
Trichoderma-Based Biopreparation with Prebiotics Supplementation for the Naturalization of Raspberry Plant Rhizosphere


Abstract
:1. Introduction
2. Results
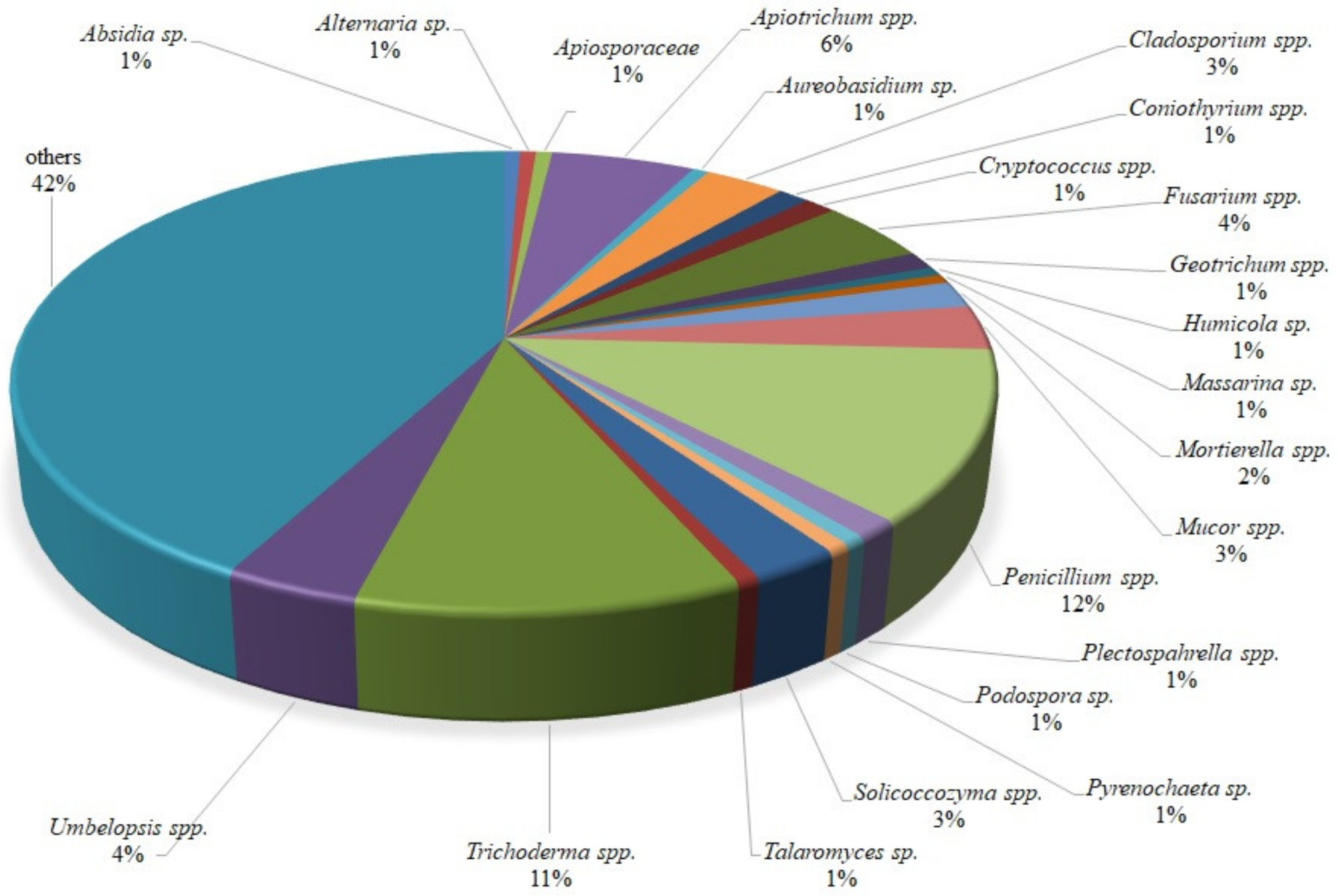
2.1. Isolates Description and Characterization
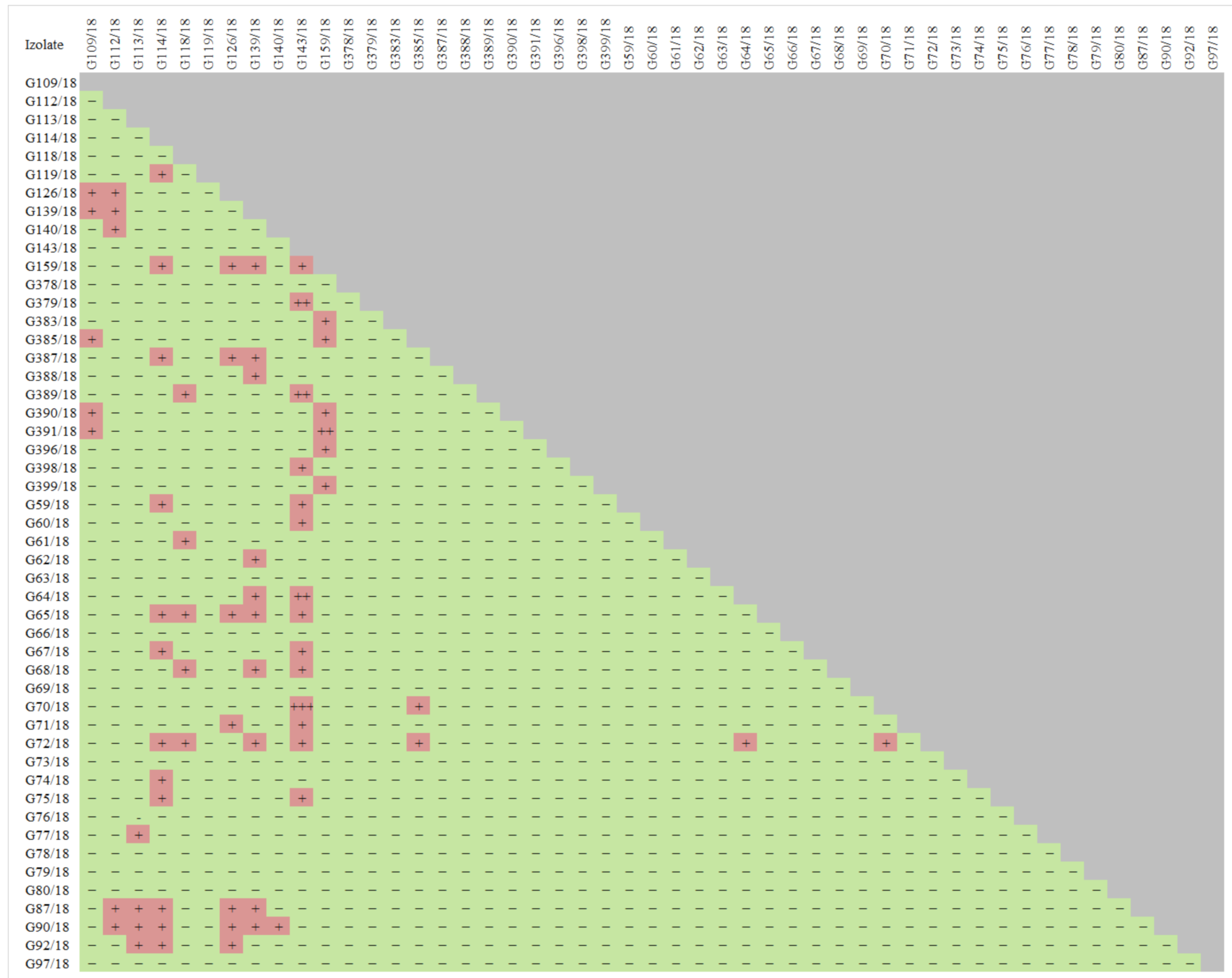
2.1.1. Antagonists Selection
2.1.2. Enzymatic Activity of Selected Trichoderma Isolates
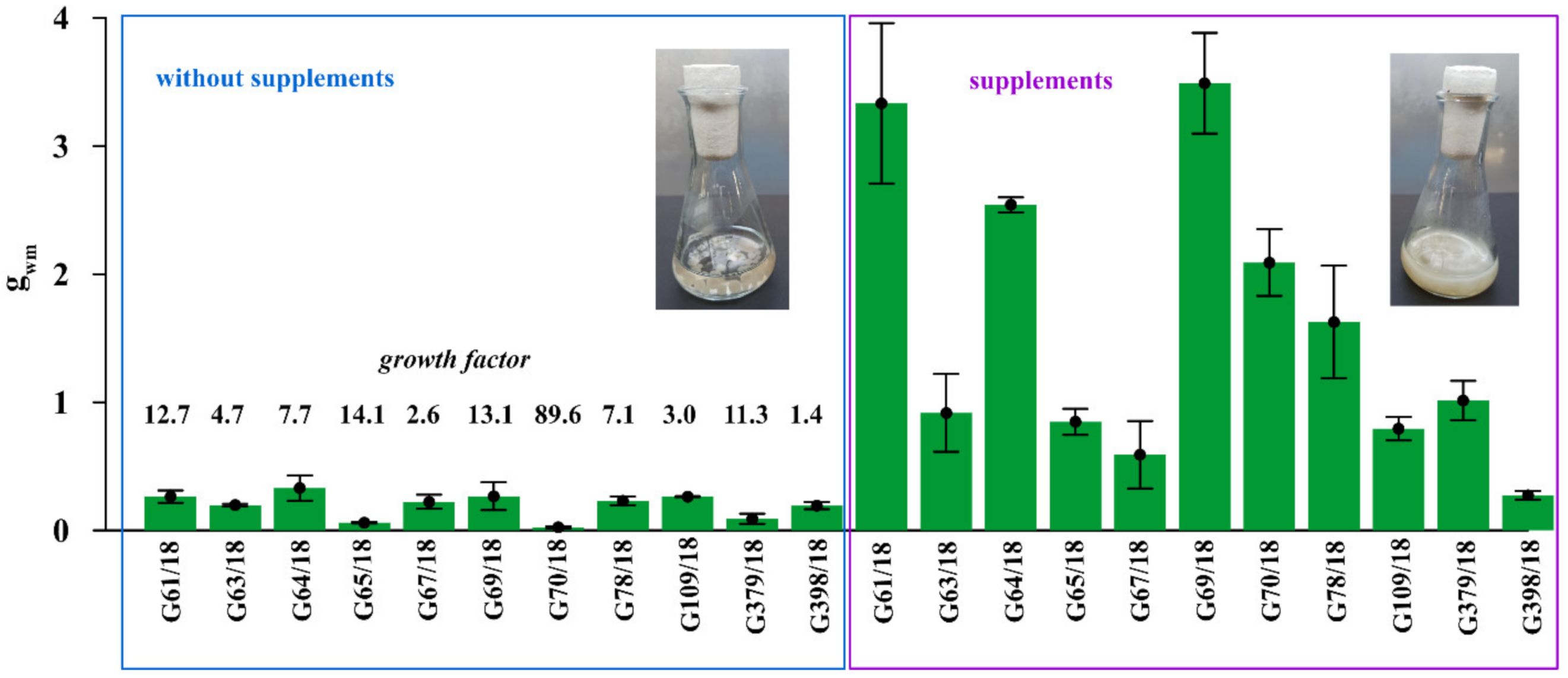
2.1.3. Effect of Supplement Addition on Trichoderma Isolate Growth
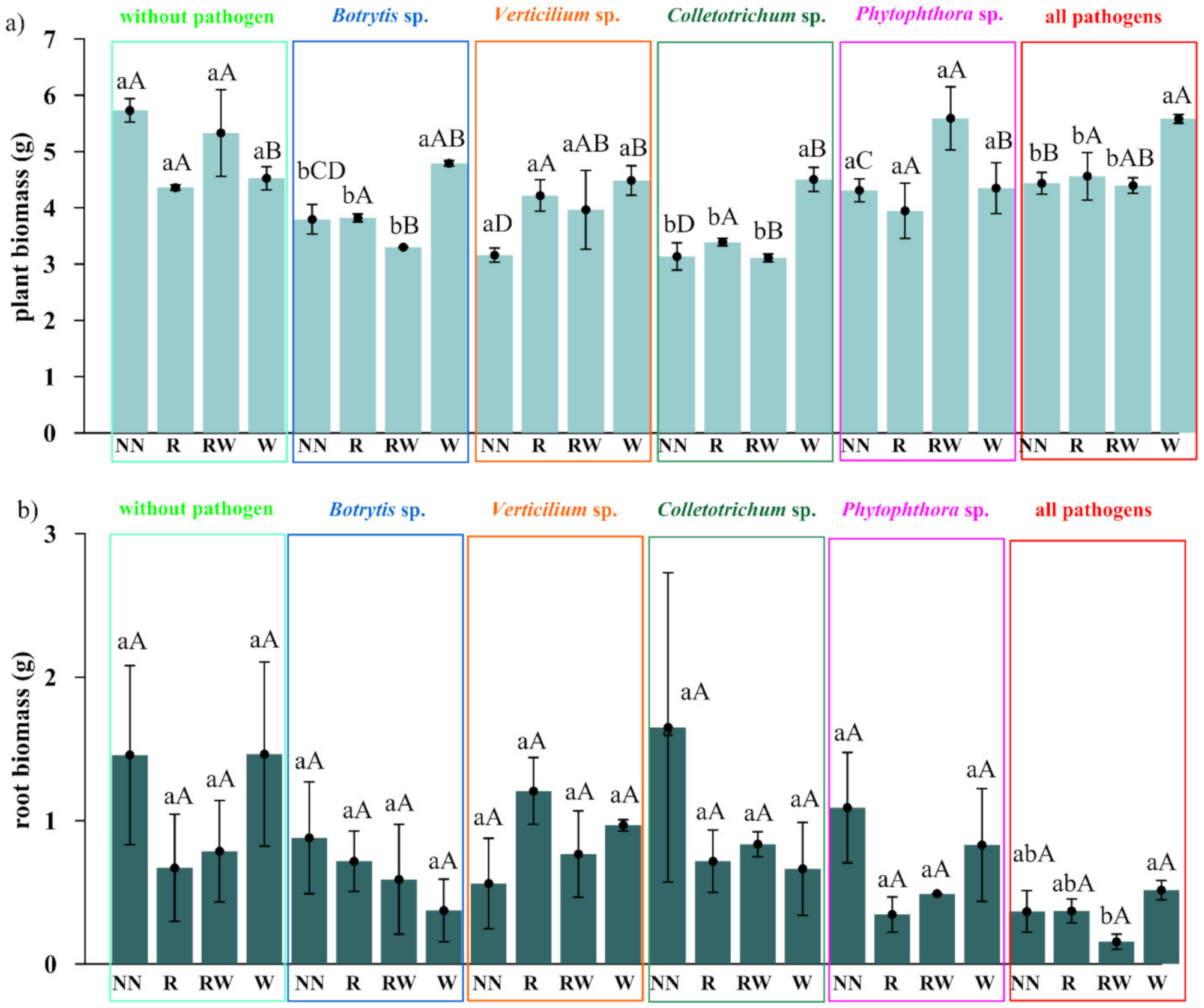
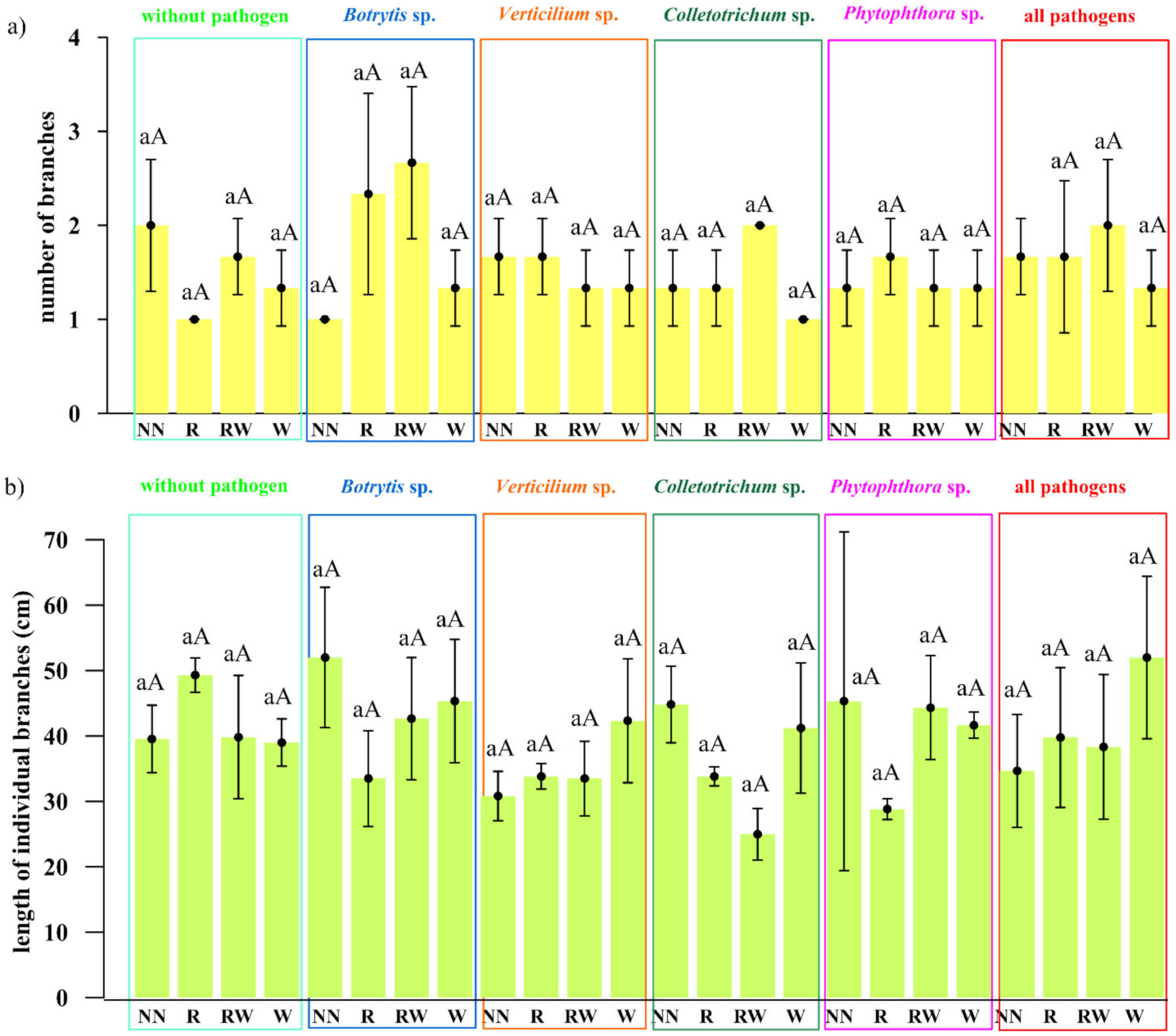
2.2. The Early Effect of the Trichoderma Isolates on Raspberry and Soil Properties
2.3. Laboratory-Scale Biopreparation Development
2.3.1. Trichoderma Sporulation Optimization
2.3.2. Formulations
3. Discussion
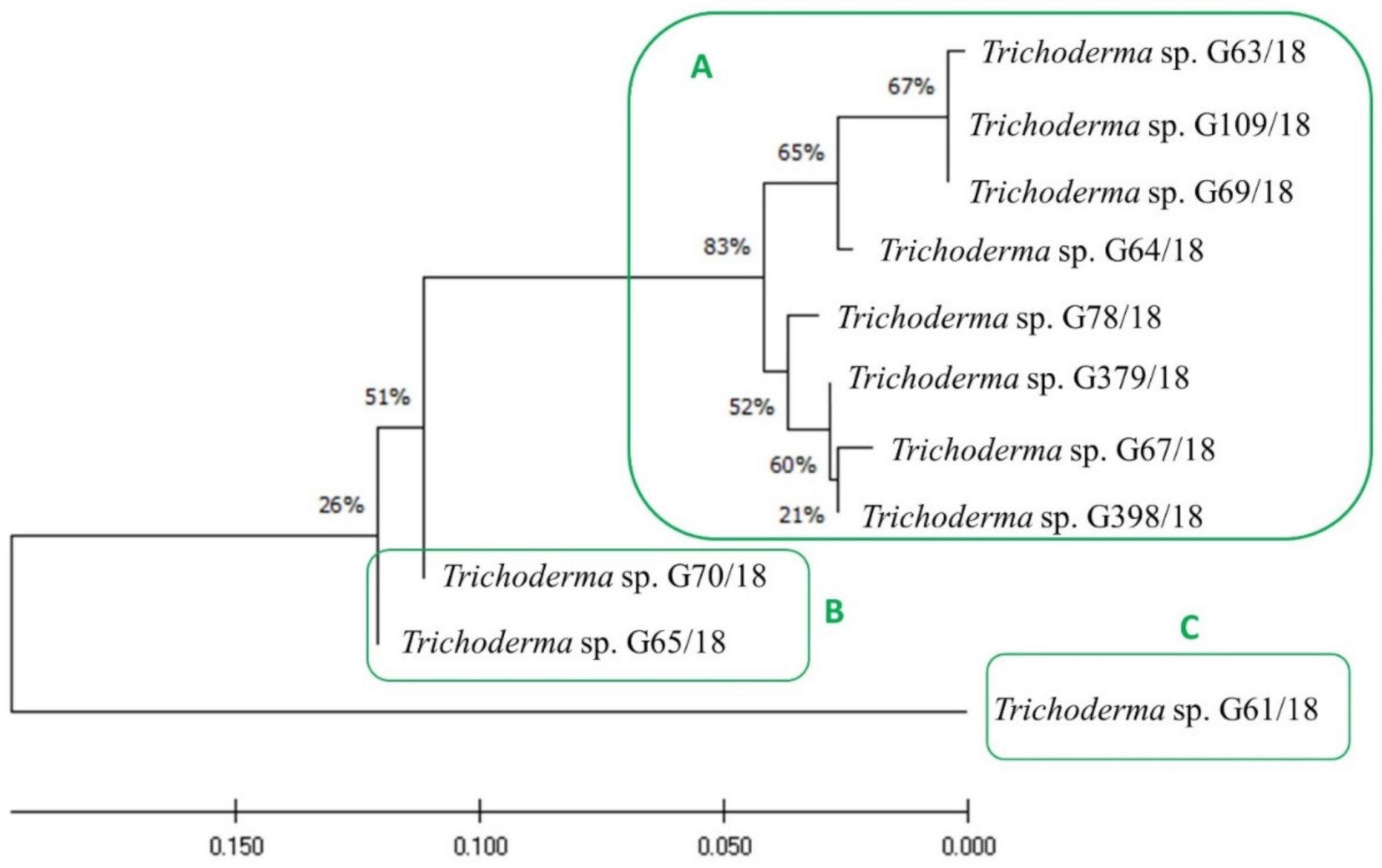
3.1. Fungal Consortium Selection for Biopreparation
3.2. Evaluation of the Influence of the Promptly Selected Trichoderma-Based Consortium on Raspberry in the Pot Experiment
3.3. The Development of the Biopreparation Targeted at Raspberry Naturalization
4. Materials and Methods
4.1. Isolates Description and Characterization
4.1.1. Isolation and Identification
4.1.2. Antagonistic Abilities of Raspberry Fungal Isolates against Phytopathogens
4.1.3. Enzymatic Activity of Selected Trichoderma Isolates
4.1.4. Effect of Supplement Addition on Trichoderma Isolate Growth
4.2. Effect of the Trichoderma Isolates on Raspberry Growth and Soil Properties
4.2.1. Phytotron Raspberry Pot Experiment Set up—Pathosystems and Naturalization Strategies
4.2.2. Plant and Soil Analyses
4.3. Laboratory-Scale Biopreparation Development
4.3.1. Trichoderma Sporulation Optimization
4.3.2. Solid Formulations—Powder and Pellet
4.3.3. Dissolvable Formulations—Powder and Gel
4.4. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- OECD-FAO. Agricultural Outlook 2019–2028. Available online: https://www.oecd-ilibrary.org/agriculture-and-food/oecd-fao-agricultural-outlook-2019-2028_agr_outlook-2019-en (accessed on 28 May 2021).
- Oszust, K.; Frąc, M. First report on the microbial communities of the wild and planted raspberry rhizosphere—A statement on the taxa, processes and a new indicator of functional diversity. Ecol. Ind. 2020, 121, 107117. [Google Scholar] [CrossRef]
- Malarczyk, D.; Panek, J.; Frac, M. Alternative molecular-based diagnostic methods of plant pathogenic fungi affecting berry crops-a review. Molecules 2019, 24, 1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oszust, K.; Cybulska, J.; Frąc, M. How do Trichoderma genus fungi win a nutritional competition battle against soft fruit pathogens? A report on niche overlap nutritional potentiates. Int. J. Mol. Sci. 2020, 21, 4235. [Google Scholar] [CrossRef] [PubMed]
- Regulation EC. Council Regulation (EC) No 834/2007. Available online: https://eur-lex.europa.eu/eli/reg/2007/834/oj (accessed on 28 May 2021).
- Communication, EU Biodiversity Strategy for 2030. COM(2020)380. Bringing Nature Back into Our Lives. Available online: https://eur-lex.europa.eu/legal-content/PL/TXT/PDF/?uri=CELEX:52020DC0380&from=EN (accessed on 28 May 2021).
- Montanarella, L.; Panagos, P. The relevance of sustainable soil management within the European Green Deal. Land Use Policy 2021, 100, 104950. [Google Scholar] [CrossRef]
- High Level Panel of Experts on Food Security and Nutrition of the Committee on World Food Security. Agroecological and Other Innovative Approaches for Sustainable Agriculture and Food Systems That Enhance Food Security and Nutrition. 2019. HLPE Report 14. Available online: http://www.fao.org/3/ca5602en/ca5602en.pdf (accessed on 28 May 2021).
- van der Heijden, M.G.; Hartmann, M. Networking in the plant microbiome. PLoS Biol. 2016, 14, e1002378. [Google Scholar] [CrossRef]
- Szczałba, M.; Kopta, T.; Gąstoł, M.; Sękara, A. Comprehensive insight into arbuscular mycorrhizal fungi, Trichoderma spp. and plant multilevel interactions with emphasis on biostimulation of horticultural crops. J. Appl. Microbiol. 2019, 127, 630–647. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Sarma, B.K.; Singh, H.B.; Upadhyay, R.S. Chapter 40—Trichoderma: A silent worker of plant rhizosphere. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 533–542. [Google Scholar]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The “secrets” of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Sharma, S.; Kour, D.; Rana, K.L.; Dhiman, A.; Thakur, S.; Thakur, P.; Thakur, S.; Thakur, N.; Sudheer, S.; Yadav, N.; et al. Trichoderma: Biodiversity, ecological significances, and industrial applications. In Recent Advancement in White Biotechnology through Fungi Volume 1: Diversity and Enzymes Perspectives; Yadav, A.N., Mishra, S., Singh, S., Gupta, A., Eds.; Springer Nature Switzerland AG: Cham, Switzerland, 2019; pp. 85–120. [Google Scholar]
- Sharma, A.K.; Sharma, P. Trichoderma. Host Pathogen Interactions and Applications; Springer: Singapore, 2020. [Google Scholar]
- Jaroszuk-Ściseł, J.; Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Majewska, M.; Hanaka, A.; Tyśkiewicz, K.; Pawlik, A.; Janusz, G. Phytohormones (auxin, gibberellin) and ACC deaminase in vitro synthesized by the mycoparasitic Trichoderma DEMTkZ3A0 strain and changes in the level of auxin and plant resistance markers in wheat seedlings inoculated with this strain conidia. Int. J. Mol. Sci. 2019, 20, 4923. [Google Scholar] [CrossRef] [Green Version]
- Oszust, K.; Pawlik, A.; Janusz, G.; Ziemiński, K.; Cyran, M.; Siczek, A.; Gryta, A.; Bilińska-Wielgus, N.; Frąc, M. Characterization and influence of a multi-enzymatic biopreparation for biogas yield enhancement. Bioresources 2017, 12, 6187–6206. [Google Scholar] [CrossRef] [Green Version]
- Oszust, K.; Frąc, M. Apple pomace microbiome carrying fungal load against phytopathogens—Considerations regarding application in agriculture and horticulture. Bioresources 2020, 15, 945–966. [Google Scholar] [CrossRef]
- Zin, N.A.; Badaluddin, N.A. Biological functions of Trichoderma spp. for agriculture applications. Ann. Agric. Sci. 2020, 65, 168–178. [Google Scholar] [CrossRef]
- Pylak, M.; Oszust, K.; Frąc, M. Review report on the role of bioproducts, biopreparations, biostimulants and microbial inoculants in organic production of fruit. Rev. Environ. Sci. Biotechnol. 2019, 18, 597–616. [Google Scholar] [CrossRef] [Green Version]
- Al-Ani, L.K.T. A Patent survey of Trichoderma spp. (from 2007 to 2017). In Intellectual Property Issues in Microbiology; Singh, H.B., Keswani, C., Singh, S.P., Eds.; Springer: Singapore, 2019; pp. 163–192. [Google Scholar]
- Pylak, M.; Oszust, K.; Frąc, M. Searching for new beneficial bacterial isolates of wild raspberries for biocontrol of phytopathogens-antagonistic properties and functional characterization. Int. J. Mol. Sci. 2020, 21, 9361. [Google Scholar] [CrossRef]
- Sahai, P.; Kumar, V. Carriers and their role in plant agrosystem. In Probiotics and Plant Health; Kumar, V., Kumar, M., Sharma, S., Prasad, R., Eds.; Springer: Singapore, 2017; pp. 291–315. [Google Scholar]
- Fikri, A.S.I.; Rahman, I.A.; Nor, N.S.M.; Hamzah, A. Isolation and identification of local bacteria endophyte and screening of its antimicrobial property against pathogenic bacteria and fungi. AIP Conf. Proc. 2018, 140, 020072. [Google Scholar] [CrossRef]
- Santhanam, R.; Weinhold, A.; Goldberg, J.; Oh, Y.; Baldwin, I.T. Native root-associated bacteria rescue a plant from a sudden-wilt disease that emerged during continuous cropping. Proc. Natl. Acad. Sci. USA 2015, 112, E5013–E5020. [Google Scholar] [CrossRef] [Green Version]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Cheng, D.; Song, J.; Xie, M.; Song, D. The bidirectional relationship between host physiology and microbiota and health benefits of probiotics: A review. Trends Food. Sci. Technol. 2019, 9, 426–435. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathog. 2018, 10, 21. [Google Scholar] [CrossRef]
- Hartman, K.; van der Heijden, M.G.; Wittwer, R.A.; Banerjee, S.; Walser, J.C.; Schlaeppi, K. Cropping practices manipulate abundance patterns of root and soil microbiome members paving the way to smart farming. Microbiome 2018, 6, 1–14. [Google Scholar] [CrossRef]
- Harman, G.E. Trichoderma—not just for biocontrol anymore. Phytoparasitica 2011, 39, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Köhl, J.; Postma, J.; Nicot, P.; Ruocco, M.; Blum, B. Stepwise screening of microorganisms for commercial use in biological control of plant-pathogenic fungi and bacteria. Biol. Control. 2011, 57, 1–12. [Google Scholar] [CrossRef]
- Bastakoti, S.; Belbase, S.; Manandhar, S.; Arjyal, C. Trichoderma species as biocontrol agent against soil borne fungal pathogens. Nepal J. Biotechnol. 2017, 5, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Daguerre, Y.; Edel-Hermann, V.; Steinberg, C. Fungal Genes and Metabolites Associated with the Biocontrol of Soil-Borne Plant Pathogenic Fungi; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.A.; Asolkar, R.N.; Inderbitzin, P.; Fenical, W. Secondary metabolite chemistry of the marine-derived fungus Massarina sp., strain CNT-016. Phytochemistry 2007, 68, 1212–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Molin, A.; Minio, A.; Griggio, F.; Delledonne, M.; Infantino, A.; Aragona, M. The genome assembly of the fungal pathogen Pyrenochaeta lycopersici from single-molecule real time sequencing sheds new light on its biological complexity. PLoS ONE 2018, 13, e0200217. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Pan, Z.; Yin, C.; Su, J.; Hu, X.; Zhu, X.; Shan, Y.; Fu, F. Antifungal activity and mechanism of ε-polylysine against Geotrichum citri-aurantii. Food Sci. 2020, 41, 221–229. [Google Scholar] [CrossRef]
- de León, Y.M.P.; Muñoz-Castellanos, L.N.; Ornelas-Paz, C.H.A.; Berlanga-Reyes, D.I.; Rios-Velasco, C. Morphological and molecular identification of Mortierella species associated to rhizosphere of apple trees with symptoms of root diseases. Rev. Mex. Fitopat 2018, 36, 184–195. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Reyes, M.F.; Carrasco, H.; Olea, A.F.; Silva-Moreno, E. Natural compounds: A sustainable alternative to the phytopathogens control. J. Chil. Chem. Soc. 2019, 64, 4459–4465. [Google Scholar] [CrossRef]
- Cerqueira, A.E.S.; Silva, T.H.; Nunes, A.C.S.; Nunes, D.D.; Lobato, L.C.; Veloso, T.G.R.; De Paula, S.O.; Kasuya, M.C.M.; Silva, C.C. Amazon basin pasture soils reveal susceptibility to phytopathogens and lower fungal community dissimilarity than forest. Appl. Soil. Ecol. 2018, 131, 1–11. [Google Scholar] [CrossRef]
- Pretscher, J.; Fischkal, T.; Branscheidt, S.; Jäger, L.; Kahl, S.; Schlander, M.; Thines, E.; Claus, H. Yeasts from different habitats and their potential as biocontrol agents. Fermentation 2018, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Oszust, K.; Pylak, M.; Frąc, M. Sposób otrzymywania biopreparatu do naturalizacji ryzosfery roślin malin o właściwościach antagonistycznych w stosunku do fitopatogenów grzybowych należących do rodzaju Botrytis, Verticillium, Colletotrichum i Phytophthora z zastosowaniem szczepów grzyba z rodzaju Trichoderma, biopreparat do naturalizacji ryzosfery roślin malin, sposób prowadzenia hodowli szczepów grzybów z rodzaju Trichoderma do zastosowania w biopreparacie oraz kompozycja podłoża namnażającego dla grzybów z rodzaju Trichoderma. Patent Application No. P.434148, 2020. [Google Scholar]
- Kumar, N.; Khurana, S.P. Trichoderma-plant-pathogen interactions for benefit of agriculture and environment. Biocontrol agents and secondary metabolites: Applications and immunization for plant growth and protection. Woodhead Publ. 2020, 41, 1–726. [Google Scholar]
- Keswani, C.; Bisen, K.; Singh, S.; Sarma, B.; Singh, H. A proteomic approach to understand the tripartite interactions between plant-Trichoderma-pathogen: Investigating the potential for efficient biological control. In Plant, Soil and Microbes; Springer: Cham, Switzerland, 2016; pp. 79–93. [Google Scholar] [CrossRef]
- Macías-Rodríguez, L.; Guzmán-Gómez, A.; García-Juárez, P.; Contreras-Cornejo, H.A. Trichoderma atroviride promotes tomato development and alters the root exudation of carbohydrates, which stimulates fungal growth and the biocontrol of the phytopathogen Phytophthora cinnamomi in a tripartite interaction system. FEMS Microb. Ecol. 2018, 94, fiy137. [Google Scholar] [CrossRef] [Green Version]
- Schuster, A.; Schmoll, M. Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechn. 2010, 87, 787–799. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Boer, W.; Folman, L.B.; Summerbell, R.C.; Boddy, L. Living in a fungal world: Impact of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 2005, 29, 795–811. [Google Scholar] [CrossRef] [Green Version]
- Macías-Rodríguez, L.; Contreras-Cornejo, H.A.; Adame-Garnica, S.G.; del-Val, E.; Larsen, J. The interactions of Trichoderma at multiple trophic levels: Inter-kingdom communication. Microbiol. Res. 2020, 240, 126552. [Google Scholar] [CrossRef]
- Lasa, A.V.; Mašínová, T.; Baldrian, P.; Fernández-López, M. Bacteria from the endosphere and rhizosphere of Quercus spp. use mainly cell wall-associated enzymes to decompose organic matter. PLoS ONE 2019, 14, e0214422. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Hu, Y.; Zhang, S.; Noll, L.; Böckle, T.; Dietrich, M.; Herbold, C.W.; Eichorst, S.A.; Woebken, D.; Richter, A.; et al. Soil multifunctionality is affected by the soil environment and by microbial community composition and diversity. Soil. Biol. Biochem. 2019, 136, 107521. [Google Scholar] [CrossRef]
- Comerford, N.B.; Franzluebbers, A.J.; Stromberger, M.E.; Morris, L.; Markewitz, D.; Moore, R. Assessment and evaluation of soil ecosystem services. Soil Horizons. 2013, 54, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Harman, G.E.; Petzoldt, R.; Comis, A.; Chen, J. Interactions between Trichoderma harzianum strain T22 and maize inbred line Mo17 and effects of these interactions on diseases caused by Pythium ultimum and Colletotrichum graminicola. Phytopathology 2004, 94, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Eslahi, N.; Kowsari, M.; Motallebi, M.; Zamani, M.R.; Moghadasi, Z. Influence of recombinant Trichoderma strains on growth of bean (Phaseolus vulgaris L.) by increased root colonization and induction of root growth related genes. Sci. Hort. 2020, 261, 108932. [Google Scholar] [CrossRef]
- Poveda, J.; Eugui, D.; Abril-Urias, P. Could Trichoderma be a plant pathogen? Successful root colonization. In Trichoderma: Host Pathogen Interactions and Applications; Sharma, A.K., Sharma, P., Eds.; Springer: Singapore, 2020; pp. 35–59. [Google Scholar] [CrossRef]
- Lombardi, N.; Caira, S.; Troise, A.D.; Scaloni, A.; Vitaglione, P.; Vinale, F.; Marra, R.; Salzano, A.M.; Lorito, M.; Woo, S.L. Trichoderma applications on strawberry plants modulate the physiological processes positively affecting fruit production and quality. Front. Microbiol. 2020, 11, 1364. [Google Scholar] [CrossRef] [PubMed]
- Jangir, M.; Sharma, S.; Sharma, S. Non-target Effects of Trichoderma on plants and soil microbial communities. In Plant Microbe Interface; Springer: Cham, Switzerland, 2019; pp. 239–251. [Google Scholar] [CrossRef]
- Akkaya, Ö.; Şeker, M.G.; Çiftçi, Y.Ö. Plant growth-promoting microbiome network. In Beneficial Microbes for Sustainable Agriculture and Environmental Management; Sangeetha, J., Thangadurai, D., Islam, S., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 27–80. [Google Scholar]
- Naliukhin, A.; Glinushkin, A.; Khamitova, S.; Avdeev, Y.M. The influence of biomodified fertilizers on the productivity of crops and biological properties of soddy-podzolic soils. Entomol. Appl. Sci. Lett. 2018, 5, 1–7. [Google Scholar] [CrossRef]
- Halob, A.A.; Gatea, I.H.; Khalaf, M.K.; Sabar, A.B. Biopreparation for antimicrobial material from mixture of nano silver and olive leaves extract. IOP Conf. Ser. Mater. Sci. Eng. 2020, 928, 062008. [Google Scholar] [CrossRef]
- Fidelis, O.I. Role of microorganisms in bioconversion of solid wastes and economic development. Int. J. Med. Sci. Clin. Res. Rev. 2018, 1, 1–6. [Google Scholar]
- Jain, M.; Pal, A. Organic waste management. SSRN 2020. [Google Scholar] [CrossRef]
- Mercy, S.; Mubsira, B.S.; Jenifer, I. Application of different fruit peels formulations as a natural fertilizer for plant growth. Int. J. Sci. Technol. Res. 2014, 3, 300–307. [Google Scholar]
- Smolinska, U.; Kowalska, B.; Kowalczyk, W.; Szczech, M. The use of agro-industrial wastes as carriers of Trichoderma fungi in the parsley cultivation. Sci. Hort. 2014, 179, 1–8. [Google Scholar] [CrossRef]
- Oleszek, M.; Pecio, Ł.; Kozachok, S.; Lachowska-Filipiuk, Ż.; Oszust, K.; Frąc, M. Phytochemicals of apple pomace as prospect bio-fungicide agents against mycotoxigenic fungal species—In vitro experiments. Toxins 2019, 11, 361. [Google Scholar] [CrossRef] [Green Version]
- Naeimi, S.; Khosravi, V.; Varga, A.; Vágvölgyi, C.; Kredics, L. Screening of organic substrates for solid-state fermentation, viability and bioefficacy of Trichoderma harzianum AS12-2, a biocontrol strain against rice sheath blight disease. Agronomy 2020, 10, 1258. [Google Scholar] [CrossRef]
- Malusá, E.; Sas-Paszt, L.; Ciesielska, J. Technologies for beneficial microorganisms inocula used as biofertilizers. Sci. World J. 2012, 491206. [Google Scholar] [CrossRef]
- Doni, F.; Zain, C.R.; Isahak, A.; Fathurrahman, F.; Anhar, A.; Mohamad, W.N.W.; Yusoff, W.M.W.; Uphoff, N. A simple, efficient, and farmer-friendly Trichoderma-based biofertilizer evaluated with the SRI Rice Management System. Org. Agric. 2018, 8, 207–223. [Google Scholar] [CrossRef]
- van Diepen, L.T.; Frey, S.D.; Landis, E.A.; Morrison, E.W.; Pringle, A. Fungi exposed to chronic nitrogen enrichment are less able to decay leaf litter. Ecology 2017, 98, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Vassilev, N.; Vassileva, M.; Martos, V.; Del Moral, L.F.G.; Kowalska, J.; Tylkowski, B.; Malusa, E. Formulation of microbial inoculants by encapsulation in natural polysaccharides: Focus on beneficial properties of carrier additives and derivatives. Front. Plant Sci. 2020, 1, 270. [Google Scholar] [CrossRef]
- Akter, N. Rheological Behaviour of Probiotic Bacteria Dispersed in Maltodextrin and Sucrose Solutions; Malmö Universitet: Malmo, Sweden, 2020. [Google Scholar]
- Samedi, L.; Charles, A.L. Viability of 4 probiotic bacteria microencapsulated with arrowroot starch in the simulated gastrointestinal tract (GIT) and yoghurt. Foods 2019, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Hofman, D.L.; Van Buul, V.J.; Brouns, F.J. Nutrition, health, and regulatory aspects of digestible maltodextrins. Crit. Rev. Food Sci. Nutr. 2016, 56, 2091–2100. [Google Scholar] [CrossRef] [PubMed]
- Lobo, C.B.; Tomás, M.S.J.; Viruel, E.; Ferrero, M.A.; Lucca, M.E. Development of low-cost formulations of plant growth-promoting bacteria to be used as inoculants in beneficial agricultural technologies. Microbiol. Res. 2019, 219, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Baena-Aristizábal, C.M.; Foxwell, M.; Wright, D.; Villamizar-Rivero, L. Microencapsulation of Rhizobium leguminosarum bv. trifolii with guar gum: Preliminary approach using spray drying. J. Biotechnol. 2019, 302, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Pertile, G.; Lamorski, K.; Bieganowski, A.; Boguta, P.; Brzezińska, M.; Polakowski, C.; Skic, K.; Sokołowska, Z.; Baranowski, P.; Gackiewicz, B.; et al. Immediate effects of the application of various fungal strains with urea fertiliser on microbiome structure and functions and their relationships with the physicochemical parameters of two different soil types. Appl. Soil Ecol. 2021, 103972. [Google Scholar] [CrossRef]
- Gryta, A.; Frąc, M.; Oszust, K. Genetic and metabolic diversity of soil microbiome in response to exogenous organic matter amendments. Agronomy 2020, 10, 546. [Google Scholar] [CrossRef] [Green Version]
- Raut, I.; Constantin, M.; Vasilescu, G.; Arsene, M.; Jecu, L.; Sesan, T. Optimization of Trichoderma strain cultivation for biocontrol activity. Sci. Bull. Seria F. Biotechnol. 2013, 154–159. [Google Scholar]
- Oszust, K.; Pawlik, A.; Siczek, A.; Janusz, G.; Gryta, A.; Bilińska-Wielgus, N.; Frąc, M. Efficient cellulases production by Trichoderma atroviride G79/11 in submerged culture based on soy flour-cellulose-lactose. BioResources 2017, 12, 8468–8489. [Google Scholar] [CrossRef]
- Palomar, L.T.; Alpirez, G.M.; Díaz, L.C.; Navarro, R.F.; Moreno, M.A.; Méndez, J.V. Partial characterization of the cellulolytic enzyme produced by filamentous fungi. In Microbes in the Spotlight: Recent Progress in the Understanding of Beneficial and Harmful Microorganisms; BrownWalker Press: Boca Raton, FL, USA, 2016; p. 411. [Google Scholar]
- Liu, H.Q.; Feng, Y.; Zhao, D.Q.; Jiang, J.X. Evaluation of cellulases produced from four fungi cultured on furfural residues and microcrystalline cellulose. Biodegradation 2012, 23, 465–472. [Google Scholar] [CrossRef]
- Chinedu, S.N.; Okochi, V.; Omidiji, O. Cellulase production by wild strains of Aspergillus niger, Penicillium chrysogenum and Trichoderma harzianum grown on waste cellulosic materials. Life J. Sci. 2011, 13, 57–62. [Google Scholar]
- Motta, F.L.; Santana, M.H.A. Biomass production from Trichoderma viride in nonconventional oat medium. Biotechnol. Prog. 2012, 28, 1245–1250. [Google Scholar] [CrossRef]
- Atlas, R.M. Handbook of Microbiological Media, 4th ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar] [CrossRef]
- Malarczyk, D.G.; Panek, J.; Frąc, M. Triplex real-time PCR approach for the detection of crucial fungal berry pathogens—Botrytis spp., Colletotrichum spp. and Verticillium spp. Int. J. Mol. Sci. 2020, 21, 8469. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- King, B.C.; Donnelly, M.K.; Bergstrom, G.C.; Walker, L.P.; Gibson, D.M. An optimized microplate assay system for quantitative evaluation of plant cell wall–degrading enzyme activity of fungal culture extracts. Biotechnol. Bioeng. 2009, 102, 1033–1044. [Google Scholar] [CrossRef]
- Janas, P.; Targoński, Z. Effect of temperature on the production of cellulases, xylanases and lytic enzymes by selected Trichoderma reesei mutants. Acta Mycol. 1995, 30, 255–264. [Google Scholar] [CrossRef]
- Hankin, L.; Anagnostakis, S.L. Solid media containing carboxymethylcellulose to detect Cx cellulase activity of micro-organisms. Microbiology 1977, 98, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deschamps, F.; Huet, M. β-Glucosidase production in agitated solid fermentation, study of its properties. Biotechnol. Lett. 1984, 6, 451–456. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylase α and β. Methods Enzymol. 1955, 1, 149–151. [Google Scholar] [CrossRef]
- Anson, M.L. The estimation of pepsin, trypsin, papain, and cathepsin with hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Mandels, M.; Andreotti, R. Problems and challenges in the cellulose to cellulase fermentation. Process. Biochem. 1978, 13, 6–13. [Google Scholar]
- PN-ISO-10390. Oznaczanie pH Gleby; Polish Committee of Standardization: Warszawa, Poland, 1997. [Google Scholar]
- PN-R-04022:1996+Az1:2002. Zawartość Potasu Przyswajalnego; Polish Committee of Standardization: Warszawa, Poland, 2002. [Google Scholar]
- PN-R-04020:1994+Az1:2004. Zawartość Magnezu Przyswajalnego; Polish Committee of Standardization: Warszawa, Poland, 2004. [Google Scholar]
- Waszkiewicz-Robak, B.; Swiderski, F. Hydrokoloidy pochodzenia roślinnego jako zamienniki żelatyny. Bezpieczna Żywność 2001, 1, 31–37. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Code | GenBank Sequences Accession Number | Identification | Isolation Compartment and Institution | GPS Coordinates | Forest District | Agar Medium |
|---|---|---|---|---|---|---|
| G25/18 | MW151041 | Apiotrichum porosum | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G26/18 | MW151042 | Apiotrichum porosum | Wild raspberry rhizosphere, IA PAS, Poland | - | Łuków | Sabouraud |
| G27/18 | MW151043 | Cryptococcus sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°01’08.3” N 22°01’57.8” E | Kraśnik | Rose Bengal |
| G28/18 | MW151044 | Aureobasidium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.7” N 21°95’99.3” E | Kraśnik | Rose Bengal |
| G29/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°01’08.3” N 22°01’57.8” E | Kraśnik | Sabouraud |
| G30/18 | MW150994.1 | Trichoderma sp. | Wild raspberry roots, IA PAS, Poland | 51°00’90.3” N 22°01’13.9” E | Kraśnik | Rose Bengal |
| G31/18 | MW150994 | Trichoderma sp. | Wild raspberry roots, IA PAS, Poland | 51°41’39.5” N 22°24’40.2” E | Łuków | Rose Bengal |
| G32/18 | MW150995 | Apiotrichum porosum | Wild raspberry roots, IA PAS, Poland | 51°00’93.6” N 22°01’22.0” E | Kraśnik | Rose Bengal |
| G33/18 | MW151045 | Solicoccozyma sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Łuków | Sabouraud |
| G34/18 | MW151046 | Apiotrichum porosum | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G35/18 | MW151047 | Cryptococcus sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Sabouraud |
| G36/18 | MW151048 | Solicoccozyma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Sabouraud |
| G37/18 | MW151049 | Solicoccozyma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.7” N 21°95’99.3” E | Kraśnik | Sabouraud |
| G38/18 | MW151050 | Solicoccozyma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.7” N 21°95’99.3” E | Kraśnik | Sabouraud |
| G39/18 | MW151051 | Umbelopsis vinacea | Wild raspberry rhizosphere, IA PAS, Poland | 51°01’08.3” N 22°01’77.8” E | Kraśnik | Sabouraud |
| G40/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G41/18 | MW150996 | Umbelopsis sp. | Wild raspberry roots, IA PAS, Poland | 51°79’04.0” N 22°20’68.1” E | Łuków | Sabouraud |
| G42/18 | MW151052 | Umbelopsis isabelina | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Sabouraud |
| G43/18 | MW151053 | Apiotrichum porosum | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Rose Bengal |
| G44/18 | MW151054 | Apiotrichum porosum | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Rose Bengal |
| G45/18 | MW151055 | Mortierella sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°00’90.3” N 22°01’13.9” E | Kraśnik | Sabouraud |
| G46/18 | MW151056 | Umbelopsis sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Sabouraud |
| G47/18 | MW151057 | Umbelopsis sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Łuków | Sabouraud |
| G48/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°01’08.3” N 22°01’77.8” E | Kraśnik | Sabouraud |
| G49/18 | MW151058 | Apiotrichum porosum | Wild raspberry rhizosphere, IA PAS, Poland | - | Łuków | Sabouraud |
| G50/18 | MW151059 | Apiotrichum porosum | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Sabouraud |
| G51/18 | MW151060 | Geotrichum europaeum | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’27.5” N 22°24’36.6” E | Janów Lubelski | Sabouraud |
| G52/18 | MW151061 | Geotrichum europaeum | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Rose Bengal |
| G53/18 | MW151062 | Apiotrichum porosum | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Rose Bengal |
| G54/18 | MW151063 | Plectospahrella cucumerina | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G55/18 | MW257185 | Plectospahrella sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G56/18 | MW257186 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Sabouraud |
| G57/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Sabouraud |
| G58/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’10.0” N 21°95’99.8” E | Kraśnik | Sabouraud |
| G59/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°00’90.3” N 22°01’13.9” E | Kraśnik | Rose Bengal |
| G60/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’27.5” N 22°24’36.6” E | Janów Lubelski | Rose Bengal |
| G61/18 | MW233576 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | Siedlce | Rose Bengal | |
| G62/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°01’05.1” N 22°01’67.2” E | Kraśnik | Rose Bengal |
| G63/18 | MT558561 | Trichoderma sp. | Wild raspberry roots, IA PAS, Poland | 51°54’46.2” N 22°28’30.6” E | Świdnik | Rose Bengal |
| G64/18 | MT558562 | Trichoderma sp. | Wild raspberry roots, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G65/18 | MW233577 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.2” N 22°28’30.6” E | Świdnik | Rose Bengal |
| G66/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G67/18 | MW205828 | Trichoderma sp. | Wild raspberry roots, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Rose Bengal |
| G68/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Sabouraud |
| G69/18 | MT559294 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Rose Bengal |
| G70/18 | MW233578 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Rose Bengal |
| G71/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | Janów Lubelski | Rose Bengal | |
| G72/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Rose Bengal |
| G73/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Sabouraud |
| G74/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Rose Bengal |
| G75/18 | MT558563.1 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Rose Bengal |
| G76/18 | MW257187 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.7” N 21°95’99.3” E | Kraśnik | Rose Bengal |
| G77/18 | MW257194 | Trichoderma sp. | Wild raspberry roots, IA PAS, Poland | 50°39’27.3” N 22°24’10.7” E | Janów Lubelski | Sabouraud |
| G78/18 | MW205829 | Trichoderma sp. | Wild raspberry roots, IA PAS, Poland | 50°39’27.3” N 22°24’10.7” E | Janów Lubelski | Rose Bengal |
| G79/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Rose Bengal |
| G80/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Rose Bengal |
| G81/18 | MW257188 | Mortierella sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Rose Bengal |
| G83/18 | MW257189 | Mortierella sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G84/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°17’10.0” N 21°95’99.8” E | Kraśnik | Sabouraud |
| G85/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G86/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G87/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°00’93.6” N 22°01’22.0” E | Kraśnik | Rose Bengal |
| G88/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°00’93.6” N 22°01’22.0” E | Kraśnik | Rose Bengal |
| G89/18 | MW257190 | Mucor sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G90/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’10.0” N 21°95’99.8” E | Kraśnik | Sabouraud |
| G91/18 | MW257191 | Mucor sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°00’92.6” N 22°01’12.0” E | Kraśnik | Sabouraud |
| G92/18 | MW257192 | Absidia sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°00’90.3” N 22°01’13.9” E | Kraśnik | Sabouraud |
| G93/18 | MW257193 | Mucor sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°00’90.3” N 22°01’13.9” E | Kraśnik | Rose Bengal |
| G94/18 | MW257196 | Fusarium sp. | Wild raspberry roots, IA PAS, Poland | 50°39’27.5” N 22°24’36.6” E | Janów Lubelski | Rose Bengal |
| G95/18 | MW257197 | Fusarium sp. | Wild raspberry roots, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G96/18 | MW257195 | Umbelopsis sp. | Wild raspberry roots, IA PAS, Poland | 51°41’39.5” N 22°24’40.2” E | Łuków | Sabouraud |
| G97/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’27.3” N 22°24’10.7” E | Janów Lubelski | Rose Bengal |
| G98/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G99/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G100/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G101/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°00’90.3” N 22°01’13.9” E | Kraśnik | Sabouraud |
| G102/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Sabouraud |
| G103/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Sabouraud |
| G104/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°00’90.3” N 22°01’13.9” E | Kraśnik | Sabouraud |
| G105/18 | - | - | Wild raspberry roots, IA PAS, Poland | Siedlce | Sabouraud | |
| G106/18 | MW150997 | Fusarium venenatum | Wild raspberry roots, IA PAS, Poland | 51°17’13.7” N 21°95’99.3” E | Kraśnik | Sabouraud |
| G107/18 | MW150998 | Fusarium sp. | Wild raspberry roots, IA PAS, Poland | 51°54’46.2” N 22°28’30.6” E | Świdnik | Sabouraud |
| G108/18 | MW150999 | Fusarium roseum | Wild raspberry roots, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Sabouraud |
| G109/18 | MW233579 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.7” N 21°95’99.3” E | Kraśnik | Sabouraud |
| G110/18 | MW151000 | Fusarium sp. | Wild raspberry roots, IA PAS, Poland | 50°39’27.3” N 22°24’10.7” E | Janów Lubelski | Sabouraud |
| G111/18 | - | - | Wild raspberry roots, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Sabouraud |
| G112/18 | - | - | Wild raspberry roots, IA PAS, Poland | - | Siedlce | Rose Bengal |
| G113/18 | MW151064 | Talaromyces sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Sabouraud |
| G114/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G115/18 | MW151001 | Penicillium coprobium | Wild raspberry roots, IA PAS, Poland | 51°17’10.0” N 21°95’99.8” E | Kraśnik | Rose Bengal |
| G116/18 | MW151065 | Penicillium coprobium | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’27.3” N 22°24’10.7” E | Janów Lubelski | Rose Bengal |
| G117/18 | MW151066 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°41’39.5” N 22°24’40.2” E | Łuków | Rose Bengal |
| G118/18 | MW151067 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’27.5” N 22°24’36.6” E | Janów Lubelski | Rose Bengal |
| G119/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°01’05.1” N 22°01’67.2” E | Kraśnik | Rose Bengal |
| G120/18 | MW151068 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G121/18 | MW151069 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.2” N 22°28’30.6” E | Świdnik | Rose Bengal |
| G122/18 | MW151070 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°01’05.1” N 22°01’67.2” E | Kraśnik | Rose Bengal |
| G123/18 | MW151071 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Rose Bengal |
| G124/18 | MW151072 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Rose Bengal |
| G125/18 | MW151002 | Penicillium sp. | Wild raspberry roots, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G126/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G127/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Rose Bengal |
| G128/18 | MW151073 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’47.9” N 22°28’44.8” E | Świdnik | Sabouraud |
| G129/18 | MW151074 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | Łuków | Sabouraud | |
| G130/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G131/18 | MW151075 | Cladosporium sp. | Wild raspberry rhizosphere, IA PAS, Poland | Łuków | Sabouraud | |
| G132/18 | MW151076 | Cladosporium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’27.5” N 22°24’36.6” E | Janów Lubelski | Rose Bengal |
| G133/18 | MW151077 | Cladosporium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’10.0” N 21°95’99.8” E | Kraśnik | Sabouraud |
| G134/18 | MW151078 | Humicola sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Rose Bengal |
| G135/18 | MW151079 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G136/18 | MW151080 | Massarina sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G137/18 | MW151081 | Cladosporium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G138/18 | MW151082 | Cladosporium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Sabouraud |
| G139/18 | MW151083 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Sabouraud |
| G140/18 | MW250233 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’27.3” N 22°24’10.7” E | Janów Lubelski | Rose Bengal |
| G141/18 | - | - | Wild raspberry roots, IA PAS, Poland | 50°39’27.3” N 22°24’10.7” E | Janów Lubelski | Rose Bengal |
| G142/18 | MW151003 | Alternaria sp. | Wild raspberry roots, IA PAS, Poland | 51°17’10.0” N 21°95’99.8” E | Kraśnik | Sabouraud |
| G143/18 | - | - | Wild raspberry roots, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Sabouraud |
| G144/18 | MW151004 | Mucor moelleri | Wild raspberry roots, IA PAS, Poland | 51°01’05.1” N 22°01’67.2” E | Kraśnik | Sabouraud |
| G145/18 | MW151005 | Podospora sp. | Wild raspberry roots, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Sabouraud |
| G146/18 | MW151006 | Apiosporaceae | Wild raspberry roots, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Sabouraud |
| G147/18 | MW151084 | Coniothyrium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Rose Bengal |
| G148/18 | MW151085 | Coniothyrium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G149/18 | MW151086 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | 51°17’13.0” N 21°95’99.7” E | Kraśnik | Sabouraud |
| G150/18 | MW151087 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G151/18 | MW151088 | Pyrenochaeta sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Rose Bengal |
| G152/18 | MW151089 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Rose Bengal |
| G153/18 | MW151090 | Penicillium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Sabouraud |
| G154/18 | MW151091 | Fusarium sp. | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Sabouraud |
| G155/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Rose Bengal |
| G157/18 | MW151007 | Mucor sp. | Wild raspberry roots, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Sabouraud |
| G158/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Siedlce | Rose Bengal |
| G159/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 51°54’46.6” N 22°28’29.9” E | Świdnik | Sabouraud |
| G375/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G376/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Pikovskaya |
| G377/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G378/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G379/18 | MT559285 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Pikovskaya |
| G380/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Pikovskaya |
| G381/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Pikovskaya |
| G382/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G383/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G384/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G385/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G386/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G387/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G388/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G389/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G390/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G391/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G392/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G393/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G394/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Janów Lubelski | Pikovskaya |
| G395/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G396/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G397/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G398/18 | MT559286 | Trichoderma sp. | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Pikovskaya |
| G399/18 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | - | Kraśnik | Pikovskaya |
| G6/19 | - | - | Wild raspberry rhizosphere, IA PAS, Poland | 50°39’42.1” N 22°24’21.0” E | Janów Lubelski | Potato Dextrose |
| G172/18 | MT126803 | Colletotrichum sp. | Strawberry fruit, IA PAS, Poland | - | - | Potato Dextrose |
| G371/18 | MT558572.1 | Colletotrichum sp. | Research Institute of Horticulture | - | - | Potato Dextrose |
| G166/18 | MT126798 | Colletotrichum sp. | Strawberry roots, IA PAS, Poland | - | - | Potato Dextrose |
| G293/18 | MT133324 | Verticillium sp. | Strawberry roots, IA PAS, Poland | - | - | Potato Dextrose |
| G296/18 | MT133320 | Verticillium sp. | Strawberry roots, IA PAS, Poland | - | - | Potato Dextrose |
| G297/18 | MT133316 | Verticillium sp. | Strawberry roots, IA PAS, Poland | - | - | Potato Dextrose |
| G368/18 | MT558571 | Phytophthora sp. | Research Institute of Horticulture | - | - | - |
| G408/18 | MT126670 | Phytophthora sp. | Strawberry roots, IA PAS, Poland | - | - | Potato Dextrose |
| G373/18 | - | Phytophthora sp. | Research Institute of Horticulture | - | - | - |
| G369/18 | MT558729 | Phytophthora cactorum | Research Institute of Horticulture | - | - | - |
| G275/16 | MT154302 | Botrytis sp. | Strawberry roots, IA PAS, Poland | - | - | Potato Dextrose |
| G277/18 | MT154304 | Botrytis sp. | Strawberry roots, IA PAS, Poland | - | - | Potato Dextrose |
| G276/18 | MT154303 | Botrytis sp. | Strawberry roots, IA PAS, Poland | - | - | Potato Dextrose |
| Trichoderma spp. | Botrytis spp. | Colletotrichum spp. | Verticillium spp. | Phytophthora spp. | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Growth Inhibition (mm) | Sporulation Inhibition (mm) | Growth Inhibition (mm) | Sporulation Inhibition (mm) | Growth Inhibition (mm) | Sporulation Inhibition (mm) | Growth Inhibition (mm) | Sporulation Inhibition (mm) | |||||
| G61/18 | G275/18 | 46.5 ± 3.5 | - | G172/18 | 46.7 ± 18.1 | - | G293/18 | 22.5 ± 1.6 | 70.6 ± 3.8 | G368/18 | - | 49.3 ± 9.4 |
| G63/18 | 63.9 ± 5.5 | - | 45.5 ± 8.0 | - | 44.2 ± 13.0 | 67.5 ± 1.2 | 33.5 ± 0.0 | - | ||||
| G64/18 | 69.9 ± 4.9 | - | 45.9 ± 2.6 | - | 42.3 ± 5.1 | 72.9 ± 3.8 | 21.9 ± 1.8 | 55.7 ± 2.4 | ||||
| G65/18 | 39.0 ± 8.5 | - | 32.1 ± 1.8 | - | 24.2 ± 1.2 | 55.8 ± 1.8 | 12.7 ± 0.6 | - | ||||
| G67/18 | 60.4 ± 13.5 | - | 22.3 ± 2.0 | - | 47.6 ± 10.3 | - | 24.7 ± 3.1 | - | ||||
| G69/18 | 51.0 ± 15.6 | - | 38.3 ± 7.0 | - | 34.7 ± 1.8 | 63.4 ± 7.3 | 29.7 ± 1.4 | 58.8 ± 10.8 | ||||
| G70/18 | 32.5 ± 7.8 | - | 23.9 ± 4.7 | - | 40.0 ± 0.0 | - | 90.0 ± 0.0 | - | ||||
| G78/18 | 45.6 ± 15.1 | - | 31.3 ± 1.8 | - | 20.7 ± 0.6 | 62.4 ± 1.1 | 37.6 ± 9.3 | - | ||||
| G109/18 | 38.1 ± 4.5 | - | 17.1 ± 1.2 | - | 53.0 ± 13.2 | - | 67.9 ± 14.7 | - | ||||
| G379/18 | 40.1 ± 1.4 | - | - | 18.0 ± 0.0 | 40.5 ± 3.5 | - | - | 34.4 ± 2.3 | ||||
| G398/18 | 38.0 ± 1.9 | - | - | 24.0 ± 2.8 | 30.0 ± 1.4 | 55.6 ± 3.4 | - | - | ||||
| G61/18 | G277/18 | 66.8 ± 16.4 | - | G371/18 | 58.9 ± 6.7 | - | G296/18 | 23.4 ± 4.4 | 66.6 ± 8.1 | G373/18 | 48.3 ± 1.3 | - |
| G63/18 | 52.8 ± 15.6 | - | 38.7 ± 5.9 | - | 33.8 ± 8.6 | 72.3 ± 11.6 | 48.3 ± 9.4 | - | ||||
| G64/18 | 60.4 ± 15.9 | - | 50.4 ± 11.5 | - | 36.9 ± 9.7 | 63.4 ± 6.1 | 54.2 ± 16.4 | - | ||||
| G65/18 | 55.2 ± 11.2 | - | 36.8 ± 12.7 | - | 25.3 ± 0.6 | 59.7 ± 5.2 | 39.4 ± 8.0 | - | ||||
| G67/18 | 22.5 ± 0.7 | 54.0 ± 9.9 | 29.2 ± 2.2 | - | 55.1 ± 8.9 | - | 25.7 ± 3.9 | 66.6 ± 14.9 | ||||
| G69/18 | 39.6 ± 5.5 | - | 27.8 ± 6.8 | - | 37.4 ± 12.1 | 68.0 ± 10.8 | 49.0 ± 21.2 | - | ||||
| G70/18 | 38.7 ± 5.5 | - | 20.1 ± 3.0 | - | 37.6 ± 0.6 | - | 37.7 ± 10.9 | - | ||||
| G78/18 | 37.0 ± 1.4 | - | 36.2 ± 5.9 | - | 21.4 ± 0.6 | 56.8 ± 10.1 | 39.7 ± 1.4 | - | ||||
| G109/18 | 29.1 ± 8.0 | - | 15.3 ± 7.2 | - | 27.8 ± 9.3 | - | 50.2 ± 4.0 | - | ||||
| G379/18 | 36.1 ± 2.2 | - | - | 23.0 ± 1.4 | - | 55.0 ± 15.6 | - | 49.6 ± 1.3 | ||||
| G398/18 | 30.0 ± 2.8 | - | - | 16.0 ± 4.8 | 31.2 ± 0.7 | 49.6 ± 3.6 | - | 56.7 ± 12.7 | ||||
| G61/18 | G276/18 | 40.2 ± 9.0 | - | G166/18 | 90.0 ±0.0 | G297/18 | 29.0 ± 2.2 | 73.2 ± 12.7 | G369/18 | 54.9 ± 12.1 | - | |
| G63/18 | 31.2 ± 2.5 | 70.8 ± 6.9 | 45.8 ± 20.8 | - | 47.1 ± 6.1 | 79.3 ± 15.2 | 35.2 ± 0.0 | - | ||||
| G64/18 | 71.5 ± 17.0 | - | 64.0 ± 13.1 | - | 55.6 ± 12.3 | 83.6 ± 9.1 | 20.5 ± 2.9 | 62.4 ± 0.0 | ||||
| G65/18 | 35.5 ± 2.0 | 55.9 ± 4.0 | 30.1 ± 1.8 | - | - | 65.8 ± 14.2 | 14.2 ± 0.5 | - | ||||
| G67/18 | 14.5 ± 3.3 | 38.4 ± 27.8 | 33.0 ± 5.5 | - | 61.8 ± 4.4 | - | 25.9 ± 3.1 | - | ||||
| G69/18 | 33.5 ± 3.4 | 61.3 ± 4.5 | 27.0 ±0.0 | - | 37.1 ± 2.7 | 60.0 ± 8.5 | 30.3 ± 1.5 | 76.4 ± 1.5 | ||||
| G70/18 | 31.3 ± 4.0 | - | 46.7 ± 30.6 | - | 40.7 ± 0.6 | - | 90.0 ± 0.0 | |||||
| G78/18 | 34.4 ± 14.9 | - | 25.8 ± 3.5 | - | 24.7 ± 0.0 | 56.2 ± 1.7 | 61.0 ± 13.3 | - | ||||
| G109/18 | 34.7 ± 13.1 | - | 19.4 ± 3.3 | - | 25.8 ± 5.9 | - | 83.5 ± 9.2 | - | ||||
| G379/18 | - | 44.5 ± 0.0 | - | 23.04 ± 5.3 | 63.1 ± 11.7 | - | - | 48.2 ± 0.0 | ||||
| G398/18 | - | 47.4 ± 6.7 | - | 28.08 ± 0.0 | - | 69.3 ± 8.9 | - | 14.02 ± 1.3 | ||||
| Isolate | FPU | CMC | BGL | XYL | PRO | AMY | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (μmol min−1 mL−1) | ||||||||||||
| 37 °C, pH 7 | 37 °C, pH 4.5 | 50 °C, pH 7 | 50 °C, pH 4.5 | 37°C, pH 7 | 37°C, pH 4.5 | 50°C, pH 7 | 50°C, pH 4.5 | 37°C, pH 5.0 | 30°C, pH 4.8 | 37°C, pH 7.5 | 30°C, pH 5.0 | |
| G61/18 | 0.75 ± 0.02 | 0.16 ± 0.01 | 0.23 ± 0.02 | 0.29 ± 0.04 | 0.32 ± 0.01 | 0.15 ± 0.01 | - | - | 48.17 ± 0.03 | 139.56 ± 0.10 | - | 915.58 ± 0.17 |
| G63/18 | 0.32 ± 0.03 | - | - | - | 0.56 ± 0.02 | 0.08 ± 0.00 | 0.77 ± 0.08 | 0.09 ± 0.00 | 15.85 ± 0.02 | 616.35 ± 0.06 | 0.02 ± 0.02 | - |
| G64/18 | 0.36 ± 0.01 | 0.10 ± 0.00 | 1.23 ± 0.16 | - | 0.40 ± 0.03 | 0.09 ± 0.00 | 0.38 ± 0.00 | 0.53 ± 0.04 | 63.50 ± 0.16 | 214.23 ± 0.16 | 0.69 ± 0.04 | 585.11 ± 0.30 |
| G65/18 | 0.15 ± 0.10 | - | 0.80 ± 0.11 | 0.29 ± 0.00 | 0.52 ± 0.02 | - | - | 85.28 ± 0.06 | 2750.12 ± 0.08 | - | 441.53 ± 0.34 | |
| G67/18 | 0.33 ± 0.01 | - | 0.65 ± 0.02 | 0.39 ± 0.02 | - | - | 0.20 ± 0.02 | 0.48 ± 0.01 | 38.16 ± 0.13 | 1294.56 ± 0.16 | 0.37 ± 0.08 | 959 ± 0.35 |
| G69/18 | - | - | 0.03 ± 0.03 | 0.68 ± 0.02 | 1.15 ± 0.04 | - | 0.26 ± 0.06 | - | 87.05 ± 0.05 | - | 0.36 ± 0.02 | 541.73 ± 0.22 |
| G70/18 | 0.75 ± 0.02 | - | - | 0.74 ± 0.02 | - | - | - | - | 254.00 ± 0.07 | 1708.88 ± 0.41 | 0.77 ± 0.02 | 1130.11 ± 0.11 |
| G78/18 | - | 0.19 ± 0.02 | 0.87 ± 0.10 | 0.26 ± 0.01 | 0.23 ± 0.00 | 0.13 ± 0.00 | - | 0.17 ± 0.04 | 82.98 ± 0.02 | 141.38 ± 0.08 | 0.17 ± 0.05 | 443.40 ± 0.25 |
| G109/18 | 1.00 ± 0.02 | - | 0.94 ± 0.02 | 0.20 ± 0.02 | 0.02 ± 0.00 | - | - | - | 23.30 ± 0.01 | 1367.21 ± 0.10 | 0.23 ± 0.03 | 732.40 ± 0.10 |
| G379/18 | 0.26 ± 0.07 | 0.39 ± 0.01 | 0.30 ± 0.05 | - | 0.42 ± 0.01 | - | - | 0.40 ± 0.01 | 26.20 ± 0.02 | 135.32 ± 0.17 | - | - |
| G398/18 | 0.13 ± 0.03 | 0.99 ± 0.04 | - | 2.02 ± 0.03 | 0.58 ± 0.02 | - | - | - | 24.15 ± 0.05 | 288.83 ± 0.14 | 0.08 ± 0.00 | 508.72 ± 0.29 |
| Pathosystem | Naturalization | Macronutrients in Stems and Leaves | Absorbable Forms of Minerals in Soil | Nitrogen Forms in Soil | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | P2O5 | K2O | Mg | N-NO3 | N-NH4 | Nmin | ||
| % DM | mg/100 g soil | mg/kg DM | kg/ha | |||||||||
| Without pathogen | NN | 1.44 | 0.29 | 2.47 | 1.22 | 0.44 | 18.3 | 32.5 | 14.3 | 4.41 | 6.26 | 45.9 |
| R | 1.58 | 0.30 | 2.63 | 1.27 | 0.46 | 18.3 | 32.2 | 12.8 | 5.53 | 4.76 | 44.2 | |
| RW | 1.43 | 0.26 | 1.96 | 1.17 | 0.44 | 19.4 | 31.6 | 13.7 | 3.14 | 4.25 | 31.8 | |
| W | 1.36 | 0.26 | 2.33 | 1.17 | 0.46 | 19.5 | 34.8 | 12.3 | 8.91 | 4.17 | 56.2 | |
| Botrytis sp. | NN | 1.41 | 0.28 | 2.12 | 1.13 | 0.46 | 19.6 | 35.6 | 12.6 | 3.65 | 7.71 | 48.8 |
| R | 1.25 | 0.28 | 1.96 | 1.19 | 0.44 | 19.7 | 35.2 | 11.7 | 4.59 | 7.84 | 53.4 | |
| RW | 1.41 | 0.31 | 2.47 | 1.21 | 0.42 | 20.2 | 34.8 | 13.3 | 3.10 | 7.19 | 44.2 | |
| W | 1.21 | 0.28 | 1.94 | 1.12 | 0.36 | 16.4 | 34.8 | 14.9 | 3.59 | 6.32 | 42.6 | |
| Verticillium sp. | NN | 1.51 | 0.29 | 2.04 | 1.20 | 0.42 | 16.1 | 31.8 | 14.0 | 2.23 | 9.37 | 49.9 |
| R | 1.28 | 0.28 | 1.75 | 1.10 | 0.38 | 15.9 | 32.5 | 13.7 | 1.66 | 7.46 | 39.2 | |
| RW | 1.53 | 0.27 | 1.91 | 1.20 | 0.36 | 16.2 | 32.9 | 13.9 | 1.81 | 7.33 | 39.3 | |
| W | 1.31 | 0.27 | 2.24 | 1.23 | 0.36 | 16.8 | 31.7 | 12.3 | 1.32 | 9.71 | 41.8 | |
| Colletotrichum sp. | NN | 1.18 | 0.26 | 2.01 | 1.33 | 0.38 | 17.5 | 31.8 | 12.8 | 2.78 | 7.45 | 44.0 |
| R | 1.57 | 0.29 | 2.56 | 1.34 | 0.44 | 17.1 | 32.1 | 12.9 | 6.68 | 4.20 | 46.8 | |
| RW | 1.60 | 0.27 | 2.42 | 1.41 | 0.44 | 17.8 | 31.2 | 12.9 | 13.37 | 2.01 | 66.1 | |
| W | 1.08 | 0.29 | 2.03 | 1.26 | 0.40 | 20.4 | 34.5 | 13.7 | 3.53 | 6.96 | 45.1 | |
| Phytophthora sp. | NN | 1.60 | 0.28 | 2.56 | 1.36 | 0.40 | 18.5 | 32.3 | 14.4 | 4.4 | 6.61 | 47.3 |
| R | 1.48 | 0.25 | 2.35 | 1.27 | 0.38 | 20.0 | 37.2 | 14.9 | 8.76 | 8.25 | 73.1 | |
| RW | 1.51 | 0.26 | 2.00 | 1.27 | 0.40 | 15.3 | 33.0 | 14.9 | 2.12 | 6.68 | 37.8 | |
| W | 1.51 | 0.28 | 2.58 | 1.34 | 0.40 | 17.7 | 31.3 | 12.4 | 7.20 | 4.81 | 51.6 | |
| All pathogens | NN | 1.54 | 0.27 | 2.23 | 1.24 | 0.39 | 16.7 | 30.4 | 12.9 | 4.32 | 9.15 | 57.9 |
| R | 1.44 | 0.30 | 2.22 | 1.15 | 0.38 | 18.7 | 36.2 | 14.5 | 2.30 | 10.14 | 53.5 | |
| RW | 1.56 | 0.29 | 2.12 | 1.26 | 0.36 | 18.9 | 38.7 | 13.7 | 1.84 | 8.89 | 46.1 | |
| W | 1.14 | 0.27 | 2.15 | 1.29 | 0.40 | 16.8 | 34.0 | 13.8 | 2.54 | 11.53 | 60.5 | |
| Medium | Trichoderma spp. | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G61/18 | G63/18 | G64/18 | G65/18 | G67/18 | G69/18 | G70/18 | G78/18 | G109/18 | G379/18 | G398/18 | ||
| Agar medium | MSCL-A | |||||||||||
| TR | ||||||||||||
| MN | ||||||||||||
| MA | ||||||||||||
| WBAP-A | ||||||||||||
| WBAP-B | ||||||||||||
| WBAP-C | ||||||||||||
| WBAP-D | ||||||||||||
| Solid-state medium | WBAP-E | |||||||||||
| WBAP-F | ||||||||||||
| WBAP-G | ||||||||||||
| WBAP-H | ||||||||||||
| Liquid and commercial agar medium | MSCL-L | |||||||||||
| PDA | ||||||||||||
| CM | ||||||||||||
| MEA | ||||||||||||
| Average spores’ number per plate * | 1.31 × 1010 | 1.66 × 1010 | 6.67 × 1010 | 1.14 × 1010 | 6.53 × 109 | 3.83 × 1010 | 6.61 × 1010 | 1 × 1010 | 3.614 × 109 | 0 | 0 | |
| Gelling Agent Concentration (%) | Water Solution (%) | Xanthan Gum | Citrus Pectin | CarboxymethylCellulose | Sodium Alginate | Guar Gum | Locust Bean Gum | Apple Pectin | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adhering Suspension (g) | Viscosity (mPa) | Adhering Suspension (g) | Viscosity (mPa) | Adhering Suspension (g) | Viscosity (mPa) | Adhering Suspension (g) | Viscosity (mPa) | Adhering Suspension (g) | Viscosity (mPa) | Adhering Suspension (g) | Viscosity (mPa) | Adhering Suspension (g) | Viscosity (mPa) | ||
| 0.5% | 1% | <1.00 | 3.62 | <1.00 | 4.54 | <1.00 | 4.28 | <1.00 | 4.10 | <1.00 | 6.41 | <1.00 | 3.54 | - | - |
| 5% | <1.00 | 8.10 | <1.00 | 4.98 | - | - | - | - | - | - | - | - | - | - | |
| 10% | 1.44 | 9.53 | <1.00 | 5.60 | 1.36 | 7.25 | <1.00 | 7.80 | <1.00 | 8.54 | <1.00 | 5.45 | - | - | |
| 15% | 1.51 | 21.17 | 1.14 | 7.36 | - | - | - | - | - | - | - | - | - | - | |
| 20% | 1.91 | 32.01 | 1.42 | 9.09 | 1.81 | 10.82 | 1.32 | 11.04 | 1.02 | 9.82 | <1.00 | 7.80 | - | - | |
| 1% | 1% | <1.00 | 4.54 | <1.00 | 4.32 | - | - | - | - | - | - | - | - | - | - |
| 5% | 1.22 | 8.76 | <1.00 | 5.16 | - | - | - | - | - | - | - | - | - | - | |
| 10% | 1.70 | 14.19 | <1.00 | 6.37 | - | - | - | - | - | - | - | - | - | - | |
| 15% | 2.03 | 32.12 | <1.00 | 8.36 | - | - | - | - | - | - | - | - | - | - | |
| 20% | 2.68 | 53.38 | 1.10 | 9.93 | - | - | - | - | - | - | - | - | - | - | |
| 2% | 1% | <1.00 | 6.04 | <1.00 | 5.34 | <1.00 | 5.71 | <1.00 | 6.78 | <1.00 | 4.94 | <1.00 | 4.06 | - | - |
| 5% | 1.88 | 16.36 | <1.00 | 6.11 | - | - | - | - | - | - | - | - | - | - | |
| 10% | 2.13 | 32.96 | <1.00 | 7.33 | <1.00 | 9.64 | 1.23 | 12.98 | <1.00 | 9.13 | <1.00 | 7.44 | - | - | |
| 15% | 3.21 | 65.61 | <1.00 | 9.64 | - | - | - | - | - | - | - | - | - | - | |
| 20% | 5.42 | 116.8 | 1.38 | 14.23 | 1.11 | 29.51 | 1.72 | 55.99 | 1.51 | 28.3 | 1.34 | 10.41 | - | - | |
| 4% | 1% | <1.00 | 8.43 | <1.00 | 4.72 | - | - | - | - | - | - | - | - | - | - |
| 5% | 1.66 | 28.37 | <1.00 | 6.34 | - | - | - | - | - | - | - | - | - | - | |
| 10% | 3.79 | 75.51 | <1.00 | 8.83 | - | - | - | - | - | - | - | - | - | - | |
| 15% | 6.71 | 144.50 | <1.00 | 15.11 | - | - | - | - | - | - | - | - | - | - | |
| 20% | 7.90 | 177.90 | 2.31 | 36.74 | - | - | - | - | - | - | - | - | - | - | |
| 6% | 1% | 1.07 | 9.13 | <1.00 | 5.67 | <1.00 | 7.25 | <1.00 | 9.02 | <1.00 | <1.00 | <1.00 | 3.76 | - | - |
| 5% | 2.40 | 45.56 | <1.00 | 7.73 | - | - | - | - | - | - | - | - | - | - | |
| 10% | 3.25 | 103.30 | <1.00 | 11.99 | 1.61 | 30.06 | 1.86 | 54.63 | 3.37 | 72.33 | 2.56 | 11.62 | - | - | |
| 15% | 7.60 | 195.30 | 2.89 | 30.13 | - | - | - | - | - | - | - | - | - | - | |
| 20% | 10.14 | 335.70 | 3.41 | 65.53 | 2.68 | 181.40 | 4.32 | >365.00 | 7.23 | 238.60 | 3.76 | 60.61 | - | - | |
| 15% | 20% | - | - | 5.24 | >365.00 | - | - | - | - | - | - | - | - | 3.90 | 172.50 |
| 20% | 15% | - | - | 6.21 | 353.80 | - | - | - | - | - | - | - | - | 2.98 | 168.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oszust, K.; Pylak, M.; Frąc, M. Trichoderma-Based Biopreparation with Prebiotics Supplementation for the Naturalization of Raspberry Plant Rhizosphere. Int. J. Mol. Sci. 2021, 22, 6356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126356
Oszust K, Pylak M, Frąc M. Trichoderma-Based Biopreparation with Prebiotics Supplementation for the Naturalization of Raspberry Plant Rhizosphere. International Journal of Molecular Sciences. 2021; 22(12):6356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126356
Chicago/Turabian StyleOszust, Karolina, Michał Pylak, and Magdalena Frąc. 2021. "Trichoderma-Based Biopreparation with Prebiotics Supplementation for the Naturalization of Raspberry Plant Rhizosphere" International Journal of Molecular Sciences 22, no. 12: 6356. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22126356