
Nitric Oxide Synthase Blockade Impairs Spontaneous Calcium Activity in Mouse Primary Hippocampal Culture Cells

Abstract
:1. Introduction
2. Results
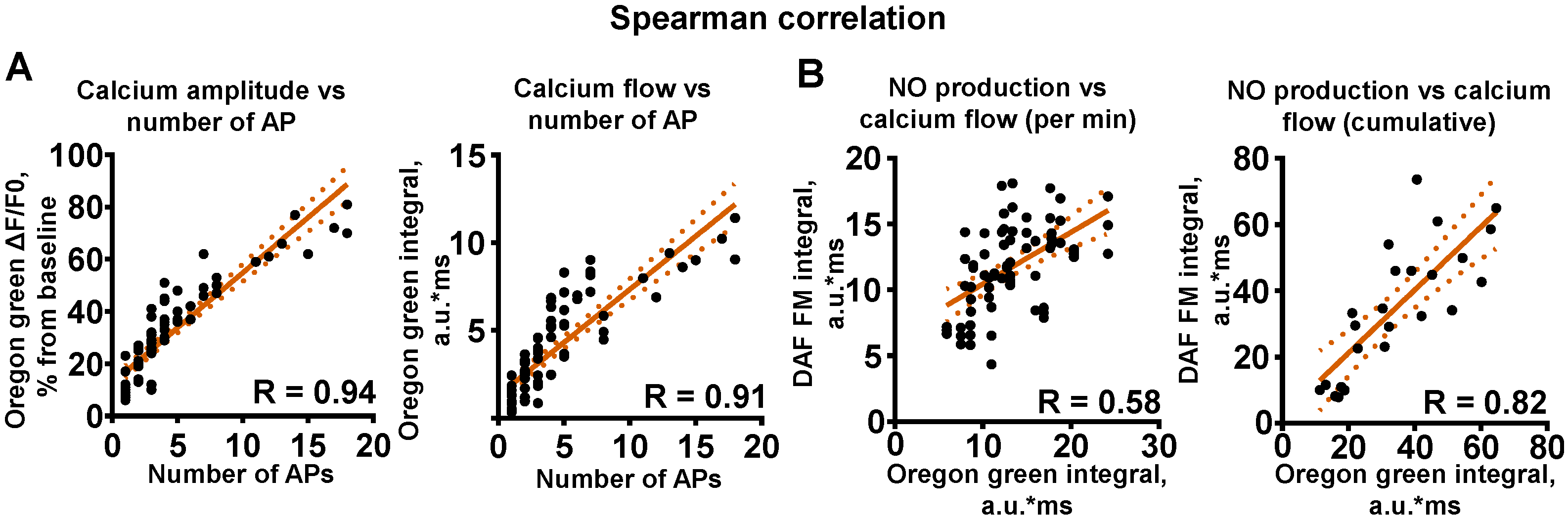
2.1. NO Synthesis in Hippocampal Primary Cultures
2.2. Spontaneous Electric Activity in Cultured Neurons Is Closely Related to Calcium Oscillations
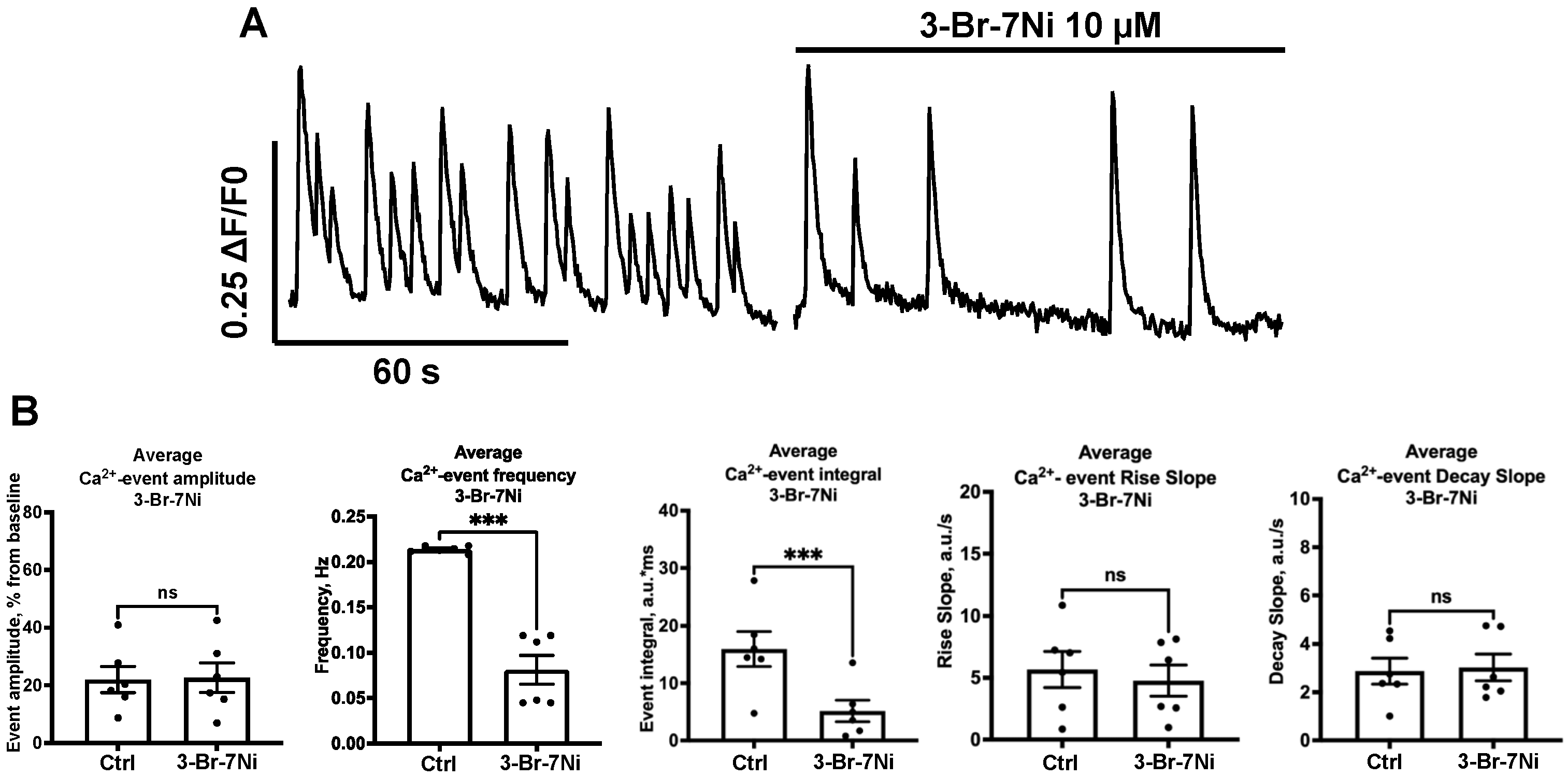
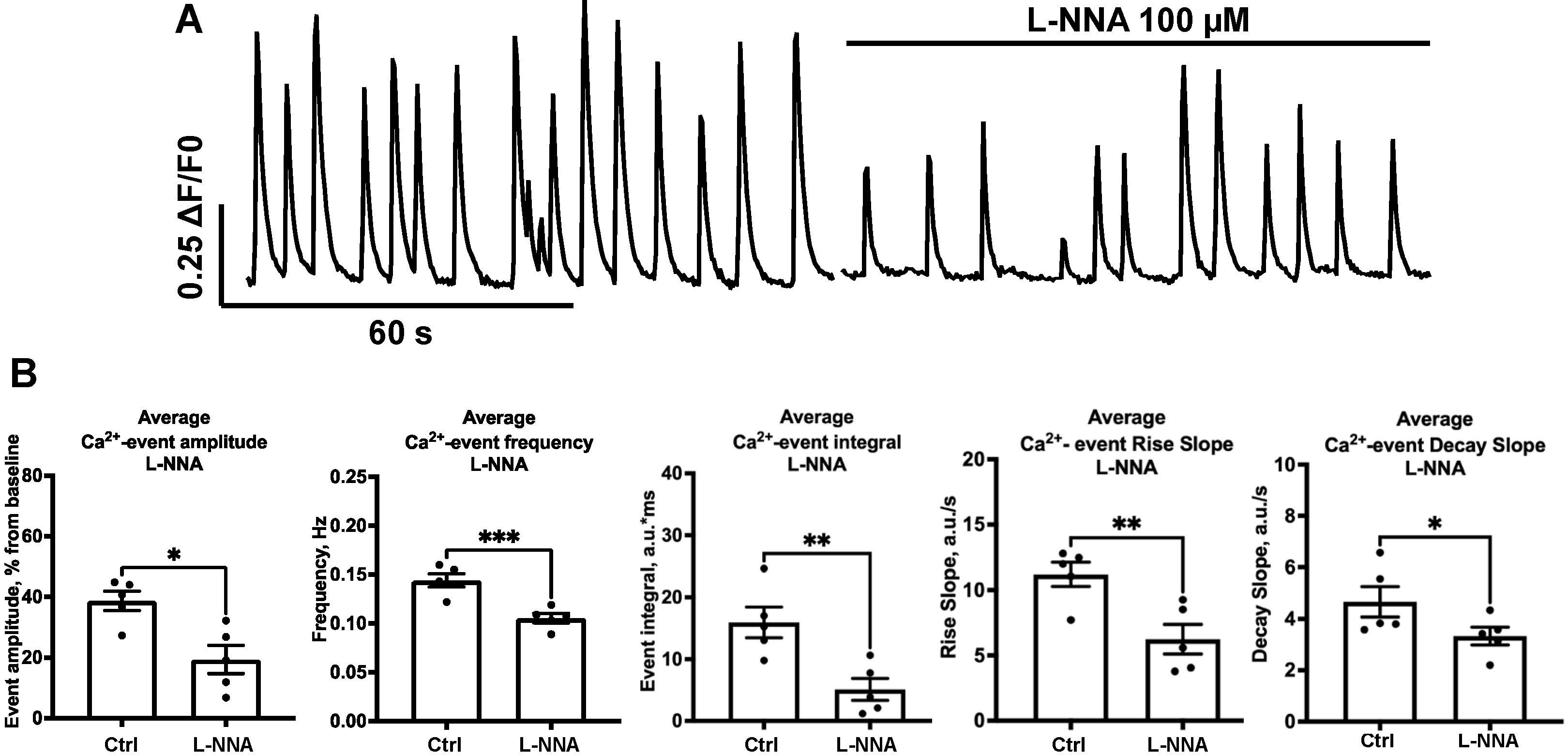
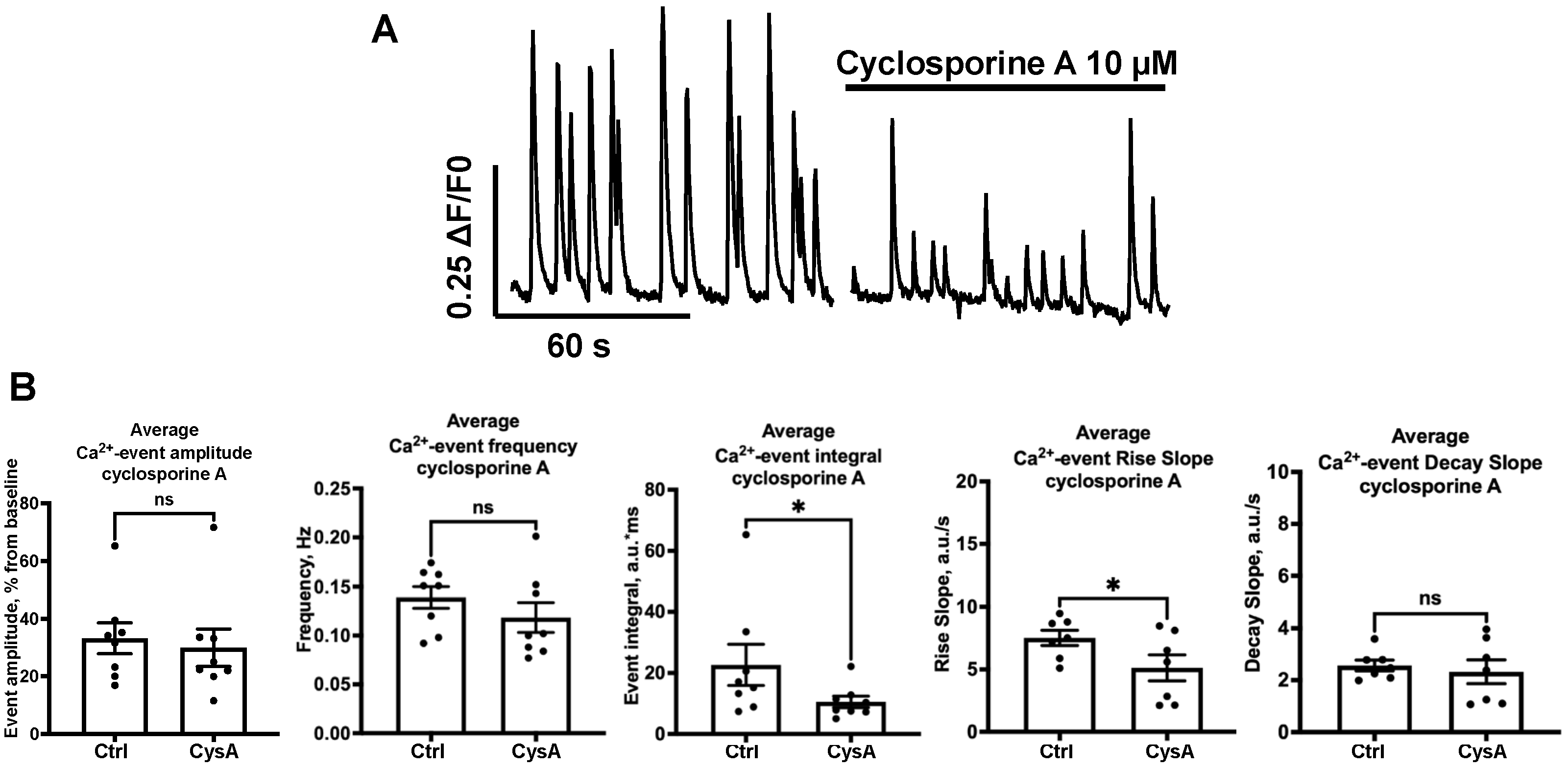
2.3. Effect of NOS-Blockade on Calcium Activity Is Associated with Spontaneous Neuronal Firing
3. Discussion
4. Materials and Methods
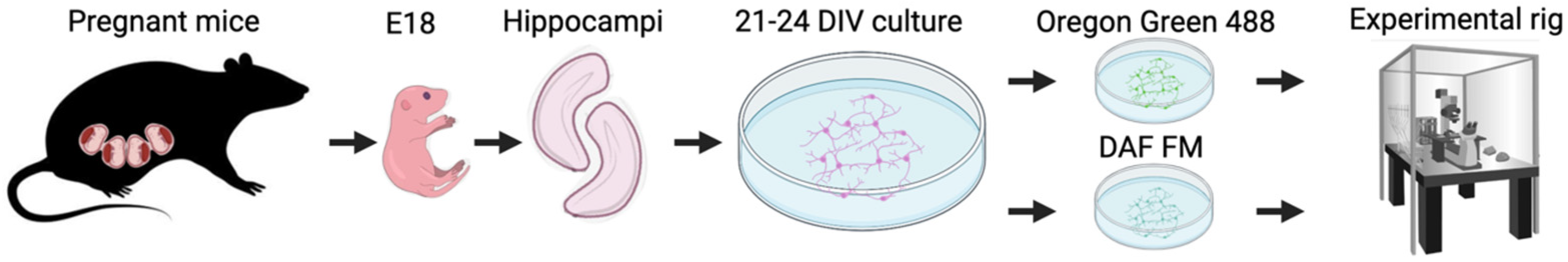
4.1. Cell Culture Preparation
4.2. Intracellular Calcium Recordings
4.3. Intracellular NO Recordings
4.4. Whole-Cell Patch-Clamp Recordings
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murad, F. Nitric Oxide: The Coming of the Second Messenger. Rambam Maimonides Med. J. 2011, 2, e0038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farah, C.; Michel, L.Y.M.; Balligand, J.-L. Nitric oxide signalling in cardiovascular health and disease. Nat. Rev. Cardiol. 2018, 15, 292–316. [Google Scholar] [CrossRef] [PubMed]
- Gregg, A.R. Mouse Models and the Role of Nitric Oxide in Reproduction. Curr. Pharm. Des. 2003, 9, 391–398. [Google Scholar] [CrossRef]
- Weigert, A.; Brüne, B. Nitric oxide, apoptosis and macrophage polarization during tumor progression. Nitric Oxide 2008, 19, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Zanelli, S.; Naylor, M.; Kapur, J. Nitric oxide alters GABAergic synaptic transmission in cultured hippocampal neurons. Brain Res. 2009, 1297, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charriaut-Marlangue, C.; Bonnin, P.; Pham, H.; Loron, G.; Leger, P.-L.; Gressens, P.; Renolleau, S.; Baud, O. Nitric oxide signaling in the brain: A new target for inhaled nitric oxide? Ann. Neurol. 2013, 73, 442–448. [Google Scholar] [CrossRef]
- Zhou, L.; Zhu, D.-Y. Neuronal nitric oxide synthase: Structure, subcellular localization, regulation, and clinical implications. Nitric Oxide 2009, 20, 223–230. [Google Scholar] [CrossRef]
- Yin, Y.; Zhang, N.; Diao, Y.; Hua, R.; Wang, J.; Han, S.; Li, J. Nitric oxide-mediated pathways and its role in the degenerative diseases. Front. Biosci. 2017, 22, 824–834. [Google Scholar] [CrossRef] [Green Version]
- Lovick, T.A.; Brown, L.A.; Key, B.J. Neurovascular relationships in hippocampal slices: Physiological and anatomical studies of mechanisms underlying flow-metabolism coupling in intraparenchymal microvessels. Neuroscience 1999, 92, 47–60. [Google Scholar] [CrossRef]
- Hardingham, N.; Dachtler, J.; Fox, K. The role of nitric oxide in pre-synaptic plasticity and homeostasis. Front. Cells Neurosci. 2013, 7, 190. [Google Scholar] [CrossRef]
- Brenman, J.E.; Bredt, D.S. Synaptic signaling by nitric oxide. Curr. Opin. Neurobiol. 1997, 7, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Araki, S.; Osuka, K.; Takata, T.; Tsuchiya, Y.; Watanabe, Y. Coordination between Calcium/Calmodulin-Dependent Protein Kinase II and Neuronal Nitric Oxide Synthase in Neurons. Int. J. Mol. Sci. 2020, 21, 7997. [Google Scholar] [CrossRef] [PubMed]
- Bredt, D.S.; Ferris, C.D.; Snyder, S.H. Nitric oxide synthase regulatory sites. Phosphorylation by cyclic AMP-dependent protein kinase, protein kinase C, and calcium/calmodulin protein kinase; identification of flavin and calmodulin binding sites. J. Biol. Chem. 1992, 267, 10976–10981. [Google Scholar] [CrossRef]
- Dinerman, J.L.; Steiner, J.P.; Dawson, T.M.; Dawson, V.; Snyder, S.H. Cyclic nucleotide dependent phosphorylation of neuronal nitric oxide synthase inhibits catalytic activity. Neuropharmacology 1994, 33, 1245–1251. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, Y.; Nishio, M.; Naito, Y.; Yokokura, H.; Nimura, Y.; Hidaka, H.; Watanabe, Y. Regulation of Neuronal Nitric-oxide Synthase by Calmodulin Kinases. J. Biol. Chem. 1999, 274, 20597–20602. [Google Scholar] [CrossRef] [Green Version]
- Komeima, K.; Hayashi, Y.; Naito, Y.; Watanabe, Y. Inhibition of Neuronal Nitric-oxide Synthase by Calcium/ Calmodulin-dependent Protein Kinase IIα through Ser847 Phosphorylation in NG108-15 Neuronal Cells. J. Biol. Chem. 2000, 275, 28139–28143. [Google Scholar] [CrossRef] [Green Version]
- Steinert, J.R.; Chernova, T.; Forsythe, I.D. Nitric Oxide Signaling in Brain Function, Dysfunction, and Dementia. Neuroscientist 2010, 16, 435–452. [Google Scholar] [CrossRef]
- Dawson, T.M.; Steiner, J.P.; Dawson, V.L.; Dinerman, J.L.; Uhl, G.R.; Snyder, S.H. Immunosuppressant FK506 enhances phosphorylation of nitric oxide synthase and protects against glutamate neurotoxicity. Proc. Natl. Acad. Sci. USA 1993, 90, 9808–9812. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Krukoff, T.L. Adrenomedullin Stimulates Nitric Oxide Production from Primary Rat Hypothalamic Neurons: Roles of Calcium and Phosphatases. Mol. Pharmacol. 2007, 72, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Itakura, M.; Watanabe, I.; Sugaya, T.; Takahashi, M. Direct association of the unique C -terminal tail of transmembrane AMPA receptor regulatory protein γ-8 with calcineurin. FEBS J. 2014, 281, 1366–1378. [Google Scholar] [CrossRef]
- Schuh, K.; Uldrijan, S.; Telkamp, M.; Röthlein, N.; Neyses, L. The plasmamembrane calmodulin–dependent calcium pump: A major regulator of nitric oxide synthase I. J. Cell Biol. 2001, 155, 201–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, W.; Zhou, J.; Li, W.; Zhou, T.; Chen, Q.; Yang, F.; Wei, T. Plasma membrane calcium ATPase 4b inhibits nitric oxide generation through calcium-induced dynamic interaction with neuronal nitric oxide synthase. Protein Cell 2013, 4, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Kodama, T.; Koyama, Y. Nitric oxide from the laterodorsal tegmental neurons: Its possible retrograde modulation on norepinephrine release from the axon terminal of the locus coeruleus neurons. Neuroscience 2005, 138, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, A.V.; Bal, N.V.; Balaban, P.M. Serine/Threonine Phosphatases in LTP: Two B or Not to Be the Protein Synthesis Blocker-Induced Impairment of Early Phase. Int. J. Mol. Sci. 2021, 22, 4857. [Google Scholar] [CrossRef] [PubMed]
- Pigott, B.M.; Garthwaite, J. Nitric Oxide Is Required for L-Type Ca2+ Channel-Dependent Long-Term Potentiation in the Hippocampus. Front. Synaptic Neurosci. 2016, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Maltsev, A.V.; Bal, N.V.; Balaban, P.M. LTP suppression by protein synthesis inhibitors is NO-dependent. Neuropharmacology 2018, 146, 276–288. [Google Scholar] [CrossRef]
- Sammut, S.; Threlfell, S.; West, A.R. Nitric oxide-soluble guanylyl cyclase signaling regulates corticostriatal transmission and short-term synaptic plasticity of striatal projection neurons recorded in vivo. Neuropharmacology 2010, 58, 624–631. [Google Scholar] [CrossRef] [Green Version]
- Kuchibhotla, K.V.; Lattarulo, C.R.; Hyman, B.T.; Bacskai, B.J. Synchronous Hyperactivity and Intercellular Calcium Waves in Astrocytes in Alzheimer Mice. Science 2009, 323, 1211–1215. [Google Scholar] [CrossRef] [Green Version]
- Maltsev, A.V.; Kokoz, Y.M. Cardiomyocytes generating spontaneous Ca(2+)-transients as tools for precise estimation of sarcoplasmic reticulum Ca(2+) transport. Arch. Biochem. Biophys. 2020, 693, 108542. [Google Scholar] [CrossRef]
- Zumerle, S.; Calì, B.; Munari, F.; Angioni, R.; Di Virgilio, F.; Molon, B.; Viola, A. Intercellular Calcium Signaling Induced by ATP Potentiates Macrophage Phagocytosis. Cell Rep. 2019, 27, 1–10. [Google Scholar] [CrossRef]
- Brini, M.; Calì, T.; Ottolini, D.; Carafoli, E. Neuronal calcium signaling: Function and dysfunction. Cells Mol. Life Sci. 2014, 71, 2787–2814. [Google Scholar] [CrossRef]
- Pchitskaya, E.; Popugaeva, E.; Bezprozvanny, I. Calcium signaling and molecular mechanisms underlying neurodegenerative diseases. Cell Calcium 2018, 70, 87–94. [Google Scholar] [CrossRef]
- Gleichmann, M.; Mattson, M.P. Neuronal Calcium Homeostasis and Dysregulation. Antioxid. Redox Signal. 2011, 14, 1261–1273. [Google Scholar] [CrossRef] [Green Version]
- Merino, J.J.; Arce, C.; Naddaf, A.; Bellver-Landete, V.; Oset-Gasque, M.J.; González, M.P. The Nitric Oxide Donor SNAP-Induced Amino Acid Neurotransmitter Release in Cortical Neurons. Effects of Blockers of Voltage-Dependent Sodium and Calcium Channels. PLoS ONE 2014, 9, e90703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahern, G.P.; Hsu, S.; Jackson, M.B. Direct actions of nitric oxide on rat neurohypophysial K + channels. J. Physiol. 1999, 520, 165–176. [Google Scholar] [CrossRef]
- Ahern, G.P.; Klyachko, V.A.; Jackson, M.B. cGMP and S-nitrosylation: Two routes for modulation of neuronal excitability by NO. Trends Neurosci. 2002, 25, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Biel, M.; Sautter, A.; Ludwig, A.; Hofmann, F.; Zong, X. Cyclic nucleotide-gated channels—Mediators of NO:cGMP-regulated processes. Naunyn Schmiedeberg’s Arch. Pharmacol. 1998, 358, 140–144. [Google Scholar] [CrossRef] [PubMed]
- González-Forero, D.; Portillo, F.; Gómez, L.; Montero, F.; Kasparov, S.; Moreno-López, B. Inhibition of Resting Potassium Conductances by Long-Term Activation of the NO/cGMP/Protein Kinase G Pathway: A New Mechanism Regulating Neuronal Excitability. J. Neurosci. 2007, 27, 6302–6312. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Dempo, Y.; Ohashi, A.; Saito, M.; Toyoda, H.; Sato, H.; Koshino, H.; Maeda, Y.; Hirai, T. Nitric Oxide Activates Leak K+ Currents in the Presumed Cholinergic Neuron of Basal Forebrain. J. Neurophysiol. 2007, 98, 3397–3410. [Google Scholar] [CrossRef] [Green Version]
- Kawano, T.; Zoga, V.; Kimura, M.; Liang, M.-Y.; Wu, H.-E.; Gemes, G.; McCallum, J.B.; Kwok, W.-M.; Hogan, Q.H.; Sarantopoulos, C.D. Nitric Oxide Activates ATP-Sensitive Potassium Channels in Mammalian Sensory Neurons: Action by Direct S-Nitrosylation. Mol. Pain 2009, 5, 12. [Google Scholar] [CrossRef]
- Sanders, K.M.; Koh, S.D. Two-pore-domain potassium channels in smooth muscles: New components of myogenic regulation. J. Physiol. 2005, 570, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.-H.; Bavencoffe, A.; Pan, H.-L. Molecular Basis of Regulating High Voltage-Activated Calcium Channels by S-Nitrosylation. J. Biol. Chem. 2015, 290, 30616–30623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mele, M.; Vieira, R.; Correia, B.; De Luca, P.; Duarte, F.V.; Pinheiro, P.S.; Duarte, C.B. Transient incubation of cultured hippocampal neurons in the absence of magnesium induces rhythmic and synchronized epileptiform-like activity. Sci. Rep. 2021, 11, 11374. [Google Scholar] [CrossRef]
- Molnár, E. Long-term potentiation in cultured hippocampal neurons. Semin. Cell Dev. Biol. 2011, 22, 506–513. [Google Scholar] [CrossRef] [PubMed]
- A Wagenaar, D.; Pine, J.; Potter, S.M. An extremely rich repertoire of bursting patterns during the development of cortical cultures. BMC Neurosci. 2006, 7, 11. [Google Scholar] [CrossRef]
- Sokolov, R.A.; Mukhina, I.V. Spontaneous Ca2+ events are linked to the development of neuronal firing during maturation in mice primary hippocampal culture cells. Arch. Biochem. Biophys. 2022, 727, 109330. [Google Scholar] [CrossRef]
- Bredt, D.S.; Hwang, P.M.; Snyder, S.H. Localization of nitric oxide synthase indicating a neural role for nitric oxide. Nature 1990, 347, 768–770. [Google Scholar] [CrossRef]
- Feng, M.-G.; Prieto, M.C.; Navar, L.G. Nebivolol-induced vasodilation of renal afferent arterioles involves β3-adrenergic receptor and nitric oxide synthase activation. Am. J. Physiol. Physiol. 2012, 303, F775–F782. [Google Scholar] [CrossRef] [Green Version]
- Negri, S.; Faris, P.; Pellavio, G.; Botta, L.; Orgiu, M.; Forcaia, G.; Sancini, G.; Laforenza, U.; Moccia, F. Group 1 metabotropic glutamate receptors trigger glutamate-induced intracellular Ca2+ signals and nitric oxide release in human brain microvascular endothelial cells. Cells Mol. Life Sci. 2020, 77, 2235–2253. [Google Scholar] [CrossRef]
- Tiburcio-Félix, R.; Cisneros, B.; Hernández-Kelly, L.C.R.; Hernández-Contreras, M.A.; Luna-Herrera, J.; Rea-Hernández, I.; Jiménez-Aguilar, R.; Olivares-Bañuelos, T.N.; Ortega, A. Neuronal Nitric Oxide Synthase in Cultured Cerebellar Bergmann Glia: Glutamate-Dependent Regulation. ACS Chem. Neurosci. 2019, 10, 2668–2675. [Google Scholar] [CrossRef]
- Kakizawa, S.; Yamazawa, T.; Chen, Y.; Ito, A.; Murayama, T.; Oyamada, H.; Kurebayashi, N.; Sato, O.; Watanabe, M.; Mori, N.; et al. Nitric oxide-induced calcium release via ryanodine receptors regulates neuronal function. EMBO J. 2011, 31, 417–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoyanovsky, D.; Murphy, T.; Anno, P.R.; Kim, Y.-M.; Salama, G. Nitric oxide activates skeletal and cardiac ryanodine receptors. Cell Calcium 1997, 21, 19–29. [Google Scholar] [CrossRef]
- Willmott, N.J.; Galione, A.; Smith, P.A. Nitric oxide induces intracellular Ca2+ mobilization and increases secretion of incorporated 5-hydroxytryptamine in rat pancreatic β-cells. FEBS Lett. 1995, 371, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanderson, J.L.; Freund, R.K.; Gorski, J.A.; Dell’Acqua, M.L. β-Amyloid disruption of LTP/LTD balance is mediated by AKAP150-anchored PKA and Calcineurin regulation of Ca2+-permeable AMPA receptors. Cell Rep. 2021, 37, 109786. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Cat.# |
|---|---|
| Abcam GTP trisodium salt | ab146528 |
| Applichem Calcium Chloride | A3652,1000 |
| Applichem D-glucose | 141341 |
| Applichem Sodium Chloride | 141659 |
| Applichem Magnesium Chloride 6-hydrate | 141396 |
| Applichem Potassium Chloride | A2939,1000 |
| Sigma Adenosine 5′-triphosphate disodium salt hydrate | A26209-5G |
| Sigma HEPES | H3375-250G |
| Sigma L-Ascorbic acid | A4544-25G |
| Sigma Potassium D-gluconate | G4500-100G |
| Tocris 3-Bromo-7-nitroindazole | 0735 |
| Sigma Nω-Nitro-L-arginine | N5501 |
| Tocris Cyclosporine A | 1101 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokolov, R.A.; Jappy, D.; Podgorny, O.V.; Mukhina, I.V. Nitric Oxide Synthase Blockade Impairs Spontaneous Calcium Activity in Mouse Primary Hippocampal Culture Cells. Int. J. Mol. Sci. 2023, 24, 2608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032608
Sokolov RA, Jappy D, Podgorny OV, Mukhina IV. Nitric Oxide Synthase Blockade Impairs Spontaneous Calcium Activity in Mouse Primary Hippocampal Culture Cells. International Journal of Molecular Sciences. 2023; 24(3):2608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032608
Chicago/Turabian StyleSokolov, Rostislav A., David Jappy, Oleg V. Podgorny, and Irina V. Mukhina. 2023. "Nitric Oxide Synthase Blockade Impairs Spontaneous Calcium Activity in Mouse Primary Hippocampal Culture Cells" International Journal of Molecular Sciences 24, no. 3: 2608. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032608