
Cross−Talk between Transcriptome Analysis and Physiological Characterization Identifies the Genes in Response to the Low Phosphorus Stress in Malus mandshurica

 , ,
, ,
Abstract
:1. Introduction
2. Results
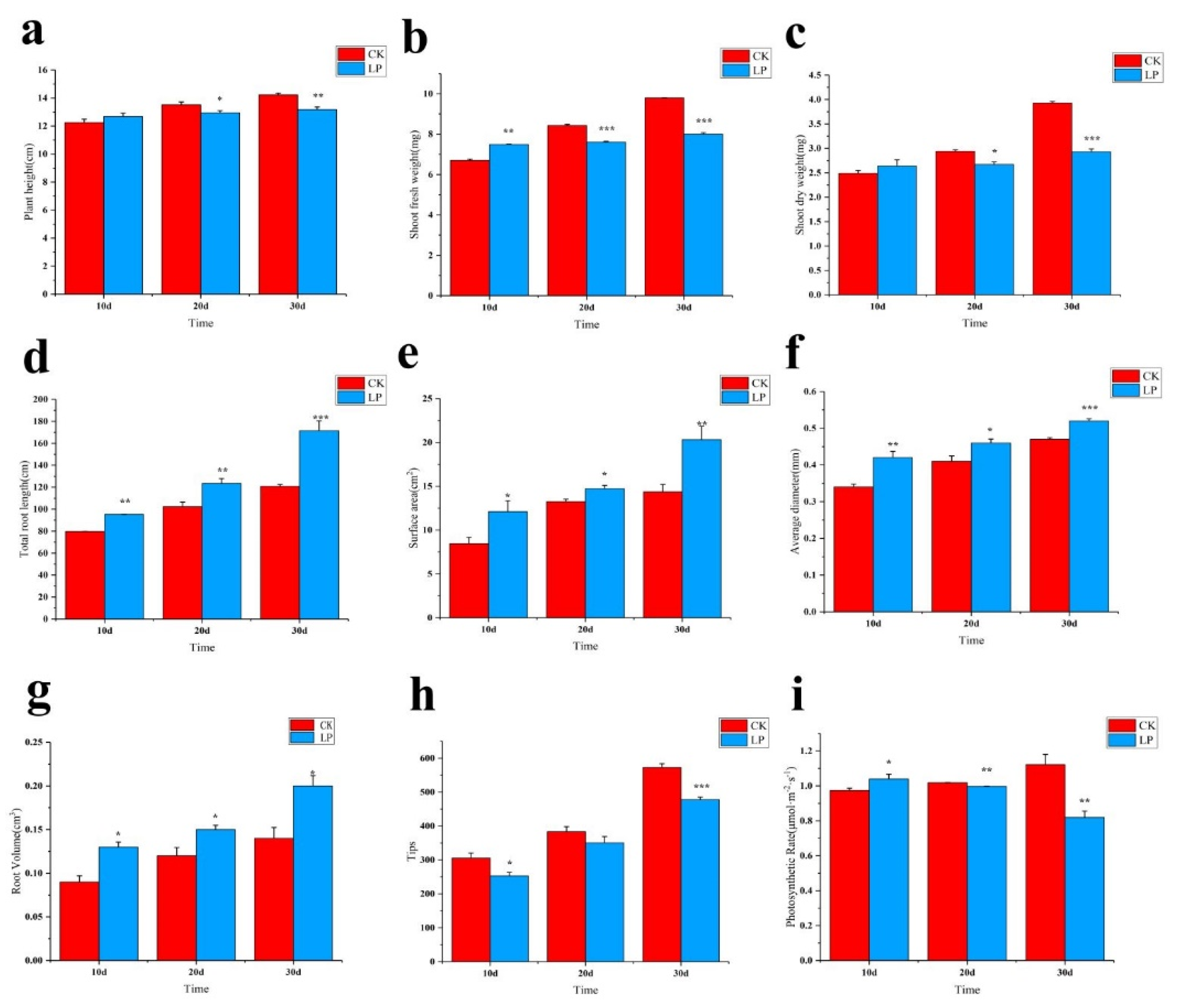
2.1. Morphological Traits to Low Phosphorus Treatment
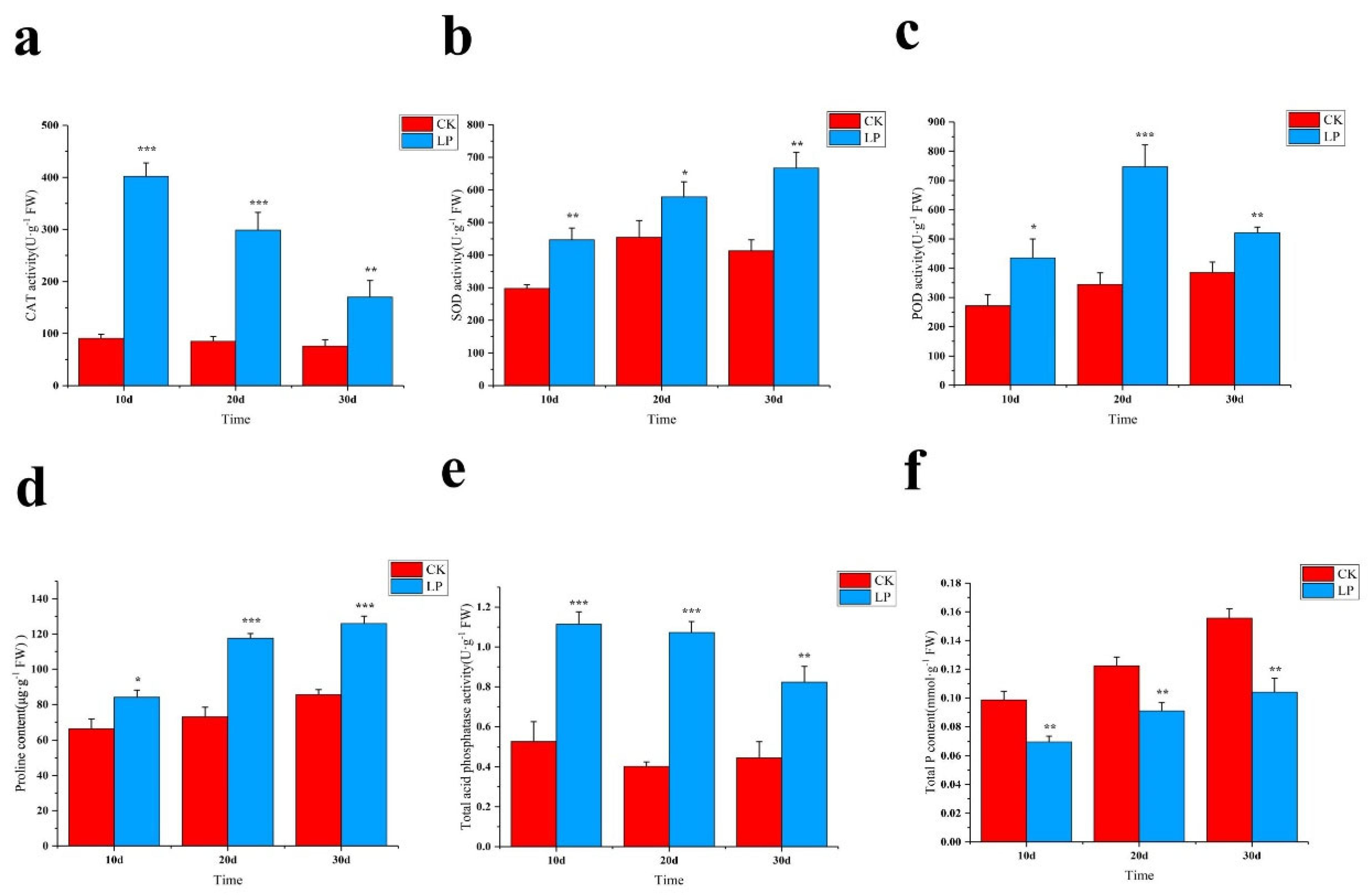
2.2. Physiological Responses to LowPhosphorus Treatment
2.3. Transcriptome Profiling and Validation of RNA Sequencing Data of M. mandshurica to LowPhosphorus Treatment
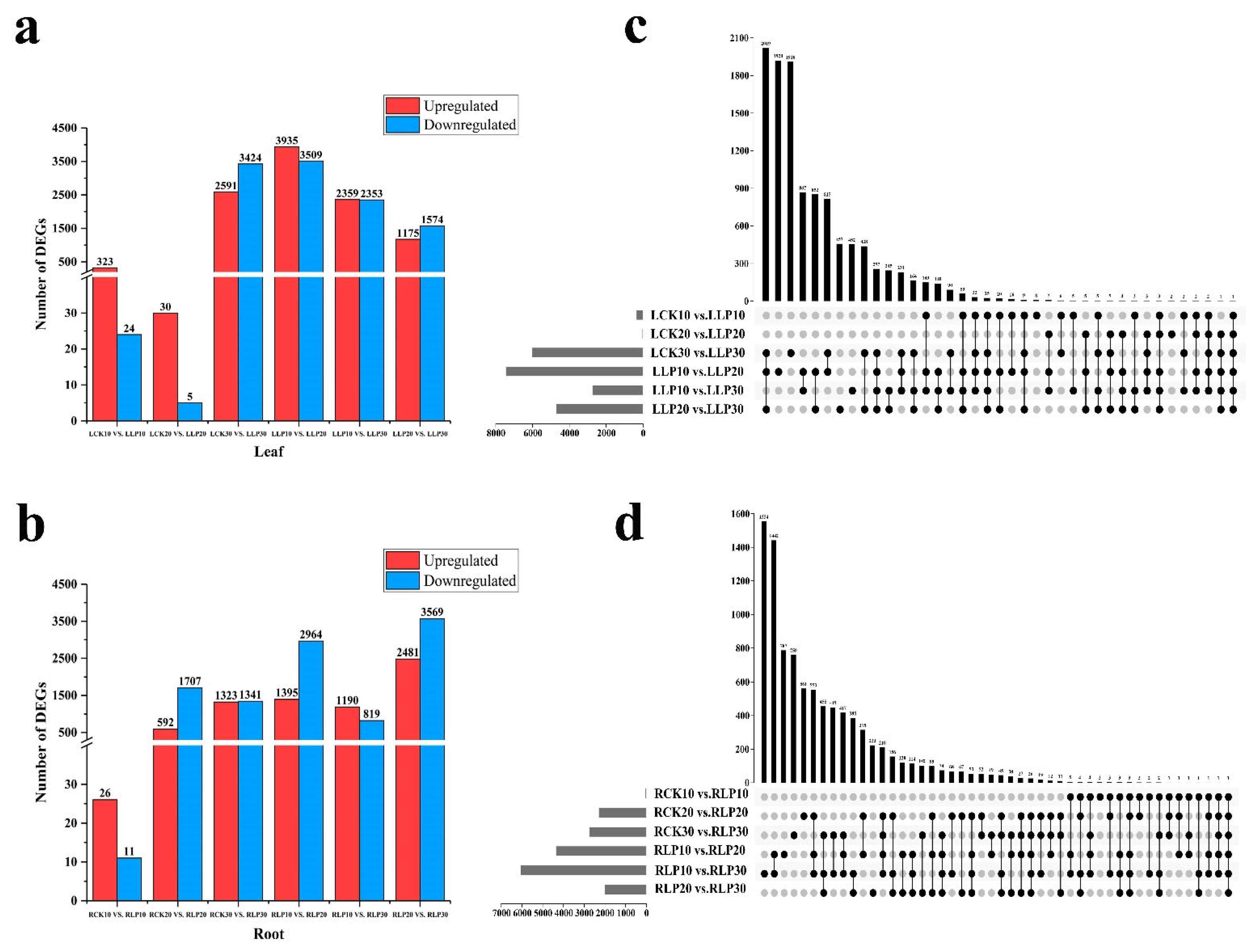
2.4. Identification of DEGs to Low Phosphorus Treatment
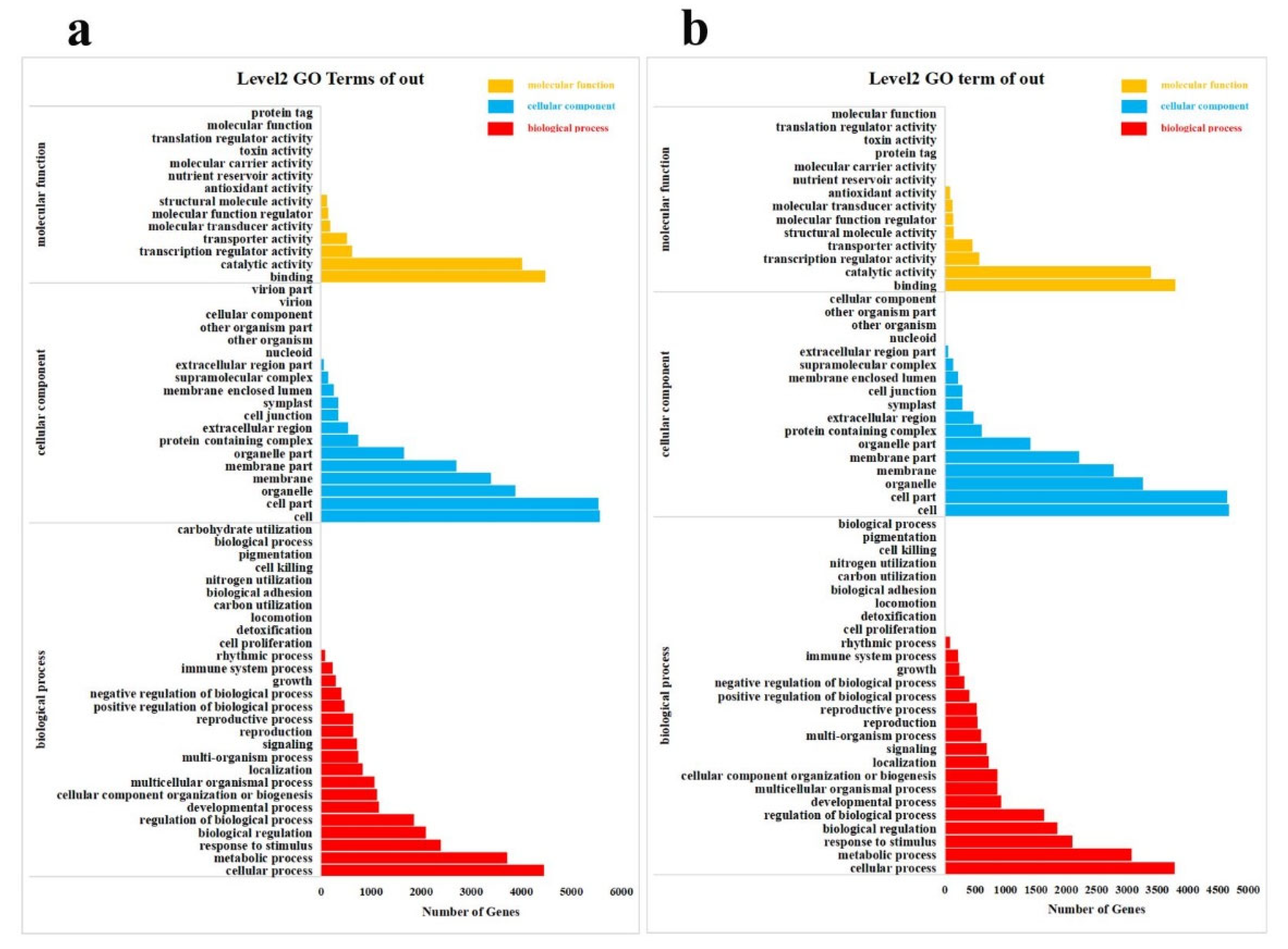
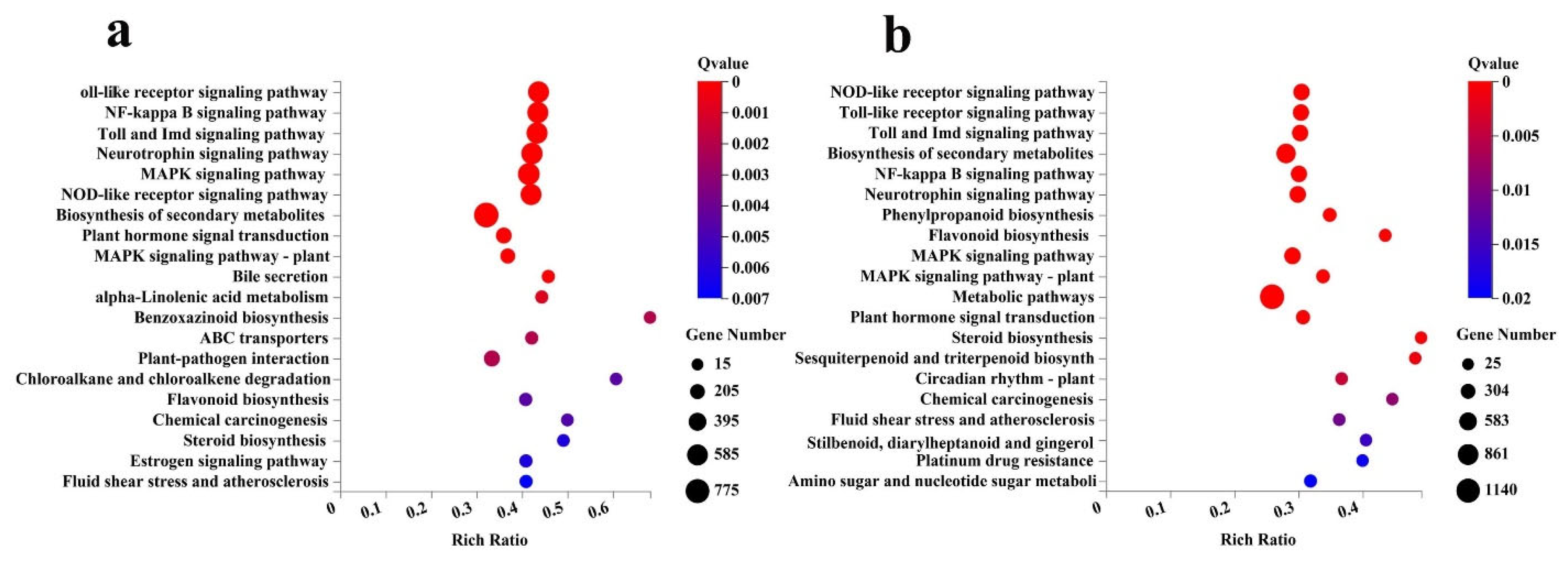
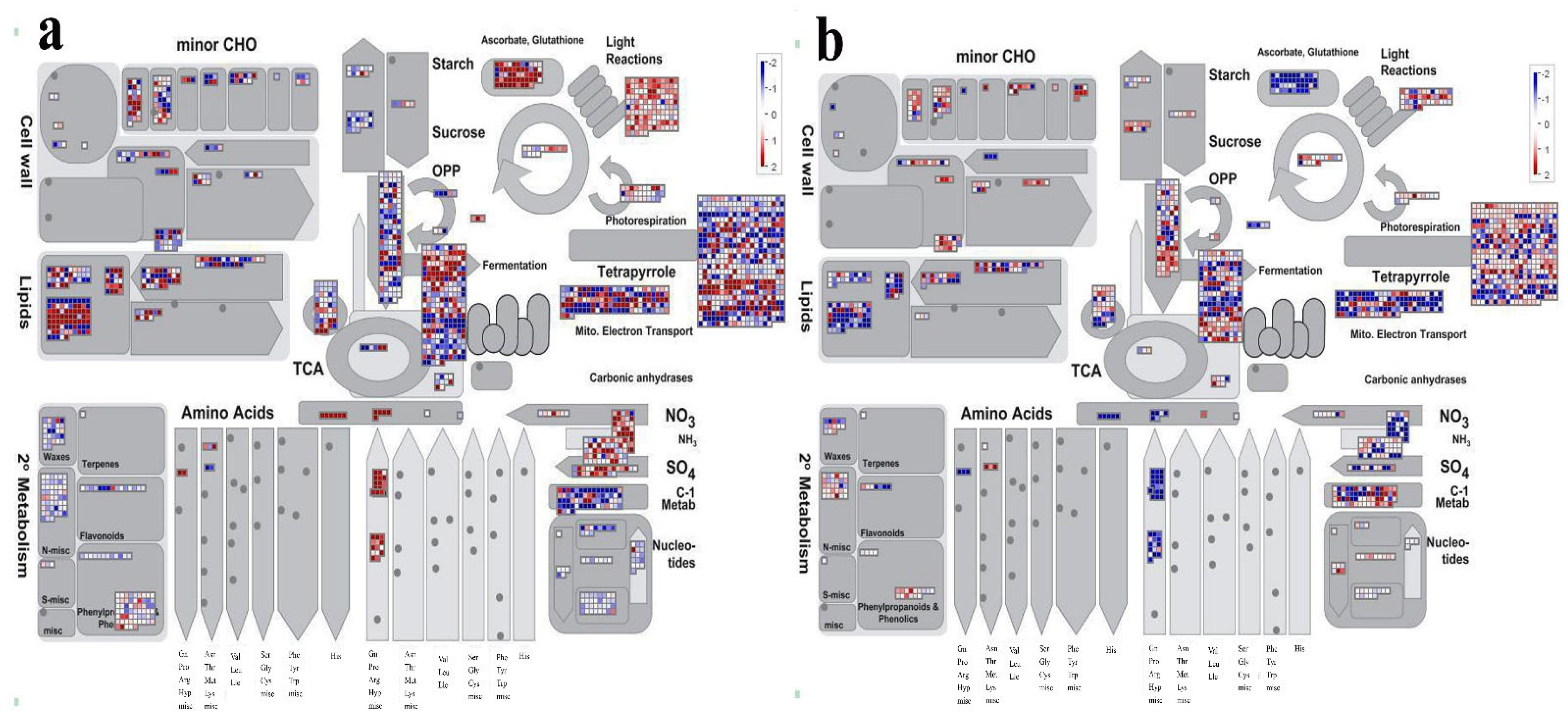
2.5. Classification, Enrichment and Metabolism Overview Analysis of DEGs
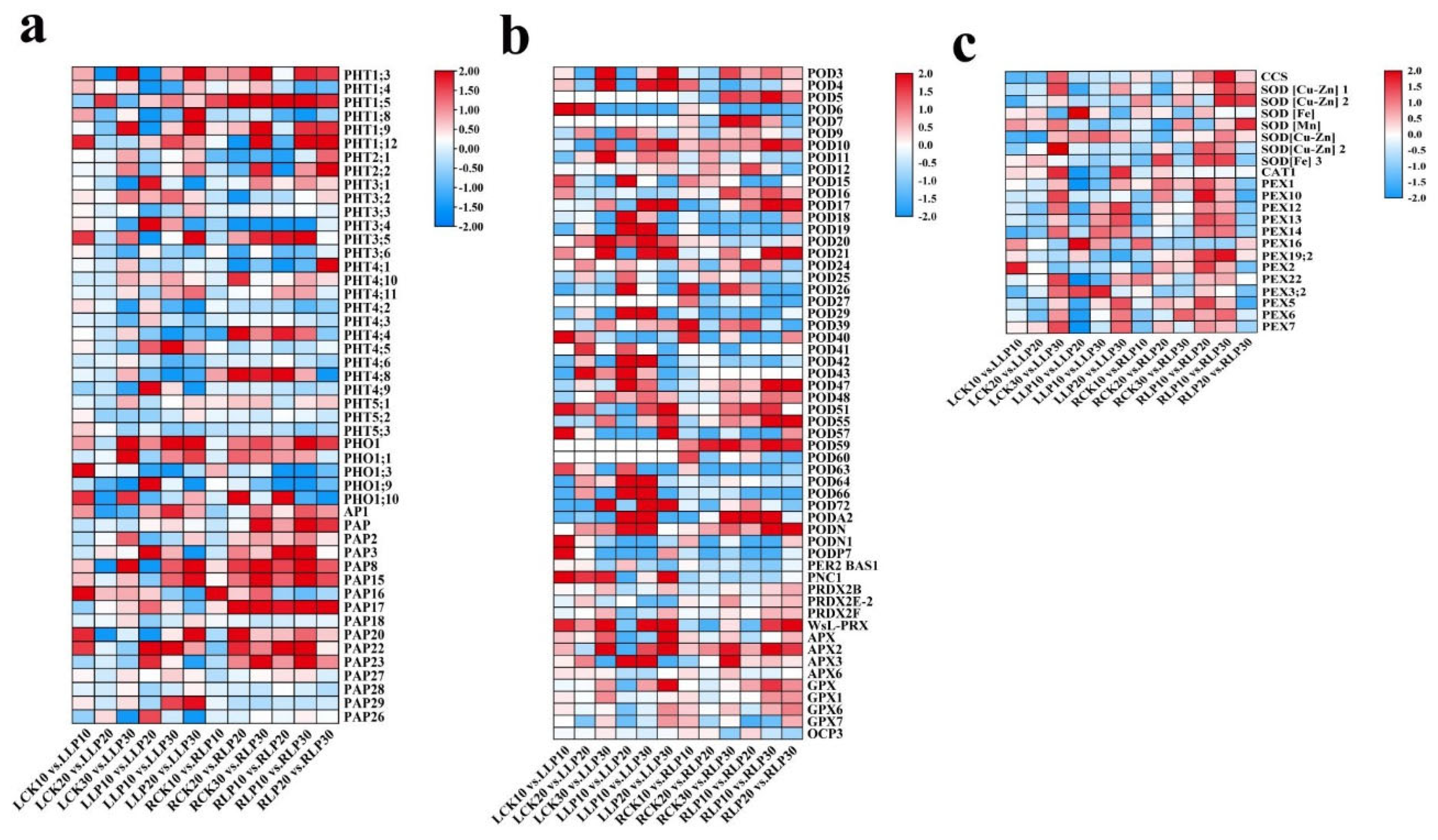
2.6. Expression Profiles of Physiologically Relevant DEGs to Low Phosphorus Treatment
2.7. Analysis of Pi Transporter Genes to Low-Pi Stress
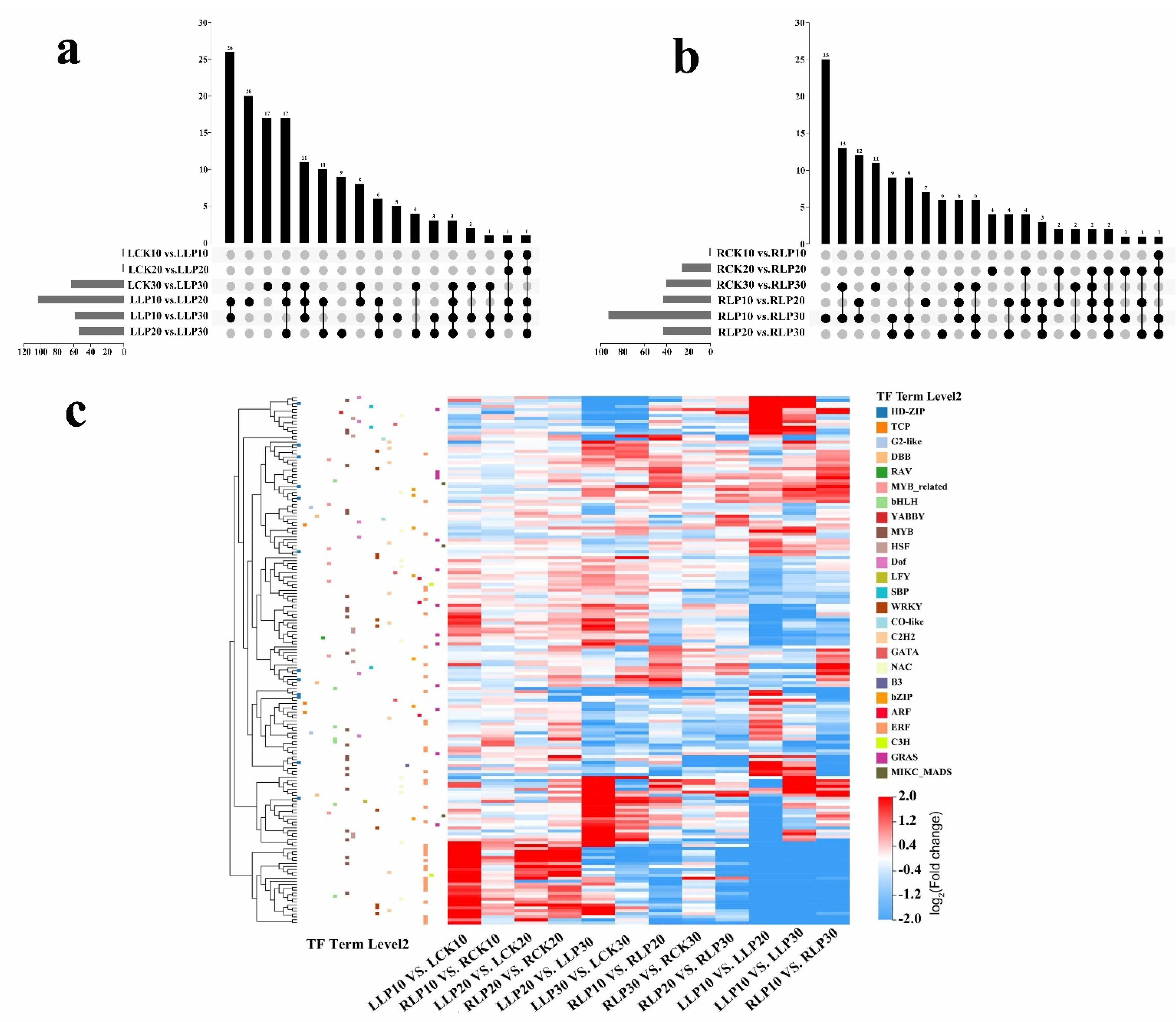
2.8. Identification of Differentially Expressed TFs to Low Phosphorus Treatment
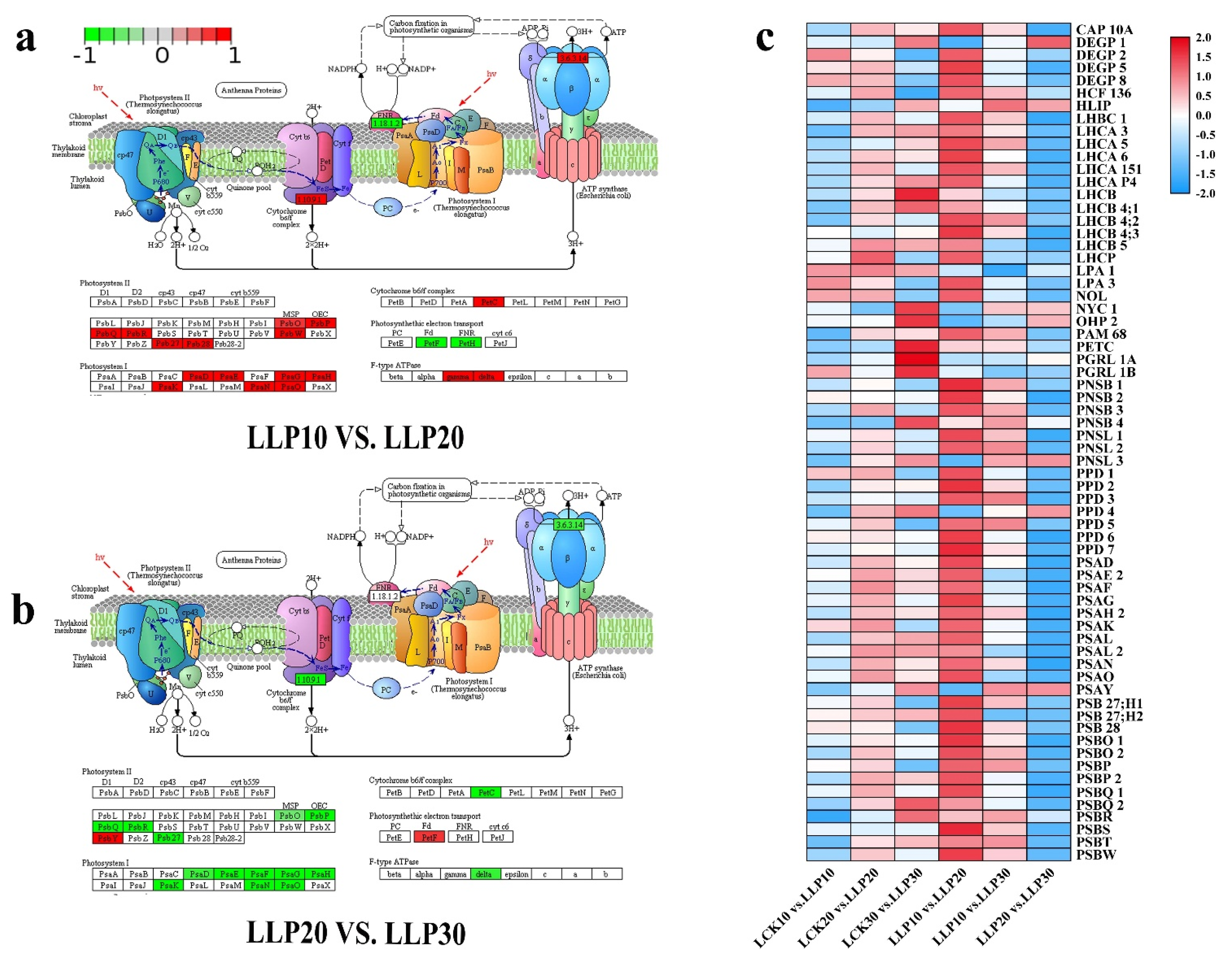
2.9. PSI and PSII System Genes in Response to Low Phosphorus Treatment
3. Discussion
3.1. Characteristics of Resistance to Low Phosphorus Stress in M. mandshurica
3.2. Antioxdative Genes Co−Ordinate the Low Phosphorus Tolerance in M. mandshurica
3.3. MYB TFs Involved in Low Phosphorus Tolerance in M. mandshurica
4. Materials and Methods
4.1. Plant Materials and Low Phosphorus Treatments
4.2. Determination of Morphological and Physiological Parameters
4.3. Quantification of Plant Total Phosphorus Content
4.4. RNA Extraction and Illumina Sequencing
4.5. RNA-Seq Data Processing and DEGs Analysis
4.6. Quantitative Real−Time PCR (qRT−PCR) Analysis
4.7. Statistical Analysis
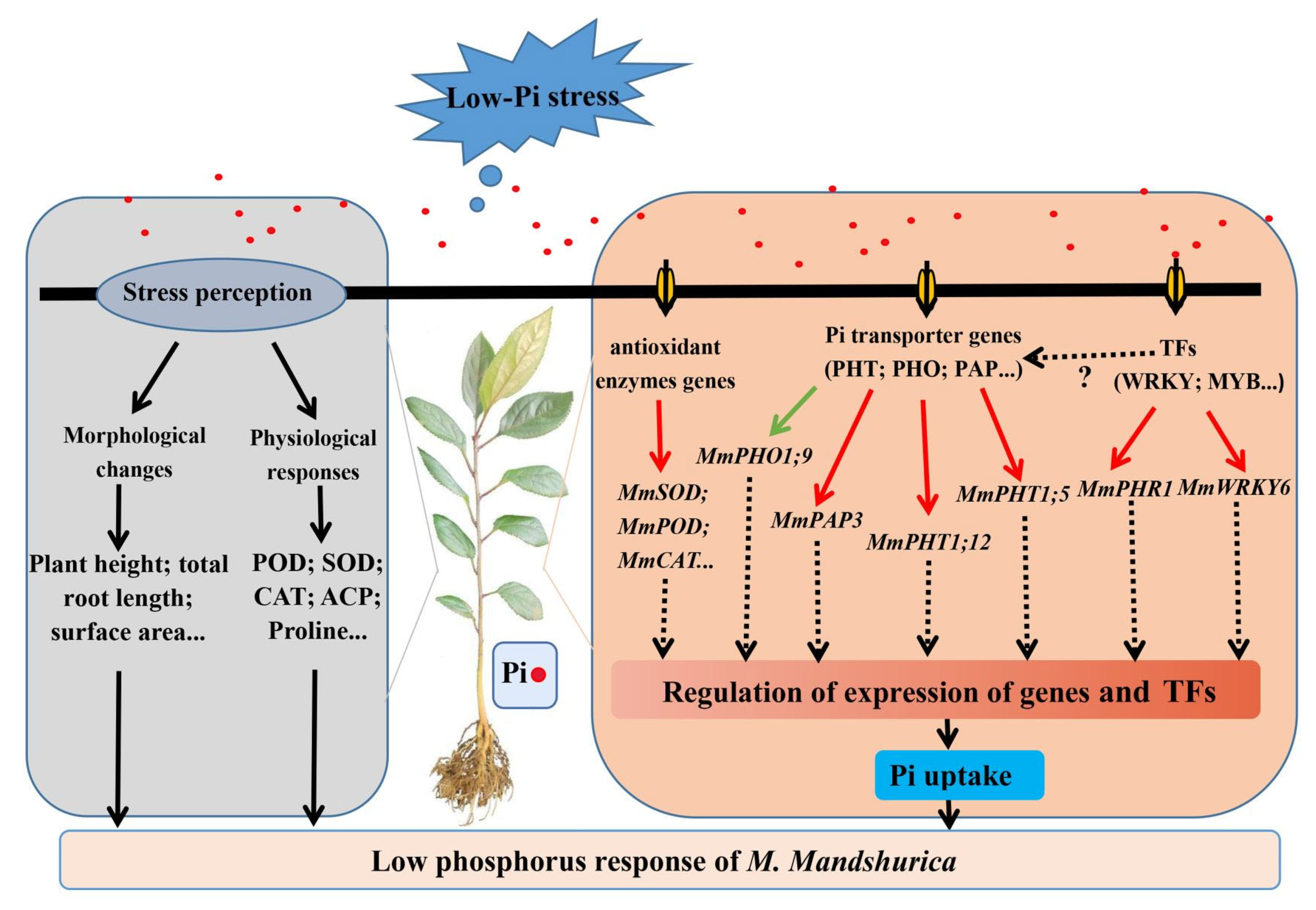
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jin, C.; Fang, C.; Yuan, H.; Wang, S.; Wu, Y.; Liu, X.; Zhang, Y.; Luo, J. Interaction between carbon metabolism and phosphate accumulation is revealed by a mutation of a cellulose synthase−like protein, CSLF6. J. Exp. Bot. 2015, 66, 2557–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovill, W.D.; Huang, C.Y.; Mcdonald, G.K. Genetic approaches to enhancing phosphorus−use efficiency (PUE) in crops: Challenges and directions. Crop Pasture Sci. 2013, 64, 179–198. [Google Scholar] [CrossRef] [Green Version]
- Faucon, M.P.; Houben, D.; Reynoird, J.P.; Mercadal−Dulaurent, A.M.; Lambers, H. Advances and Perspectives to Improve the Phosphorus Availability in Cropping Systems for Agroecological Phosphorus Management. Adv. Agron. 2015, 134, 51–79. [Google Scholar] [CrossRef]
- López−Bucio, J.; Cruz−Ramırez, A.; Herrera−Estrella, L. The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 2003, 6, 280–287. [Google Scholar] [CrossRef]
- Deng, Q.W.; Luo, X.D.; Chen, Y.L.; Zhou, Y.; Zhang, F.T.; Hu, B.L.; Xie, J.K. Transcriptome analysis of phosphorus stress responsiveness in the seedlings of Dongxiang wild rice (Oryza rufipogon Griff.). Biol. Res. 2018, 51, 7. [Google Scholar] [CrossRef] [Green Version]
- Li, L.Q.; Huang, L.P.; Pan, G.; Liu, L.; Wang, X.Y.; Lu, L.M. Identifying the genes regulated by AtWRKY6 using comparative transcript and proteomic analysis under phosphorus deficiency. Int. J. Mol. Sci. 2017, 18, 1046. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Liao, H.; Lucas, W.J. Molecular mechanisms underlying phosphate sensing, signaling, and adaptation in plants. J. Integr. Plant Biol. 2014, 56, 192–220. [Google Scholar] [CrossRef]
- Wang, F.; Deng, M.; Xu, J.; Zhu, X.; Mao, C. Molecular mechanisms of phosphate transport and signaling in higher plants. In Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 74, pp. 114–122. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.; Liao, L.; Xu, J.; Liang, X.; Liu, W. Genome−Wide Identification and Functional Characterization of the Phosphate Transporter Gene Family in Sorghum. Biomolecules 2019, 9, 670. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Liu, F.; Zhou, B. Genome−Wide Identification and Expression Profile Analysis of the PHT1 Gene Family in Gossypium hirsutum and Its Two Close Relatives of Subgenome Donor Species. Int. J. Mol. Sci. 2020, 21, 4905. [Google Scholar] [CrossRef]
- Bhosale, R.; Giri, J.; Pandey, B.K.; Giehl, R.F.; Hartmann, A.; Traini, R.; Truskina, J.; Leftley, N.; Hanlon, M.; Swarup, K.; et al. A mechanistic framework for auxin dependent Arabidopsis root hair elongation to low external phosphate. Nat. Commun. 2018, 9, 1409. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Sun, K.; Su, X.; Pan, Y.X.; Zhang, Y.F.; Wang, X.P. Physiological responses and tolerance mechanisms to Pb in two xerophils: Salsola passerina Bunge and Chenopodium album L. J. Hazard. Mater. 2012, 205, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Song, H.; Cheng, H.; Hao, D.; Wang, H.; Kan, G.; Jin, H.; Yu, D. The Acid Phosphatase-Encoding Gene GmACP1 Contributes to Soybean Tolerance to Low−Phosphorus Stress. PLoS Genet. 2014, 10, e1004061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Liao, H.; Trull, M.C.; Beebe, S.E.; Lynch, J.P. Induction of a Major Leaf Acid Phosphatase Does Not Confer Adaptation to Low Phosphorus Availability in Common Bean. Plant Physiol. 2001, 125, 1901–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Chang, X.J.; Ye, Y.; Xie, W.B.; Wu, P.; Lian, X.M. Comprehensive Sequence and Whole−Life−Cycle Expression Profile Analysis of the Phosphate Transporter Gene Family in Rice. Mol. Plant 2011, 4, 1105–1122. [Google Scholar] [CrossRef] [Green Version]
- Fan, F.; Cui, B.; Zhang, T.; Qiao, G.; Ding, G.; Wen, X. The temporal transcriptomic response of Pinus massoniana seedlings to phosphorus deficiency. PLoS ONE 2014, 9, e105068. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Li, M.; Shao, Y.; Yu, L.; Ma, F. Comprehensive genomic identification and expression analysis of the phosphate transporter (PHT) gene family in apple. Front. Plant Sci. 2017, 8, 426. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.; Zhu, Z.; Peng, L.; Chen, Q.; Jiang, Y. Soil Nutrient Status and Leaf Nutrient Diagnosis in the Main Apple Producing Regions in China. Hortic. Plant J. 2018, 4, 89–93. [Google Scholar] [CrossRef]
- Qaswar, M.; Ahmed, W.; Huang, J.; Fan, H.; Shi, X.; Jiang, X.; Liu, K.; Xu, Y.; He, Z.; Asghar, W.; et al. Soil carbon (C), nitrogen (N) and phosphorus (P) stoichiometry drives phosphorus lability in paddy soil under long−term fertilization: A fractionation and path analysis study. PLoS ONE 2019, 14, e0218195. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Cao, G.; Wang, X.; Zhao, J.; Yan, X.; Liao, H. Isolation and application of effective nitrogen fixation rhizobial strains on low−phosphorus acid soils in South China. Chin. Sci. Bull. 2009, 54, 412–420. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Liang, Y.; Xing, L.; Mao, J.; Liu, Z.; Dong, F.; Meng, Y.; Han, M.; Zhao, C.; Bao, L.; et al. Transcriptome Analysis Reveals Multiple Hormones, Wounding and Sugar Signaling Pathways Mediate Adventitious Root Formation in Apple Rootstock. Int. J. Mol. Sci. 2018, 19, 2201. [Google Scholar] [CrossRef] [Green Version]
- Divi, U.K.; El Tahchy, A.; Vanhercke, T.; Petrie, J.R.; Robles-Martinez, J.A.; Singh, S.P. Transcriptional and biochemical responses of monoacylglycerol acyltransferase−mediated oil synthesis and associated senescence-like responses in Nicotiana Benthamiana. Front. Plant Sci. 2014, 5, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.; Cheng, D.; Yang, Y.; Zhang, G.; Qin, M.; Chen, J.; Chen, Y.; Jiang, M. Co−expression network analysis of the transcriptomes of rice roots exposed to various cadmium stresses reveals universal cadmium−responsive genes. BMC Plant Biol. 2017, 17, 194. [Google Scholar] [CrossRef]
- Grene, R. Oxidative stress and acclimation mechanisms in plants. In The Arabidopsis Book; The American Society of Plant Biologists: Rockville, MD, USA, 2002; Volume 1, p. e0036. [Google Scholar] [CrossRef] [Green Version]
- Péret, B.; Clément, M.; Nussaume, L.; Desnos, T. Root developmental adaptation to phosphate starvation: Better safe than sorry. Trends Plant Sci. 2011, 16, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Hocking, P.J.; Simpson, R.J.; George, T.S. Plant mechanisms to optimise access to soil phosphorus. Crop Pasture Sci. 2009, 60, 124–143. [Google Scholar] [CrossRef]
- Ryan, P.R.; Liao, M.; Delhaize, E.; Rebetzke, G.J.; Weligama, C.; Spielmeyer, W.; James, R.A. Early vigour improves phosphate uptake in wheat. J. Exp. Bot. 2015, 66, 7089–7100. [Google Scholar] [CrossRef] [Green Version]
- Carswell, C.; Grant, B.R.; Theodorou, M.E.; Harris, J.; Niere, J.O.; Plaxton, W.C. The fungicide phosphonate disrupts the phosphate-starvation response in Brassica nigra seedlings. Plant Physiol. 1996, 110, 105–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gao, H.; He, L.; Zhu, W.; Yan, L.; Chen, Q.; He, C. The PHOSPHATE1 genes participate in salt and Pi signaling pathways and play adaptive roles during soybean evolution. BMC Plant Biol. 2019, 19, 353. [Google Scholar] [CrossRef] [Green Version]
- Anandan, A.; Parameswaran, C.; Mahender, A.; Nayak, A.K.; Vellaikumar, S.; Balasubramaniasai, C.; Ali, J. Trait variations and expression profiling of OsPHT1 gene family at the early growth-stages under phosphorus−limited conditions. Sci. Rep. 2021, 11, 13563. [Google Scholar] [CrossRef]
- Roy, S.; Verma, B.C.; Banerjee, A.; Kumar, J.; Ray, U.S.; Mandal, N.P. Genetic diversity for drought and low−phosphorus tolerance in rice (Oryza sativa L.) varieties and donors adapted to rainfed drought-prone ecologies. Sci. Rep. 2021, 11, 13671. [Google Scholar] [CrossRef]
- Sun, T.; Zhang, J.; Zhang, Q.; Li, X.; Li, M.; Yang, Y.; Zhou, J.; Wei, Q.; Zhou, B. Transcriptome and metabolome analyses revealed the response mechanism of apple to different phosphorus stresses. Plant Physiol. Biochem. 2021, 167, 639–650. [Google Scholar] [CrossRef]
- Teng, K.; Tan, P.; Guo, W.; Yue, Y.; Fan, X.; Wu, J. Heterologous expression of a novel Zoysia japonica C2H2 zinc finger gene, ZjZFN1, improved salt tolerance in Arabidopsis. Front. Plant Sci. 2018, 9, 1159. [Google Scholar] [CrossRef] [PubMed]
- Rohman, M.M.; Islam, M.R.; Monsur, M.B.; Amiruzzaman, M.; Fujita, M.; Hasanuzzaman, M. Trehalose protects maize plants from salt stress and phosphorus deficiency. Plants 2019, 8, 568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veronica, N.; Subrahmanyam, D.; Vishnu Kiran, T.; Yugandhar, P.; Bhadana, V.P.; Padma, V.; Jayasree, G.; Voleti, S.R. Influence of low phosphorus concentration on leaf photosynthetic characteristics and antioxidant response of rice genotypes. Photosynthetica 2017, 55, 285–293. [Google Scholar] [CrossRef]
- Tewari, R.K.; Kumar, P.; Sharma, P.N. Oxidative stress and antioxidant responses in young leaves of mulberry plants grown under nitrogen, phosphorus or potassium deficiency. J. Integr. Plant Biol. 2007, 49, 313–322. [Google Scholar] [CrossRef]
- Juszczuk, I.; Malusa, M.; Rychter, A. Phosphate deficiency induced oxidative stress in bean (Phaseolus vulgaris L.). In Plant Nutrition; Springer: Berlin/Heidelberg, Germany, 2001; pp. 148–149. [Google Scholar] [CrossRef]
- Wu, W.; Lin, Y.; Liu, P.; Chen, Q.; Tian, J.; Liang, C. Association of extracellular dNTP utilization with a GmPAP1-like protein identified in cell wall proteomic analysis of soybean roots. J. Exp. Bot. 2018, 69, 603–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Gao, Y.; Lynch, J.P. Large crown root number improves topsoil foraging and phosphorus acquisition. Plant Physiol. 2018, 177, 90–104. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Versaw, W.K.; Pumplin, N.; Gomez, S.K.; Blaylock, L.A.; Harrison, M.J. Closely related members of the Medicago truncatula PHT1 phosphate transporter gene family encode phosphate transporters with distinct biochemical activities. J. Biol. Chem. 2008, 283, 24673–24681. [Google Scholar] [CrossRef] [Green Version]
- Teng, W.; Zhao, Y.Y.; Zhao, X.Q.; He, X.; Ma, W.Y.; Deng, Y.; Chen, X.P.; Tong, Y.P. Genome−wide identification, characterization, and expression analysis of PHT1 phosphate transporters in wheat. Front. Plant Sci. 2017, 8, 543. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Wang, X.; Hu, R.; Wang, Y.; Xiao, C.; Jiang, Y.; Zhang, X.; Zheng, C.; Fu, Y.F. The pattern of Phosphate transporter 1 genes evolutionary divergence in Glycine max L. BMC Plant Biol. 2013, 13, 48. [Google Scholar] [CrossRef] [Green Version]
- Grace, E.; Cotsaftis, O.; Tester, M.; Smith, F.; Smith, S. Arbuscular mycorrhizal inhibition of growth in barley cannot be attributed to extent of colonization, fungal phosphorus uptake or effects on expression of plant phosphate transporter genes. New Phytol. 2009, 181, 938–949. [Google Scholar] [CrossRef]
- Rausch, C.; Daram, P.; Brunner, S.; Jansa, J.; Laloi, M.; Leggewie, G.; Amrhein, N.; Bucher, M. A phosphate transporter expressed in arbuscule−containing cells in potato. Nature 2001, 414, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Cui, P.J.; Tian, Y.; Huang, Y.; Wang, H.F.; Liu, F.; Chen, Y.F. Maize ZmPT7 regulates Pi uptake and redistribution which is modulated by phosphorylation. Plant Biotechnol. J. 2020, 18, 2406–2419. [Google Scholar] [CrossRef] [PubMed]
- Loth-Pereda, V.; Orsini, E.; Courty, P.E.; Lota, F.; Kohler, A.; Diss, L.; Blaudez, D.; Chalot, M.; Nehls, U.; Bucher, M.; et al. Structure and expression profile of the phosphate Pht1 transporter gene family in mycorrhizal Populus trichocarpa. Plant Physiol. 2011, 156, 2141–2154. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Yuan, J.; Chang, X.; Yang, M.; Zhang, L.; Lu, K.; Lian, X. The phosphate transporter gene OsPht1; 4 is involved in phosphate homeostasis in rice. PLoS ONE 2015, 10, e0126186. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, S.; Upadhyay, M.K.; Srivastava, A.K.; Abdelrahman, M.; Suprasanna, P.; Tran, L.P. Cellular and subcellular phosphate transport machinery in plants. Int. J. Mol. Sci. 2018, 19, 1914. [Google Scholar] [CrossRef] [Green Version]
- Lapis−Gaza, H.R.; Jost, R.; Finnegan, P.M. Arabidopsis PHOSPHATE TRANSPORTER1 genes PHT1;8 and PHT1;9 are involved in root-to-shoot translocation of orthophosphate. BMC Plant Biol. 2014, 14, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.; Li, X.; Sun, D.; Chen, Y.; Xu, G.; Cao, Y.; Ma, L.Q. Expressing phosphate transporter PvPht2;1 enhances P transport to the chloroplasts and increases arsenic tolerance in Arabidopsis thaliana. Environ. Sci. Technol. 2021, 55, 2276–2284. [Google Scholar] [CrossRef]
- Liu, X.L.; Wang, L.; Wang, X.W.; Yan, Y.; Yang, X.L.; Xie, M.Y.; Hu, Z.; Shen, X.; Ai, H.; Lin, H.H.; et al. Mutation of the chloroplast−localized phosphate transporter OsPHT2; 1 reduces flavonoid accumulation and UV tolerance in rice. Plant J. 2020, 102, 53–67. [Google Scholar] [CrossRef]
- Guo, C.; Zhao, X.; Liu, X.; Zhang, L.; Gu, J.; Li, X.; Lu, W.; Xiao, K. Function of wheat phosphate transporter gene TaPHT2;1 in Pi translocation and plant growth regulation under replete and limited Pi supply conditions. Planta 2013, 237, 1163–1178. [Google Scholar] [CrossRef]
- Lin, W.Y.; Lin, S.I.; Chiou, T.J. Molecular regulators of phosphate homeostasis in plants. J. Exp. Bot. 2009, 60, 1427–1438. [Google Scholar] [CrossRef] [Green Version]
- Baek, D.; Kim, M.C.; Chun, H.J.; Kang, S.; Park, H.C.; Shin, G.; Park, J.; Shen, M.; Hong, H.; Kim, W.Y.; et al. Regulation of miR399f transcription by AtMYB2 affects phosphate starvation responses in Arabidopsis. Plant Physiol. 2013, 161, 362–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhou, X.; Xu, Y.; Yao, M.; Xie, F.; Gai, J.; Li, Y.; Yang, S. Soybean SPX1 is an important component of the response to phosphate deficiency for phosphorus homeostasis. Plant Sci. 2016, 248, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Osorio, M.B.; Ng, S.; Berkowitz, O.; De Clercq, I.; Mao, C.; Shou, H.; Whelan, J.; Jost, R. SPX4 Acts on PHR1-Dependent and -Independent Regulation of Shoot Phosphorus Status in Arabidopsis. Plant Physiol. 2019, 181, 332–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Sun, J.; Miao, J.; Guo, J.; Shi, Z.; He, M.; Chen, Y.; Zhao, X.; Li, B.; Han, F.; et al. A phosphate starvation response regulator Ta−PHR1 is involved in phosphate signalling and increases grain yield in wheat. Ann. Bot. 2013, 111, 1139–1153. [Google Scholar] [CrossRef] [Green Version]
- Ruan, W.; Guo, M.; Cai, L.; Hu, H.; Li, C.; Liu, Y.; Wu, Z.; Mao, C.; Yi, K.; Wu, P.; et al. Genetic manipulation of a high−affinity PHR1 target cis-element to improve phosphorous uptake in Oryza sativa L. Plant Mol. Biol. 2015, 87, 429–440. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, F.; Cui, W.; Chen, K.; Zhao, R.; Zhang, Z. The FvPHR1 transcription factor control phosphate homeostasis by transcriptionally regulating miR399a in woodland strawberry. Plant Sci. 2019, 280, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Luo, Y.; Wang, W.; Fan, N.; Wang, D.; Yang, C.; Jia, H. NtMYB12 positively regulates flavonol biosynthesis and enhances tolerance to low Pi stress in Nicotiana tabacum. Front. Plant Sci. 2020, 10, 1683. [Google Scholar] [CrossRef]
- Lu, M.; Cheng, Z.; Zhang, X.M.; Huang, P.; Fan, C.; Yu, G.; Chen, F.; Xu, K.; Chen, Q.; Miao, Y.; et al. Spatial divergence of PHR-PHT1 modules maintains phosphorus homeostasis in soybean nodules. Plant Physiol. 2020, 184, 236–250. [Google Scholar] [CrossRef]
- Wang, J.; Pan, W.; Nikiforov, A.; King, W.; Hong, W.; Li, W.; Han, Y.; Patton-Vogt, J.; Shen, J.; Cheng, L. Identification of two glycerophosphodiester phosphodiesterase genes in maize leaf phosphorus remobilization. Crop J. 2021, 9, 95–108. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Ren, Y.R.; Zheng, P.F.; Qu, F.J.; Song, L.Q.; You, C.X.; Wang, X.F.; Hao, Y.J. Functional identification of apple MdMYB2 gene in phosphate−starvation response. J. Plant Physiol. 2020, 244, 153089. [Google Scholar] [CrossRef]
- Miura, K.; Rus, A.; Sharkhuu, A.; Yokoi, S.; Karthikeyan, A.S.; Raghothama, K.G.; Baek, D.; Koo, Y.D.; Jin, J.B.; Bressan, R.A.; et al. The Arabidopsis SUMO E3 ligase SIZ1 controls phosphate deficiency responses. Proc. Natl. Acad. Sci. USA 2005, 102, 7760–7765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juszczuk, I.; Malusà, E.; Rychter, A.M. Oxidative stress during phosphate deficiency in roots of bean plants (Phaseolus vulgaris L.). J. Plant Physiol. 2001, 10, 1299–1305. [Google Scholar] [CrossRef]
- Shin, R.; Berg, R.H.; Schachtman, D.P. Reactive oxygen species and root hairs in Arabidopsis root response to nitrogen, phosphorus and potassium deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, W.M.; Epstein, E.; Bloom, A. Mineral Nutrition of Plants: Principles and Perspectives, 2nd ed.; Sinauer Associates, Inc. Publishers: Sunderland, MA, USA, 2004; ISBN 0-87893-172-4. [Google Scholar] [CrossRef]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Goth, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Civello, P.M.; Martinez, G.A.; Chaves, A.R.; Anon, M.C. Peroxidase from strawberry fruit (Fragaria ananassa Duch.): Partial purification and determination of some properties. J. Agric. Food Chem. 1995, 43, 2596–2601. [Google Scholar] [CrossRef]
- Dissing, J.; Dahl, O.; Svensmark, O. Phosphonic and arsonic acids as inhibitors of human red cell acid phosphatase and their use in affinity chromatography. Biochim. Biophys. Acta (BBA)-Enzymol. 1979, 569, 159–176. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I. Rapid determination of free proline for water−stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Kumar, N.; Kumar, S.; Vats, S.K.; Ahuja, P.S. Effect of altitude on the primary products of photosynthesis and the associated enzymes in barley and wheat. Photosynth. Res. 2006, 88, 63. [Google Scholar] [CrossRef]
- Matula, J. Determination of dissolved reactive and dissolved total phosphorus in water extract of soils. Plant Soil Environ. 2011, 57, 1–6. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S.L.; Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quan.ntification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Raw | Total Clean | Clean Reads | Q20% | Genome Total | Gene Total |
|---|---|---|---|---|---|---|
| Reads (M) Reads (M) (%) | Mapping (%) Mapping (%) | |||||
| LCK | 46.23 | 44.15 | 95.59 | 98.16 | 83.67 | 79.10 |
| LLP | 46.41 | 44.09 | 95.06 | 97.99 | 83.56 | 79.36 |
| RCK | 46.60 | 44.38 | 95.28 | 98.21 | 71.08 | 64.78 |
| RLP | 45.72 | 44.50 | 97.34 | 98.03 | 66.47 | 61.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Wu, Y.; Shen, L.; Hou, Q.; Wu, R.; Li, Z.; Deng, L.; Wen, X. Cross−Talk between Transcriptome Analysis and Physiological Characterization Identifies the Genes in Response to the Low Phosphorus Stress in Malus mandshurica. Int. J. Mol. Sci. 2022, 23, 4896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094896
Zhao H, Wu Y, Shen L, Hou Q, Wu R, Li Z, Deng L, Wen X. Cross−Talk between Transcriptome Analysis and Physiological Characterization Identifies the Genes in Response to the Low Phosphorus Stress in Malus mandshurica. International Journal of Molecular Sciences. 2022; 23(9):4896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094896
Chicago/Turabian StyleZhao, Hong, Yawei Wu, Luonan Shen, Qiandong Hou, Rongju Wu, Zhengchun Li, Lin Deng, and Xiaopeng Wen. 2022. "Cross−Talk between Transcriptome Analysis and Physiological Characterization Identifies the Genes in Response to the Low Phosphorus Stress in Malus mandshurica" International Journal of Molecular Sciences 23, no. 9: 4896. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094896