
Sera from Patients with NMOSD Reduce the Differentiation Capacity of Precursor Cells in the Central Nervous System

 ,
,  , , ,
, , , 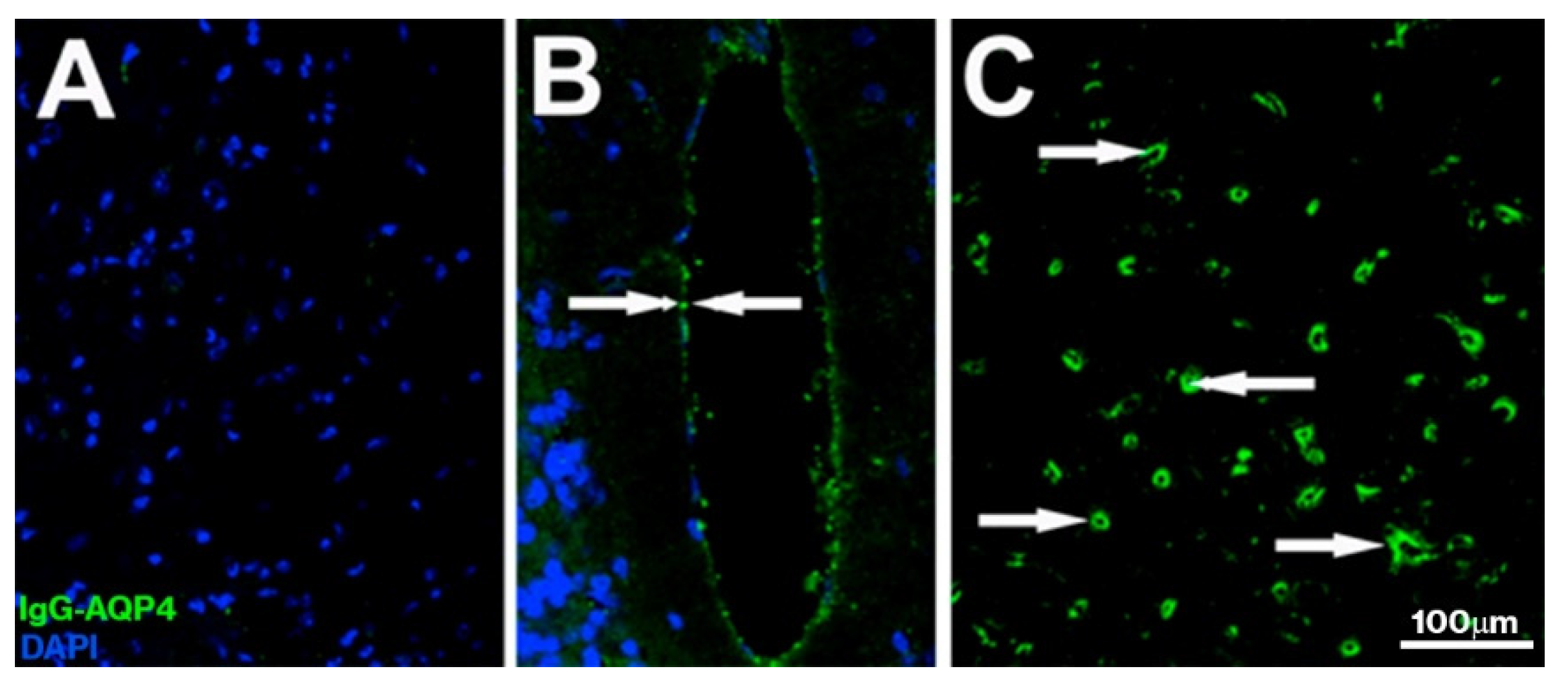{kind=link}
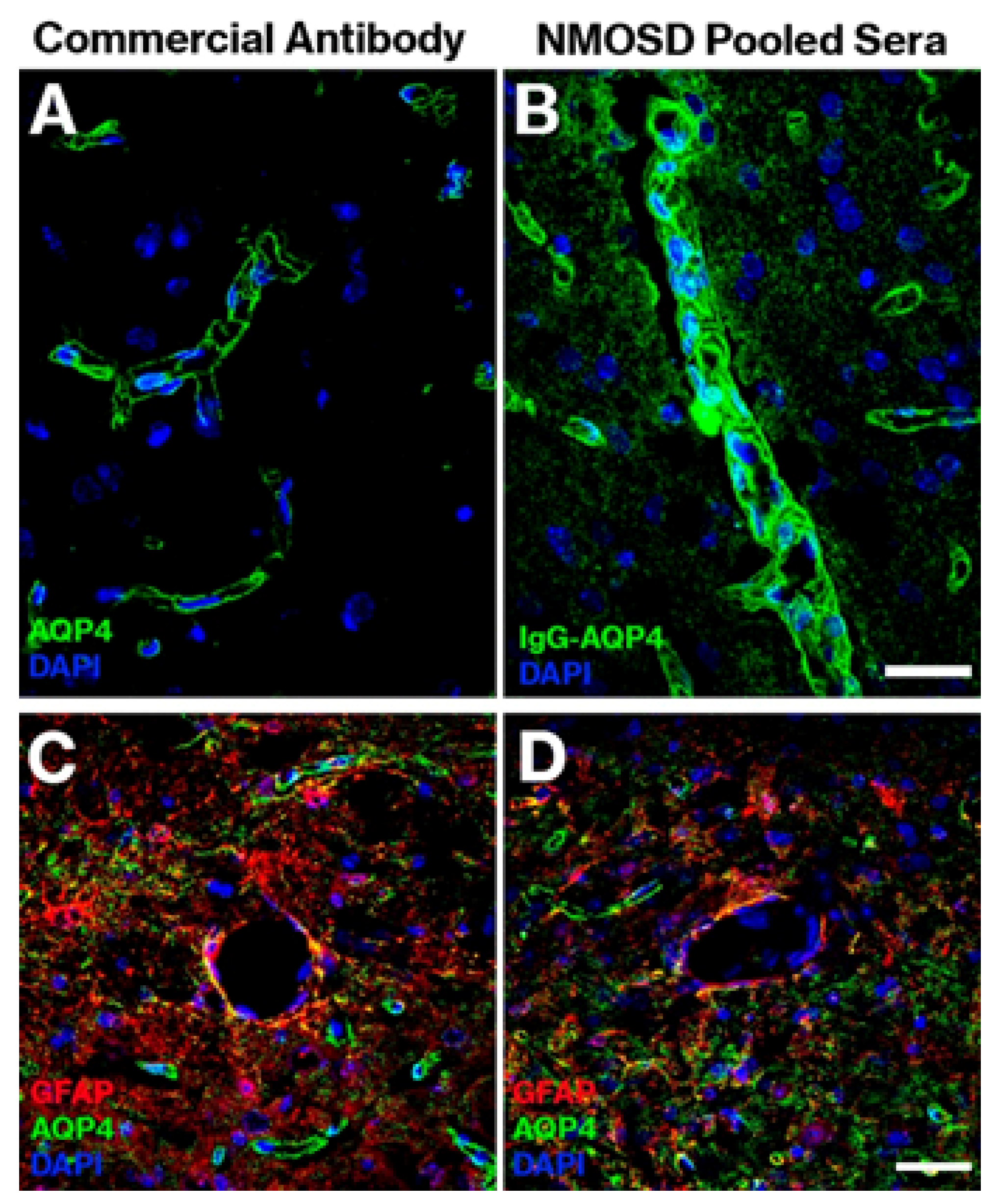{kind=link}
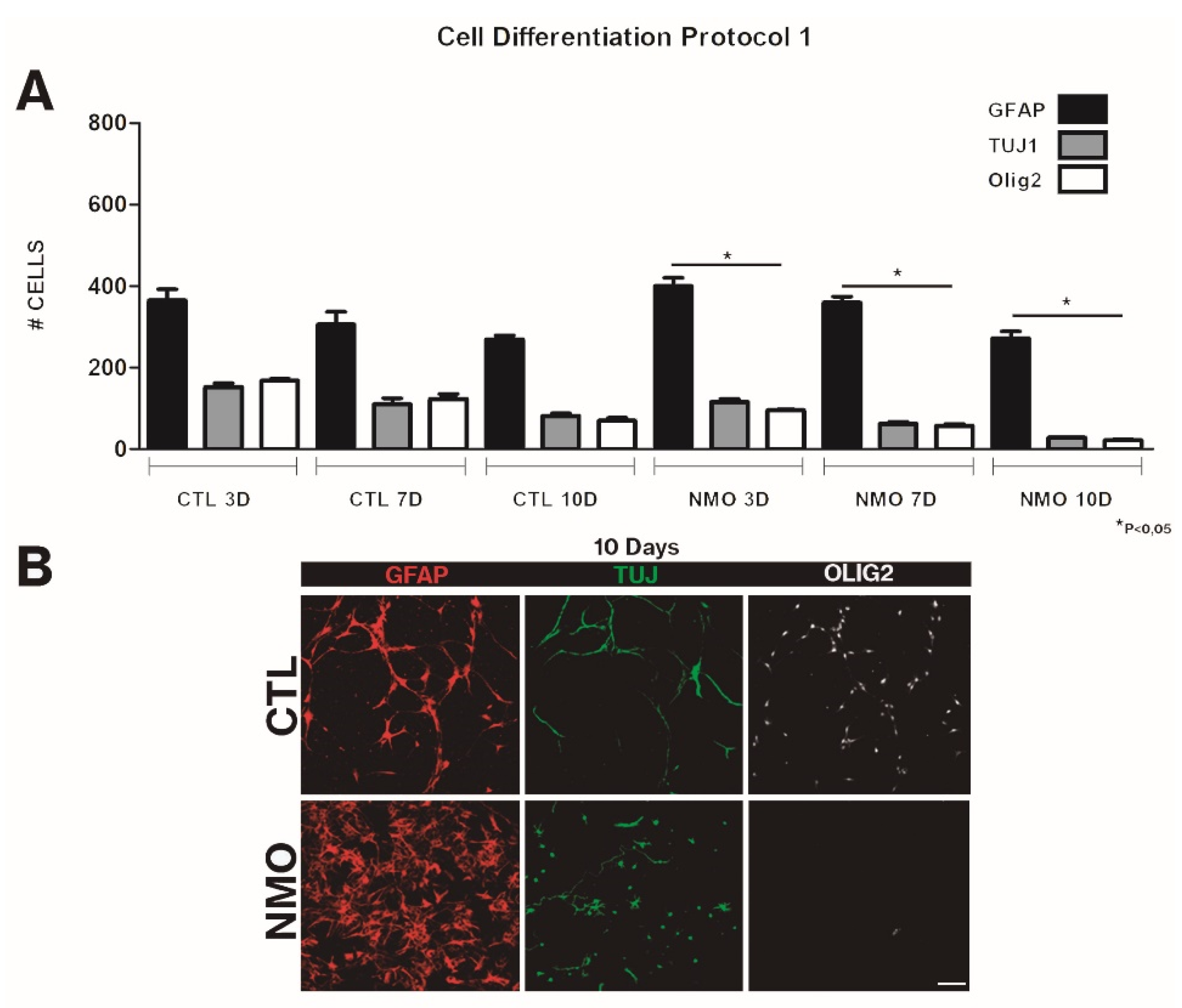{kind=link}
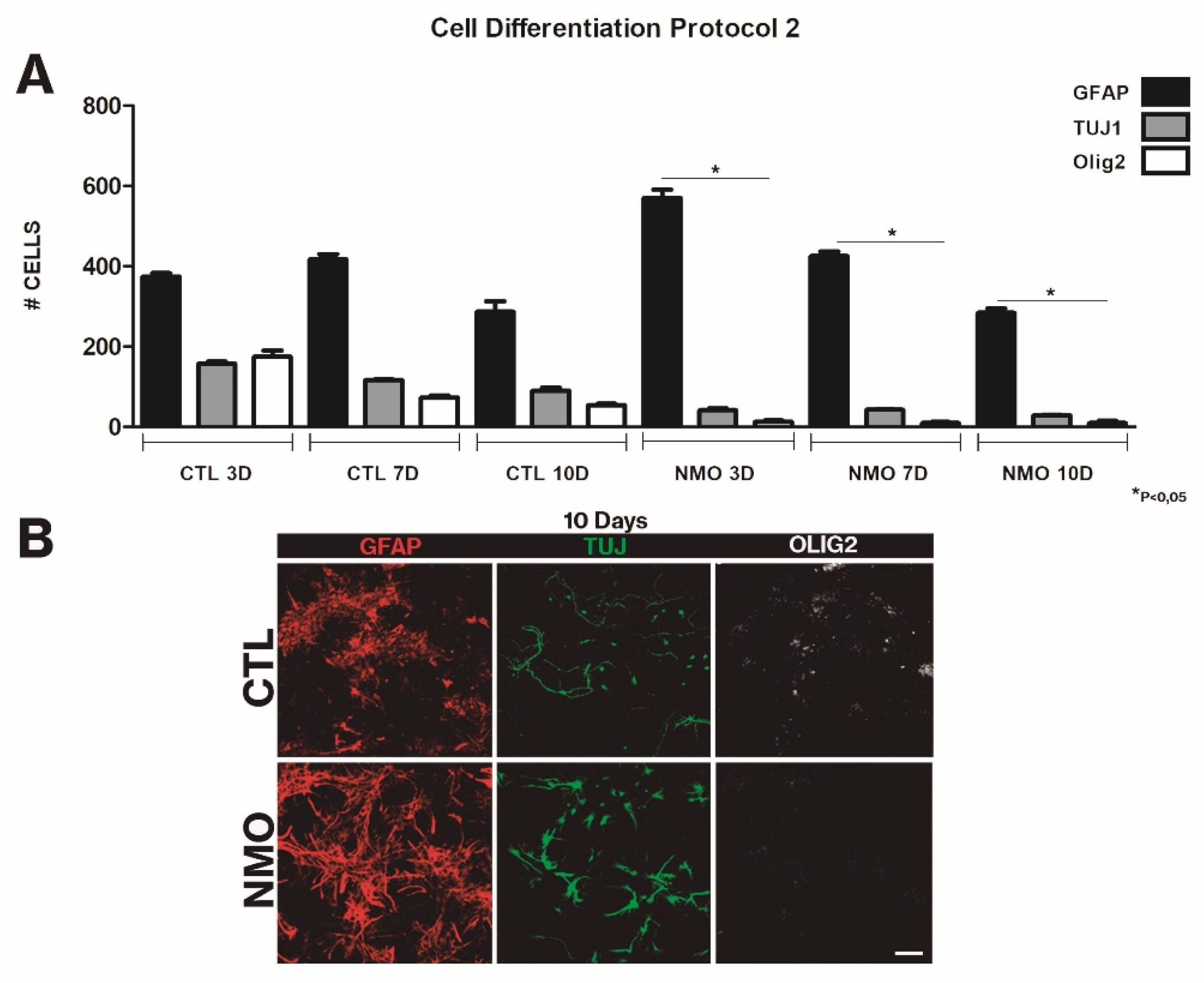{kind=link}
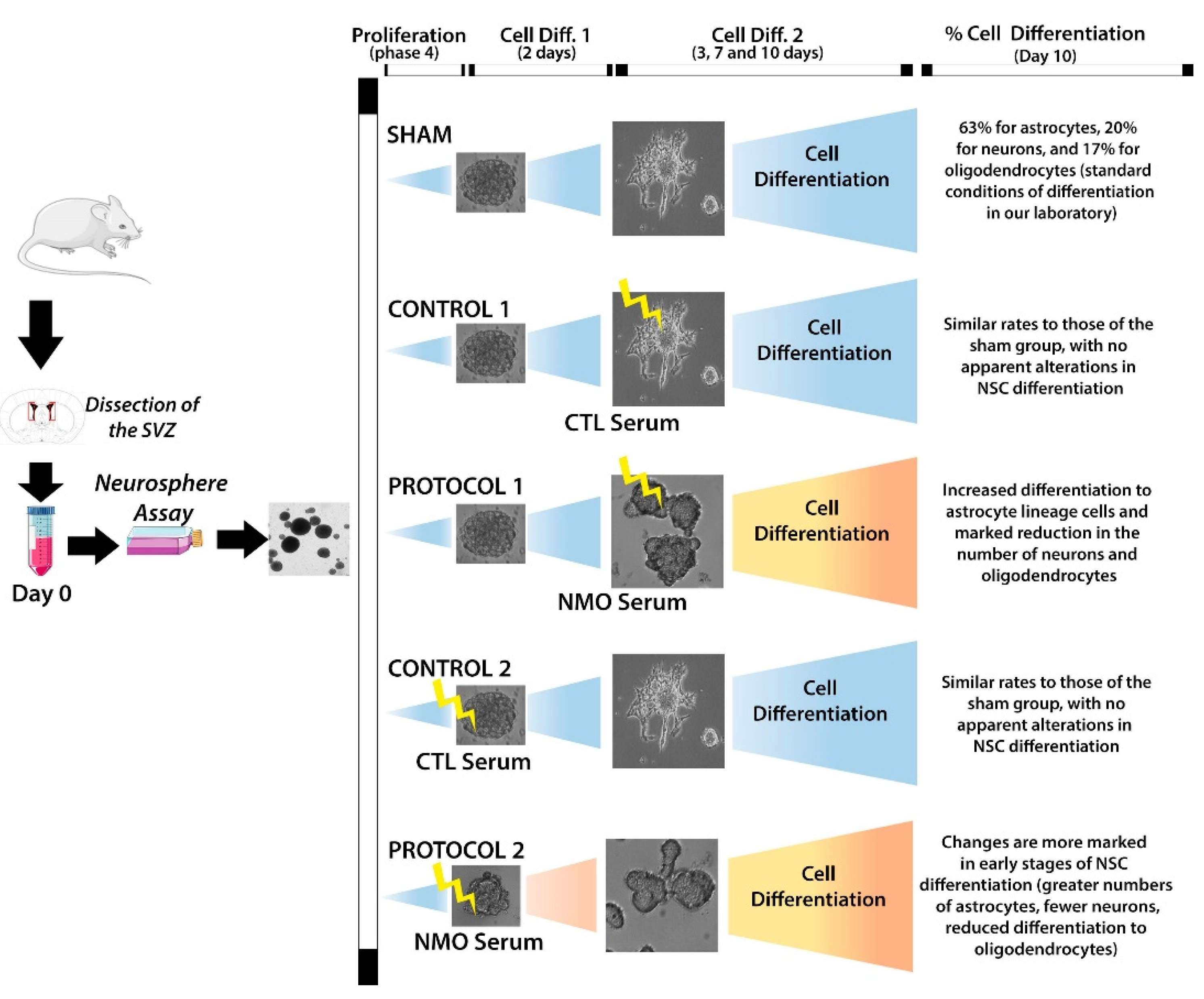{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Patients and Controls
2.2. Effect on SVZ Cell Differentiation
3. Discussion
4. Material and Methods
4.1. Study Groups
4.2. Experimental Animals
4.3. Sample Extraction and Storage, and Confirmation of the Presence of AQP4–IgG in Serum Samples and Mouse Cerebella Treated with the Serum
4.4. Primary Culture of Neurospheres from the Subventricular Zone
4.5. Effects of the Serum Sample on Differentiation
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, Y.; Zhong, L.; Geng, J. Neuromyelitis optica spectrum disorder: Pathogenesis, treatment, and experimental models. Mult. Scler. Relat. Disord. 2019, 27, 412–418. [Google Scholar] [CrossRef]
- Hor, J.Y.; Asgari, N.; Nakashima, I.; Broadley, S.A.; Leite, M.I.; Kissani, N.; Jacob, A.; Marignier, R.; Weinshenker, B.G.; Paul, F.; et al. Epidemiology of Neuromyelitis Optica Spectrum Disorder and Its Prevalence and Incidence Worldwide. Front. Neurol. 2020, 11, 501. [Google Scholar] [CrossRef]
- Waters, P.J.; Pittock, S.J.; Bennett, J.L.; Jarius, S.; Weinshenker, B.G.; Wingerchuk, D.M. Evaluation of aquaporin-4 antibody assays. Clin. Exp. Neuroimmunol. 2014, 5, 290–303. [Google Scholar] [CrossRef] [Green Version]
- Kessler, R.A.; Mealy, M.A.; Levy, M.Z. Treatment of Neuromyelitis Optica Spectrum Disorder: Acute, Preventive, and Symptomatic. Curr. Treat. Options Neurol. 2016, 18, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicchia, G.P.; Mastrototaro, M.; Rossi, A.; Pisani, F.; Tortorella, C.; Ruggieri, M.; Lia, A.; Trojano, M.; Frigeri, A.; Svelto, M. Aquaporin-4 orthogonal arrays of particles are the target for neuromyelitis optica autoantibodies. Glia 2009, 57, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Pisani, F.; Mastrototaro, M.; Rossi, A.; Nicchia, G.P.; Tortorella, C.; Ruggieri, M.; Trojano, M.; Frigeri, A.; Svelto, M. Identification of Two Major Conformational Aquaporin-4 Epitopes for Neuromyelitis Optica Autoantibody Binding. J. Biol. Chem. 2011, 286, 9216–9224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, F.; Takeshita, Y.; Hamamoto, Y.; Nishihara, H.; Sano, Y.; Honda, M.; Sato, R.; Maeda, T.; Takahashi, T.; Fujikawa, S.; et al. GRP 78 antibodies are associated with clinical phenotype in neuromyelitis optica. Ann. Clin. Transl. Neurol. 2019, 6, 2079–2087. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.L.; Owens, G.P. Neuromyelitis Optica: Deciphering a Complex Immune-Mediated Astrocytopathy. J. Neuroophthalmol 2017, 37, 291–299. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, M.C.; Verkman, A. Aquaporin 4 and neuromyelitis optica. Lancet Neurol. 2012, 11, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Verkman, A.S.; Phuan, P.-W.; Asavapanumas, N.; Tradtrantip, L. Biology of AQP4 and Anti-AQP4 Antibody: Therapeutic Implications for NMO. Brain Pathol. 2013, 23, 684–695. [Google Scholar] [CrossRef] [Green Version]
- Verkman, A.S.; Anderson, M.O.; Papadopoulos, M.C. Aquaporins: Important but elusive drug targets. Nat. Rev. Drug Discov. 2014, 13, 259–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abir-Awan, M.; Kitchen, P.; Salman, M.M.; Conner, M.T.; Conner, A.C.; Bill, R.M. Inhibitors of Mammalian Aquaporin Water Channels. Int. J. Mol. Sci. 2019, 20, 1589. [Google Scholar] [CrossRef] [Green Version]
- Matías-Guiu, J.; Gomez-Pinedo, U.; Matias-Guiu, J.A. Novedades en esclerosis múltiple: La remielinización como objetivo terapéutico. Med. Clínica 2017, 148, 377–380. [Google Scholar] [CrossRef]
- Wrzos, C.; Winkler, A.; Metz, I.; Kayser, D.M.; Thal, D.R.; Wegner, C.; Brück, W.; Nessler, S.; Bennett, J.L.; Stadelmann, C. Early loss of oligodendrocytes in human and experimental neuromyelitis optica lesions. Acta Neuropathol. 2014, 127, 523–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brufck, W.; Popescu, B.; Lucchinetti, C.F.; Markovic-Plese, S.; Gold, R.; Thal, D.R.; Metz, I. Neuromyelitis optica lesions may inform multiple sclerosis heterogeneity debate. Ann. Neurol. 2012, 72, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Misu, T.; Höftberger, R.; Fujihara, K.; Wimmer, I.; Takai, Y.; Nishiyama, S.; Nakashima, I.; Konno, H.; Bradl, M.; Garzuly, F.; et al. Presence of six different lesion types suggests diverse mechanisms of tissue injury in neuromyelitis optica. Acta Neuropathol. 2013, 125, 815–827. [Google Scholar] [CrossRef] [Green Version]
- Masaki, K.; Suzuki, S.O.; Matsushita, T.; Matsuoka, T.; Imamura, S.; Yamasaki, R.; Suzuki, M.; Suenaga, T.; Iwaki, T.; Kira, J.-I. Connexin 43 Astrocytopathy Linked to Rapidly Progressive Multiple Sclerosis and Neuromyelitis Optica. PLoS ONE 2013, 8, e72919. [Google Scholar] [CrossRef]
- Richard, C.; Ruiz, A.; Cavagna, S.; Bigotte, M.; Vukusic, S.; Masaki, K.; Suenaga, T.; Kira, J.-I.; Giraudon, P.; Marignier, R. Connexins in neuromyelitis optica: A link between astrocytopathy and demyelination. Brain 2020, 143, 2721–2732. [Google Scholar] [CrossRef]
- Yao, X.; Su, T.; Verkman, A.S. Clobetasol promotes remyelination in a mouse model of neuromyelitis optica. Acta Neuropathol. Commun. 2016, 4, 42. [Google Scholar] [CrossRef] [Green Version]
- Blakemore, W.F.; Gilson, J.M.; Crang, A. The presence of astrocytes in areas of demyelination influences remyelination following transplantation of oligodendrocyte progenitors. Exp. Neurol. 2003, 184, 955–963. [Google Scholar] [CrossRef]
- Li, J.; Zhang, L.; Chu, Y.; Namaka, M.; Deng, B.; Kong, J.; Bi, X. Astrocytes in Oligodendrocyte Lineage Development and White Matter Pathology. Front. Cell. Neurosci. 2016, 10. [Google Scholar] [CrossRef] [Green Version]
- Marignier, R.; Nicolle, A.; Watrin, C.; Touret, M.; Cavagna, S.; Varrin-Doyer, M.; Cavillon, G.; Rogemond, V.; Confavreux, C.; Honnorat, J.; et al. Oligodendrocytes are damaged by neuromyelitis optica immunoglobulin G via astrocyte injury. Brain 2010, 133, 2578–2591. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, P.; Day, R.E.; Salman, M.M.; Conner, M.T.; Bill, R.M.; Conner, A.C. Beyond water homeostasis: Diverse functional roles of mammalian aquaporins. Biochim. Biophys. Acta 2015, 1850, 2410–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belenguer, G.; Domingo-Muelas, A.; Ferrón, S.R.; Morante-Redolat, J.M.; Fariñas, I. Isolation, culture and analysis of adult subependymal neural stem cells. Differentiation 2016, 91, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Hara-Chikuma, M.; Verkman, A.S. Physiological roles of glycerol-transporting aquaporins: The aquaglyceroporins. Cell. Mol. Life Sci. 2006, 63, 1386–1392. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, P.; Salman, M.M.; Pickel, S.U.; Jennings, J.; Törnroth-Horsefield, S.; Conner, M.T.; Bill, R.M.; Conner, A.C. Water channel pore size determines exclusion properties but not solute selectivity. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeshima-Kataoka, H.; Yasui, M. Correlation between astrocyte activity and recovery from blood–brain barrier breakdown caused by brain injury. NeuroReport 2016, 27, 894–900. [Google Scholar] [CrossRef]
- Ishibashi, K.; Hara, S.; Kondo, S. Aquaporin water channels in mammals. Clin. Exp. Nephrol. 2009, 13, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Badaut, J.; Lasbennes, F.; Magistretti, P.J.; Regli, L. Aquaporins in Brain: Distribution, Physiology, and Pathophysiology. Br. J. Pharmacol. 2002, 22, 367–378. [Google Scholar] [CrossRef]
- Salman, M.M.; Kitchen, P.; Woodroofe, M.N.; Brown, J.E.; Bill, R.M.; Conner, A.C.; Conner, M.T. Hypothermia increases aquaporin 4 (AQP4) plasma membrane abundance in human primary cortical astrocytes via a calcium/transient receptor potential vanilloid 4 (TRPV4)- and calmodulin-mediated mechanism. Eur. J. Neurosci. 2017, 46, 2542–2547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitchen, P.; Salman, M.M.; Halsey, A.M.; Clarke-Bland, C.; Macdonald, J.A.; Ishida, H.; Vogel, H.J.; Almutiri, S.; Logan, A.; Kreida, S.; et al. Targeting Aquaporin-4 Subcellular Localization to Treat Central Nervous System Edema. Cell 2020, 181, 784–799. [Google Scholar] [CrossRef]
- Simone, L.; Pisani, F.; Mola, M.G.; De Bellis, M.; Merla, G.; Micale, L.; Frigeri, A.; Vescovi, A.L.; Svelto, M.; Nicchia, G.P. AQP4 Aggregation State Is a Determinant for Glioma Cell Fate. Cancer Res. 2019, 79, 2182–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylvain, N.J.; Salman, M.M.; Pushie, M.J.; Hou, H.; Meher, V.; Herlo, R.; Peeling, L.; Kelly, M.E. The effects of trifluoperazine on brain edema, aquaporin-4 expression and metabolic markers during the acute phase of stroke using photothrombotic mouse model. Biochim. Biophys. Acta 2021, 1863, 183573. [Google Scholar] [CrossRef]
- Cavazzin, C.; Ferrari, D.; Facchetti, F.; Russignan, A.; Vescovi, A.L.; La Porta, C.A.; Gritti, A. Unique expression and localization of aquaporin-4 and aquaporin-9 in murine and human neural stem cells and in their glial progeny. Glia 2006, 53, 167–181. [Google Scholar] [CrossRef]
- Porta, C.A.; Gena, P.; Gritti, A.; Fascio, U.; Svelto, M.; Calamita, G. Adult murine CNS stem cells express aquaporin channels. Biol. Cell 2006, 98, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, G.-Q.; Li, Y.; Gu, Y.; Chen, X.-M.; Zhou, Y.; Zhao, S.-Z.; Shen, J. Beyond water channel: Aquaporin-4 in adult neurogenesis. Neurochem. Int. 2010, 56, 651–654. [Google Scholar] [CrossRef]
- Kong, H.; Sha, L.-L.; Fan, Y.; Xiao, M.; Ding, J.-H.; Wu, J.; Hu, G. Requirement of AQP4 for Antidepressive Efficiency of Fluoxetine: Implication in Adult Hippocampal Neurogenesis. Neuropsychopharmacology 2008, 34, 1263–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mack, A.F.; Wolburg, H. A Novel Look at Astrocytes: Aquaporins, ionic homeostasis, and the role of the microenvironment for regeneration in the CNS. Neuroscience 2012, 19, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Hu, G. Involvement of Aquaporin 4 in Astrocyte Function and Neuropsychiatric Disorders. CNS Neurosci. Ther. 2014, 20, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, W.; Barnett, M.H.; Prain, K.; Broadley, S.A. Molecular Pathogenesis of Neuromyelitis Optica. Int. J. Mol. Sci. 2012, 13, 12970–12993. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.; Fan, Y.; Xie, J.; Ding, J.; Sha, L.; Shi, X.; Sun, X.; Hu, G. AQP4 knockout impairs proliferation, migration and neuronal differentiation of adult neural stem cells. J. Cell Sci. 2008, 121, 4029–4036. [Google Scholar] [CrossRef] [Green Version]
- Liang, R.; Yong, S.; Huang, X.; Kong, H.; Hu, G.; Fan, Y. Aquaporin-4 Mediates the Suppressive Effect of Lipopolysaccharide on Hippocampal Neurogenesis. Neuroimmunomodulation 2016, 23, 309–317. [Google Scholar] [CrossRef]
- Lan, Y.-L.; Fang, D.-Y.; Zhao, J.; Ma, T.-H.; Li, S. A research update on the potential roles of aquaporin 4 in neuroinflammation. Acta Neurol. Belg. 2015, 116, 127–134. [Google Scholar] [CrossRef]
- Borsini, A.; Zunszain, P.A.; Thuret, S.; Pariante, C.M. The role of inflammatory cytokines as key modulators of neurogenesis. Trends Neurosci. 2015, 38, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Harlow, D.E.; Given, K.S.; Owens, G.P.; Macklin, W.B.; Bennett, J.L. Variable sensitivity to complement-dependent cytotoxicity in murine models of neuromyelitis optica. J. Neuroinflammation 2016, 13, 301. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Given, K.S.; Owens, G.P.; Macklin, W.B.; Bennett, J.L. Distinct patterns of glia repair and remyelination in antibody-mediated demyelination models of multiple sclerosis and neuromyelitis optica. Glia 2018, 66, 2575–2588. [Google Scholar] [CrossRef]
- Ikeshima-Kataoka, H. Neuroimmunological Implications of AQP4 in Astrocytes. Int. J. Mol. Sci. 2016, 17, 1306. [Google Scholar] [CrossRef] [PubMed]
- Ikota, H.; Iwasaki, A.; Kawarai, M.; Nakazato, Y. Neuromyelitis optica with intraspinal expansion of Schwann cell remyelination. Neuropathology 2009, 30, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.L.; Lam, C.; Kalluri, S.R.; Bs, P.S.; Ms, K.B.; DuPree, C.; Ms, M.G.; Case, D.; Antel, J.P.; Owens, G.P.; et al. Intrathecal pathogenic anti–aquaporin-4 antibodies in early neuromyelitis optica. Ann. Neurol. 2009, 66, 617–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Pinedo, U.; Sirerol-Piquer, M.S.; Durán-Moreno, M.; García-Verdugo, J.M.; Matias-Guiu, J. Alexander Disease Mutations Produce Cells with Coexpression of Glial Fibrillary Acidic Protein and NG2 in Neurosphere Cultures and Inhibit Differentiation into Mature Oligodendrocytes. Front. Neurol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Matías-Guiu, J.; Matías-Guiu, J.A.; Montero-Escribano, P.; Barcia, J.A.; Canales-Aguirre, A.A.; Mateos-Diaz, J.C.; Gómez-Pinedo, U. Particles Containing Cells as a Strategy to Promote Remyelination in Patients with Multiple Sclerosis. Front. Neurol. 2020, 11, 638. [Google Scholar] [CrossRef] [PubMed]
- Ojeda-Hernández, D.D.; Canales-Aguirre, A.A.; Matias-Guiu, J.; Gomez-Pinedo, U.; Mateos-Díaz, J.C. Potential of Chitosan and Its Derivatives for Biomedical Applications in the Central Nervous System. Front. Bioeng. Biotechnol. 2020, 8, 389. [Google Scholar] [CrossRef]
- Ader, M.; Tanaka, E.M. Modeling human development in 3D culture. Curr. Opin. Cell Biol. 2014, 31, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Salman, M.M.; Marsh, G.; Kusters, I.; Delincé, M.; Di Caprio, G.; Upadhyayula, S.; De Nola, G.; Hunt, R.; Ohashi, K.G.; Gray, T.; et al. Design and Validation of a Human Brain Endothelial Microvessel-on-a-Chip Open Microfluidic Model Enabling Advanced Optical Imaging. Front. Bioeng. Biotechnol. 2020, 8, 573775. [Google Scholar] [CrossRef] [PubMed]
- Aldewachi, H.; Al-Zidan, R.; Conner, M.; Salman, M. High-Throughput Screening Platforms in the Discovery of Novel Drugs for Neurodegenerative Diseases. Bioengineering 2021, 8, 30. [Google Scholar] [CrossRef]
- Del Palacio, J.P.; Díaz, C.; De La Cruz, M.; Annang, F.; Martín, J.; Pérez-Victoria, I.; González-Menéndez, V.; De Pedro, N.; Tormo, J.R.; Algieri, F.; et al. High-Throughput Screening Platform for the Discovery of New Immunomodulator Molecules from Natural Product Extract Libraries. J. Biomol. Screen. 2016, 21, 567–578. [Google Scholar] [CrossRef]
- Wingerchuk, D.M.; Banwell, B.; Bennett, J.L.; Cabre, P.; Carroll, W.; Chitnis, T.; De Seze, J.; Fujihara, K.; Greenberg, B.M.; Jacob, A.; et al. International consensus diagnostic criteria for neuromyelitis optica spectrum disorders. Neurology 2015, 85, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Jung, S.-W.; Kim, Y.; Park, Y.-J.; Han, K.; Oh, E.-J. Detection of Anti-Aquaporin-4 Antibodies in Neuromyelitis Optica: Comparison of Tissue-Based and Cell-Based Indirect Immunofluorescence Assays and ELISA. J. Clin. Lab. Anal. 2012, 26, 184–189. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Pinedo, U.; García-Ávila, Y.; Gallego-Villarejo, L.; Matías-Guiu, J.A.; Benito-Martín, M.S.; Esteban-García, N.; Sanclemente-Alamán, I.; Pytel, V.; Moreno-Jiménez, L.; Sancho-Bielsa, F.; et al. Sera from Patients with NMOSD Reduce the Differentiation Capacity of Precursor Cells in the Central Nervous System. Int. J. Mol. Sci. 2021, 22, 5192. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105192
Gómez-Pinedo U, García-Ávila Y, Gallego-Villarejo L, Matías-Guiu JA, Benito-Martín MS, Esteban-García N, Sanclemente-Alamán I, Pytel V, Moreno-Jiménez L, Sancho-Bielsa F, et al. Sera from Patients with NMOSD Reduce the Differentiation Capacity of Precursor Cells in the Central Nervous System. International Journal of Molecular Sciences. 2021; 22(10):5192. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105192
Chicago/Turabian StyleGómez-Pinedo, Ulises, Yolanda García-Ávila, Lucía Gallego-Villarejo, Jordi A. Matías-Guiu, María Soledad Benito-Martín, Noelia Esteban-García, Inmaculada Sanclemente-Alamán, Vanesa Pytel, Lidia Moreno-Jiménez, Francisco Sancho-Bielsa, and et al. 2021. "Sera from Patients with NMOSD Reduce the Differentiation Capacity of Precursor Cells in the Central Nervous System" International Journal of Molecular Sciences 22, no. 10: 5192. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105192