
Utilizing Red Spotted Apollo Butterfly Transcriptome to Identify Antimicrobial Peptide Candidates against Porphyromonas gingivalis

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains Preparation
2.2. P. bremeri Rearing and RNA Isolation
2.3. Next Generation Sequencing and Assembly
2.4. AMP Screening
2.5. Peptide Synthesis and Antimicrobial Activity
2.6. Cell Viability Assays
3. Results
3.1. In Silico AMP Prediction
3.2. Antimicrobial Activity
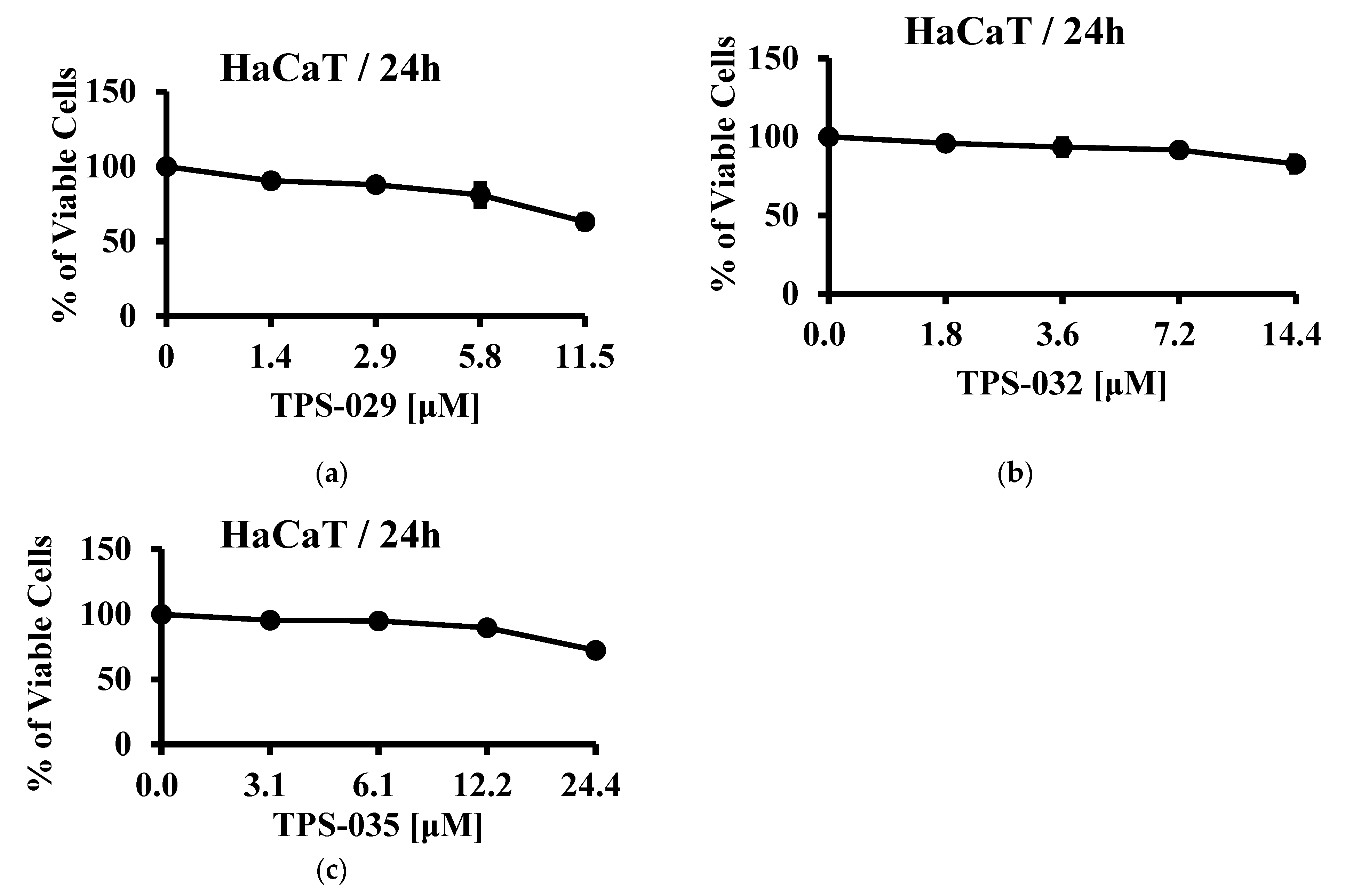
3.3. Cell Viability
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef] [PubMed]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial Resistance in Bacteria: Mechanisms, Evolution, and Persistence. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Lopez Romo, A.; Quiros, R. Appropriate use of antibiotics: An unmet need. Ther. Adv. Urol. 2019, 11, 1756287219832174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Chung, H.; Shin, Y.P.; Kim, M.A.; Natarajan, S.; Veerappan, K.; Kim, S.H.; Park, J.; Hwang, J.S. Deciphering Novel Antimicrobial Peptides from the Transcriptome of Papilio xuthus. Insects 2020, 11, 776. [Google Scholar] [CrossRef]
- Lee, J.H.; Chung, H.; Shin, Y.P.; Kim, I.W.; Natarajan, S.; Veerappan, K.; Seo, M.; Park, J.; Hwang, J.S. Transcriptome Analysis of Psacothea hilaris: De Novo Assembly and Antimicrobial Peptide Prediction. Insects 2020, 11, 676. [Google Scholar] [CrossRef]
- Park, Y.; Kim, Y.; Park, G.-W.; Lee, J.-O.; Lee, K.-W. Supercooling capacity along with up-regulation of glycerol content in an overwintering butterfly, Parnassius bremeri. J. Asia-Pac. Entomol. 2017, 20, 949–954. [Google Scholar] [CrossRef]
- Brady, D.; Grapputo, A.; Romoli, O.; Sandrelli, F. Insect Cecropins, Antimicrobial Peptides with Potential Therapeutic Applications. Int. J. Mol. Sci. 2019, 20, 5862. [Google Scholar] [CrossRef] [Green Version]
- Buonocore, F.; Fausto, A.M.; Della Pelle, G.; Roncevic, T.; Gerdol, M.; Picchietti, S. Attacins: A Promising Class of Insect Antimicrobial Peptides. Antibiotics 2021, 10, 212. [Google Scholar] [CrossRef]
- Badapanda, C.; Chikara, S.K. Lepidopteran Antimicrobial Peptides (AMPs): Overview, Regulation, Modes of Action, and Therapeutic Potentials of Insect-Derived AMPs. In Short Views on Insect Genomics and Proteomics: Insect Proteomics; Raman, C., Goldsmith, M.R., Agunbiade, T.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; Volume 2, pp. 141–163. [Google Scholar] [CrossRef]
- Willis, J.R.; Gabaldon, T. The Human Oral Microbiome in Health and Disease: From Sequences to Ecosystems. Microorganisms 2020, 8, 308. [Google Scholar] [CrossRef] [Green Version]
- Suwandecha, T.; Srichana, T.; Balekar, N.; Nakpheng, T.; Pangsomboon, K. Novel antimicrobial peptide specifically active against Porphyromonas gingivalis. Arch. Microbiol. 2015, 197, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ai, L.; Zhang, Y.; Cheng, J.; Yu, H.; Li, C.; Zhang, D.; Pan, Y.; Lin, L. The Effects of Antimicrobial Peptide Nal-P-113 on Inhibiting Periodontal Pathogens and Improving Periodontal Status. Biomed Res. Int. 2018, 2018, 1805793. [Google Scholar] [CrossRef] [Green Version]
- Taniguchi, M.; Ochiai, A.; Takahashi, K.; Nakamichi, S.I.; Nomoto, T.; Saitoh, E.; Kato, T.; Tanaka, T. Antimicrobial activity against Porphyromonas gingivalis and mechanism of action of the cationic octadecapeptide AmyI-1-18 and its amino acid-substituted analogs. J. Biosci. Bioeng. 2016, 122, 652–659. [Google Scholar] [CrossRef] [PubMed]
- FASTQC. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 16 April 2021).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, O.; Hara, Y.; Kuraku, S. gVolante for standardizing completeness assessment of genome and transcriptome assemblies. Bioinformatics 2017, 33, 3635–3637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Escamilla, A.M.; Rousseau, F.; Schymkowitz, J.; Serrano, L. Prediction of sequence-dependent and mutational effects on the aggregation of peptides and proteins. Nat. Biotechnol. 2004, 22, 1302–1306. [Google Scholar] [CrossRef] [PubMed]
- Conchillo-Sole, O.; de Groot, N.S.; Aviles, F.X.; Vendrell, J.; Daura, X.; Ventura, S. AGGRESCAN: A server for the prediction and evaluation of "hot spots" of aggregation in polypeptides. BMC Bioinform. 2007, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrent, M.; Di Tommaso, P.; Pulido, D.; Nogues, M.V.; Notredame, C.; Boix, E.; Andreu, D. AMPA: An automated web server for prediction of protein antimicrobial regions. Bioinformatics 2012, 28, 130–131. [Google Scholar] [CrossRef] [Green Version]
- Waghu, F.H.; Gopi, L.; Barai, R.S.; Ramteke, P.; Nizami, B.; Idicula-Thomas, S. CAMP: Collection of sequences and structures of antimicrobial peptides. Nucleic Acids Res. 2014, 42, D1154–D1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.T.; Lee, C.C.; Yang, J.R.; Lai, J.Z.; Chang, K.Y. A large-scale structural classification of antimicrobial peptides. Biomed Res. Int. 2015, 2015, 475062. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Park, S.; Shin, Y.-K.; Kang, H.; Kim, K.-Y. In vitro antibacterial activity of macelignan and corosolic acid against the bacterial bee pathogens Paenibacillus larvae and Melissococcus plutonius. Acta Vet. Brno 2018, 87, 277–284. [Google Scholar] [CrossRef]
- Park, S.; Kim, J.; Shin, Y.-K.; Kim, K.-Y. Antimicrobial activity of 4-hydroxyderricin, sophoraflavanone G, acetylshikonin, and kurarinone against the bee pathogenic bacteria Paenibacillus larvae and Melissococcus plutonius. J. Apic. Res. 2020, 1–5. [Google Scholar]
- Jenssen, H.; Hamill, P.; Hancock, R.E. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahlapuu, M.; Hakansson, J.; Ringstad, L.; Bjorn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [Green Version]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial peptides: Key components of the innate immune system. Crit. Rev. Biotechnol. 2012, 32, 143–171. [Google Scholar] [CrossRef] [Green Version]
- Torrent, M.; Andreu, D.; Nogues, V.M.; Boix, E. Connecting peptide physicochemical and antimicrobial properties by a rational prediction model. PLoS ONE 2011, 6, e16968. [Google Scholar] [CrossRef]
- Bostanci, N.; Belibasakis, G.N. Porphyromonas gingivalis: An invasive and evasive opportunistic oral pathogen. FEMS Microbiol. Lett. 2012, 333, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Zhou, W.; Wang, H.; Liang, S. Roles of Porphyromonas gingivalis and its virulence factors in periodontitis. Adv. Protein Chem. Struct. Biol. 2020, 120, 45–84. [Google Scholar] [CrossRef] [PubMed]
- Olsen, I.; Taubman, M.A.; Singhrao, S.K. Porphyromonas gingivalis suppresses adaptive immunity in periodontitis, atherosclerosis, and Alzheimer’s disease. J. Oral. Microbiol. 2016, 8, 33029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanagasingam, S.; Chukkapalli, S.S.; Welbury, R.; Singhrao, S.K. Porphyromonas gingivalis is a Strong Risk Factor for Alzheimer’s Disease. J. Alzheimers Dis. Rep. 2020, 4, 501–511. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Stains | Description | Medium | Temperature (°C) | Atmosphere |
|---|---|---|---|---|
| Staphylococcus epidermidis | KACC 12454 | NB | 30 | Aerobic |
| Klebsiella oxytoca | KCTC 1686 | NB | 37 | Aerobic |
| Salmonella typhimurium | CCARM 0240 | NB | 37 | Aerobic |
| Escherichia coli | KACC 11598 | TSB | 37 | Aerobic |
| Enterococcus faecalis | CCARM 5511 | TSB | 37 | Aerobic |
| Enterococcus faecium | KACC 11954 | TSB | 37 | Aerobic |
| Pseudomonas aeruginosa | KACC 14021 | TSB | 37 | Aerobic |
| Staphylococcus aureus | CCARM 3505 | TSB | 37 | Aerobic |
| Streptococcus mutans | KACC 16833 | TSB | 37 | Aerobic |
| Staphylococcus epidermidis | KACC 13234 | TSB | 37 | Aerobic |
| Fusobacterium nucleatum subsp. Nucleatum | KCTC 2640 | BHI | 37 | Aerobic |
| Actinomyces viscosus | KCTC 9146 | BHI | 37 | Anaerobic |
| Propionibacterium acnes | CCARM 9009 | BHI | 37 | Anaerobic |
| Porphyromonas gingivalis | KCTC 5352 | Modified TSB | 37 | Anaerobic |
| Streptococcus sobrinus | KCTC 5809 | Modified TSB | 37 | Anaerobic |
| Candida albicans | KCTC 7965 | YPD | 30 | Aerobic |
| Candida tropicalis | KCTC 7212 | YPD | 30 | Aerobic |
| Candida parapsilosis | KACC 49573 | YPD | 30 | Aerobic |
| Candida tropicalis var. tropicalis | KCTC 17762 | YPD | 30 | Aerobic |
| Candida parapsilosis var. parapsilosis | KACC 45480 | YPD | 30 | Aerobic |
| Candida glabrata | KCTC 7219 | YPD | 30 | Aerobic |
| Pichia guilliermondii | KCTC 7211 | YPD | 30 | Aerobic |
| Filobasidiella neoformans var. bacillispora | KCTC 17528 | YPD | 30 | Aerobic |
| Propensity | Tools | Descriptions/Parameters | Cutoff | No. of Sequences |
|---|---|---|---|---|
| Total Protein fragments | 266,300 | |||
| Physicochemical | Pepstats | Peptide Length | ≥2 to 50 | 189,818 |
| Pepstats | Charge | >0 (+) | 128,625 | |
| Pepstats | Isoelectric Point(pI) | ≥8 to ≤12 | 88,951 | |
| AMPA | Stretch | ≥1 | 152,758 | |
| Aggregation (Invivo) | Tango | AGG | ≤500 | 157,940 |
| Tango | Helix | ≥0 Helix ≤25 | 179,511 | |
| Tango | Beta | ≥25 Beta ≤100 | 88,470 | |
| Aggregation (Invitro) | Aggrescan | Na4vSS | ≥−40 Na4vSS ≤60 | 149,072 |
| Similarity | BlastP | Similarity | <80 | 25,246 |
| AMP | CAMP ADAM | Support Vector Machine (SVM) classifier | >0.5, AMP | 7574 |
| Random Forest Classifier | >0.5, AMP | 8571 | ||
| Artificial Neural Network (ANN) classifier | AMP | 12,051 | ||
| Discriminant Analysis classifier | >0.5, AMP | 9127 | ||
| Support Vector Machine (SVM) classifier | >0.5, AMP | 17,976 | ||
| Final | 3570 |
| Sequence | Mw (Da) | Porphyromonas gingivalis (KCTC5352) | Filobasidiella neoformans var. Bacillispora (KCTC17528) | Pichia guilliemondii (KCTC7211) | |
|---|---|---|---|---|---|
| Oxytetracycline | 460.4 | 108.6 (50) | ND | ND | |
| Miconazole | 416.1 | ND | 7.5 (3.125) | 7.5 (3.125) | |
| TPS-029 | RLFNYGLFSSKIIKHTIK | 2165.76 | >46.2 (>100) | 23.1 (50) | >46.2 (>100) |
| TPS-032 | RVLTHVFKCKLKLR | 1741.41 | 14.4 (25) | >57.4 (>100) | 28.7 (50) |
| TPS-035 | RCCKLVFR | 1024.42 | >97.6 (>100) | >97.6 (>100) | 97.6 (100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, K.-W.; Kim, J.-G.; Veerappan, K.; Chung, H.; Natarajan, S.; Kim, K.-Y.; Park, J. Utilizing Red Spotted Apollo Butterfly Transcriptome to Identify Antimicrobial Peptide Candidates against Porphyromonas gingivalis. Insects 2021, 12, 466. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050466
Lee K-W, Kim J-G, Veerappan K, Chung H, Natarajan S, Kim K-Y, Park J. Utilizing Red Spotted Apollo Butterfly Transcriptome to Identify Antimicrobial Peptide Candidates against Porphyromonas gingivalis. Insects. 2021; 12(5):466. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050466
Chicago/Turabian StyleLee, Kang-Woon, Jae-Goo Kim, Karpagam Veerappan, Hoyong Chung, Sathishkumar Natarajan, Ki-Young Kim, and Junhyung Park. 2021. "Utilizing Red Spotted Apollo Butterfly Transcriptome to Identify Antimicrobial Peptide Candidates against Porphyromonas gingivalis" Insects 12, no. 5: 466. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050466