
Melatonin Inhibits Osteoclastogenesis and Osteolytic Bone Metastasis: Implications for Osteoporosis

 ,
,  and
and
Abstract
:1. Introduction
2. Melatonin Metabolically Reprograms HSPC Self-Renewal, Stimulates Osteoblast Differentiation, and Inhibits Osteoclastogenesis
3. Melatonin Shows Potential for Osteoporosis—Preclinical and Clinical Evidence
4. Melatonin Shows Potential in Bone Cancer Treatment and Osteolytic Bone Metastasis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AC | Adenylate cyclase |
| BM | Bone marrow |
| BMD | Bone mineral density |
| BV/TV | Bone volume/tissue volume |
| Ca2+ | Calcium |
| cAMP | Cyclic adenosine monophosphate |
| cGMP | Cyclic guanosine monophosphate |
| CREB | cAMP response element-binding protein |
| DAG | Diacylglycerol |
| EAE | Experimental autoimmune encephalomyelitis |
| GC | Guanylate cyclase |
| HSPCs | Hematopoietic stem and progenitor cells |
| IL | Interleukin |
| IP3 | Inositol triphosphate |
| LAN | Light at night |
| MAPK | Mitogen-activated protein kinase |
| micro-CT | Micro-computed tomography |
| MS | Multiple sclerosis |
| MSCs | Mesenchymal stem cells |
| MSDK | Melatonin, strontium (citrate), vitamin D3 and vitamin K2 |
| MT1/2 | High-affinity G-protein-coupled melatonin receptors |
| OCN | Osteocalcin |
| NFATc1 | Nuclear factor of activated T cell cytoplasmic 1 |
| NF-κB | Nuclear factor kappa B |
| NTX | N-terminal telopeptide |
| PKA | Protein kinase A |
| PKG | Protein kinase G |
| PLC | Phospholipase C |
| PPARγ | Peroxisome proliferator-activated receptor gamma |
| RANKL | Receptor activator of nuclear factor-kappa B ligand |
| ROR | Retinoid-related orphan receptor |
| s.c. | Subcutaneous |
| SMI | Structure Model Index |
| Tb.N | Trabecular number |
| Tb.Sp | Trabecular Separation/Spacing |
| Tb.Th | Trabecular thickness |
| TNF | Tumor necrosis factor |
| Wk | Week |
References
- Bonmati-Carrion, M.A.; Tomas-Loba, A. Melatonin and Cancer: A Polyhedral Network Where the Source Matters. Antioxidants 2021, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef] [PubMed]
- Cipolla-Neto, J.; Amaral, F.G.D. Melatonin as a Hormone: New Physiological and Clinical Insights. Endocr. Rev. 2018, 39, 990–1028. [Google Scholar] [CrossRef] [Green Version]
- Ferlazzo, N.; Andolina, G.; Cannata, A.; Costanzo, M.G.; Rizzo, V.; Curro, M.; Ientile, R.; Caccamo, D. Is Melatonin the Cornucopia of the 21st Century? Antioxidants 2020, 9, 1088. [Google Scholar] [CrossRef] [PubMed]
- Hisatsune, C.; Nakamura, K.; Kuroda, Y.; Nakamura, T.; Mikoshiba, K. Amplification of Ca2+ signaling by diacylglycerol-mediated inositol 1,4,5-trisphosphate production. J. Biol. Chem. 2005, 280, 11723–11730. [Google Scholar] [CrossRef] [Green Version]
- Swulius, M.T.; Waxham, M.N. Ca (2+)/calmodulin-dependent protein kinases. Cell Mol. Life Sci. 2008, 65, 2637–2657. [Google Scholar] [CrossRef] [Green Version]
- Nagahama, T.; Hayashi, K.; Ozawa, Y.; Takenaka, T.; Saruta, T. Role of protein kinase C in angiotensin II-induced constriction of renal microvessels. Kidney Int. 2000, 57, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangel, L.B.; Caruso-Neves, C.; Lara, L.S.; Lopes, A.G. Angiotensin II stimulates renal proximal tubule Na(+)-ATPase activity through the activation of protein kinase C. Biochim. Biophys. Acta 2002, 1564, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Hattangady, N.G.; Olala, L.O.; Bollag, W.B.; Rainey, W.E. Acute and chronic regulation of aldosterone production. Mol. Cell Endocrinol. 2012, 350, 151–162. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, I.J.; Huang, C.C.; Liu, S.C.; Tang, C.H. Reconsidering the Role of Melatonin in Rheumatoid Arthritis. Int. J. Mol. Sci. 2020, 21, 2877. [Google Scholar] [CrossRef]
- Liu, P.I.; Chang, A.C.; Lai, J.L.; Lin, T.H.; Tsai, C.H.; Chen, P.C.; Jiang, Y.J.; Lin, L.W.; Huang, W.C.; Yang, S.F.; et al. Melatonin interrupts osteoclast functioning and suppresses tumor-secreted RANKL expression: Implications for bone metastases. Oncogene 2021, 40, 1503–1515. [Google Scholar] [CrossRef]
- Foley, H.M.; Steel, A.E. Adverse events associated with oral administration of melatonin: A critical systematic review of clinical evidence. Complement. Ther. Med. 2019, 42, 65–81. [Google Scholar] [CrossRef]
- Seabra, M.L.; Bignotto, M.; Pinto, L.R., Jr.; Tufik, S. Randomized, double-blind clinical trial, controlled with placebo, of the toxicology of chronic melatonin treatment. J. Pineal Res. 2000, 29, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, L.P.; Gogenur, I.; Rosenberg, J.; Reiter, R.J. The Safety of Melatonin in Humans. Clin. Drug Investig. 2016, 36, 169–175. [Google Scholar] [CrossRef]
- Golan, K.; Kumari, A.; Kollet, O.; Khatib-Massalha, E.; Subramaniam, M.D.; Ferreira, Z.S.; Avemaria, F.; Rzeszotek, S.; Garcia-Garcia, A.; Xie, S.; et al. Daily Onset of Light and Darkness Differentially Controls Hematopoietic Stem Cell Differentiation and Maintenance. Cell Stem Cell 2018, 23, 572–585.e577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golan, K.; Kollet, O.; Markus, R.P.; Lapidot, T. Daily light and darkness onset and circadian rhythms metabolically synchronize hematopoietic stem cell differentiation and maintenance: The role of bone marrow norepinephrine, tumor necrosis factor, and melatonin cycles. Exp. Hematol. 2019, 78, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Kim, Y.-M.; Kim, H.S.; Lee, K.Y. Melatonin promotes osteoblast differentiation by regulating Osterix protein stability and expression. Sci. Rep. 2017, 7, 5716. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jang, A.R.; Park, M.J.; Kim, D.I.; Park, J.H. Melatonin Inhibits Osteoclastogenesis and Bone Loss in Ovariectomized Mice by Regulating PRMT1-Mediated Signaling. Endocrinology 2021, 162. [Google Scholar] [CrossRef]
- Zhou, L.; Chen, X.; Yan, J.; Li, M.; Liu, T.; Zhu, C.; Pan, G.; Guo, Q.; Yang, H.; Pei, M.; et al. Melatonin at pharmacological concentrations suppresses osteoclastogenesis via the attenuation of intracellular ROS. Osteoporos. Int. 2017, 28, 3325–3337. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, L.; Wang, Z.; Li, C.; Li, S.; Li, L.; Fan, Q.; Zheng, L. Melatonin Suppresses Estrogen Deficiency-Induced Osteoporosis and Promotes Osteoblastogenesis by Inactivating the NLRP3 Inflammasome. Calcif. Tissue Int. 2018, 103, 400–410. [Google Scholar] [CrossRef]
- Lee, S.; Le, N.H.; Kang, D. Melatonin alleviates oxidative stress-inhibited osteogenesis of human bone marrow-derived mesenchymal stem cells through AMPK activation. Int. J. Med. Sci. 2018, 15, 1083–1091. [Google Scholar] [CrossRef] [Green Version]
- Maria, S.; Samsonraj, R.M.; Munmun, F.; Glas, J.; Silvestros, M.; Kotlarczyk, M.P.; Rylands, R.; Dudakovic, A.; van Wijnen, A.J.; Enderby, L.T.; et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: Implications of a role for MT2 melatonin receptors, MEK1/2, and MEK5 in melatonin-mediated osteoblastogenesis. J. Pineal Res. 2018, 64, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Su, P.; Xu, C.; Chen, C.; Liang, A.; Du, K.; Peng, Y.; Huang, D. Melatonin inhibits adipogenesis and enhances osteogenesis of human mesenchymal stem cells by suppressing PPARgamma expression and enhancing Runx2 expression. J. Pineal Res. 2010, 49, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Rhee, Y.H.; Ahn, J.C. Melatonin attenuated adipogenesis through reduction of the CCAAT/enhancer binding protein beta by regulating the glycogen synthase 3 beta in human mesenchymal stem cells. J. Physiol. Biochem. 2016, 72, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Alvarez-Garcia, V.; Martinez-Campa, C.; Alonso-Gonzalez, C.; Cos, S. Melatonin promotes differentiation of 3T3-L1 fibroblasts. J. Pineal Res. 2012, 52, 12–20. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, H.J.; Bae, M.K.; Kim, Y.D. Suppression of Osteoclastogenesis by Melatonin: A Melatonin Receptor-Independent Action. Int. J. Mol. Sci. 2017, 18, 1142. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Gong, Z.; Zhao, R.; Zhu, Y. Melatonin inhibits RANKL-induced osteoclastogenesis through the miR-882/Rev-erbα axis in Raw264.7 cells. Int. J. Mol. Med. 2021, 47, 633–642. [Google Scholar] [CrossRef]
- Ikegame, M.; Hattori, A.; Tabata, M.J.; Kitamura, K.I.; Tabuchi, Y.; Furusawa, Y.; Maruyama, Y.; Yamamoto, T.; Sekiguchi, T.; Matsuoka, R.; et al. Melatonin is a potential drug for the prevention of bone loss during space flight. J. Pineal Res. 2019, 67, e12594. [Google Scholar] [CrossRef]
- Thenmozhi, A.; Nagalakshmi, K.; Shila, S.; Rasappan, P. Bone Restoration in Diabetic Osteolysis and Therapeutic Targets. Diabetes Case Rep. 2017, 2, 1000132. [Google Scholar] [CrossRef] [Green Version]
- Jahanban-Esfahlan, R.; Mehrzadi, S.; Reiter, R.J.; Seidi, K.; Majidinia, M.; Baghi, H.B.; Khatami, N.; Yousefi, B.; Sadeghpour, A. Melatonin in regulation of inflammatory pathways in rheumatoid arthritis and osteoarthritis: Involvement of circadian clock genes. Br. J. Pharmacol. 2018, 175, 3230–3238. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.C.; Chiou, C.H.; Liu, S.C.; Hu, S.L.; Su, C.M.; Tsai, C.H.; Tang, C.H. Melatonin attenuates TNF-alpha and IL-1beta expression in synovial fibroblasts and diminishes cartilage degradation: Implications for the treatment of rheumatoid arthritis. J. Pineal Res. 2019, 66, e12560. [Google Scholar] [CrossRef]
- Coyle, D. Cost-Effectiveness of Pharmacological Treatments for Osteoporosis Consistent with the Revised Economic Evaluation Guidelines for Canada. MDM Policy Pract. 2019, 4, 2381468318818843. [Google Scholar] [CrossRef] [Green Version]
- Tarantino, U.; Saturnino, L.; Scialdoni, A.; Feola, M.; Liuni, F.M.; Tempesta, V.; Pistillo, P. Fracture healing in elderly patients: New challenges for antiosteoporotic drugs. Aging Clin. Exp. Res. 2013, 25 (Suppl. 1), S105–S108. [Google Scholar] [CrossRef]
- Zacchetti, G.; Dayer, R.; Rizzoli, R.; Ammann, P. Systemic treatment with strontium ranelate accelerates the filling of a bone defect and improves the material level properties of the healing bone. BioMed Res. Int. 2014, 2014, 549785. [Google Scholar] [CrossRef]
- Lian, C.; Wu, Z.; Gao, B.; Peng, Y.; Liang, A.; Xu, C.; Liu, L.; Qiu, X.; Huang, J.; Zhou, H.; et al. Melatonin reversed tumor necrosis factor-alpha-inhibited osteogenesis of human mesenchymal stem cells by stabilizing SMAD1 protein. J. Pineal Res. 2016, 61, 317–327. [Google Scholar] [CrossRef]
- Jin, Y.Z.; Lee, J.H.; Xu, B.; Cho, M. Effect of medications on prevention of secondary osteoporotic vertebral compression fracture, non-vertebral fracture, and discontinuation due to adverse events: A meta-analysis of randomized controlled trials. BMC Musculoskelet. Disord. 2019, 20, 399. [Google Scholar] [CrossRef] [Green Version]
- Gürler, E.B.; Çilingir-Kaya, Ö.T.; Peker Eyüboglu, I.; Ercan, F.; Akkiprik, M.; Reiter, R.J.; Yegen, B. Melatonin supports alendronate in preserving bone matrix and prevents gastric inflammation in ovariectomized rats. Cell Biochem. Funct. 2019, 37, 102–112. [Google Scholar] [CrossRef]
- Zhu, F.; Liu, Z.; Ren, Y. Mechanism of melatonin combined with calcium carbonate on improving osteoporosis in aged rats. Exp. Ther. Med. 2018, 16, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Ladizesky, M.G.; Cutrera, R.A.; Boggio, V.; Somoza, J.; Centrella, J.M.; Mautalen, C.; Cardinali, D.P. Effect of melatonin on bone metabolism in ovariectomized rats. Life Sci. 2001, 70, 557–565. [Google Scholar] [CrossRef]
- Ladizesky, M.G.; Boggio, V.; Albornoz, L.E.; Castrillon, P.O.; Mautalen, C.; Cardinali, D.P. Melatonin increases oestradiol-induced bone formation in ovariectomized rats. J. Pineal Res. 2003, 34, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Ladizesky, M.G.; Boggio, V.; Cutrera, R.A.; Mondelo, N.; Mastaglia, S.; Somoza, J.; Cardinali, D.P. Melatonin effect on bone metabolism in rats treated with methylprednisolone. J. Pineal Res. 2006, 40, 297–304. [Google Scholar] [CrossRef]
- Satomura, K.; Tobiume, S.; Tokuyama, R.; Yamasaki, Y.; Kudoh, K.; Maeda, E.; Nagayama, M. Melatonin at pharmacological doses enhances human osteoblastic differentiation in vitro and promotes mouse cortical bone formation in vivo. J. Pineal Res. 2007, 42, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Munoz, F.; Lopez-Pena, M.; Mino, N.; Gomez-Moreno, G.; Guardia, J.; Cutando, A. Topical application of melatonin and growth hormone accelerates bone healing around dental implants in dogs. Clin. Implant Dent. Relat. Res. 2012, 14, 226–235. [Google Scholar] [CrossRef]
- Sharan, K.; Lewis, K.; Furukawa, T.; Yadav, V.K. Regulation of bone mass through pineal-derived melatonin-MT2 receptor pathway. J. Pineal Res. 2017, 63, e12423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, Z.M.; Li, H.B.; Sun, S.X.; Jiang, Y.C.; Wang, B.; Dong, Y.F. Melatonin promotes osteoblast differentiation of bone marrow mesenchymal stem cells in aged rats. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 4446–4456. [Google Scholar]
- Liu, H.; Liu, Z.; Man, C.W.; Guo, J.; Han, X.; Hu, Z.; Ng, T.B.; Zhao, Z.; Li, J.; Wang, W.; et al. The effect of exogenous melatonin on reducing scoliotic curvature and improving bone quality in melatonin-deficient C57BL/6J mice. Sci. Rep. 2019, 9, 6202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Liang, T.; Zhu, Y.; Qiu, J.; Qiu, X.; Lian, C.; Gao, B.; Peng, Y.; Liang, A.; Zhou, H.; et al. Melatonin prevents bone destruction in mice with retinoic acid-induced osteoporosis. Mol. Med. 2019, 25, 43. [Google Scholar] [CrossRef]
- Zhou, W.; Liu, Y.; Shen, J.; Yu, B.; Bai, J.; Lin, J.; Guo, X.; Sun, H.; Chen, Z.; Yang, H.; et al. Melatonin Increases Bone Mass around the Prostheses of OVX Rats by Ameliorating Mitochondrial Oxidative Stress via the SIRT3/SOD2 Signaling Pathway. Oxid. Med. Cell. Longev. 2019, 2019, 4019619. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, T.; Liu, X.; Li, Z.; Zhou, D.; Xu, W. Melatonin suppresses epithelial-to-mesenchymal transition in the MG-63 cell line. Mol. Med. Rep. 2020, 21, 1356–1364. [Google Scholar] [CrossRef]
- Da, W.; Tao, L.; Wen, K.; Tao, Z.; Wang, S.; Zhu, Y. Protective Role of Melatonin Against Postmenopausal Bone Loss via Enhancement of Citrate Secretion from Osteoblasts. Front. Pharmacol 2020, 11, 667. [Google Scholar] [CrossRef]
- Xiao, L.; Lin, J.; Chen, R.; Huang, Y.; Liu, Y.; Bai, J.; Ge, G.; Shi, X.; Chen, Y.; Shi, J.; et al. Sustained Release of Melatonin from GelMA Liposomes Reduced Osteoblast Apoptosis and Improved Implant Osseointegration in Osteoporosis. Oxid. Med. Cell Longev. 2020, 2020, 6797154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, X.; Wang, G.; Chen, L.; Yang, H.; He, F.; Lin, J. Melatonin Rescues the Ti Particle-Impaired Osteogenic Potential of Bone Marrow Mesenchymal Stem Cells via the SIRT1/SOD2 Signaling Pathway. Calcif. Tissue Int. 2020, 107, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, C.; Si, J.; Wang, B.; Zhang, D.; Ding, D.; Zhang, J.; Wang, H. Melatonin up-regulates bone marrow mesenchymal stem cells osteogenic action but suppresses their mediated osteoclastogenesis via MT2 -inactivated NF-kappaB pathway. Br. J. Pharmacol. 2020, 177, 2106–2122. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Feng, C.; Gao, M.; Jin, M.; Liu, T.; Yuan, Y.; Yan, G.; Gong, R.; Sun, Y.; He, M.; et al. MicroRNA-92b-5p modulates melatonin-mediated osteogenic differentiation of bone marrow mesenchymal stem cells by targeting ICAM-1. J. Cell. Mol. Med. 2019, 23, 6140–6153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, X.; Wang, X.; Qiu, J.; Zhu, Y.; Liang, T.; Gao, B.; Wu, Z.; Lian, C.; Peng, Y.; Liang, A.; et al. Melatonin Rescued Reactive Oxygen Species-Impaired Osteogenesis of Human Bone Marrow Mesenchymal Stem Cells in the Presence of Tumor Necrosis Factor-Alpha. Stem Cells Int. 2019, 2019, 6403967. [Google Scholar] [CrossRef]
- Ma, H.; Wang, X.; Zhang, W.; Li, H.; Zhao, W.; Sun, J.; Yang, M. Melatonin Suppresses Ferroptosis Induced by High Glucose via Activation of the Nrf2/HO-1 Signaling Pathway in Type 2 Diabetic Osteoporosis. Oxid. Med. Cell. Longev. 2020, 2020, 9067610. [Google Scholar] [CrossRef]
- Zhou, R.; Ma, Y.; Tao, Z.; Qiu, S.; Gong, Z.; Tao, L.; Zhu, Y. Melatonin Inhibits Glucose-Induced Apoptosis in Osteoblastic Cell Line Through PERK-eIF2alpha-ATF4 Pathway. Front. Pharmacol. 2020, 11, 602307. [Google Scholar] [CrossRef]
- Zhao, R.; Tao, L.; Qiu, S.; Shen, L.; Tian, Y.; Gong, Z.; Tao, Z.B.; Zhu, Y. Melatonin rescues glucocorticoid-induced inhibition of osteoblast differentiation in MC3T3-E1 cells via the PI3K/AKT and BMP/Smad signalling pathways. Life Sci. 2020, 257, 118044. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, T.; Deng, Z.; Gao, W.; Liang, T.; Qiu, X.; Gao, B.; Wu, Z.; Qiu, J.; Zhu, Y.; et al. Melatonin promotes bone marrow mesenchymal stem cell osteogenic differentiation and prevents osteoporosis development through modulating circ_0003865 that sponges miR-3653-3p. Stem Cell Res. Ther. 2021, 12, 150. [Google Scholar] [CrossRef]
- Chen, W.; Chen, X.; Chen, A.C.; Shi, Q.; Pan, G.; Pei, M.; Yang, H.; Liu, T.; He, F. Melatonin restores the osteoporosis-impaired osteogenic potential of bone marrow mesenchymal stem cells by preserving SIRT1-mediated intracellular antioxidant properties. Free Radic. Biol. Med. 2020, 146, 92–106. [Google Scholar] [CrossRef] [PubMed]
- Kotlarczyk, M.P.; Lassila, H.C.; O’Neil, C.K.; D’Amico, F.; Enderby, L.T.; Witt-Enderby, P.A.; Balk, J.L. Melatonin osteoporosis prevention study (MOPS): A randomized, double-blind, placebo-controlled study examining the effects of melatonin on bone health and quality of life in perimenopausal women. J. Pineal Res. 2012, 52, 414–426. [Google Scholar] [CrossRef] [PubMed]
- Amstrup, A.K.; Sikjaer, T.; Heickendorff, L.; Mosekilde, L.; Rejnmark, L. Melatonin improves bone mineral density at the femoral neck in postmenopausal women with osteopenia: A randomized controlled trial. J. Pineal Res. 2015, 59, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Amstrup, A.K.; Sikjaer, T.; Mosekilde, L.; Rejnmark, L. The effect of melatonin treatment on postural stability, muscle strength, and quality of life and sleep in postmenopausal women: A randomized controlled trial. Nutr. J. 2015, 14, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria, S.; Swanson, M.H.; Enderby, L.T.; D’Amico, F.; Enderby, B.; Samsonraj, R.M.; Dudakovic, A.; van Wijnen, A.J.; Witt-Enderby, P.A. Melatonin-micronutrients Osteopenia Treatment Study (MOTS): A translational study assessing melatonin, strontium (citrate), vitamin D3 and vitamin K2 (MK7) on bone density, bone marker turnover and health related quality of life in postmenopausal osteopenic women following a one-year double-blind RCT and on osteoblast-osteoclast co-cultures. Aging 2017, 9, 256–285. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, Z.; Wang, J.; Li, Z.; Cui, H.; Dai, G.; Chen, S.; Zhang, M.; Zheng, Z.; Zhan, Z.; et al. Wnt4 signaling mediates protective effects of melatonin on new bone formation in an inflammatory environment. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 10126–10139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igarashi-Migitaka, J.; Seki, A.; Ikegame, M.; Honda, M.; Sekiguchi, T.; Mishima, H.; Shimizu, N.; Matsubara, H.; Srivastav, A.K.; Hirayama, J.; et al. Oral administration of melatonin contained in drinking water increased bone strength in naturally aged mice. Acta Histochem. 2020, 122, 151596. [Google Scholar] [CrossRef]
- Tao, Z.S.; Lu, H.L.; Ma, N.F.; Zhang, R.T.; Li, Y.; Yang, M.; Xu, H.G. Rapamycin could increase the effects of melatonin against age-dependent bone loss. Z. Gerontol. Geriatr. 2020, 53, 671–678. [Google Scholar] [CrossRef]
- Ghareghani, M.; Scavo, L.; Arnoult, D.; Zibara, K.; Farhadi, N. Melatonin therapy reduces the risk of osteoporosis and normalizes bone formation in multiple sclerosis. Fundam. Clin. Pharmacol. 2018, 32, 181–187. [Google Scholar] [CrossRef]
- Ren, H.; Sun, R.; Wang, J. Relationship of melatonin level, oxidative stress and inflammatory status with osteoporosis in maintenance hemodialysis of chronic renal failure. Exp. Ther. Med. 2018, 15, 5183–5188. [Google Scholar] [CrossRef]
- Giudice, A.; Crispo, A.; Grimaldi, M.; Polo, A.; Bimonte, S.; Capunzo, M.; Amore, A.; D’Arena, G.; Cerino, P.; Budillon, A.; et al. The Effect of Light Exposure at Night (LAN) on Carcinogenesis via Decreased Nocturnal Melatonin Synthesis. Molecules 2018, 23, 1308. [Google Scholar] [CrossRef] [Green Version]
- Anbalagan, M.; Dauchy, R.; Xiang, S.; Robling, A.; Blask, D.; Rowan, B.; Hill, S. SAT-337 Disruption Of The Circadian Melatonin Signal By Dim Light At Night Promotes Bone-lytic Breast Cancer Metastases. J. Endocr. Soc. 2019, 3. [Google Scholar] [CrossRef]
- Anbalagan, M. Abstract LB-212: Circadian melatonin signal disruption by exposure to artificial light at night promotes bone lytic breast cancer metastases. Cancer Res. 2020, 80, LB-212. [Google Scholar] [CrossRef]
- Xiang, S.; Dauchy, R.T.; Hauch, A.; Mao, L.; Yuan, L.; Wren, M.A.; Belancio, V.P.; Mondal, D.; Frasch, T.; Blask, D.E.; et al. Doxorubicin resistance in breast cancer is driven by light at night-induced disruption of the circadian melatonin signal. J. Pineal Res. 2015, 59, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.; Dauchy, R.T.; Hoffman, A.E.; Pointer, D.; Frasch, T.; Blask, D.E.; Hill, S.M. Epigenetic inhibition of the tumor suppressor ARHI by light at night-induced circadian melatonin disruption mediates STAT3-driven paclitaxel resistance in breast cancer. J. Pineal Res. 2019, 67, e12586. [Google Scholar] [CrossRef]
- Dauchy, R.T.; Xiang, S.; Mao, L.; Brimer, S.; Wren, M.A.; Yuan, L.; Anbalagan, M.; Hauch, A.; Frasch, T.; Rowan, B.G.; et al. Circadian and melatonin disruption by exposure to light at night drives intrinsic resistance to tamoxifen therapy in breast cancer. Cancer Res. 2014, 74, 4099–4110. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.H.; Lin, R.C.; Yang, J.S.; Yang, W.E.; Reiter, R.J.; Yang, S.F. Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma. Cells 2019, 8, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Zhao, J.; Bai, J.; Shen, H.; Zhang, B.; Deng, L.; Sun, C.; Liu, Y.; Zhang, J.; Zheng, J. Risk and clinicopathological features of osteosarcoma metastasis to the lung: A population-based study. J. Bone Oncol. 2019, 16, 100230. [Google Scholar] [CrossRef]
- Ahmed, G.; Zamzam, M.; Kamel, A.; Ahmed, S.; Salama, A.; Zaki, I.; Kamal, N.; Elshafiey, M. Effect of timing of pulmonary metastasis occurrence on the outcome of metastasectomy in osteosarcoma patients. J. Pediatr. Surg. 2019, 54, 775–779. [Google Scholar] [CrossRef]
- Panzer, A. Melatonin in osteosarcoma: An effective drug? Med. Hypotheses 1997, 48, 523–525. [Google Scholar] [CrossRef]
- Wang, Y.P.; Yang, Z.P. Effects of melatonin combined with Cis-platinum or methotrexate on the proliferation of osteosarcoma cell line SaOS-2. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2015, 37, 215–220. [Google Scholar] [CrossRef]
- Lu, K.H.; Su, S.C.; Lin, C.W.; Hsieh, Y.H.; Lin, Y.C.; Chien, M.H.; Reiter, R.J.; Yang, S.F. Melatonin attenuates osteosarcoma cell invasion by suppression of C-C motif chemokine ligand 24 through inhibition of the c-Jun N-terminal kinase pathway. J. Pineal Res. 2018, 65, e12507. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Santos, G.; Martin, V.; Rodriguez-Blanco, J.; Herrera, F.; Casado-Zapico, S.; Sanchez-Sanchez, A.M.; Antolin, I.; Rodriguez, C. Fas/Fas ligand regulation mediates cell death in human Ewing’s sarcoma cells treated with melatonin. Br. J. Cancer 2012, 106, 1288–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, M.; Cui, P.; Yu, M.; Han, J.; Li, H.; Xiu, R. Melatonin modulates the expression of VEGF and HIF-1 alpha induced by CoCl2 in cultured cancer cells. J. Pineal Res. 2008, 44, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Hwang, M.S.; Suh, S.I.; Baek, W.K. Melatonin down-regulates HIF-1 alpha expression through inhibition of protein translation in prostate cancer cells. J. Pineal Res. 2009, 46, 415–421. [Google Scholar] [CrossRef]
- Shrestha, S.; Zhu, J.; Wang, Q.; Du, X.; Liu, F.; Jiang, J.; Song, J.; Xing, J.; Sun, D.; Hou, Q.; et al. Melatonin potentiates the antitumor effect of curcumin by inhibiting IKKbeta/NF-kappaB/COX-2 signaling pathway. Int. J. Oncol. 2017, 51, 1249–1260. [Google Scholar] [CrossRef]
- Qu, H.; Xue, Y.; Lian, W.; Wang, C.; He, J.; Fu, Q.; Zhong, L.; Lin, N.; Lai, L.; Ye, Z.; et al. Melatonin inhibits osteosarcoma stem cells by suppressing SOX9-mediated signaling. Life Sci. 2018, 207, 253–264. [Google Scholar] [CrossRef]
- Wang, X.; Wang, B.; Xie, J.; Hou, D.; Zhang, H.; Huang, H. Melatonin inhibits epithelial-to-mesenchymal transition in gastric cancer cells via attenuation of IL-1β/NF-κB/MMP2/MMP9 signaling. Int. J. Mol. Med. 2018, 42, 2221–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, M.C.; Pan, Y.H.; Wu, H.L.; Lu, Y.J.; Liao, W.C.; Yeh, C.Y.; Lee, J.J.; Jeng, J.H. Stimulation of MMP-9 of oral epithelial cells by areca nut extract is related to TGF-beta/Smad2-dependent and -independent pathways and prevented by betel leaf extract, hydroxychavicol and melatonin. Aging 2019, 11, 11624–11639. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Tai, H.C.; Tang, C.H.; Lin, L.W.; Lin, T.H.; Chang, A.C.; Chen, P.C.; Chen, Y.H.; Wang, P.C.; Lai, Y.W.; et al. Melatonin impedes prostate cancer metastasis by suppressing MMP-13 expression. J. Cell Physiol. 2021, 236, 3979–3990. [Google Scholar] [CrossRef]
- Gulbahce-Mutlu, E.; Baltaci, S.B.; Menevse, E.; Mogulkoc, R.; Baltaci, A.K. The Effect of Zinc and Melatonin Administration on Lipid Peroxidation, IL-6 Levels, and Element Metabolism in DMBA-Induced Breast Cancer in Rats. Biol. Trace Elem. Res. 2021, 199, 1044–1051. [Google Scholar] [CrossRef]
- Hasan, M.; Browne, E.; Guarinoni, L.; Darveau, T.; Hilton, K.; Witt-Enderby, P.A. Novel Melatonin, Estrogen, and Progesterone Hormone Therapy Demonstrates Anti-Cancer Actions in MCF-7 and MDA-MB-231 Breast Cancer Cells. Breast Cancer 2020, 14, 1178223420924634. [Google Scholar] [CrossRef] [PubMed]
- Estaras, M.; Gonzalez-Portillo, M.R.; Martinez, R.; Garcia, A.; Estevez, M.; Fernandez-Bermejo, M.; Mateos, J.M.; Vara, D.; Blanco-Fernandez, G.; Lopez-Guerra, D.; et al. Melatonin Modulates the Antioxidant Defenses and the Expression of Proinflammatory Mediators in Pancreatic Stellate Cells Subjected to Hypoxia. Antioxidants 2021, 10, 577. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhu, Y.; Zhu, H.; Cai, R.; Wang, K.F.; Song, J.; Wang, R.X.; Zhou, R.X. Role of transforming growth factor beta1 in the inhibition of gastric cancer cell proliferation by melatonin in vitro and in vivo. Oncol. Rep. 2019, 42, 753–762. [Google Scholar] [CrossRef] [PubMed]


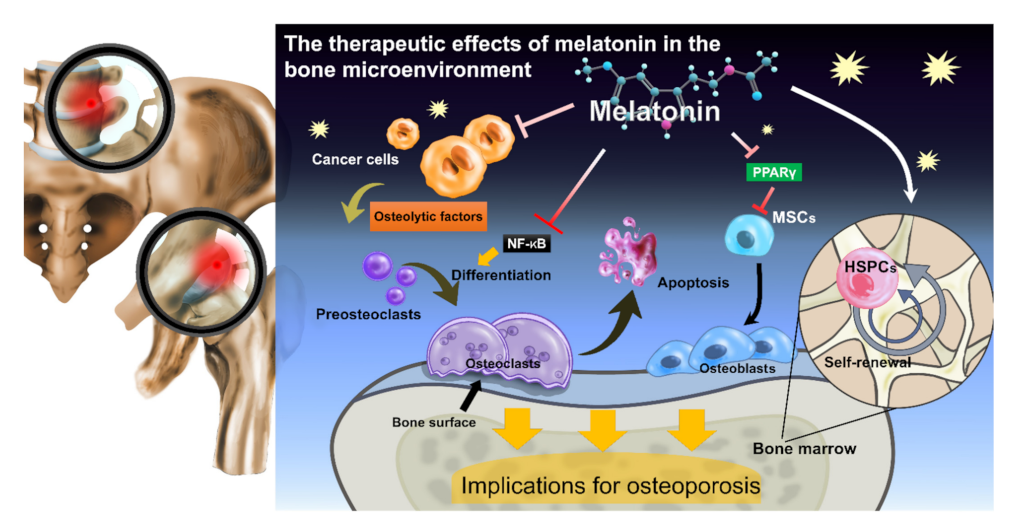{kind=link}
| Preclinical | |||||
| Cellular or Animal Model | Dosage | Administration | Outcomes | Ref. | |
| BMMSCs | 10 µmol/L | Culture | Melatonin promoted osteogenesis in BMMSCs by upregulating miR-92b-5p expression, which enhanced the differentiation of BMMSCs into mature osteoblasts by targeting ICAM-1. | [54] | |
| MC3T3-E1 cells, human bone MSCs | 1 mM | Culture | Melatonin promoted osteogenic differentiation and mineralization in inflammatory conditions; this process required Wnt4, via activation of β-catenin and p38-JNK MAPK signaling. | [65] | |
| BMMSCs | 100 μmol/L | Culture | Melatonin inhibited ROS generation during osteogenesis of BMMSCs in the presence of TNF-α and promoted the osteogenic differentiation of BMMSCs. | [55] | |
| MC3T3-E1 cells Diabetic rats | Cells: 1, 10, or 100 μM Rats: 10 or 50 mg/kg | Culture Animals: Intraperitoneal injection | Melatonin inhibited ferroptosis and improved the osteogenic capacity of MC3T3-E1 cells by activating Nrf2/HO-1 signaling in vitro and in vivo. | [56] | |
| MC3T3-E1 cells | 100 nM | Culture | Melatonin protects MC3T3-E1 cells against high glucose-induced changes (reduced viability, apoptosis and calcium influx) by inhibiting the PERK-eIF2α-ATF4-CHOP signaling pathway. | [57] | |
| MC3T3-E1 cells | 1 μM | Culture | Melatonin treatment rescued MC3T3-E1 cells from dexamethasone-induced inhibition of osteoblast differentiation via the PI3K/AKT and BMP/Smad signaling pathways. | [58] | |
| OVX rats | 25 μg/mL | Oral (melatonin in drinking water) | Melatonin and alendronate provided similar supportive effects on preservation of bone mass in OVX rats, with no additive effect on bone remodeling when these treatments were combined. However, melatonin prevented alendronate-induced gastric side effects. | [37] | |
| Raw264.7 cells | 0.1 or 1 µmol | Culture | Melatonin inhibits RANKL-induced osteoclastogenesis by increasing the expression of Rev-erbα and reducing miR-882 expression in Raw264.7 cells. | [27] | |
| BMMSCs | 100 μmol/L | Culture | Melatonin promoted BMMSC osteogenic differentiation and inhibited osteoporosis pathogenesis by suppressing the expression of circ_0003865 and increasing the expression of miR-3653-3p. | [59] | |
| Clinical | |||||
| Study Population | Route of Administration | Dosing Schedule | Outcomes | ClinicalTrials.gov. Identifier | Ref. |
| Perimenopausal women (n = 18) | Oral | Nightly placebo or melatonin (3 mg) for 6 months. | At 6 months, serum markers of bone resorption (NTX) and formation (OCN) were not significantly changed from baseline in either group, although the NTX:OCN ratio trended downward over time with melatonin (placebo showed no such trend). Melatonin also reduced osteoclast to osteoblast ratios. | NCT01152580 | [61] |
| Postmenopausal women with osteopenia* (n = 81) | Oral | Nightly placebo or melatonin (1 mg or 3 mg) for 12 months. All study participants also received a daily supplement of 800 mg calcium and 20 μg vitamin D3. | After 1 year of treatment, femoral neck BMD was increased by 0.5% with 1 mg/day melatonin and by 2.3% with 3 mg/day melatonin, compared with placebo. At 12 months, trabecular thickness was increased from baseline in the (combined) melatonin group by 2.2% compared with placebo, while volumetric BMD at the lumbar spine was increased by 3.6% in the 3 mg/day melatonin group compared with placebo. Biochemical markers of bone turnover were not affected by melatonin, although 24-h urinary calcium was decreased by 3.7% in the (combined) melatonin group compared with placebo. | NCT01690000 | [62] |
| Postmenopausal women with osteopenia * (n = 23) | Oral | Nightly placebo or MSDK supplementation: melatonin (5 mg), strontium (citrate), vitamin D3 and vitamin K2. | Over 1 year, compared with placebo, MSDK treatment increased lumbar spine BMD by 4.3% and left femoral neck BMD by 2.2%, and showed a trend towards an increase in hip BMD from baseline. The 10-year vertebral fracture risk probability fell by 6.48% with MSDK treatment, but increased by 10.8% with placebo. MSDK increased serum bone formation markers and reduced bone turnover. | NCT01870115 | [64] |
| Type of Cancer | Osteolytic Factors | Dosage | Model | Outcomes | Ref. |
|---|---|---|---|---|---|
| Pancreatic, cervical, lung | VEGF HIF-1α | Cells: 1 nM or 1 mM | Cell lines: PANC-1, HeLa and A549 | At the high concentration (1 mM), melatonin inhibited VEGF mRNA and protein levels, as well as HIF-1α protein, in all three human cancer cell lines. | [83] |
| Prostate | HIF-1α | Cells: 1 mM | Cell lines: DU145, PC-3, and LNCaP | Melatonin-induced inhibition of HIF-1α protein expression, HIF-1α transcriptional activity and the release of VEGF in all three cell lines correlated with dephosphorylation of p70S6K and its direct target RPS6. | [84] |
| Bladder | COX-2 | Cells: 1 mM | Cell lines: T24, UMUC3 and 5637 | When combined with curcumin, melatonin enhanced the inhibitory effects of curcumin on COX-2 activity and enhanced the antiproliferative, antimigratory and proapoptotic activities of curcumin in bladder cancer cells. | [85] |
| Animal: 10 mg/kg | Animal: BALB/c nude mice | ||||
| Osteosarcoma | SOX9 | Cells: 0.5 mM | Cell lines: HOS and U2-OS | Melatonin suppressed osteosarcoma cell migration and invasion and also significantly inhibited osteosarcoma metastasis in a mouse model of osteosarcoma. These effects were achieved by downregulating SOX9-mediated signaling. | [86] |
| Animal: 100 mg/kg | Animal: BALB/c nude mice | ||||
| Gastric adenocarcinoma | MMP-2 MMP-9 | Cells: 0.1, 0.5 or 1.5 mM | Cell lines: MGC80-3 and SGC-7901 | Melatonin suppressed IL-1β-induced EMT in human gastric adenocarcinoma cells by targeting IL-1β/NF-κB/MMP-2/MMP-9 signaling. | [87] |
| Osteosarcoma | CCL24 | Cells: 2 mM | Cell lines: HOS and U2OS | Melatonin inhibited the migratory potential and invasiveness of osteosarcoma HOS and U2OS cells. Melatonin also suppressed chemokine CCL24 levels in U2OS cells through the inhibition of the JNK pathway. | [81] |
| Oral | MMP-9 | Cells: 100 and 250 μg/mL | Cell line: SAS | Areca nut extract components (betel quid chewing) may contribute to tumor invasion and metastasis by stimulating MMP-9 mRNA expression and secretion of oral cancer cells, which was inhibited by melatonin. | [88] |
| Osteosarcoma | MMP-9 HIF-1αTGF-β | Cells: 50, 100, 200, 500 and 1000 nM | Cell line: MG-63 | Melatonin inhibits TGF-β1-induced EMT in osteosarcoma MG-63 cells by suppressing HIF-1α/Snail/MMP-9 signaling. | [49] |
| Prostate | MMP-13 | Cells: 1 mM | Cell lines: DU145 and PC-3 | Melatonin inhibited the migratory and invasive properties of prostate cancer cells, as well as MMP-13 expression, via the MT1 receptor and PLC, p38, and c-Jun signaling. Melatonin also inhibited prostate cancer metastasis and MMP-13 expression in an orthotopic prostate cancer model. | [89] |
| Animal: 20 or 60 mg/kg | Animal: SCID mice | ||||
| Breast | IL-6 | Animal: 5 mg/kg | Animal: Female rats with DMBA-induced breast cancer | Combined zinc and melatonin therapy helped to prevent tumor growth by significantly disrupting the metabolism of several elements (iron, magnesium, zinc and copper), and by suppressing IL-6 levels and reducing tissue damage that encourages tumor growth. | [90] |
| Lung, prostate | RANKL | Cells: 0.1, 0.3 or 0.7 mM | Cell lines: A549 and PC-3 | Melatonin inhibited RANKL production in lung and prostate cancer cells by downregulating the p38 MAPK pathway, which consequently prevented cancer-associated osteoclast differentiation. In animal models of lung and prostate bone metastasis, melatonin treatment markedly reduced tumor volumes and numbers of osteolytic lesions. | [11] |
| Animal: 20 or 60 mg/kg | Animal: BALB/c nude mice | ||||
| Breast | Integrin β 1Elf-5 | Cells: 5 mM | Cell lines: MCF-7 and MDA-MB-231 | MEMP HT (5 mg melatonin, 0.5 mg estradiol, and 50 mg progesterone [half the recommended dose] hormone therapy) showed anticancer activity in ER+ and triple negative breast cancer cells. These effects were largely attributed to the melatonin component and MEMP HT working through MEK1/2- and MEK-5-dependent intracellular signaling cascades in each cancer cell line, modulating intracellular signaling proteins that encourage the inhibition of cellular proliferative and migratory activities. | [91] |
| Pancreatic stellate cells | COX-2 IL-6 TNF-α | Cells: 1000, 100, 10 or 1 μM | Cells: Primary PSCs from Wistar rat pups (3–5 days after birth) | Pharmacological concentrations of melatonin increased ROS production and reduced levels of glutathione in PSCs under hypoxic conditions. Melatonin downregulated NF-kB phosphorylation and COX-2, IL-6, and TNF-α expression. | [92] |
| Gastric | TGF-β1 | Cells: 2 or 4 mM | Cell line: MFC | Melatonin inhibited gastric cancer cell proliferation in vitro by increasing TGF-β1 expression and also increased TGF-β1 levels in gastric cancer tumor tissues in vivo. | [93] |
| Animal: 25, 50, or 100 mg/kg | Animal: H-2Kk mice |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MacDonald, I.J.; Tsai, H.-C.; Chang, A.-C.; Huang, C.-C.; Yang, S.-F.; Tang, C.-H. Melatonin Inhibits Osteoclastogenesis and Osteolytic Bone Metastasis: Implications for Osteoporosis. Int. J. Mol. Sci. 2021, 22, 9435. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179435
MacDonald IJ, Tsai H-C, Chang A-C, Huang C-C, Yang S-F, Tang C-H. Melatonin Inhibits Osteoclastogenesis and Osteolytic Bone Metastasis: Implications for Osteoporosis. International Journal of Molecular Sciences. 2021; 22(17):9435. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179435
Chicago/Turabian StyleMacDonald, Iona J., Hsiao-Chi Tsai, An-Chen Chang, Chien-Chung Huang, Shun-Fa Yang, and Chih-Hsin Tang. 2021. "Melatonin Inhibits Osteoclastogenesis and Osteolytic Bone Metastasis: Implications for Osteoporosis" International Journal of Molecular Sciences 22, no. 17: 9435. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179435