. Identification of 5-hydroxytryptamine7 receptor binding sites in rat hypothalamus: Sensitivity to chronic antidepressant treatment. Mol. Pharmacol. 1995, 47, 99–103. [Google Scholar]
- Mork, A.; Pehrson, A.; Brennum, L.T.; Nielsen, S.M.; Zhong, H.; Lassen, A.B.; Miller, S.; Westrich, L.; Boyle, N.J.; Sanchez, C.; et al. Pharmacological effects of Lu AA21004: A novel multimodal compound for the treatment of major depressive disorder. J. Pharmacol. Exp. Ther. 2012, 340, 666–675. [Google Scholar] [CrossRef] [Green Version]
- Andreetta, F.; Carboni, L.; Grafton, G.; Jeggo, R.; Whyment, A.D.; van den Top, M.; Hoyer, D.; Spanswick, D.; Barnes, N.M. Hippocampal 5-HT7 receptors signal phosphorylation of the GluA1 subunit to facilitate AMPA receptor mediated-neurotransmission in vitro and in vivo. Br. J. Pharmacol. 2016, 173, 1438–1451. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, J.; Alonso, J.M.; Andres, J.I.; Cid, J.M.; Diaz, A.; Iturrino, L.; Gil, P.; Megens, A.; Sipido, V.K.; Trabanco, A.A. Discovery of new tetracyclic tetrahydrofuran derivatives as potential broad-spectrum psychotropic agents. J. Med. Chem. 2005, 48, 1709–1712. [Google Scholar] [CrossRef]
- Roth, B.L.; Craigo, S.C.; Choudhary, M.S.; Uluer, A.; Monsma, F.J., Jr.; Shen, Y.; Meltzer, H.Y.; Sibley, D.R. Binding of typical and atypical antipsychotic agents to 5-hydroxytryptamine-6 and 5-hydroxytryptamine-7 receptors. J. Pharmacol. Exp. Ther. 1994, 268, 1403–1410. [Google Scholar] [PubMed]
- Renner, U.; Zeug, A.; Woehler, A.; Niebert, M.; Dityatev, A.; Dityateva, G.; Gorinski, N.; Guseva, D.; Abdel-Galil, D.; Frohlich, M.; et al. Heterodimerization of serotonin receptors 5-HT1A and 5-HT7 differentially regulates receptor signalling and trafficking. J. Cell Sci. 2012, 125, 2486–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pergola, G.; Danet, L.; Pitel, A.L.; Carlesimo, G.A.; Segobin, S.; Pariente, J.; Suchan, B.; Mitchell, A.S.; Barbeau, E.J. The Regulatory Role of the Human Mediodorsal Thalamus. Trends Cogn. Sci. 2018, 22, 1011–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, D.A.; Wang, Z. Serotonin and noradrenaline excite GABAergic neurones of the guinea-pig and cat nucleus reticularis thalami. J. Physiol. 1991, 442, 235–255. [Google Scholar] [CrossRef] [PubMed]
- Russchen, F.; Amaral, D.G.; Price, J. The afferent input to the magnocellular division of the mediodorsal thalamic nucleus in the monkey, Macaca fascicularis. J. Comp. Neurol. 1987, 256, 175–210. [Google Scholar] [CrossRef] [PubMed]
- Porrino, L.; Crane, A.; Goldman-Rakic, P. Direct and indirect pathways from the amygdala to the frontal lobe in rhesus monkeys. J. Comp. Neurol. 1981, 198, 121–136. [Google Scholar] [CrossRef]
- Fukuyama, K.; Okada, M. Age-Dependent and Sleep/Seizure-Induced Pathomechanisms of Autosomal Dominant Sleep-Related Hypermotor Epilepsy. Int. J. Mol. Sci. 2020, 21, 8142. [Google Scholar]
- Fukuyama, K.; Fukuzawa, M.; Shiroyama, T.; Okada, M. Pathogenesis and pathophysiology of autosomal dominant sleep-related hypermotor epilepsy with S284L-mutant alpha4 subunit of nicotinic ACh receptor. Br. J. Pharmacol. 2020, 177, 2143–2162. [Google Scholar] [CrossRef]
- Fukuyama, K.; Fukuzawa, M.; Shiroyama, T.; Okada, M. Pathomechanism of nocturnal paroxysmal dystonia in autosomal dominant sleep-related hypermotor epilepsy with S284L-mutant alpha4 subunit of nicotinic ACh receptor. Biomed. Pharmacother. 2020, 126, 110070. [Google Scholar]
- Fukuyama, K.; Fukuzawa, M.; Okubo, R.; Okada, M. Upregulated Connexin 43 Induced by Loss-of-Functional S284L-Mutant alpha4 Subunit of Nicotinic ACh Receptor Contributes to Pathomechanisms of Autosomal Dominant Sleep-Related Hypermotor Epilepsy. Pharmaceuticals 2020, 13, 58. [Google Scholar] [CrossRef] [Green Version]
- Fukuyama, K.; Fukuzawa, M.; Okada, M. Upregulated and Hyperactivated Thalamic Connexin 43 Plays Important Roles in Pathomechanisms of Cognitive Impairment and Seizure of Autosomal Dominant Sleep-Related Hypermotor Epilepsy with S284L-Mutant alpha4 Subunit of Nicotinic ACh Receptor. Pharmaceuticals 2020, 13, 99. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Fukuyama, K.; Kawano, Y.; Shiroyama, T.; Suzuki, D.; Ueda, Y. Effects of Acute and Sub-chronic Administrations of Guanfacine on Catecholaminergic Transmissions in the Orbitofrontal Cortex. Neuropharmacology 2019. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, K.; Hasegawa, T.; Okada, M. Cystine/Glutamate Antiporter and Aripiprazole Compensate NMDA Antagonist-Induced Dysfunction of Thalamocortical L-Glutamatergic Transmission. Int. J. Mol. Sci. 2018, 19, 3645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohoyama, K.; Yamamura, S.; Hamaguchi, T.; Nakagawa, M.; Motomura, E.; Shiroyama, T.; Tanii, H.; Okada, M. Effect of novel atypical antipsychotic, blonanserin, on extracellular neurotransmitter level in rat prefrontal cortex. Eur. J. Pharmacol. 2011, 653, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Ohoyama, K.; Hamaguchi, T.; Nakagawa, M.; Suzuki, D.; Matsumoto, T.; Motomura, E.; Tanii, H.; Shiroyama, T.; Okada, M. Effects of zotepine on extracellular levels of monoamine, GABA and glutamate in rat prefrontal cortex. Br. J. Pharmacol. 2009, 157, 656–665. [Google Scholar] [CrossRef]
- Furtak, S.C.; Wei, S.M.; Agster, K.L.; Burwell, R.D. Functional neuroanatomy of the parahippocampal region in the rat: The perirhinal and postrhinal cortices. Hippocampus 2007, 17, 709–722. [Google Scholar] [CrossRef]
- Nakano, T.; Hasegawa, T.; Suzuki, D.; Motomura, E.; Okada, M. Amantadine Combines Astroglial System Xc− Activation with Glutamate/NMDA Receptor Inhibition. Biomolecules 2019, 9, 191. [Google Scholar]
- Woodward, N.D.; Purdon, S.E.; Meltzer, H.Y.; Zald, D.H. A meta-analysis of neuropsychological change to clozapine, olanzapine, quetiapine, and risperidone in schizophrenia. Int. J. Neuropsychopharmacol. 2005, 8, 457–472. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, H.Y.; Share, D.B.; Jayathilake, K.; Salomon, R.M.; Lee, M.A. Lurasidone Improves Psychopathology and Cognition in Treatment-Resistant Schizophrenia. J. Clin. Psychopharmacol. 2020, 40, 240–249. [Google Scholar] [CrossRef]
- Luo, L.L.; Chen, X.; Chai, Y.; Li, J.H.; Zhang, M.; Zhang, J.N. A distinct pattern of memory and attention deficiency in patients with depression. Chin. Med. J. 2013, 126, 1144–1149. [Google Scholar]
- Shilyansky, C.; Williams, L.M.; Gyurak, A.; Harris, A.; Usherwood, T.; Etkin, A. Effect of antidepressant treatment on cognitive impairments associated with depression: A randomised longitudinal study. Lancet Psychiatry 2016, 3, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Reppermund, S.; Zihl, J.; Lucae, S.; Horstmann, S.; Kloiber, S.; Holsboer, F.; Ising, M. Persistent cognitive impairment in depression: The role of psychopathology and altered hypothalamic-pituitary-adrenocortical (HPA) system regulation. Biol. Psychiatry 2007, 62, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Knorr, U.; Madsen, J.M.; Kessing, L.V. The effect of selective serotonin reuptake inhibitors in healthy subjects revisited: A systematic review of the literature. Exp. Clin. Psychopharmacol. 2019, 27, 413–432. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, R.S.; Florea, I.; Tonnoir, B.; Loft, H.; Lam, R.W.; Christensen, M.C. Efficacy of Vortioxetine on Cognitive Functioning in Working Patients With Major Depressive Disorder. J. Clin. Psychiatry 2017, 78, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahableshwarkar, A.R.; Zajecka, J.; Jacobson, W.; Chen, Y.; Keefe, R.S. A Randomized, Placebo-Controlled, Active-Reference, Double-Blind, Flexible-Dose Study of the Efficacy of Vortioxetine on Cognitive Function in Major Depressive Disorder. Neuropsychopharmacology 2015, 40, 2025–2037. [Google Scholar] [CrossRef] [Green Version]
- Perez-Palomar, B.; Mollinedo-Gajate, I.; Berrocoso, E.; Meana, J.J.; Ortega, J.E. Serotonin 5-HT3 receptor antagonism potentiates the antidepressant activity of citalopram. Neuropharmacology 2018, 133, 491–502. [Google Scholar] [CrossRef]
- McGrath, J.C.; Drummond, G.B.; McLachlan, E.M.; Kilkenny, C.; Wainwright, C.L. Guidelines for reporting experiments involving animals: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1573–1576. [Google Scholar] [CrossRef] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotoxic Coordinates, 6th ed.; Academic Press: San Diego, CA, USA, 2007. [Google Scholar]
- Okada, M.; Yoshida, S.; Zhu, G.; Hirose, S.; Kaneko, S. Biphasic actions of topiramate on monoamine exocytosis associated with both soluble N-ethylmaleimide-sensitive factor attachment protein receptors and Ca2+-induced Ca2+-releasing systems. Neuroscience 2005, 134, 233–246. [Google Scholar] [CrossRef]
- Tanahashi, S.; Yamamura, S.; Nakagawa, M.; Motomura, E.; Okada, M. Effect of lamotrigine and carbamazepine on corticotropin-releasing factor-associated serotonergic transmission in rat dorsal raphe nucleus. Psychopharmacology 2012, 220, 599–610. [Google Scholar] [CrossRef]
- Yamamura, S.; Hoshikawa, M.; Dai, K.; Saito, H.; Suzuki, N.; Niwa, O.; Okada, M. ONO-2506 inhibits spike-wave discharges in a genetic animal model without affecting traditional convulsive tests via gliotransmission regulation. Br. J. Pharmacol. 2013, 168, 1088–1100. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, S.; Hamaguchi, T.; Ohoyama, K.; Sugiura, Y.; Suzuki, D.; Kanehara, S.; Nakagawa, M.; Motomura, E.; Matsumoto, T.; Tanii, H.; et al. Topiramate and zonisamide prevent paradoxical intoxication induced by carbamazepine and phenytoin. Epilepsy Res. 2009, 84, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, K.; Tanahashi, S.; Hoshikawa, M.; Shinagawa, R.; Okada, M. Zonisamide regulates basal ganglia transmission via astroglial kynurenine pathway. Neuropharmacology 2014, 76, 137–145. [Google Scholar] [CrossRef]
- Fukuyama, K.; Ueda, Y.; Okada, M. Effects of Carbamazepine, Lacosamide and Zonisamide on Gliotransmitter Release Associated with Activated Astroglial Hemichannels. Pharmaceuticals 2020, 13, 117. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, K.; Okada, M. Effects of levetiracetam on astroglial release of kynurenine-pathway metabolites. Br. J. Pharmacol. 2018, 175, 4253–4265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, M.J.; Alexander, S.; Cirino, G.; Docherty, J.R.; George, C.H.; Giembycz, M.A.; Hoyer, D.; Insel, P.A.; Izzo, A.A.; Ji, Y.; et al. Experimental design and analysis and their reporting II: Updated and simplified guidance for authors and peer reviewers. Br. J. Pharmacol. 2018, 175, 987–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okada, M.; Matsumoto, R.; Yamamoto, Y.; Fukuyama, K. Effects of Subchronic Administrations of Vortioxetine, Lurasidone, and Escitalopram on Thalamocortical Glutamatergic Transmission Associated with Serotonin 5-HT7 Receptor. Int. J. Mol. Sci. 2021, 22, 1351. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031351
Okada M, Matsumoto R, Yamamoto Y, Fukuyama K. Effects of Subchronic Administrations of Vortioxetine, Lurasidone, and Escitalopram on Thalamocortical Glutamatergic Transmission Associated with Serotonin 5-HT7 Receptor. International Journal of Molecular Sciences. 2021; 22(3):1351. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031351
Chicago/Turabian StyleOkada, Motohiro, Ryusuke Matsumoto, Yoshimasa Yamamoto, and Kouji Fukuyama. 2021. "Effects of Subchronic Administrations of Vortioxetine, Lurasidone, and Escitalopram on Thalamocortical Glutamatergic Transmission Associated with Serotonin 5-HT7 Receptor" International Journal of Molecular Sciences 22, no. 3: 1351. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031351