
Paclobutrazol Promotes Root Development of Difficult-to-Root Plants by Coordinating Auxin and Abscisic Acid Signaling Pathways in Phoebe bournei
 , , and
, , and 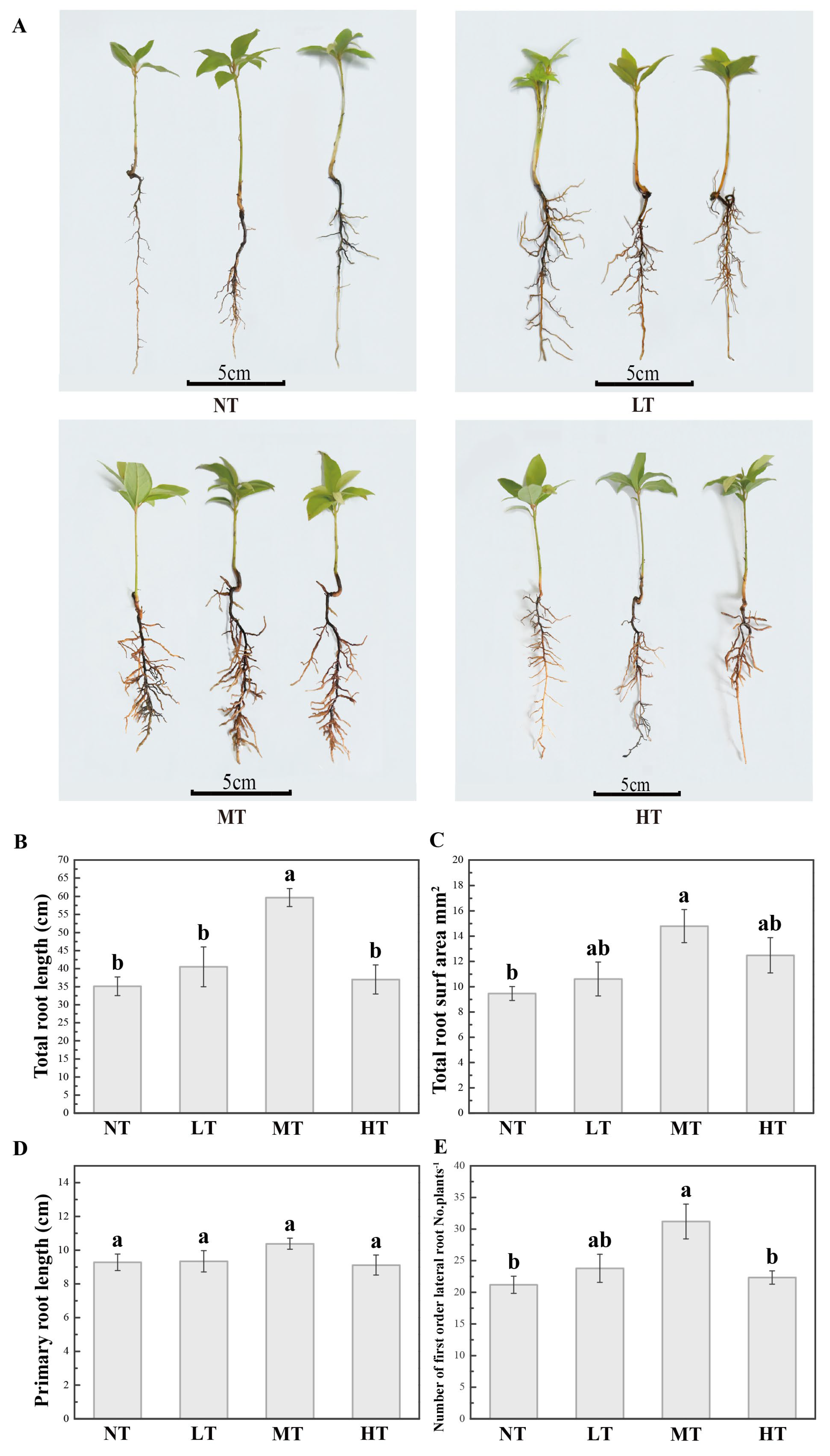{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Moderate-Concentration Paclobutrazol Promoted Root Growth and Development in P. bournei
2.2. Paclobutrazol Induced Genome-Wide Aberrant Expression of Genes in P. bournei
2.3. Paclobutrazol Regulates Root Growth and Development by Mediating IAA and ABA Biosynthesis and Signal Transduction in P. bourne
3. Discussion
3.1. Paclobutrazol Enhances Root Growth and Development by Promoting IAA Biosynthesis and Signaling Pathway in Difficult-to-Root Rare Plants
3.2. Paclobutrazol Inhibited ABA Biosynthesis and Signaling Pathway in the Roots of Difficult-to-Root Rare Plants
3.3. Interaction of ABA and IAA in Response to Paclobutrazol Treatments
3.4. Roles of GA3 in Root Growth and Response to Paclobutrazol Treatment in the Difficult-to-Root Rare Plants
4. Materials and Methods
4.1. Plant Growth and Paclobutrazol Treatment
4.2. Analysis of Root Morphological Traits
4.3. Endogenous Hormones Quantification
4.4. RNA-Sequencing and DEGs Function Analysis
4.5. Weighted Gene Co-Expression Network Analysis
4.6. qRT-PCR Validation of DEGs
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthes-Sears, U.; Larson, D.W. Limitations to seedling growth and survival by the quantity and quality of rooting space: Implications for the establishment of Thuja occidentalis on cliff faces. Int. J. Plant Sci. 1999, 160, 122–128. [Google Scholar] [CrossRef]
- Fan, B.; McHugh, A.D.; Guo, S.; Ma, Q.; Zhang, J.; Zhang, X.; Zhang, W.; Du, J.; Yu, Q.; Zhao, C. Factors influencing the natural regeneration of the pioneering shrub Calligonum mongolicum in sand dune stabilization plantations in arid deserts of northwest China. Ecol. Evol. 2018, 8, 2975–2984. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.J.; Shao, X.H.; Xue, P.P.; Tian, Y.Q.; Xiao, Z.P.; Wu, Y.P. Root sprouting ability and growth dynamics of the rootsuckers of Emmenopterys henryi, a rare and endangered plant endemic to China. For. Ecol. Manag. 2017, 389, 35–45. [Google Scholar] [CrossRef]
- Acevedo, M.; Alvarez, C.; Cartes, E.; Dumroese, R.K.; Gonzalez, M. Production and establishment techniques for the restoration of Nothofagus alessandrii, an endangered keystone species in a Mediterranean forest. New For. 2020, 51, 159–174. [Google Scholar] [CrossRef]
- Waidmann, S.; Sarkel, E.; Kleine-Vehn, J. Same same, but different: Growth responses of primary and lateral roots. J. Exp. Bot. 2020, 71, 2397–2411. [Google Scholar] [CrossRef]
- Patrick, T.M.A.; Peter, T.M.; Tendayi, G.E. Propagation potential for the conservation of Brackenridgea zanguebarica Oliv. a critically endangered plant species endemic to Vhembe District in Limpopo Province (South Africa). J. Appl. Bot. Food Qual. 2020, 93, 59–65. [Google Scholar]
- Desta, B.; Amare, G. Paclobutrazol as a plant growth regulator. Chem. Biol. Technol. Agric. 2021, 8, 1–15. [Google Scholar] [CrossRef]
- Wang, Y.L.; Bai, X.M.; Luo, R.F.; Lu, C.; Xiang, F. Effect of paclobutrazol on the extension and root characteristics of Lolium perenne L. J. Desert Res. 2010, 30, 1319–1324. [Google Scholar]
- Liu, B.; Long, S.; Liu, K.; Zhu, T.; Gong, J.; Gao, S.; Wang, R.; Zhang, L.; Liu, T.; Xu, Y. Paclobutrazol ameliorates low-light-induced damage by improving photosynthesis, antioxidant defense system, and regulating hormone levels in Tall Fescue. Int. J. Mol. Sci. 2022, 23, 9966. [Google Scholar] [CrossRef]
- Fan, Z.X.; Li, S.C.; Sun, H.L. Paclobutrazol modulates physiological and hormonal changes in Amorpha fruticosa under drought stress. Russ. J. Plant Physiol. 2020, 67, 122–130. [Google Scholar] [CrossRef]
- Nazari, F.; Javadi, T. Growth and development of Lolium perenne L.‘Barbal’in response to different concentrations of paclobutrazol. J. Biol. Environ. Sci. 2012, 6, 195–198. [Google Scholar]
- Blázquez, M.A.; Nelson, D.C.; Weijers, D. Evolution of plant hormone response pathways. Ecol. Evol. 2020, 71, 327–353. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Zhang, Y. Adaptive growth: Shaping auxin-mediated root system architecture. Trends Plant Sci. 2020, 25, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Pacifici, E.; Polverari, L.; Sabatini, S. Plant hormone cross-talk: The pivot of root growth. J. Exp. Bot. 2015, 66, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yuan, W.; Wang, Q.; Cao, Y.; Xu, F.; Dodd, I.C.; Xu, W. ABA regulation of root growth during soil drying and recovery can involve auxin response. Plant Cell Environ. 2022, 45, 871–883. [Google Scholar] [CrossRef] [PubMed]
- Olatunji, D.; Geelen, D.; Verstraeten, I. Control of endogenous auxin levels in plant root development. Int. J. Mol. Sci. 2017, 18, 2587. [Google Scholar] [CrossRef]
- Habets, M.E.J.; Offringa, R. PIN-driven polar auxin transport in plant developmental plasticity: A key target for environmental and endogenous signals. New Phytol. 2014, 203, 362–377. [Google Scholar] [CrossRef]
- Casanova-Sáez, R.; Mateo-Bonmatí, E.; Ljung, K. Auxin metabolism in plants. Csh. Perspect. Biol. 2021, 13, a039867. [Google Scholar] [CrossRef]
- Ma, W.; Li, J.; Qu, B.; He, X.; Zhao, X.; Li, B.; Fu, X.; Tong, Y. Auxin biosynthetic gene TAR2 is involved in low nitrogen-mediated reprogramming of root architecture in Arabidopsis. Plant J. 2014, 78, 70–79. [Google Scholar] [CrossRef]
- Gallei, M.; Luschnig, C.; Friml, J. Auxin signalling in growth: Schrodinger’s cat out of the bag. Curr. Opin. Plant Biol. 2020, 53, 43–49. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Tan, S.; Li, Z.; Yuan, Z.; Glanc, M.; Domjan, D.; Wang, K.; Xuan, W.; Guo, Y. Root growth adaptation is mediated by PYLs ABA receptor-PP2A protein phosphatase complex. Adv. sci. 2020, 7, 1901455. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, C.; Wu, Z.; Jia, Y.; Wang, H.; Sun, S.; Mao, C.; Wang, X. Abscisic acid regulates auxin homeostasis in rice root tips to promote root hair elongation. Front. Plant Sci. 2017, 8, 1121. [Google Scholar] [CrossRef]
- Leftley, N.; Banda, J.; Pandey, B.; Bennett, M.; Voss, U. Uncovering how auxin optimizes root systems architecture in response to environmental stresses. Cold Spring Harb. Perspect. Biol. 2021, 13, a040014. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Liang, J.; Qiao, Y.; Yan, Y.; Li, L.; Dai, Y. Involvement of G1-to-S transition and AhAUX-dependent auxin transport in abscisic acid-induced inhibition of lateral root primodia initiation in Arachis hypogaea L. J. Plant Physiol. 2012, 169, 1102–1111. [Google Scholar] [CrossRef]
- Ding, Z.; Friml, J. Auxin regulates distal stem cell differentiation in Arabidopsis roots. Proc. Natl. Acad. Sci. USA 2010, 107, 12046–12051. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, W.; Zou, W.; Jiang, D.; Liu, X. Complete chloroplast genome sequences of two endangered Phoebe (Lauraceae) species. Bot. Stud. 2017, 58, 37. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.J.; Liu, Y.J.; Shen, A.H.; Lin, X.C. Fengshui forests conserve genetic diversity: A case study of Phoebe bournei (Hemsl.) Yang in southern China. Genet. Mol. Res. 2015, 14, 1986–1993. [Google Scholar] [CrossRef]
- Han, X.; Zhang, J.; Han, S.; Chong, S.L.; Meng, G.; Song, M.; Wang, Y.; Zhou, S.; Liu, C.; Lou, L. The chromosome-scale genome of Phoebe bournei reveals contrasting fates of terpene synthase (TPS)-a and TPS-b subfamilies. Plant Commun. 2022, 15, 973–990. [Google Scholar] [CrossRef] [PubMed]
- Li, T.H.; Min, X.H. Dormancy characteristics and germination requirements of Phoebe bournei seed. Sci. Hortic. 2020, 260, 108903. [Google Scholar] [CrossRef]
- Sun, J.; Xu, Y.; Ye, S.; Jiang, H.; Chen, Q.; Liu, F.; Zhou, W.; Chen, R.; Li, X.; Tietz, O.; et al. Arabidopsis ASA1 is important for jasmonate-mediated regulation of auxin biosynthesis and transport during lateral root formation. Plant Cell 2009, 21, 1495–1511. [Google Scholar] [CrossRef]
- Liu, W.C.; Song, R.F.; Zheng, S.Q.; Li, T.T.; Zhang, B.L.; Gao, X.; Lu, Y.-T. Coordination of plant growth and abiotic stress responses by tryptophan synthase β subunit 1 through modulation of tryptophan and ABA homeostasis in Arabidopsis. Mol. Plant. 2022, 15, 973–990. [Google Scholar] [CrossRef] [PubMed]
- Grieneisen, V.A.; Xu, J.; Marée, A.F.; Hogeweg, P.; Scheres, B.J.N. Auxin transport is sufficient to generate a maximum and gradient guiding root growth. Nature 2007, 449, 1008–1013. [Google Scholar] [CrossRef] [PubMed]
- Roychoudhry, S.; Kepinski, S. Auxin in root development. Cold Spring Harb. Perspect. Biol. 2022, 14, a039933. [Google Scholar] [CrossRef] [PubMed]
- Okushima, Y.; Fukaki, H.; Onoda, M.; Theologis, A.; Tasaka, M. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar] [CrossRef]
- Liao, X.; Li, M.; Liu, B.; Yan, M.; Yu, X.; Zi, H.; Liu, R.; Yamamuro, C. Interlinked regulatory loops of ABA catabolism and biosynthesis coordinate fruit growth and ripening in woodland strawberry. Proc. Natl. Acad. Sci. USA 2018, 115, E11542–E11550. [Google Scholar] [CrossRef]
- González-Guzmán, M.; Apostolova, N.; Bellés, J.M.; Barrero, J.M.; Piqueras, P.; Ponce, M.R.; Micol, J.L.; Serrano, R.; Rodríguez, P.L. The short-chain alcohol dehydrogenase ABA2 catalyzes the conversion of xanthoxin to abscisic aldehyde. Plant Cell 2002, 14, 1833–1846. [Google Scholar] [CrossRef]
- Saito, S.; Hirai, N.; Matsumoto, C.; Ohigashi, H.; Ohta, D.; Sakata, K.; Mizutani, M. Arabidopsis CYP707A s encode (+)-abscisic acid 8′-hydroxylase, a key enzyme in the oxidative catabolism of abscisic acid. Plant Physiol. 2004, 134, 1439–1449. [Google Scholar] [CrossRef]
- Miao, R.; Yuan, W.; Wang, Y.; Garcia-Maquilon, I.; Dang, X.; Li, Y.; Zhang, J.; Zhu, Y.; Rodriguez, P.L.; Xu, W. Low ABA concentration promotes root growth and hydrotropism through relief of ABA INSENSITIVE 1-mediated inhibition of plasma membrane H(+)-ATPase 2. Sci. Adv. 2021, 7, eabd4113. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.T.; Xu, H.; Zhang, K.; Guo, T.; Lu, Y. Glucose inhibits root meristem growth via ABA INSENSITIVE 5, which represses PIN1 accumulation and auxin activity in Arabidopsis. Plant Cell Environ. 2014, 37, 1338–1350. [Google Scholar] [CrossRef]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA perception and signalling. Trends Plant Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef]
- Xie, Q.; Essemine, J.; Pang, X.; Chen, H.; Jin, J.; Cai, W. Abscisic acid regulates the root growth trajectory by reducing auxin transporter PIN2 protein levels in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 632676. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xing, L.; Wang, X.; Hou, Y.J.; Gao, J.; Wang, P.; Duan, C.G.; Zhu, X.; Zhu, J.K. The ABA receptor PYL8 promotes lateral root growth by enhancing MYB77-dependent transcription of auxin-responsive genes. Sci. Signal. 2014, 7, ra53. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Cai, F.X.; Gao, H.J.; Zhang, S.Y.; Wang, K.; Liu, T.; Wang, X. ABA plays essential roles in regulating root growth by interacting with auxin and MAPK signaling pathways and cell-cycle machinery in rice seedlings. Plant Growth Regul. 2015, 75, 535–547. [Google Scholar] [CrossRef]
- Hedden, P. The current status of research on gibberellin biosynthesis. Plant Cell Physiol. 2020, 61, 1832–1849. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.J.; Hua, C.M.; Shen, L.S.; Yu, H. New insights into gibberellin signaling in regulating flowering in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Que, F.; Xu, Z.S.; Wang, F.; Xiong, A.S. Exogenous gibberellin altered morphology, anatomic and transcriptional regulatory networks of hormones in carrot root and shoot. BMC Plant Biol. 2015, 15, 290. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.F.; Xu, N.X.; Ding, C.L.; Gu, H.R.; Zhang, W.J.; Li, X.L. Suitable gibberellic acid treatment for double-purpose rice (Oryza sativa L.) varieties at different harvest times. Field Crops Res. 2016, 193, 178–185. [Google Scholar] [CrossRef]
- Reid, J.B.; Davidson, S.E.; Ross, J.J. Auxin acts independently of DELLA proteins in regulating gibberellin levels. Plant Signal. Behav. 2011, 6, 406–408. [Google Scholar] [CrossRef]
- Chen, S.P.; Sun, W.H.; Xiong, Y.F.; Jiang, Y.T.; Liu, X.D.; Liao, X.Y.; Zhang, D.Y.; Jiang, S.Z.; Li, Y.; Liu, B.; et al. The Phoebe genome sheds light on the evolution of magnoliids. Hortic. Res. 2020, 7, 146. [Google Scholar] [CrossRef]
- Zhang, J.H.; Zhu, Y.J.; Pan, Y.; Huang, H.H.; Li, C.L.; Li, G.Z.; Tong, Z.K. Transcriptomic profiling and identification of candidate genes in two Phoebe bournei ecotypes with contrasting cold stress responses. Trees Struct. Funct. 2018, 32, 1315–1333. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Xu, P.; Zhang, B.; Song, Y.; Wen, S.; Bai, Y.; Ji, L.; Lai, Y.; He, G.; Zhang, D. Paclobutrazol Promotes Root Development of Difficult-to-Root Plants by Coordinating Auxin and Abscisic Acid Signaling Pathways in Phoebe bournei. Int. J. Mol. Sci. 2023, 24, 3753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043753
Li J, Xu P, Zhang B, Song Y, Wen S, Bai Y, Ji L, Lai Y, He G, Zhang D. Paclobutrazol Promotes Root Development of Difficult-to-Root Plants by Coordinating Auxin and Abscisic Acid Signaling Pathways in Phoebe bournei. International Journal of Molecular Sciences. 2023; 24(4):3753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043753
Chicago/Turabian StyleLi, Jing, Peiyue Xu, Baohong Zhang, Yanyan Song, Shizhi Wen, Yujie Bai, Li Ji, Yong Lai, Gongxiu He, and Dangquan Zhang. 2023. "Paclobutrazol Promotes Root Development of Difficult-to-Root Plants by Coordinating Auxin and Abscisic Acid Signaling Pathways in Phoebe bournei" International Journal of Molecular Sciences 24, no. 4: 3753. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043753