
Integrated Metabolomics and Proteomics Analyses in the Local Milieu of Islet Allografts in Rejection versus Tolerance

 , ,
, ,
Abstract
:1. Introduction
2. Results
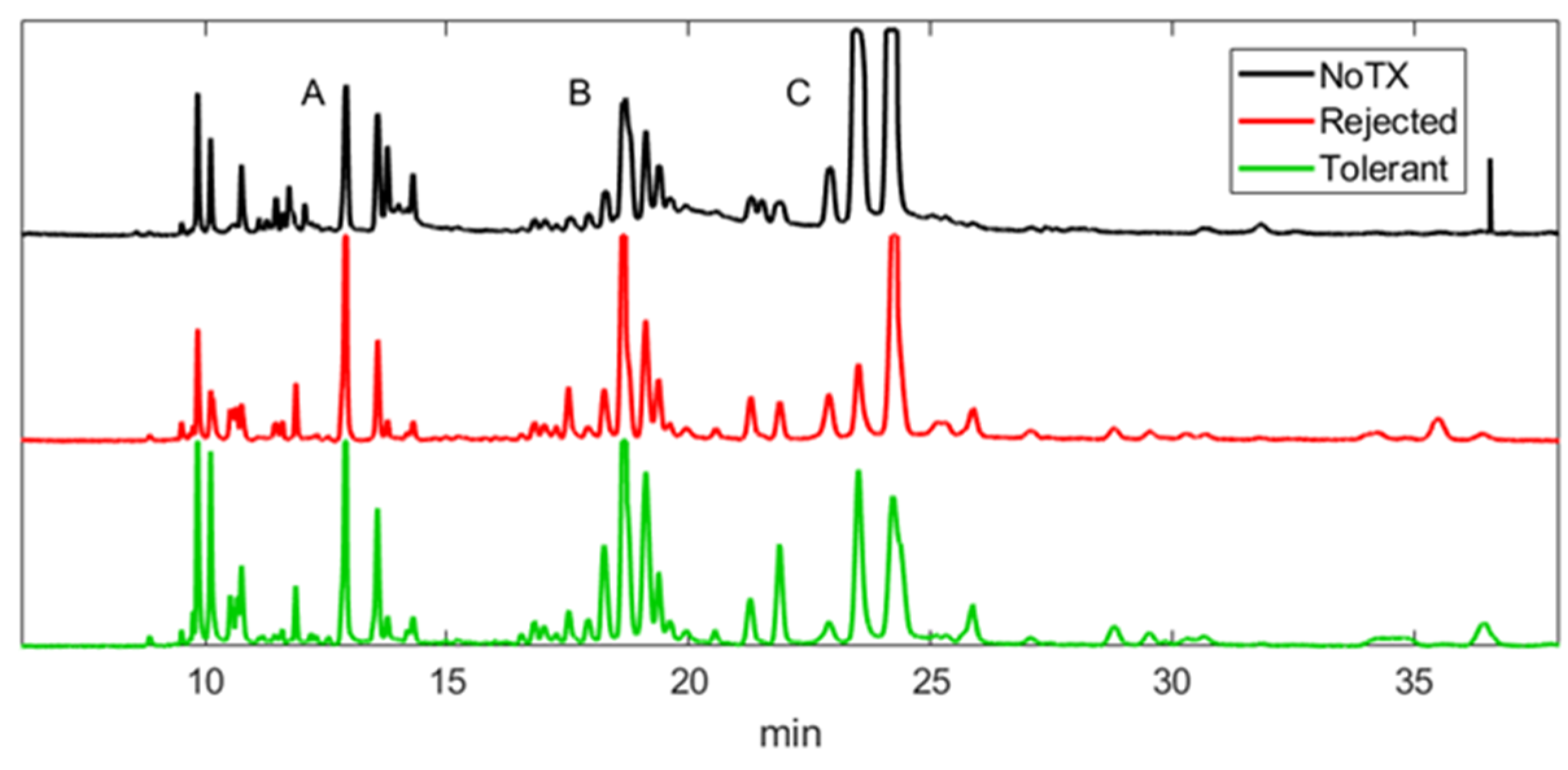
2.1. Patterns of Electropherograms Generated in Aqueous Humor Samples during Rejection versus Tolerance Are Significantly Different
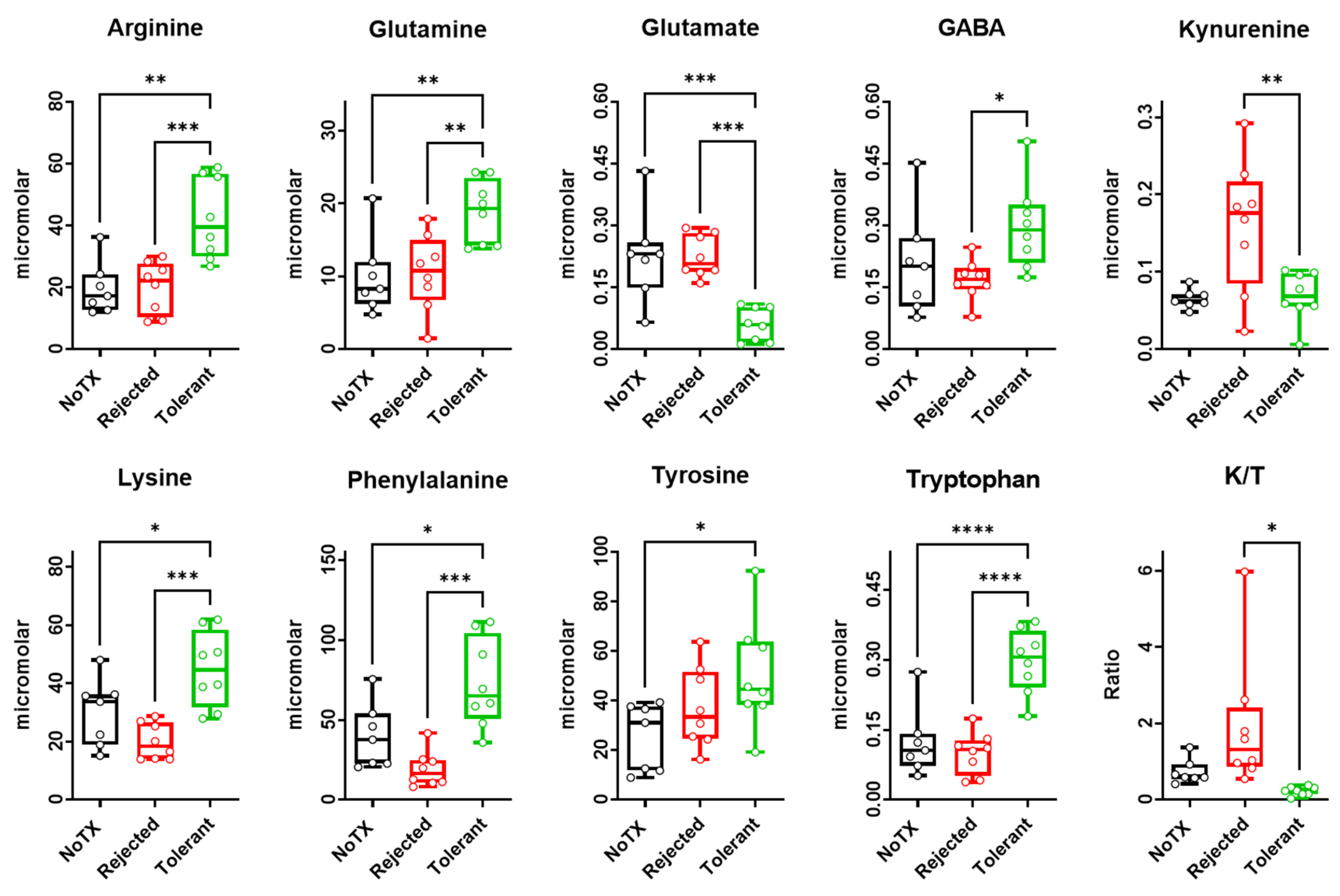
2.2. Free Amino Acids Levels in the Local Microenvironment of Rejected versus Tolerated Islet Allografts
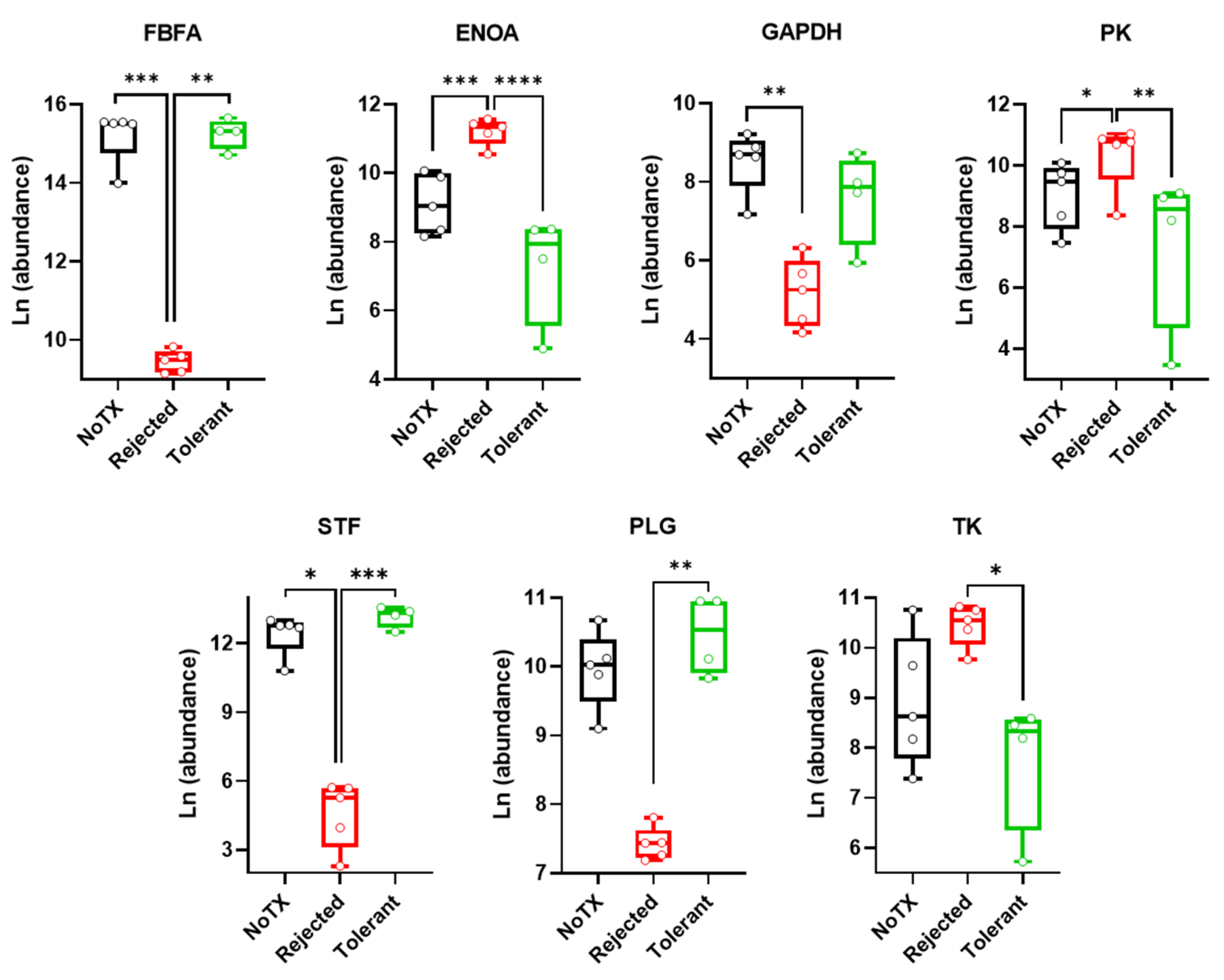
2.3. Warburg Effect-Related Protein Levels in the Local Microenvironment of Pancreatic Islet Allografts Are Significantly Different in Rejection versus Tolerance
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Islet Isolation and Transplantation
4.3. Monitoring the Survival and Rejection of the ACE-Transplanted Islets
4.4. Aqueous Humor (AQH) Samples
4.5. Sample Preparation and Metabolomics Analysis by MEKC-LIFD
4.6. Sample Preparation and Proteomics Analysis by LC-MS/MS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Warburg, O.; Wind, F.; Negelein, E. The metabolism of tumors in the body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Warburg, O.; Gawehn, K.; Geissler, A.W. Stoffwechsel der weißen Blutzellen. Z. Nat. B J. Chem. Sci. 1958, 13, 515–516. [Google Scholar] [CrossRef]
- Fukuzumi, M.; Shinomiya, H.; Shimizu, Y.; Ohishi, K.; Utsumi, S. Endotoxin-induced enhancement of glucose influx into murine peritoneal macrophages via GLUT 1. Infect. Immun. 1996, 64, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Rathmell, J.C.; Vander Heiden, M.G.; Harris, M.H.; Frauwirth, K.A.; Thompson, C.B. In the absence of extrinsic signals, nutrient utilization by lymphocytes is insufficient to maintain either cell size or viability. Mol. Cell 2000, 6, 683–692. [Google Scholar] [CrossRef]
- Kornberg, M.D. The immunologic Warburg effect: Evidence and therapeutic opportunities in autoimmunity. Wiley Interdiscip. Rev. Syst. Biol. Med. 2020, 12, e1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakaya, M.; Xiao, Y.; Zhou, X.; Chang, J.H.; Chang, M.; Cheng, X.; Blonska, M.; Lin, X.; Sun, S.C. Inflammatory T cell responses rely on amino acid transporter ASCT2 facilitation of glutamine uptake and mTORC1 kinase activation. Immunity 2014, 40, 692–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vats, D.; Mukundan, L.; Odegaard, J.I.; Zhang, L.; Smith, K.L.; Morel, C.R.; Greaves, D.R.; Murray, P.J.; Chawla, A. Oxidative metabolism and PGC-1β attenuate macrophage-mediated inflammation. Cell Metab. 2006, 4, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Hu, H.; Chang, R.; Zhong, J.; Knabel, M.; O’Meally, R.; Cole, R.N.; Pandey, A.; Semenza, G.L. Pyruvate kinase M2 is a PHD3-stimulated coactivator for hypoxia-inducible factor 1. Cell 2011, 145, 732–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palsson-Mcdermott, E.M.; Curtis, A.M.; Goel, G.; Lauterbach, M.A.R.; Sheedy, F.J.; Gleeson, L.E.; Van Den Bosch, M.W.M.; Quinn, S.R.; Domingo-Fernandez, R.; Johnson, D.G.W. Pyruvate kinase M2 regulates hif-1α activity and il-1β induction and is a critical determinant of the warburg effect in LPS-activated macrophages. Cell Metab. 2015, 21, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Shirai, T.; Nazarewicz, R.R.; Wallis, B.B.; Yanes, R.E.; Watanabe, R.; Hilhorst, M.; Tian, L.; Harrison, D.G.; Giacomini, J.C.; Assimes, T.L. The glycolytic enzyme PKM2 bridges metabolic and inflammatory dysfunction in coronary artery disease. J. Exp. Med. 2016, 213, 337–354. [Google Scholar] [CrossRef] [Green Version]
- Guillou, C.; Fréret, M.; Fondard, E.; Derambure, C.; Avenel, G.; Golinski, M.L.; Verdet, M.; Boyer, O.; Caillot, F.; Musette, P. Soluble alpha-enolase activates monocytes by CD14-dependent TLR4 signalling pathway and exhibits a dual function. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pioli, P.A.; Hamilton, B.J.; Connolly, J.E.; Brewer, G.; Rigby, W.F.C. Lactate dehydrogenase is an AU-rich element-binding protein that directly interacts with AUF1. J. Biol. Chem. 2002, 277, 35738–35745. [Google Scholar] [CrossRef] [Green Version]
- Lafferty, K.J.; Bootes, A.; Dart, G.; Talmage, D.W. Effect of organ culture on the survival of thyroid allografts in mice. Transplantation 1976, 22, 138–149. [Google Scholar] [CrossRef]
- Talmage, D.W.; Dart, G.; Radovich, J.; Lafferty, K.J. Activation of transplant immunity: Effect of donor leukocytes on thyroid allograft rejection. Science 1976, 191, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Ingulli, E. Mechanism of cellular rejection in transplantation. Pediatric Nephrol. 2010, 25, 61–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lechler, R.I.; Batchelor, J.R. Restoration of immunogenicity to passenger cell-depleted kidney allografts by the addition of donor strain dendritic cells. J. Exp. Med. 1982, 155, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Allocco, J.B.; Alegre, M.L. Exploiting immunometabolism and T cell function for solid organ transplantation. Cell. Immunol. 2020, 351, 104068. [Google Scholar] [CrossRef] [PubMed]
- Tanimine, N.; Turka, L.A.; Priyadharshini, B. Navigating T-Cell Immunometabolism in Transplantation. Transplantation 2018, 102, 230–239. [Google Scholar] [CrossRef]
- Kloc, M.; Ghobrial, R. Chronic allograft rejection: A significant hurdle to transplant success. Burn. Trauma 2014, 2, 2321–3868. [Google Scholar] [CrossRef] [Green Version]
- Abdulreda, M.H.; Caicedo, A.; Berggren, P.O. Transplantation into the anterior chamber of the eye for longitudinal, non-invasive in vivo imaging with single-cell resolution in real-time. J. Vis. Exp. 2013, 73, e50466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, V.L.; Caicedo, A.; Berman, D.M.; Arrieta, E.; Abdulreda, M.H.; Rodriguez-Diaz, R.; Pileggi, A.; Hernandez, E.; Dubovy, S.R.; Berggren, P.O.; et al. The anterior chamber of the eye as a clinical transplantation site for the treatment of diabetes: A study in a baboon model of diabetes. Diabetologia 2011, 54, 1121–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tun, S.B.B.; Chua, M.; Hasan, R.; Köhler, M.; Zheng, X.; Ali, Y.; Abdulreda, M.H.; Juntti-Berggren, L.; Barathi, V.A.; Berggren, P.O. Islet Transplantation to the Anterior Chamber of the Eye—A Future Treatment Option for Insulin-Deficient Type-2 Diabetics? A Case Report from a Nonhuman Type-2 Diabetic Primate. Cell Transplant. 2020, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shishido, A.; Caicedo, A.; Rodriguez-Diaz, R.; Pileggi, A.; Berggren, P.O.; Abdulreda, M.H. Clinical intraocular islet transplantation is not a number issue. CellR4 Repair Replace. Regen. Reprogram. 2016, 4, e2120. [Google Scholar] [PubMed]
- Alcazar, O.; Hernandez, L.F.; Tschiggfrie, A.; Muehlbauer, M.J.; Bain, J.R.; Buchwald, P.; Abdulreda, M.H. Feasibility of Localized Metabolomics in the Study of Pancreatic Islets and Diabetes. Metabolites 2019, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Alcazar, O.; Hernandez, L.F.; Tschiggfrie, A.; Muehlbauer, M.J.; Bain, J.R.; Buchwald, P.; Abdulreda, M.H. Longitudinal proteomics analysis in the immediate microenvironment of islet allografts during progression of rejection. J. Proteom. 2020, 223, 103826. [Google Scholar] [CrossRef]
- Ceballos, G.A.; Hernandez, L.F.; Paredes, D.; Betancourt, L.R.; Abdulreda, M.H. A machine learning approach to predict pancreatic islet grafts rejection versus tolerance. PLoS ONE 2020, 15, e0241925. [Google Scholar] [CrossRef]
- Hernandez, L.; Joshi, N.; Murzi, E.; Verdeguer, P.; Mifsud, J.C.; Guzman, N. Colinear laser-induced fluorescence detector for capillary electrophoresis. Analysis of glutamic acid in brain dialysates. J. Chromatogr. A 1993, 652, 399–405. [Google Scholar] [CrossRef]
- Hernandez, L.; Escalona, J.; Joshi, N.; Guzman, N. Laser-induced fluorescence and fluorescence microscopy for capillary electrophoresis zone detection. J. Chromatogr. A 1991, 559, 183–196. [Google Scholar] [CrossRef]
- Meng, Q.; Cooney, M.; Yepuri, N.; Cooney, R.N. L-arginine attenuates Interleukin-1β (IL-1β) induced nuclear factor kappa-beta (NF-κB) activation in Caco-2 cells. PLoS ONE 2017, 12, e0174441. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, F.; Peng, A.; Guo, S.; Wang, M.; Elsabagh, M.; Loor, J.J.; Wang, H. L-arginine inhibits apoptosis of ovine intestinal epithelial cells through the L-Arginine-nitric oxide pathway. J. Nutr. 2020, 150, 2051–2060. [Google Scholar] [CrossRef]
- Ryan, W.L.; Carver, M.J. Inhibition of antibody synthesis by L-phenylalanine. Science 1964, 143, 479–480. [Google Scholar] [CrossRef] [PubMed]
- Ryan, W.L. Inhibition of the Immune Response by Phenylalanine: Application to Skin Transplantation. JAMA J. Am. Med. Assoc. 1965, 191, 295–296. [Google Scholar] [CrossRef]
- Mao, Y.Y.; Bai, J.Q.; Chen, J.H.; Shou, Z.F.; He, Q.; Wu, J.Y.; Chen, Y.; Cheng, Y.Y. A pilot study of GC/MS-based serum metabolic profiling of acute rejection in renal transplantation. Transpl. Immunol. 2008, 19, 74–80. [Google Scholar] [CrossRef]
- Han, H.; Yin, J.; Wang, B.; Huang, X.; Yao, J.; Zheng, J.; Fan, W.; Li, T.; Yin, Y. Effects of dietary lysine restriction on inflammatory responses in piglets. Sci. Rep. 2018, 8, 2451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paßlack, N.; Doherr, M.G.; Zentek, J. Effects of free amino acids on cytokine secretion and proliferative activity of feline T cells in an in vitro study using the cell line MYA-1. Cytotechnology 2016, 68, 1949–1961. [Google Scholar] [CrossRef] [Green Version]
- Al-Malki, A.L. Suppression of acute pancreatitis by L-lysine in mice. BMC Complementary Altern. Med. 2015, 15, 193. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Yin, J.; Wang, B.; Huang, X.; Yao, J.; Zheng, J.; Fan, W.; Li, T.; Yin, Y.; Chen, Q.X.; et al. L-lysine confers neuroprotection by suppressing inflammatory response via microRNA-575/PTEN signaling after mouse intracerebral hemorrhage injury. Exp. Neurol. 2020, 327, 113214. [Google Scholar] [CrossRef]
- Wang, L.; Opland, D.; Tsai, S.; Luk, C.T.; Schroer, S.A.; Allison, M.B.; Elia, A.J.; Furlonger, C.; Suzuki, A.; Paige, C.J.; et al. Pten deletion in RIP-Cre neurons protects against type 2 diabetes by activating the anti-inflammatory reflex. Nat. Med. 2014, 20, 484–492. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, J.; Ye, L.; Xu, G. Serum metabolomics study of the acute graft rejection in human renal transplantation based on liquid chromatography-mass spectrometry. J. Proteome Res. 2014, 13, 2659–2667. [Google Scholar] [CrossRef]
- Günther, J.; Fallarino, F.; Fuchs, D.; Wirthgen, E. Immunomodulatory Roles of Tryptophan Metabolites in Inflammation and Cancer. Front. Immunol. 2020, 11, 1497. [Google Scholar] [CrossRef]
- Almaça, J.; Molina, J.; Menegaz, D.; Pronin, A.N.; Tamayo, A.; Slepak, V.; Berggren, P.O.; Caicedo, A. Human Beta Cells Produce and Release Serotonin to Inhibit Glucagon Secretion from Alpha Cells. Cell Rep. 2016, 17, 3281–3291. [Google Scholar] [CrossRef] [Green Version]
- Bascuñan, L.R.C.; Lyons, C.; Bennet, H.; Artner, I.; Fex, M. Serotonergic Regulation of Insulin Secretion. Acta Physiol. 2019, 225, e13101. [Google Scholar] [CrossRef]
- Boadle-Biber, M.C. Regulation of serotonin synthesis. Prog. Biophys. Mol. Biol. 1993, 60, 1–15. [Google Scholar] [CrossRef]
- Calder, P.C.; Yaqoob, P. Glutamine and the immune system. Amino Acids 1999, 17, 227–241. [Google Scholar] [CrossRef]
- Yaqoob, P.; Calder, P.C. Cytokine production by human peripheral blood mononuclear cells: Differential sensitivity to glutamine availability. Cytokine 1998, 10, 790–794. [Google Scholar] [CrossRef]
- Jain, P.; Khanna, N.K. Evaluation of anti-inflammatory and analgesic properties of l-glutamine. Agents Actions 1981, 11, 243–249. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, H. The roles of glutamine in the intestine and its implication in intestinal diseases. Int. J. Mol. Sci. 2017, 18, 1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalek, R.D.; Valerie, A.G.; Sarah, R.J.; Andrew, N.M.; Nancie, J.M.; Emily, F.M.; Sarah, A.S.; Amanda, G.N.; Jeffrey, C.R. Cutting Edge: Distinct Glycolytic and Lipid Oxidative Metabolic Programs Are Essential for Effector and Regulatory CD4 + T Cell Subsets. J. Immunol. 2011, 186, 3299–3303. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Dillon, C.P.; Shi, L.Z.; Milasta, S.; Carter, R.; Finkelstein, D.; McCormick, L.L.; Fitzgerald, P.; Chi, H.; Munger, J.; et al. The Transcription Factor Myc Controls Metabolic Reprogramming upon T Lymphocyte Activation. Immunity 2011, 35, 871–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, M.J. Influence of glucose on pyruvate carboxylase expression in pancreatic islets. Arch. Biochem. Biophys. 1995, 319, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, M.; Morimoto, R.; Yamamoto, A.; Moriyama, Y. Expression and localization of vesicular glutamate transporters in pancreatic islets, upper gastrointestinal tract, and testis. J. Histochem. Cytochem. 2003, 51, 1375–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, O.; Jacques-Silva, M.C.; Speier, S.; Yang, S.N.; Köhler, M.; Fachado, A.; Vieira, E.; Zierath, J.R.; Kibbey, R.; Berman, D.M.; et al. Glutamate Is a Positive Autocrine Signal for Glucagon Release. Cell Metab. 2008, 7, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otter, S.; Lammert, E. Exciting Times for Pancreatic Islets: Glutamate Signaling in Endocrine Cells. Trends Endocrinol. Metab. 2016, 27, 177–188. [Google Scholar] [CrossRef]
- Drucker, D.J.; Nauck, M.A. The incretin system: Glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes. Lancet 2006, 368, 1696–1705. [Google Scholar] [CrossRef]
- Wakabayashi, Y.; Yagihashi, T.; Kezuka, J.; Muramatsu, D.; Usui, M.; Iwasaki, T. Glutamate levels in aqueous humor of patients with retinal artery occlusion. Retina 2006, 26, 432–436. [Google Scholar] [CrossRef]
- Davalli, A.M.; Perego, C.; Folli, F.B. The potential role of glutamate in the current diabetes epidemic. Acta Diabetol. 2012, 49, 167–183. [Google Scholar] [CrossRef]
- Auteri, M.; Zizzo, M.G.; Serio, R. GABA and GABA receptors in the gastrointestinal tract: From motility to inflammation. Pharmacol. Res. 2015, 93, 11–21. [Google Scholar] [CrossRef]
- Jin, Z.; Mendu, S.K.; Birnir, B. GABA is an effective immunomodulatory molecule. Amino Acids 2013, 45, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, H.; Okada, Y.; Seguchi, H.; Shimada, C.; Seki, M.; Tsutou, A.; Baba, S. High concentration of gamma-aminobutyric acid in pancreatic beta cells. Diabetes 1979, 28, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Garry, D.J.; Sorenson, R.L.; Elde, R.P.; Maley, B.E.; Madsen, A. Immunohistochemical colocalization of GABA and insulin in β-cells of rat islet. Diabetes 1986, 35, 1090–1095. [Google Scholar] [CrossRef]
- Garry, D.J.; Coulter, H.D.; McIntee, T.J.; Wu, J.Y.; Sorenson, R.L. Immunoreactive GABA transaminase within the pancreatic islet is localized in mitochondria of the B-cell. J. Histochem. Cytochem. 1987, 35, 831–836. [Google Scholar] [CrossRef]
- Adeghate, E.; Ponery, A.S. GABA in the endocrine pancreas: Cellular localization and function in normal and diabetic rats. Tissue Cell 2002, 34, 1–6. [Google Scholar] [CrossRef]
- Solimena, M.; Folli, F.; Denis-Donini, S.; Comi, G.C.; Pozza, G.; De Camilli, P.; Vicari, A.M. Autoantibodies to Glutamic Acid Decarboxylase in a Patient with Stiff-Man Syndrome, Epilepsy, and Type I Diabetes Mellitus. N. Engl. J. Med. 1988, 318, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Solimena, M.; Folli, F.; Aparisi, R.; Pozza, G.; de Camilli, P. Autoantibodies to GABA-ergic Neurons and Pancreatic Beta Cells in Stiff-Man Syndrome. N. Engl. J. Med. 1990, 322, 1555–1560. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wang, Q.; Prud’homme, G.J. GABAergic system in the endocrine pancreas: A new target for diabetes treatment. Diabetes Metab. Syndr. Obes. Targets Ther. 2015, 8, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Petty, F.; Kramer, G.; Feldman, M. Is plasma GABA of peripheral origin? Biol. Psychiatry 1987, 22, 725–732. [Google Scholar] [CrossRef]
- Yazulla, S.; Studholme, K.M.; Wu, J.Y. GABAergic input to the synaptic terminals of mb1 bipolar cells in the goldfish retina. Brain Res. 1987, 411, 400–405. [Google Scholar] [CrossRef]
- Yazulla, S.; Yang, C.Y. Colocalization of GABA and glycine immunoreactivities in a subset of retinal neurons in tiger salamander. Neurosci. Lett. 1988, 95, 37–41. [Google Scholar] [CrossRef]
- Abdulreda, M.H.; Rodriguez-Diaz, R.; Cabrera, O.; Caicedo, A.; Berggren, P.O. The Different Faces of the Pancreatic Islet. Adv. Exp. Med. Biol. 2016, 938, 11–24. [Google Scholar] [CrossRef]
- Wang, Q.; Ren, L.; Wan, Y.; Prud’homme, G.J. GABAergic regulation of pancreatic islet cells: Physiology and antidiabetic effects. J. Cell. Physiol. 2019, 234, 14432–14444. [Google Scholar] [CrossRef]
- Rodriguez-Diaz, R.; Menegaz, D.; Caicedo, A. Neurotransmitters act as paracrine signals to regulate insulin secretion from the human pancreatic islet. J. Physiol. 2014, 592, 3413–3417. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Kumar, M.; Zhang, Y.; Gyulkhandanyan, A.; Xiang, Y.Y.; Ye, B.; Perrella, J.; Hyder, A.; Zhang, N.; Wheeler, M.; et al. Gamma-aminobutyric acid up- and downregulates insulin secretion from beta cells in concert with changes in glucose concentration. Diabetologia 2006, 49, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Xu, E.; Kumar, M.; Zhang, Y.; Ju, W.; Obata, T.; Zhang, N.; Liu, S.; Wendt, A.; Deng, S.; Ebina, Y.; et al. Intra-islet insulin suppresses glucagon release via GABA-GABAA receptor system. Cell Metab. 2006, 3, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Prud’Homme, G.J.; Glinka, Y.; Hasilo, C.; Paraskevas, S.; Li, X.; Wang, Q. GABA protects human islet cells against the deleterious effects of immunosuppressive drugs and exerts immunoinhibitory effects alone. Transplantation 2013, 96, 616–623. [Google Scholar] [CrossRef]
- Purwana, I.; Zheng, J.; Li, X.; Deurloo, M.; Son, D.O.; Zhang, Z.; Liang, C.; Shen, E.; Tadkase, A.; Feng, Z.P.; et al. GABA promotes human β-cell proliferation and modulates glucose homeostasis. Diabetes 2014, 63, 4197–4205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben-Othman, N.; Vieira, A.; Courtney, M.; Record, F.; Gjernes, E.; Avolio, F.; Hadzic, B.; Druelle, N.; Napolitano, T.; Navarro-Sanz, S.; et al. Long-Term GABA Administration Induces Alpha Cell-Mediated Beta-like Cell Neogenesis. Cell 2017, 168, 73–85. [Google Scholar] [CrossRef] [Green Version]
- Abdulreda, M.H.; Berman, D.M.; Shishido, A.; Martin, C.; Hossameldin, M.; Tschiggfrie, A.; Hernandez, L.F.; Hernandez, A.; Ricordi, C.; Parel, J.M.; et al. Operational immune tolerance towards transplanted allogeneic pancreatic islets in mice and a non-human primate. Diabetologia 2019, 62, 811–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miska, J.; Abdulreda, M.H.; Devarajan, P.; Lui, J.B.; Suzuki, J.; Pileggi, A.; Berggren, P.O.; Chen, Z. Real-time immune cell interactions in target tissue during autoimmune-induced damage and graft tolerance. J. Exp. Med. 2014, 211, 441–456. [Google Scholar] [CrossRef]
- Liu, W.; Son, D.O.; Lau, H.K.; Zhou, Y.; Prud’homme, G.J.; Jin, T.; Wang, Q. Combined oral administration of GABA and DPP-4 inhibitor prevents beta cell damage and promotes beta cell regeneration in mice. Front. Pharmacol. 2017, 8, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, K.; Wang, B.; Zhou, H.; Luo, Q.; Shen, J.; Xu, Y.; Zhong, Z. Amelioration of type 1 diabetes by recombinant fructose-1,6-bisphosphate aldolase and cystatin derived from Schistosoma japonicum in a murine model. Parasitol. Res. 2020, 119, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Kim, H.; Lee, N.; Won, C.; Kim, H.-R.; Hwang, Y.; Song, Y.W.; Kang, J.S.; Lee, W.J. α-Enolase Expressed on the Surfaces of Monocytes and Macrophages Induces Robust Synovial Inflammation in Rheumatoid Arthritis. J. Immunol. 2012, 189, 365–372. [Google Scholar] [CrossRef]
- Takaoka, Y.; Goto, S.; Nakano, T.; Tseng, H.P.; Yang, S.M.; Kawamoto, S.; Ono, K.; Chen, C.L. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) prevents lipopolysaccharide (LPS)-induced, sepsis-related severe acute lung injury in mice. Sci. Rep. 2014, 4, 5204. [Google Scholar] [CrossRef] [Green Version]
- Claeyssens, S.; Gangneux, C.; Brasse-Lagnel, C.; Ruminy, P.; Aki, T.; Lavoinne, A.; Salier, J.P. Amino acid control of the human glyceraldehyde 3-phosphate dehydrogenase gene transcription in hepatocyte. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G840–G849. [Google Scholar] [CrossRef]
- Shao, W.; Yeretssian, G.; Doiron, K.; Hussain, S.N.; Saleh, M. The caspase-1 digestome identifies the glycolysis pathway as a target during infection and septic shock. J. Biol. Chem. 2007, 282, 36321–36329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanman, L.E.; Qian, Y.; Eisele, N.A.; Ng, T.M.; van der Linden, W.A.; Monack, D.M.; Weerapana, E.; Bogyo, M. Disruption of glycolytic flux is a signal for inflammasome signaling and pyroptotic cell death. Elife 2016, 5, e13663. [Google Scholar] [CrossRef]
- Ferguson, E.C.; Rathmell, J.C. New roles for pyruvate kinase M2: Working out the Warburg effect. Trends Biochem. Sci. 2008, 33, 359–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christofk, H.R.; Heiden, M.G.V.; Wu, N.; Asara, J.M.; Cantley, L.C. Pyruvate kinase M2 is a phosphotyrosine-binding protein. Nature 2008, 452, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Christofk, H.R.; Vander Heiden, M.G.; Harris, M.H.; Ramanathan, A.; Gerszten, R.E.; Wei, R.; Fleming, M.D.; Schreiber, S.L.; Cantley, L.C. The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth. Nature 2008, 452, 230–233. [Google Scholar] [CrossRef]
- Anastasiou, D.; Yu, Y.; Israelsen, W.J.; Jiang, J.K.; Boxer, M.B.; Hong, B.S.; Tempel, W.; Dimov, S.; Shen, M.; Jha, A.; et al. Pyruvate kinase M2 activators promote tetramer formation and suppress tumorigenesis. Nat. Chem. Biol. 2012, 8, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.k.; Boxer, M.B.; Vander Heiden, M.G.; Shen, M.; Skoumbourdis, A.P.; Southall, N.; Veith, H.; Leister, W.; Austin, C.P.; Park, H.W.; et al. Evaluation of thieno[3,2-b]pyrrole[3,2-d]pyridazinones as activators of the tumor cell specific M2 isoform of pyruvate kinase. Bioorganic Med. Chem. Lett. 2010, 20, 3387–3393. [Google Scholar] [CrossRef] [Green Version]
- Angiari, S.; Runtsch, M.C.; Sutton, C.E.; Palsson-McDermott, E.M.; Kelly, B.; Rana, N.; Kane, H.; Papadopoulou, G.; Pearce, E.L.; Mills, K.H.G.; et al. Pharmacological Activation of Pyruvate Kinase M2 Inhibits CD4+ T Cell Pathogenicity and Suppresses Autoimmunity. Cell Metab. 2020, 31, 391–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fletcher, J.; Huehns, E.R. Function of transferrin. Nature 1968, 218, 1211–1214. [Google Scholar] [CrossRef] [PubMed]
- Trowbridge, I.S.; Omary, M.B. Human cell surface glycoprotein related to cell proliferation is the receptor for transferrin. Proc. Natl. Acad. Sci. USA 1981, 78, 3039–3043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponka, P.; Lok, C.N. The transferrin receptor: Role in health and disease. Int. J. Biochem. Cell Biol. 1999, 31, 1111–1137. [Google Scholar] [CrossRef]
- Nairz, M.; Theurl, I.; Swirski, F.K.; Weiss, G. “Pumping iron”—How macrophages handle iron at the systemic, microenvironmental, and cellular levels. Pflug. Arch. 2017, 469, 397–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Pantopoulos, K. Regulation of cellular iron metabolism. Biochem. J. 2011, 434, 365–381. [Google Scholar] [CrossRef] [Green Version]
- Solomon, E.I.; Decker, A.; Lehnert, N. Non-heme iron enzymes: Contrasts to heme catalysis. Proc. Natl. Acad. Sci. USA 2003, 100, 3589–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Gong, J.; Liu, Y. Indoleamine 2,3-dioxygenase regulation of immune response. Mol. Med. Rep. 2018, 17, 4867–4873. [Google Scholar] [CrossRef] [Green Version]
- Wittig, R.; Coy, J.F. The role of glucose metabolism and glucose-associated signalling in cancer. Perspect. Med. Chem. 2008, 1, 64–82. [Google Scholar] [CrossRef]
- Vago, J.P.; Sugimoto, M.A.; Lima, K.M.; Negreiros-Lima, G.L.; Baik, N.; Teixeira, M.M.; Perretti, M.; Parmer, R.J.; Miles, L.A.; Sousa, L.P. Plasminogen and the Plasminogen Receptor, Plg-Rkt, regulate macrophage phenotypic and functional changes. Front. Immunol. 2019, 10, 1458. [Google Scholar] [CrossRef] [PubMed]
- Pileggi, A.; Molano, R.D.; Berney, T.; Cattan, P.; Vizzardelli, C.; Oliver, R.; Fraker, C.; Ricordi, C.; Pastori, R.L.; Bach, F.H.; et al. Heme oxygenase-1 induction in islet cells results in protection from apoptosis and improved in vivo function after transplantation. Diabetes 2001, 50, 1983–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulreda, M.H.; Faleo, G.; Molano, R.D.; Lopez-Cabezas, M.; Molina, J.; Tan, Y.; Echeverria, O.A.; Zahr-Akrawi, E.; Rodriguez-Diaz, R.; Edlund, P.K.; et al. High-resolution, noninvasive longitudinal live imaging of immune responses. Proc. Natl. Acad. Sci. USA 2011, 108, 12863–12868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwanhüusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.; Matic, I.; Hilger, M.; Nagaraj, N.; Selbach, M.; Olsen, J.V.; Mann, M. A practical guide to the maxquant computational platform for silac-based quantitative proteomics. Nat. Protoc. 2009, 4, 698–705. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Metabolite | F(2/22) | R Squared | p-Value | Tol vs. Rej | Tol vs. NoTX | Rej vs. NoTX |
|---|---|---|---|---|---|---|
| Arginine | 12.32 | 0.552 | 0.0003 | *** | ** | ns |
| Glutamine | 7.784 | 0.4377 | 0.0032 | ** | ** | ns |
| Glutamate | 13.55 | 0.5755 | 0.0002 | *** | *** | ns |
| GABA | 3.744 | 0.2724 | 0.0416 | * | ns | ns |
| Kynurenine | 7.685 | 0.4354 | 0.0033 | ** | ns | ** |
| Lysine | 11 | 0.5238 | 0.0006 | *** | * | ns |
| Phenylalanine | 13.33 | 0.5713 | 0.0002 | *** | * | ns |
| Tyrosine | 3.685 | 0.2693 | 0.0434 | ns | * | ns |
| Tryptophan | 22.78 | 0.6949 | <0.0001 | **** | **** | ns |
| K/T ratio | 5.336 | 0.3479 | 0.0139 | * | ns | ns |
| Protein | F(2/11) | R Squared | p-Value | Tol vs. Rej | Tol vs. NoTX | Rej vs. NoTX |
|---|---|---|---|---|---|---|
| Fructose-bisphosphate aldolase A (FBFA) | 16.28 | 0.7475 | 0.0005 | ** | ns | *** |
| Alphaenolase (ENOA) | 29.7 | 0.8438 | <0.0001 | *** | ns | **** |
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | 7.962 | 0.5915 | 0.0073 | ns | ns | ** |
| Pyruvate kinase (PK) | 8.484 | 0.6067 | 0.0059 | ** | ns | * |
| Serotransferrin (STF) | 17.3 | 0.7588 | 0.0004 | *** | ns | * |
| Plasminogen (PLG) | 9.401 | 0.6309 | 0.0042 | ** | ns | ns |
| Transketolase (TK) | 6.951 | 0.5583 | 0.0112 | * | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez, L.F.; Betancourt, L.R.; Nakayasu, E.S.; Ansong, C.; Ceballos, G.A.; Paredes, D.; Abdulreda, M.H. Integrated Metabolomics and Proteomics Analyses in the Local Milieu of Islet Allografts in Rejection versus Tolerance. Int. J. Mol. Sci. 2021, 22, 8754. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168754
Hernandez LF, Betancourt LR, Nakayasu ES, Ansong C, Ceballos GA, Paredes D, Abdulreda MH. Integrated Metabolomics and Proteomics Analyses in the Local Milieu of Islet Allografts in Rejection versus Tolerance. International Journal of Molecular Sciences. 2021; 22(16):8754. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168754
Chicago/Turabian StyleHernandez, Luis F., Luis R. Betancourt, Ernesto S. Nakayasu, Charles Ansong, Gerardo A. Ceballos, Daniel Paredes, and Midhat H. Abdulreda. 2021. "Integrated Metabolomics and Proteomics Analyses in the Local Milieu of Islet Allografts in Rejection versus Tolerance" International Journal of Molecular Sciences 22, no. 16: 8754. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22168754