
Impressic Acid Ameliorates Atopic Dermatitis-Like Skin Lesions by Inhibiting ERK1/2-Mediated Phosphorylation of NF-κB and STAT1


 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. IPA Attenuated DNCB-Induced AD-Like Skin Severity in Mice
2.2. IPA Attenuated DNCB-Induced Increased Levels of Thymic Stromal Lymphopoietin (TSLP) in Serum and Ear Lesions
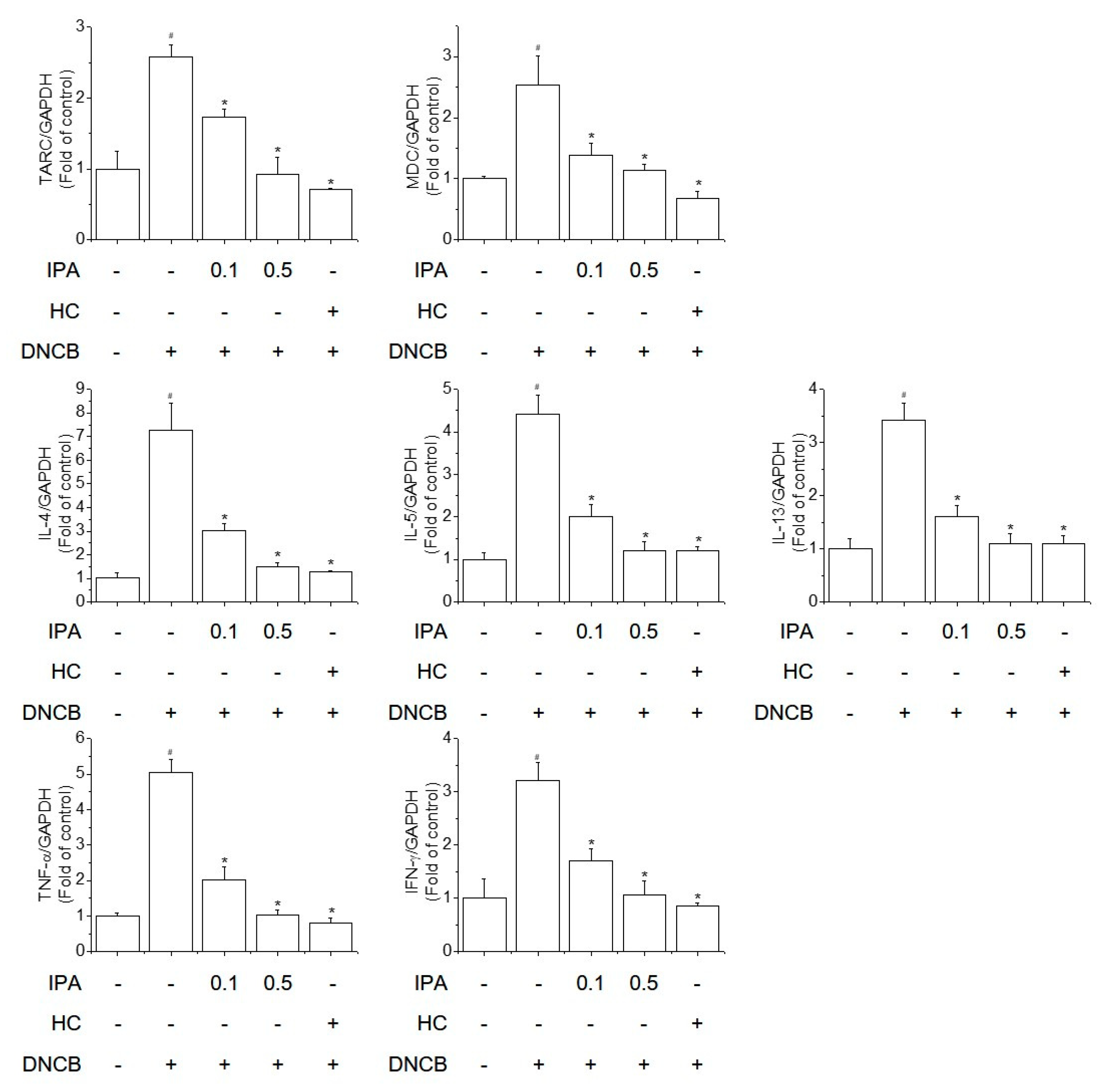
2.3. IPA Suppressed the DNCB-Increased mRNA Levels of Th1/Th2 Cytokines and Chemokines in Ear Lesions
2.4. IPA Inhibited the DNCB-Induced Increased Activation of NF-κB in Ear Lesions
2.5. IPA Suppressed the TNF-α/IFN-γ-Induced Increased mRNA Levels of TSLP, TARC and MDC in Keratinocytes
2.6. IPA Inhibited TNF-α/IFN-γ-Induced NF-κB, MAPK, and STAT1 Activation in Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Animals and Treatment
4.3. Histopathological Analysis of Ear Tissue
4.4. Measurement of Ear Thickness
4.5. ELISA
4.6. Cell Culture and Treatment
4.7. Assays of Cell Viability and Cytotoxicity
4.8. RNA Extraction and Quantitative Real-Time Reverse Transcription PCR
4.9. Transient Transfection and Luciferase Assay
4.10. Western Blotting
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AD | Atopic dermatitis |
| DNCB | 2,4-dinitrochlorobenzene |
| ERK1/2 | Extracellular signal-regulated protein kinase |
| IFN-γ | Interferon-gamma |
| IgE | Immunoglobulin E |
| IL-4 | Interleukin-4 |
| IL-5 | Interleukin-5 |
| IL-13 | Interleukin-13 |
| IPA | Impressic acid |
| IκBα | Inhibitor of nuclear factor kappa B |
| MDC | Macrophage derived chemokine |
| NF-κB | Nuclear factor-kappa B |
| STAT1 | Signal transducer and activator of transcription 1 |
| TARC | Thymus and activation regulated Chemokine |
| TNF-α | Tumor necrosis factor-alpha |
| TSLP | Thymic stromal lymphopoietin |
References
- Boguniewicz, M.; Leung, D.Y. Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, S.H. Epidermal permeability barrier defects and barrier repair therapy in atopic dermatitis. Allergy Asthma Immunol. Res. 2014, 6, 276–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czarnowicki, T.; Krueger, J.G.; Guttman-Yassky, E. Novel concepts of prevention and treatment of atopic dermatitis through barrier and immune manipulations with implications for the atopic march. J. Allergy Clin. Immunol. 2017, 139, 1723–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, E.B.; Sivaprasad, U. Th2 Cytokines and Atopic Dermatitis. J. Clin. Cell. Immunol. 2011, 2, 110. [Google Scholar] [CrossRef]
- Blakely, K.; Gooderham, M.; Papp, K.; Dupilumab, A. Monoclonal Antibody for Atopic Dermatitis: A Review of Current Literature. Skin Therapy Lett. 2016, 21, 1–5. [Google Scholar]
- Shah, P.P.; Desai, P.R.; Singh, M. Effect of oleic acid modified polymeric bilayered nanoparticles on percutaneous delivery of spantide II and ketoprofen. J. Control. Release 2012, 158, 336–345. [Google Scholar] [CrossRef] [Green Version]
- Misery, L. Therapeutic perspectives in atopic dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 267–271. [Google Scholar] [CrossRef]
- Berke, R.; Singh, A.; Guralnick, M. Atopic dermatitis: An overview. Am. Fam. Physician. 2012, 86, 35–42. [Google Scholar]
- Zhou, Z.; Shi, T.; Hou, J.; Li, M. Ferulic acid alleviates atopic dermatitis-like symptoms in mice via its potent anti-inflammatory effect. Immunopharmacol. Immunotoxicol. 2020, 42, 156–164. [Google Scholar] [CrossRef]
- Arisawa, M.; Fujita, A.; Saga, M.; Hayashi, T.; Morita, N.; Kawano, N.; Koshimura, S. Studies on cytotoxic constituents in pericarps of Mallotus japonicus, Part II. J. Nat. Prod. 1986, 49, 298–302. [Google Scholar] [CrossRef]
- Park, S.H.; Nhiem, N.X.; Kiem, P.V.; Choi, E.M.; Kim, J.A.; Kim, Y.H. A new norlupane triterpene from the leaves of Acanthopanax koreanum increases the differentiation of osteoblastic MC3T3-e1 cells. Arch. Pharm. Res. 2010, 33, 75–80. [Google Scholar] [CrossRef]
- Kim, J.A.; Yang, S.Y.; Koo, J.E.; Koh, Y.S.; Kim, Y.H. Lupane-type triterpenoids from the steamed leaves of Acanthopanax koreanum and their inhibitory effects on the LPS-stimulated pro-inflammatory cytokine production in bone marrow-derived dendritic cells. Bioorg. Med. Chem. Lett. 2010, 20, 6703–6707. [Google Scholar] [CrossRef]
- Jin, S.W.; Pham, H.T.; Choi, J.H.; Lee, G.H.; Han, E.H.; Cho, Y.H.; Chung, Y.C.; Kim, Y.H.; Jeong, H.G. Impressic Acid, a Lupane-Type Triterpenoid from Acanthopanax koreanum, Attenuates TNF-alpha-Induced Endothelial Dysfunction via Activation of eNOS/NO Pathway. Int. J. Mol. Sci. 2019, 20, 5772. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.; Min, D.S.; Yun, H.E.; Kim, K.T.; Sun, Y.N.; Dat, L.D.; Kim, Y.H.; Kim, H.P. Impressic acid from Acanthopanax koreanum, possesses matrix metalloproteinase-13 down-regulating capacity and protects cartilage destruction. J. Ethnopharmacol. 2017, 209, 73–81. [Google Scholar] [CrossRef]
- Kim, J.A.; Yang, S.Y.; Song, S.B.; Kim, Y.H. Effects of impressic acid from Acanthopanax koreanum on NF-kappaB and PPARgamma activities. Arch. Pharm. Res. 2011, 34, 1347–1351. [Google Scholar] [CrossRef]
- Lee, G.H.; Kim, J.Y.; Jin, S.W.; Pham, T.H.; Park, J.S.; Kim, C.Y.; Choi, J.H.; Han, E.H.; Kim, Y.H.; Jeong, H.G. Impressic Acid Attenuates the Lipopolysaccharide-Induced Inflammatory Response by Activating the AMPK/GSK3β/Nrf2 Axis in RAW264.7 Macrophages. Int. J. Mol. Sci. 2021, 22, 762. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jin, S.W.; Lee, G.H.; Cho, S.M.; Jeong, H.G. Orostachys japonicus ethanol extract inhibits 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in NC/Nga mice and TNF-alpha/IFN-gamma-induced TARC expression in HaCaT cells. Toxicol. Res. 2020, 36, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Ahn, E.K.; Ko, H.J.; Lee, J.Y.; Hwang, S.M.; Ko, S.; Oh, J.S. Korean red ginseng water extract alleviates atopic dermatitis-like inflammatory responses by negative regulation of mitogen-activated protein kinase signaling pathway in vivo. Biomed. Pharmacother. 2019, 117, 109066. [Google Scholar] [CrossRef]
- Wu, P.C.; Chuo, W.H.; Lin, S.C.; Lehman, C.W.; Lien, C.Z.; Wu, C.S.; Lin, C.C. Sclareol attenuates the development of atopic dermatitis induced by 2,4-dinitrochlorobenzene in mice. Immunopharmacol. Immunotoxicol. 2019, 41, 109–116. [Google Scholar] [CrossRef]
- Yang, G.; Lee, H.E.; Lim, K.M.; Choi, Y.K.; Kim, K.B.; Lee, B.M.; Lee, J.Y. Potentiation of skin TSLP production by a cosmetic colorant leads to aggravation of dermatitis symptoms. Chem. Biol. Interact. 2018, 284, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Hong, C. Osthole attenuates mouse atopic dermatitis by inhibiting thymic stromal lymphopoietin production from keratinocytes. Exp. Dermatol. 2019, 28, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Choi, Y.A.; Yu, J.H.; Jung, H.D.; Lee, S.; Park, P.H.; Lee, H.S.; Kwon, T.K.; Shin, T.Y.; Lee, S.W.; Rho, M.C.; et al. Inhibitory effect of ethanol extract of Ampelopsis brevipedunculata rhizomes on atopic dermatitis-like skin inflammation. J. Ethnopharmacol. 2019, 238, 111850. [Google Scholar] [CrossRef]
- Klonowska, J.; Glen, J.; Nowicki, R.J.; Trzeciak, M. New Cytokines in the Pathogenesis of Atopic Dermatitis-New Therapeutic Targets. Int. J. Mol. Sci. 2018, 19, 3086. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Jeong, Y.; Ashraf, M.U.; Bae, Y.S. Dendritic Cell-Mediated Th2 Immunity and Immune Disorders. Int. J. Mol. Sci. 2019, 20, 2159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giustizieri, M.L.; Mascia, F.; Frezzolini, A.; De Pita, O.; Chinni, L.M.; Giannetti, A.; Girolomoni, G.; Pastore, S. Keratinocytes from patients with atopic dermatitis and psoriasis show a distinct chemokine production profile in response to T cell-derived cytokines. J. Allergy Clin. Immunol. 2001, 107, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Kohno, K.; Koya-Miyata, S.; Harashima, A.; Ariyasu, T.; Ushio, S. NK-4 exerts selective regulatory effects on the activation and function of allergy-related Th2 cells. PLoS ONE 2018, 13, e0199666. [Google Scholar] [CrossRef]
- Lee, B.; Heo, J.; Hong, S.; Kim, E.Y.; Sohn, Y.J.; Jung, H.S. dl-Malic acid as a component of alpha-hydroxy acids: Effect on 2,4-dinitrochlorobenzene-induced inflammation in atopic dermatitis-like skin lesions in vitro and in vivo. Immunopharmacol. Immunotoxicol. 2019, 41, 614–621. [Google Scholar] [CrossRef]
- Yang, J.H.; Do, H.J.; Lee, E.; Yim, N.H.; Cho, W.K.; Park, K.I.; Ma, J.Y. Jageum-Jung improves 2,4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice and suppresses pro-inflammatory chemokine production by inhibiting TNF-alpha/IFN-gamma-induced STAT-1 and NFkappaB signaling in HaCaT cells. J. Ethnopharmacol. 2018, 221, 48–55. [Google Scholar] [CrossRef]
- Hulshof, L.; Van’t Land, B.; Sprikkelman, A.B.; Garssen, J. Role of Microbial Modulation in Management of Atopic Dermatitis in Children. Nutrients 2017, 9, 854. [Google Scholar] [CrossRef] [Green Version]
- Kapur, S.; Watson, W.; Carr, S. Atopic dermatitis. Allergy Asthma Clin. Immunol. 2018, 14, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The Traditional Medicine and Modern Medicine from Natural Products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, S.; Park, W. Modulation of IL-12 and IFN-γ Secretions by Eleutheroside E, Tortoside A, and Syringaresinol from Acanthopanax koreanum Nakai. Biomol. Ther. 2010, 18, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Cai, X.F.; Lee, I.S.; Shen, G.; Dat, N.T.; Lee, J.J.; Kim, Y.H. Triterpenoids from Acanthopanax koreanum root and their inhibitory activities on NFAT transcription. Arch. Pharm. Res. 2004, 27, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Seong, G.S.; Choung, S.Y. Fermented Morinda citrifolia (Noni) Alleviates DNCB-Induced Atopic Dermatitis in NC/Nga Mice through Modulating Immune Balance and Skin Barrier Function. Nutrients 2020, 12, 249. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.C.; Cho, K.; Lee, J.H.; Subedi, L.; Yumnam, S.; Kim, S.Y. Effect of Resveratrol-Enriched Rice on Skin Inflammation and Pruritus in the NC/Nga Mouse Model of Atopic Dermatitis. Int. J. Mol. Sci. 2019, 20, 1428. [Google Scholar] [CrossRef] [Green Version]
- Cianferoni, A.; Spergel, J. The importance of TSLP in allergic disease and its role as a potential therapeutic target. Expert Rev. Clin. Immunol. 2014, 10, 1463–1474. [Google Scholar] [CrossRef] [Green Version]
- Indra, A.K. Epidermal TSLP: A trigger factor for pathogenesis of atopic dermatitis. Expert Rev. Proteomics 2013, 10, 309–311. [Google Scholar] [CrossRef] [Green Version]
- Rerknimitr, P.; Otsuka, A.; Nakashima, C.; Kabashima, K. The etiopathogenesis of atopic dermatitis: Barrier disruption, immunological derangement, and pruritus. Inflamm. Regen. 2017, 37, 14. [Google Scholar] [CrossRef]
- Brunner, P.M.; Guttman-Yassky, E.; Leung, D.Y. The immunology of atopic dermatitis and its reversibility with broad-spectrum and targeted therapies. J. Allergy Clin. Immunol. 2017, 139, S65–S76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biedermann, T.; Skabytska, Y.; Kaesler, S.; Volz, T. Regulation of T Cell Immunity in Atopic Dermatitis by Microbes: The Yin and Yang of Cutaneous Inflammation. Front. Immunol. 2015, 6, 353. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Hwang, M.H.; Cho, Y.R.; Hong, S.S.; Kang, J.S.; Kim, W.H.; Yang, S.H.; Seo, D.W.; Oh, J.S.; Ahn, E.K. Combretum quadrangulare Extract Attenuates Atopic Dermatitis-Like Skin Lesions through Modulation of MAPK Signaling in BALB/c Mice. Molecules 2020, 25, 2003. [Google Scholar] [CrossRef] [PubMed]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-kappaB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407–419. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.H.; Suh, B.; Shin, I.; Kim, E.H.; Kim, D.; Shin, Y.J.; Chang, S.Y.; Baek, S.H.; Kim, H.; Bae, O.N. Inhibitory Effects of a Novel Chrysin-Derivative, CPD 6, on Acute and Chronic Skin Inflammation. Int. J. Mol. Sci. 2019, 20, 2607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, T.Y.; Kang, Y.M.; Eom, Y.J.; Hong, C.H.; An, H.J. Anti-Atopic Dermatitis Effect of Seaweed Fulvescens Extract via Inhibiting the STAT1 Pathway. Mediators Inflamm. 2019, 2019, 3760934. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Park, B.H.; Kim, H.G.; Hwang, Y.P.; Han, E.H.; Jin, S.W.; Seo, J.K.; Chung, Y.C.; Jeong, H.G. Inhibitory effect of Psidium guajava water extract in the development of 2,4-dinitrochlorobenzene-induced atopic dermatitis in NC/Nga mice. Food Chem Toxicol. 2012, 50, 2923. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, J.H.; Lee, G.H.; Jin, S.W.; Kim, J.Y.; Hwang, Y.P.; Han, E.H.; Kim, Y.H.; Jeong, H.G. Impressic Acid Ameliorates Atopic Dermatitis-Like Skin Lesions by Inhibiting ERK1/2-Mediated Phosphorylation of NF-κB and STAT1. Int. J. Mol. Sci. 2021, 22, 2334. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052334
Choi JH, Lee GH, Jin SW, Kim JY, Hwang YP, Han EH, Kim YH, Jeong HG. Impressic Acid Ameliorates Atopic Dermatitis-Like Skin Lesions by Inhibiting ERK1/2-Mediated Phosphorylation of NF-κB and STAT1. International Journal of Molecular Sciences. 2021; 22(5):2334. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052334
Chicago/Turabian StyleChoi, Jae Ho, Gi Ho Lee, Sun Woo Jin, Ji Yeon Kim, Yong Pil Hwang, Eun Hee Han, Young Ho Kim, and Hye Gwang Jeong. 2021. "Impressic Acid Ameliorates Atopic Dermatitis-Like Skin Lesions by Inhibiting ERK1/2-Mediated Phosphorylation of NF-κB and STAT1" International Journal of Molecular Sciences 22, no. 5: 2334. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22052334