
Lactate Like Fluconazole Reduces Ergosterol Content in the Plasma Membrane and Synergistically Kills Candida albicans


Abstract
:1. Introduction
2. Results
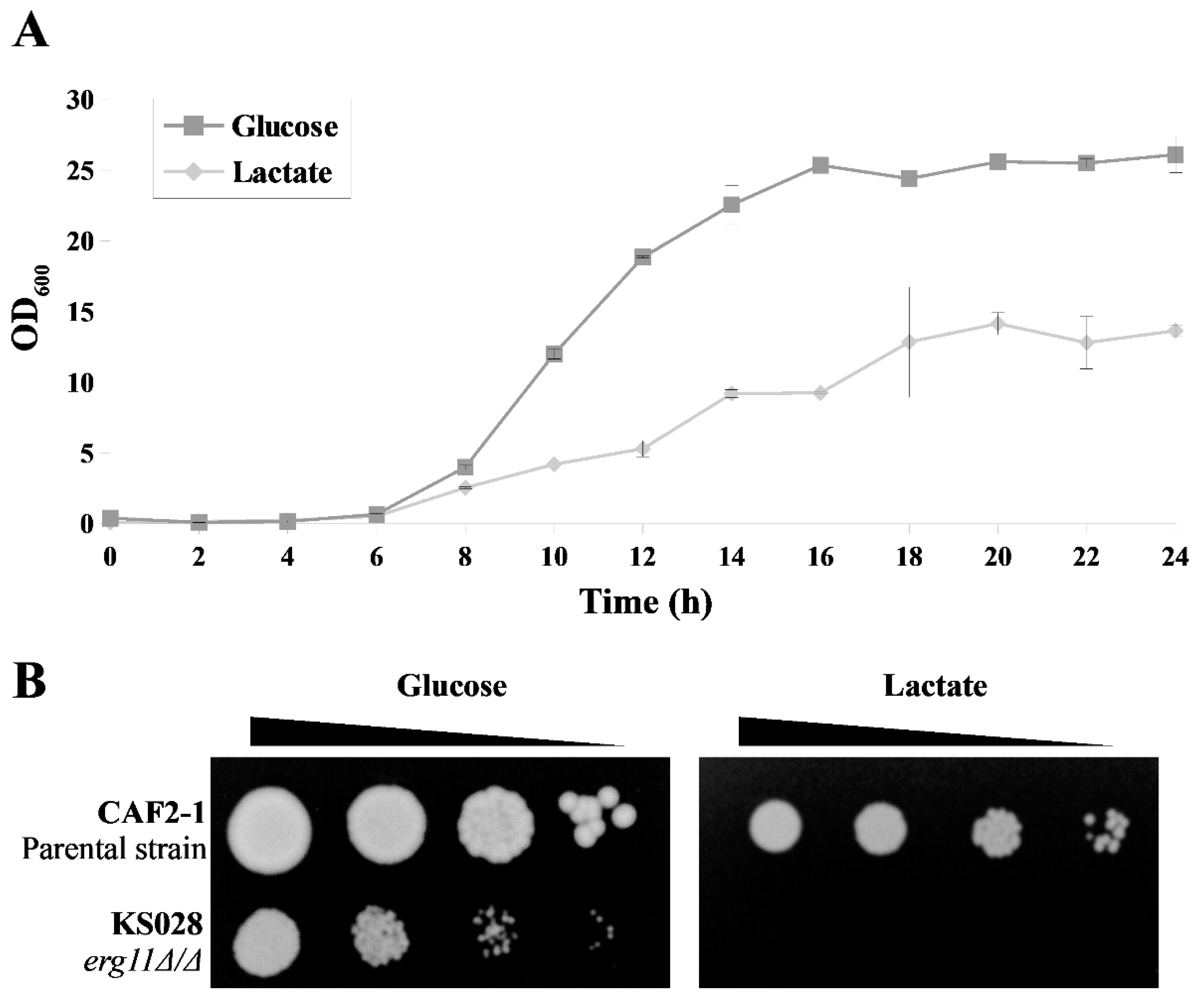
2.1. Lactate Affects the Resistance of C. albicans to Fluconazole Depending on Ergosterol—The First Observations
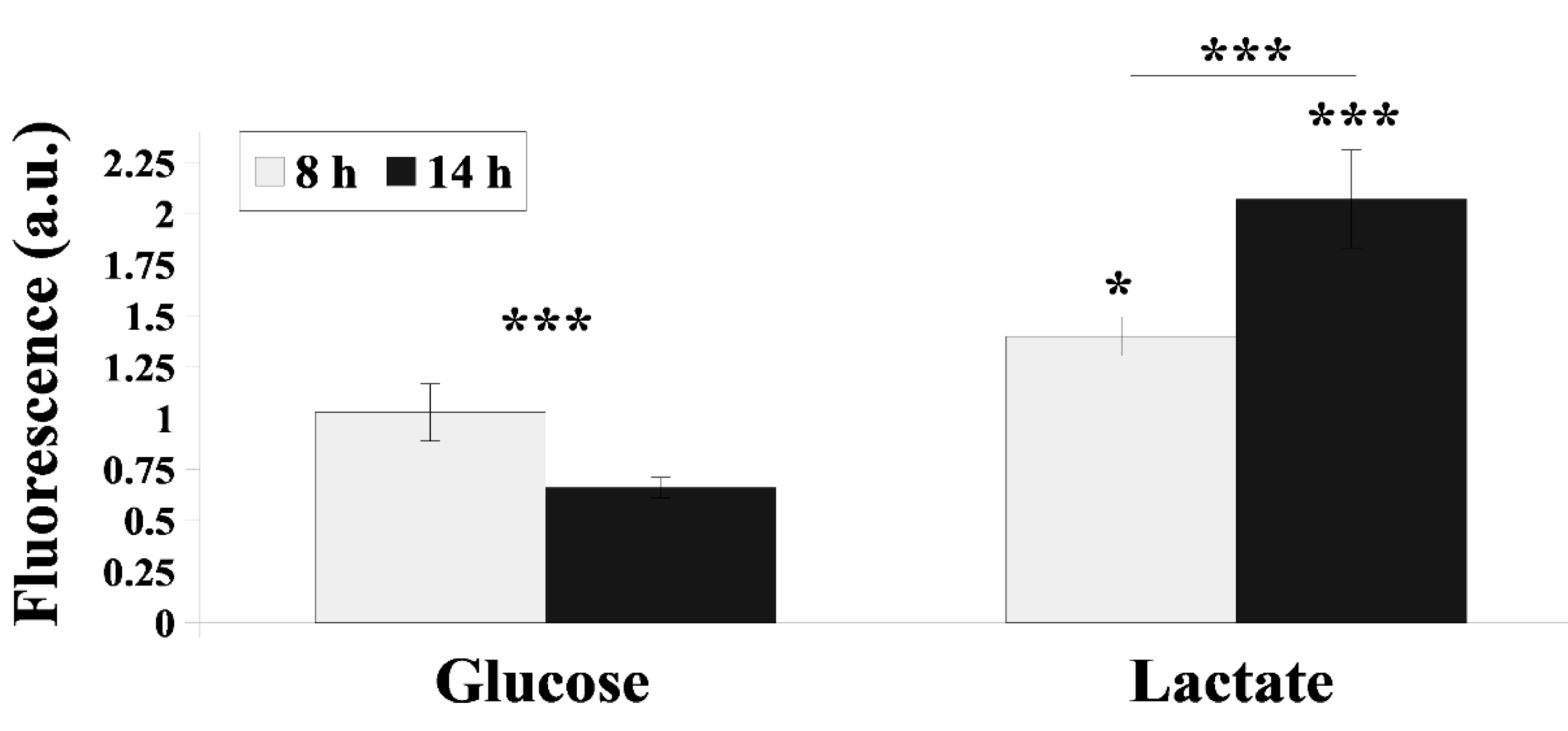
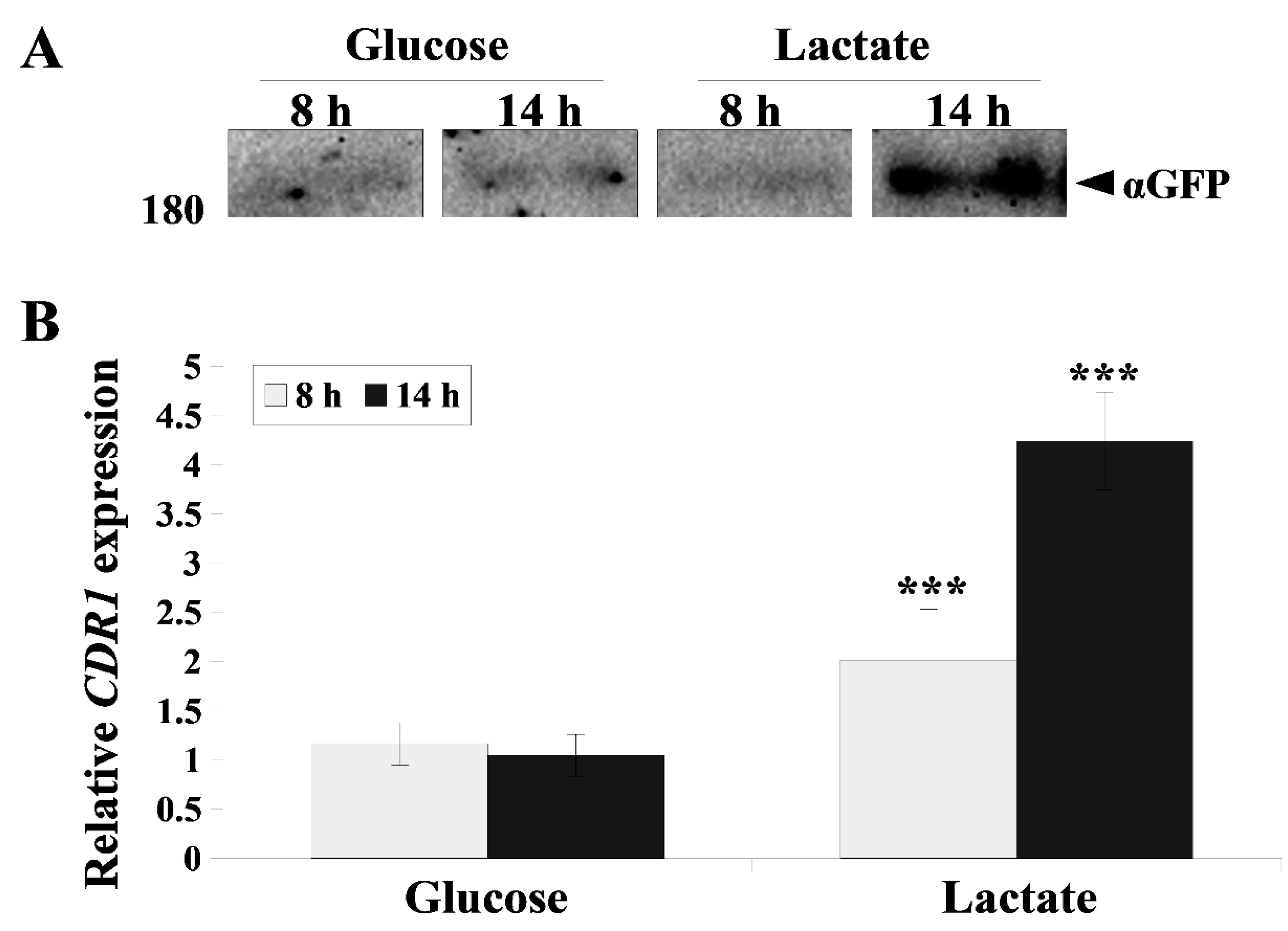
2.2. Cdr1 Transporter Activity and CDR1 Gene Expression Are More Efficiently Upregulated in the Presence Lactate Compared with Glucose
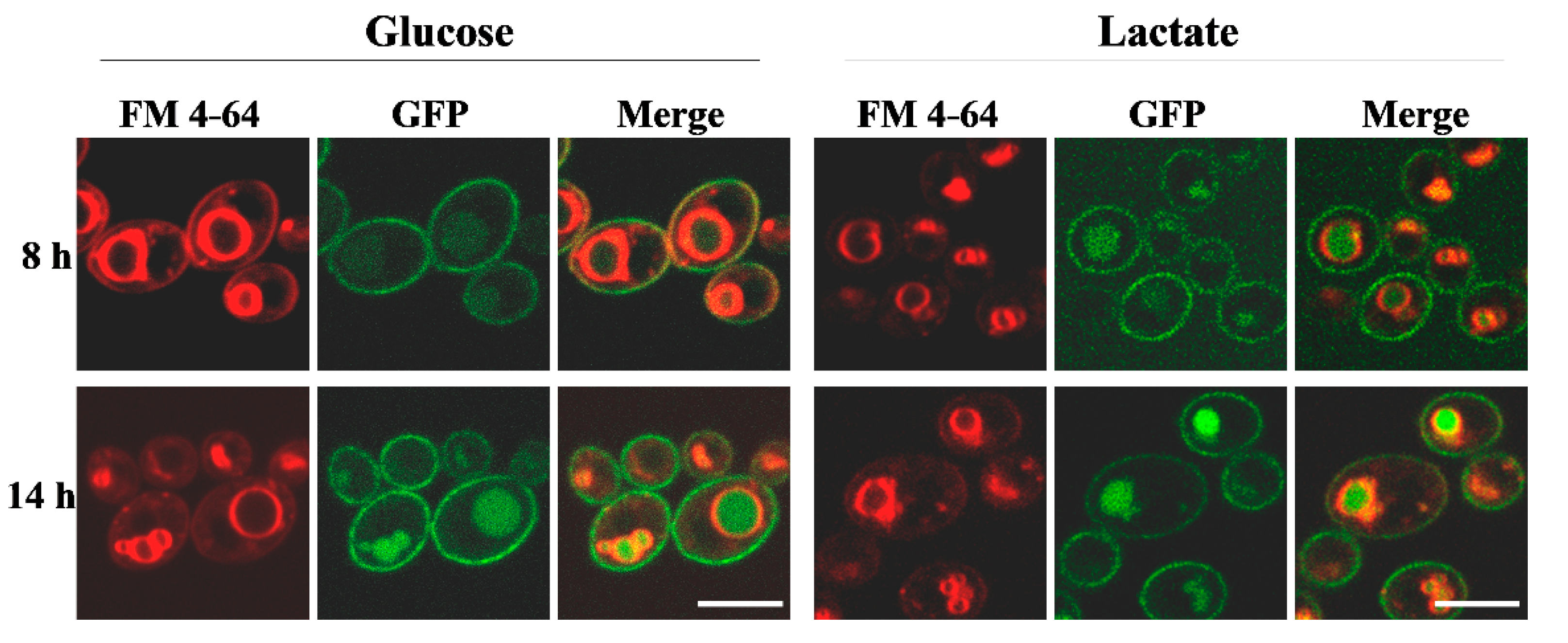
2.3. Lactate Accelerates Delocalization of Cdr1 Transporter from PM into Vacuoles
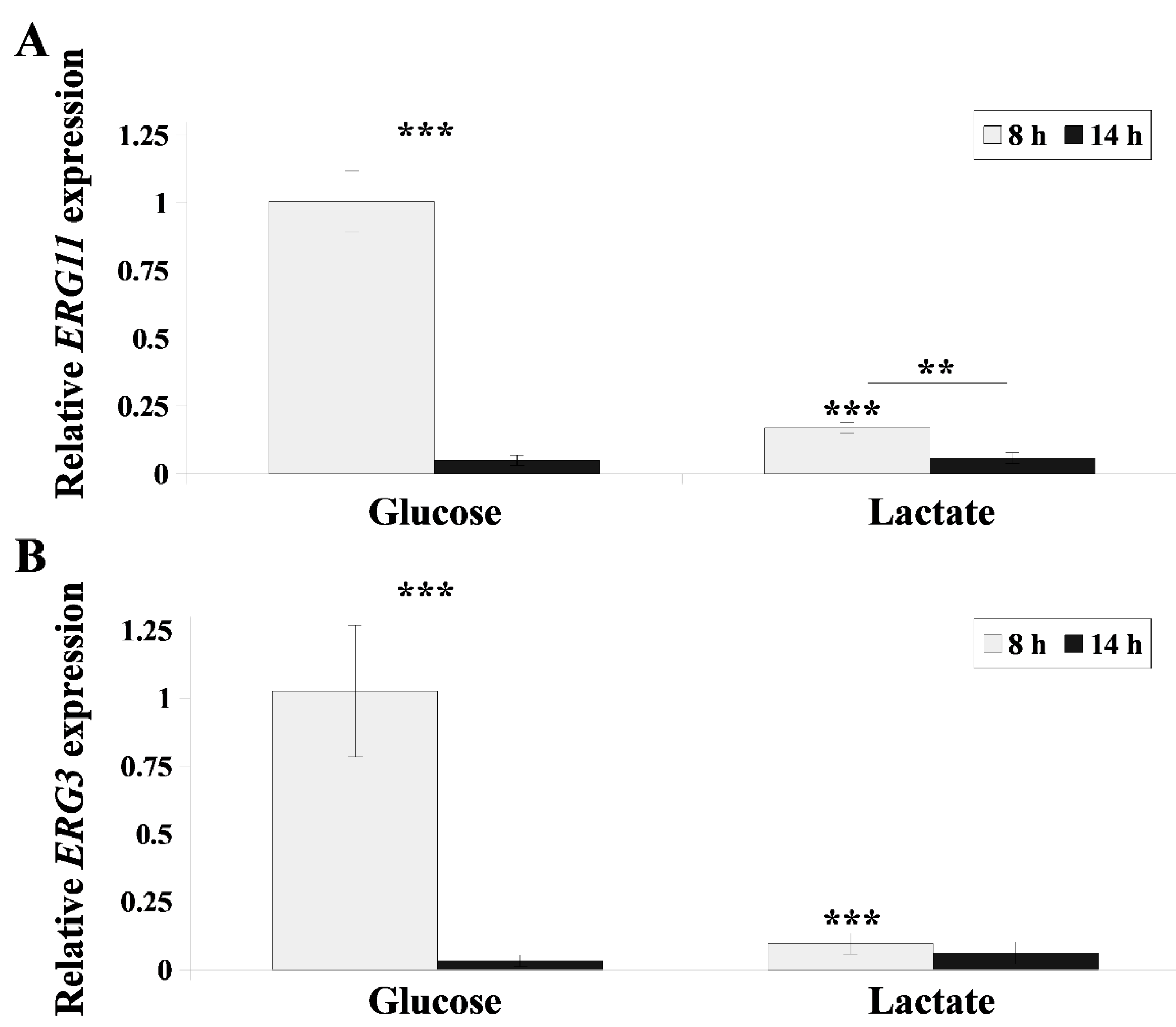
2.4. Lactate Inhibits Ergosterol Synthesis in C. albicans Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Strains and Growth Conditions
4.3. MIC50 Determination
4.4. Microscopic Studies
4.5. Western Blot
4.6. Real-Time PCR
4.7. Efflux Activity of Cdr1 Transporter
4.8. Phenotype Assay
4.9. PM Isolation and Sterol Analysis
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sobel, J. Vulvovaginal candidosis. Lancet 2007, 369, 1961–1971. [Google Scholar] [CrossRef]
- Ahmad, K.; Kokosar, J.; Guo, X.; Gu, Z.; Ishchuk, O.; Piskur, J. Genome structure and dynamics of the yeast pathogen Candida glabrata. FEMS Yeast Res. 2014, 14, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Brunke, S.; Hube, B. Two unlike cousins: Candida albicans and C. glabrata infection strategies. Cell. Microbiol. 2013, 15, 701–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, B.; FeErreira, C.; Alves, C.; Henriques, M.; Azeredo, J.; Silva, S. Vulvovaginal candidiasis: Epidemiology, microbiology and risk factors. Crit. Rev. Microbiol. 2016, 42, 905–927. [Google Scholar] [CrossRef] [Green Version]
- Sobel, J. Recurrent vulvovaginal candidiasis. Am. J. Obstet. Gynecol. 2016, 214, 15–21. [Google Scholar] [CrossRef]
- Boris, S.; Barbes, C. Role played by lactobacilli in controlling the population of vaginal pathogens. Microb. Infect. 2000, 2, 543–546. [Google Scholar] [CrossRef]
- Boskey, E.; Telsch, K.; Whaley, K.; Moench, T.; Cone, R. Acid production by vaginal flora in vitro is consistent with the rate and extent of vaginal acidification. Infect. Immun. 1999, 67, 5170–5175. [Google Scholar] [CrossRef] [Green Version]
- Owen, D.; Katz, D. A vaginal fluid simulant. Contraception 1999, 59, 91–95. [Google Scholar] [CrossRef]
- O’Hanlon, D.; Moench, T.; Cone, R. Vaginal pH and microbicidal lactic acid when lactobacilli dominate the microbiota. PLoS ONE 2013, 8, e80074. [Google Scholar] [CrossRef]
- Matsubara, V.; Wang, Y.; Bandara, H.; Mayer, M.; Samaranayake, L. Probiotic lactobacilli inhibit early stages of Candida albicans biofilm development by reducing their growth, cell adhesion, and filamentation. Appl. Microbiol. Biotechnol. 2016, 100, 6415–6426. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Xu, S.; He, Y.; Deng, G.; Sheng, H.; Huang, X.; Ouyang, C.; Zhou, H. Diverse vaginal microbiomes in reproductive-age women with vulvovaginal candidiasis. PLoS ONE 2013, 8, e79812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zangl, I.; Pap, I.; Aspöck, C.; Schüller, C. The role of Lactobacillus species in the control of Candida via biotrophic interactions. Microb. Cell 2020, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Alves, R.; Mota, S.; Silva, S.; Rodrigues, C.; Brown, A.; Henriques, M.; Casal, M.; Paiva, S. The carboxylic acid transporters Jen1 and Jen2 affect the architecture and fluconazole susceptibility of Candida albicans biofilm in the presence of lactate. Biofouling 2017, 33, 943–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, A.; Pedro, N.; Salazar, S.; Mira, N. Effect of acetic acid and lactic acid at low pH in growth and azole resistance of Candida albicans and Candida glabrata. Front. Microbiol. 2019, 9, 3265. [Google Scholar] [CrossRef] [Green Version]
- Ene, I.; Adya, A.; Wehmeier, S.; Brand, A.; MacCallum, D.; Gow, N.; Brown, A. Host carbon surces modulate cell wall architecture, drug resistance and virulence in a fungal pathogen. Cell Microbiol. 2012, 14, 1319–1335. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.; Lewis, M. Pathogenesis and treatment of oral candidosis. J. Oral Microbiol. 2011, 1, 5771. [Google Scholar] [CrossRef] [Green Version]
- Gray, K.; Palacios, D.; Dailey, I.; Endo, M.; Uno, B.; Wilcock, B.; Burke, M. Amphotericin primarily kills yeast by simply binding ergosterol. Proc. Natl. Acad. Sci. USA 2012, 109, 2234–2239. [Google Scholar] [CrossRef] [Green Version]
- Suchodolski, J.; Krasowska, A. Fructose induces fluconazole resistance in Candida albicans through activation of Mdr1 and Cdr1 transporters. Int. J. Mol. Sci. 2021, 22, 2127. [Google Scholar] [CrossRef] [PubMed]
- Lyons, C.; White, T. Transcriptional analyses of antifungal drug resistance in Candida albicans. Antimicrob. Agents Chemother. 2000, 44, 2296–2303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mira, N.; Teixeira, M.; Sa-Correia, I. Adaptive response and tolerance to weak acids in Saccharomyces cerevisiae: A genome-wide view. Omics J. Integr. Biol. 2010, 14, 525–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchodolski, J.; Muraszko, J.; Bernat, P.; Krasowska, A. A crucial role for ergosterol in plasma membrane composition, localisation, and activity of Cdr1p and H+-ATPase in Candida albicans. Microorganisms 2019, 7, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchodolski, J.; Muraszko, J.; Korba, A.; Bernat, P.; Krasowska, A. Lipid composition and cell surface hydrophobicity of Candida albicans influence the efficacy of fluconazole–gentamicin treatment. Yeast 2020, 37, 117–129. [Google Scholar] [CrossRef]
- Suchodolski, J.; Derkacz, D.; Bernat, P.; Krasowska, A. Capric acid secreted by Saccharomyces boulardii influences the susceptibility of Candida albicans to fluconazole and amphotericin B. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Moosa, M.; Sobel, J.; Elhalis, H.; Du, W.; Akins, R. Fungicidal activity of fluconazole against Candida albicans in a synthetic vagina-simulative medium. Antimicrob. Agents Chemother. 2004, 48, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Kasper, L.; Miramon, P.; Jablonowski, N.; Wisgott, S.; Wilson, D.; Brunke, S.; Hube, B. Antifungal activity of clotrimazole against Candida albicans depends on carbon sources, growth phase and morphology. J. Med. Microbiol. 2015, 64, 714–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Ende, M.; Wijnants, S.; Van Dijck, P. Sugar sensing and signaling in Candida albicans and Candida glabrata. Front. Microbiol. 2019, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sandai, D.; Yin, Z.; Selway, L.; Stead, D.; Walker, J.; Leach, M.D.; Bohovych, I.; Ene, I.V.; Kastora, S.; Budge, S.; et al. The Evolutionary Rewiring of Ubiquitination Targets Has Reprogrammed the Regulation of Carbon Assimilation in the Pathogenic Yeast Candida albicans. MBio 2012, 3, e00495-12. [Google Scholar] [CrossRef] [Green Version]
- Karababa, M.; Coste, A.; Rognon, B.; Bille, J.; Sanglard, D. Comparison of gene expression profiles of Candida albicans azole-resistant clinical isolates and laboratory strains exposed to drugs inducing multidrug transporters. Antimicrob. Agents Chemother. 2004, 48, 3064–3079. [Google Scholar] [CrossRef] [Green Version]
- Niimi, M.; Niimi, K.; Takano, Y.; Holmes, A.; Fischer, F.; Uehara, Y.; Cannon, R. Regulated overexpression of CDR1 in Candida albicans confers multidrug resistance. J. Antimicrob. Chemother. 2004, 54, 999–1006. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Harry, J.; Eastman, R.; Oliver, B.; White, T. The Candida albicans Lanosterol 14-α-Demethylase (ERG11) Gene Promoter Is Maximally Induced after Prolonged Growth with Antifungal Drugs. Antimicrob. Agents Chemother. 2004, 48, 1136–1144. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Xia, J.; Nie, K.; Wang, F.; Deng, L. Outline of the biosynthesis and regulation of ergosterol in yeast. World J. Microbiol. Biotechnol. 2019, 35, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Neugebauer, T.; Zill, P.; Lass-Flörl, C.; Bracher, F.; Binder, U. Sterol composition of clinically relevant Mucorales and changes resulting from posaconazole treatment. Molecules 2018, 23, 1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, C.; Parker, J.; Bader, O.; Weig, M.; Gross, U.; Warrilow, A.; Rolley, N.; Kelly, D.; Kelly, S. Identification and characterization of four azole-resistant erg3 mutants of Candida albicans. Antimicrob. Agents Chemother. 2010, 54, 4527–4533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, L.; Han, J.; Ferreira, R.; Lolić, P.; Borchers, C.; Finlay, B. Effect of antibiotic treatment on the intestinal metabolome. Antimicrob. Agents Chemother. 2011, 55, 1494–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barko, P.; McMichael, M.; Swanson, K.; Williams, D. The gastrointestinal microbiome: A review. J. Vet. Int. Med. 2018, 32, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Ehrström, S.; Daroczy, K.; Rylander, E.; Samuelsson, C.; Johannesson, U.; Anzén, B.; Påhlson, C. Lactic acid bacteria colonization and clinical outcome after probiotic supplementation in conventionally treated bacterial vaginosis and vulvovaginal candidiasis. Microb. Infect. 2010, 12, 691–699. [Google Scholar] [CrossRef]
- Fonzi, W.; Irwin, M. Isogenic Strain Construction and Gene Mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar] [CrossRef]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Susceptibilities of Candida albicans Multidrug Transporter Mutants to Various Antifungal Agents and Other Metabolic Inhibitors. Antimicrob. Agents Chemother. 1996, 40, 2300–2305. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Cloning of Candida albicans genes conferring resistance to azole antifungal agents: Characterization of CDR2, a new multidrug ABC transporter gene. Microbiology 1997, 143, 405–416. [Google Scholar] [CrossRef] [Green Version]
- Szczepaniak, J.; Łukaszewicz, M.; Krasowska, A. Estimation of Candida albicans ABC Transporter Behavior in Real-Time via Fluorescence. Front. Microbiol. 2015, 6, 1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vida, T.; Emr, S. A new vital stain for visualizing vacuolar membrane dynamics and endocytosis in yeast. J. Cell Biol. 1995, 128, 779–792. [Google Scholar] [CrossRef] [PubMed]
- Szczepaniak, J.; Cieślik, W.; Romanowska, A.; Musioł, R.; Krasowska, A. Blocking and dislocation of Candida albicans Cdr1p transporter by styrylquinolines. Int. J. Antimicrob. Agents 2017, 50, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Suchodolski, J.; Derkacz, D.; Muraszko, J.; Panek, J.J.; Jezierska, A. Fluconazole and Lipopeptide Surfactin Interplay During Candida albicans Plasma Membrane and Cell Wall Remodeling Increases Fungal Immune System Exposure. Pharmaceutics 2020, 12, 314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; MacKenzie, A.; Girnun, G.; Del Poeta, M. Analysis of sphingolipids, sterols, and phospholipids in human pathogenic Cryptococcus strains. J. Lipid Res. 2017, 58, 2017–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain C. albicans | Medium | Fluconazole | Brefeldin A | Fluphenazine |
|---|---|---|---|---|
| WT | YPD | 2 | 16 | 125 |
| YPL | 0.5 | 32 | >250 | |
| cdr1Δ | YPD | 0.25 | 4 | 125 |
| YPL | 0.063 | 4 | 250 | |
| cdr2Δ | YPD | 2 | 16 | 125 |
| YPL | 0.5 | 32 | >250 | |
| cdr1Δcdr2Δ | YPD | 0.25 | 4 | 62.5 |
| YPL | 0.063 | 4 | 62.5 |
| Glucose | Lactate | |||
|---|---|---|---|---|
| 8 h | 14 h | 8 h | 14 h | |
| Ergosterol | 39.1 ± 2.3 | 65.2 ± 1.1 *** | 22.9 ± 0.3 ** | 17.8 ± 4.8 *** |
| Lanosterol | 8.5 ± 0.2 | 5.4 ± 0.6 ** | 5.9 ± 0.5 ** | 5.1 ± 0.8 ** |
| 4-methylfecosterol | ND | ND | 3.9 ± 0.5 | 3.9 ± 0.2 |
| Ergosta-5,7-dienol | ND | ND | 1.1 ± 0.1 | 0.8 ± 0.2 |
| 4,4-dimethylcholesta-8,24-dien-3b-ol | ND | ND | 1.3 ± 0.3 | 1.1 ± 0.1 |
| Total sterols | 47.6 ± 2.5 | 70.6 ± 1.7 *** | 35.1 ± 1.7 | 28.7 ± 6.1 ** |
| Strain | Genotype | Reference |
|---|---|---|
| CAF2-1 | ura3Δ::imm434/URA3 | [37] |
| DSY448 | ura3∆::imm434/ura3∆::imm434 cdr1∆::hisG/cdr1∆::hisG-URA3-hisG | [38] |
| DSY653 | ura3∆::imm434/ura3∆::imm434 cdr2∆::hisG/cdr2∆::hisG-URA3-hisG | [39] |
| DSY654 | ura3∆::imm434/ura3∆::imm434 cdr1∆::hisG/cdr1∆::hisG cdr2∆::hisG/cdr2∆::hisG-URA3-hisG | [39] |
| ASCa1 | ura3Δ::imm434/ura3Δ::imm434 CDR1/CDR1-yEGFP-URA3 | [40] |
| KS028 | ura3Δ::imm434/ura3Δ::imm434 erg11Δ::SAT1-FLIP/erg11Δ::FRT | [21] |
| Primer | Sequence 5′–3′ |
|---|---|
| RDN18F | AGAAACGGCTACCACATCCAA |
| RDN18R | GGGCCCTGTATCGTTATTTATTGT |
| ERG11F | TTTGGTGGTGGTAGACATA |
| ERG11R | GAACTATAATCAGGGTCAGG |
| ERG3F | CCATCATGAATCATGACAGTCC |
| ERG3R | TGCTTCTCATGCTTTCCATC |
| CDR1F | TTTAGCCAGAACTTTCACTCATGATT |
| CDR1R | TATTTATTTCTTCATGTTCATATGGATTGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suchodolski, J.; Muraszko, J.; Bernat, P.; Krasowska, A. Lactate Like Fluconazole Reduces Ergosterol Content in the Plasma Membrane and Synergistically Kills Candida albicans. Int. J. Mol. Sci. 2021, 22, 5219. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105219
Suchodolski J, Muraszko J, Bernat P, Krasowska A. Lactate Like Fluconazole Reduces Ergosterol Content in the Plasma Membrane and Synergistically Kills Candida albicans. International Journal of Molecular Sciences. 2021; 22(10):5219. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105219
Chicago/Turabian StyleSuchodolski, Jakub, Jakub Muraszko, Przemysław Bernat, and Anna Krasowska. 2021. "Lactate Like Fluconazole Reduces Ergosterol Content in the Plasma Membrane and Synergistically Kills Candida albicans" International Journal of Molecular Sciences 22, no. 10: 5219. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22105219