Glycerolipid Composition of the Red Macroalga Agarophyton Chilensis and Comparison to the Closely Related Agarophyton Vermiculophyllum Producing Different Types of Eicosanoids †


Abstract
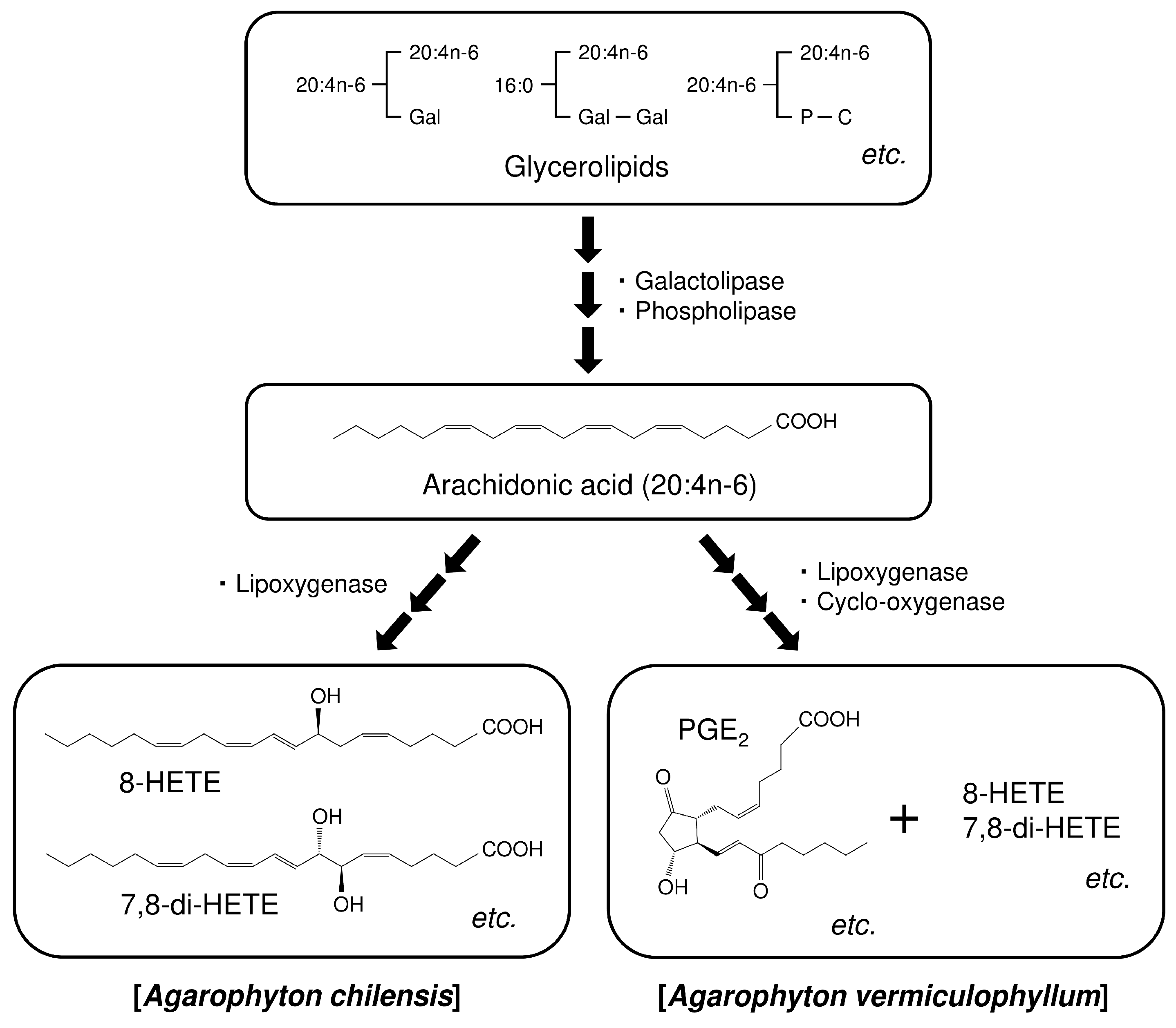
:1. Introduction
2. Results and Discussion
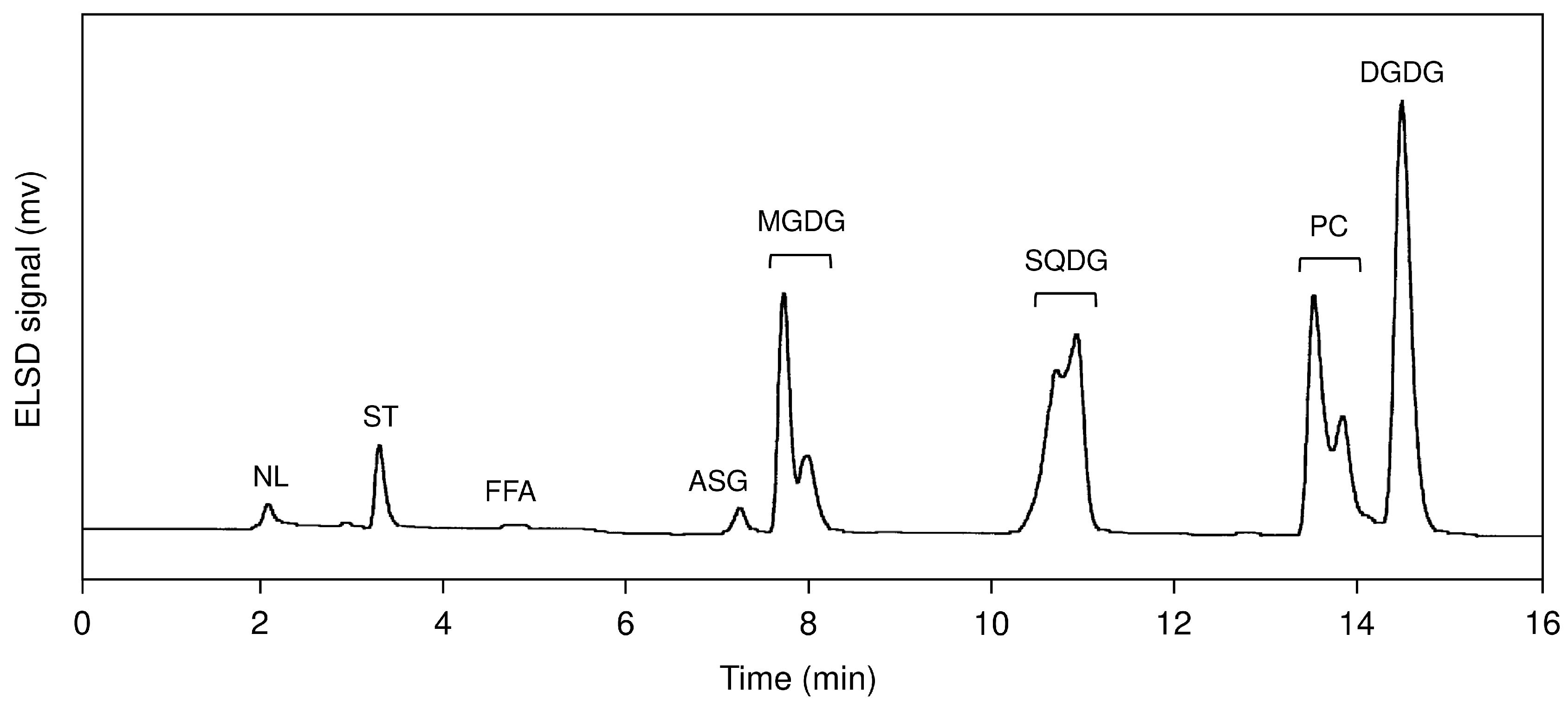
2.1. Lipid Class Composition
2.2. Fatty Acid Composition of Glycerolipids
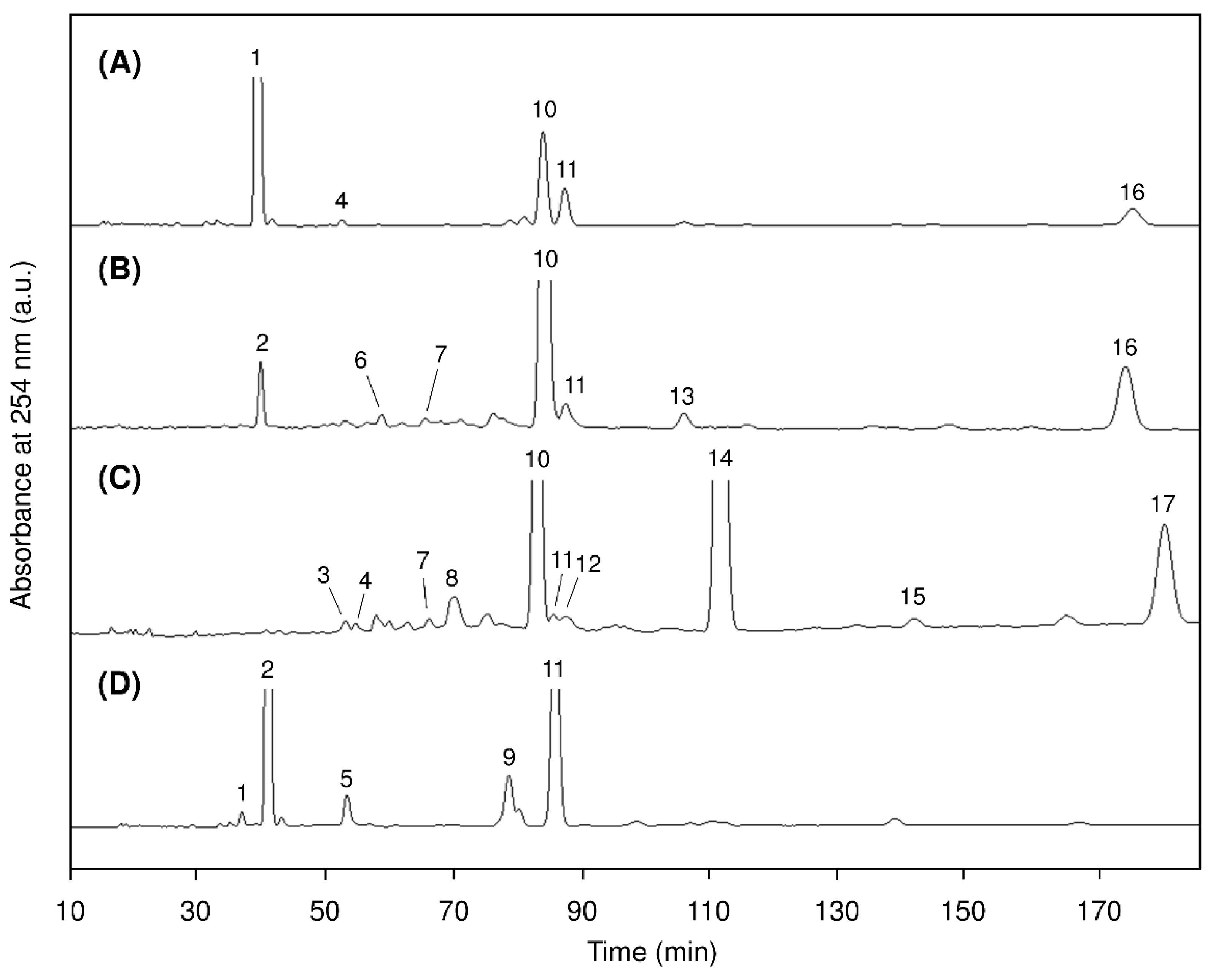
2.3. Positional Isomers of Glycerolipids
2.4. Molecular Species of Glycerolipids
3. Materials and Methods
3.1. Materials
3.2. Lipid Extraction
3.3. Lipid Class Analysis
3.4. Isolation of Glycerolipids
3.5. Fatty Acid Analysis
3.6. Release of DAG from Glycerolipids
3.7. Preparation of 3,5-Dinitrophenylurethane Derivatives
3.8. Chiral-Phase HPLC
3.9. Reversed-Phase HPLC
3.10. Reversed-Phase HPLC–ESI–MS
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gurgel, C.F.D.; Norris, J.N.; Schmidt, W.E.; Le, H.N.; Fredericq, S. Systematics of the Gracilariales (Rhodophyta) including new subfamilies, tribes, subgenera, and two new genera, Agarophyton gen. nov. and Crassa gen. nov. Phytotaxa 2018, 374, 1–23. [Google Scholar]
- Buschmann, A.H.; Briganti, F.; Retamales, C.A. Intertidal cultivation of Gracilaria chilensis (Rhodophyta) in southern Chile: Long term invertebrate abundance patterns. Aquaculture 1997, 156, 269–278. [Google Scholar] [CrossRef]
- Byrne, K.; Zuccarello, G.C.; West, J.; Liao, M.L.; Kraft, G.T. Gracilaria species (Gracilariaceae, Rhodophyta) from southeastern Australia, including a new species, Gracilaria perplexa sp. nov.: Morphology, molecular relationships and agar content. Phycol. Res. 2002, 50, 295–311. [Google Scholar] [CrossRef]
- Rempt, M.; Weinberger, F.; Grosser, K.; Pohnert, G. Conserved and species-specific oxylipin pathways in the wound-activated chemical defense of the noninvasive red alga Gracilaria chilensis and the invasive Gracilaria vermiculophylla. Beilstein J. Org. Chem. 2012, 8, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Lion, U.; Wiesemeier, T.; Weinberger, F.; Beltrán, J.; Flores, V.; Faugeron, S.; Correa, J.; Pohnert, G. Phospholipases and galactolipases trigger oxylipin-mediated wound-activated defence in the red alga Gracilaria chilensis against epiphytes. ChemBioChem 2006, 7, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N.; Hashimoto, K. Prostaglandin E2: A candidate for causative agent of “ogonori” poisoning. Nippon Suisan Gakkaishi 1984, 50, 465–469. [Google Scholar] [CrossRef]
- Nakajima, I.; Suzaki, K.; Oba, K. Production of prostaglandins by the marine alga, Gracilaria verrucosa. J. Jpn. Oil Chem. Soc. 1998, 47, 753–758. [Google Scholar] [CrossRef]
- Imbs, A.B.; Vologodskaya, A.V.; Nevshupova, N.V.; Khotimchenko, S.V.; Titlyanov, E.A. Response of prostaglandin content in the red alga Gracilaria verrucosa to season and solar irradiance. Phytochemistry 2001, 58, 1067–1072. [Google Scholar] [CrossRef]
- Kodama, S.; Nakajima, S.; Ozaki, H.; Takemoto, R.; Itabashi, Y.; Kuksis, A. Enantioseparation of hydroxyeicosatetraenoic acids by hydroxypropyl-γ-cyclodextrin-modified micellar electrokinetic chromatography. Electrophoresis 2016, 37, 3196–3205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacquemoud, D.; Pohnert, G. Extraction and analysis of oxylipins from macroalgae illustrated on the example Gracilaria vermiculophylla. In Natural Products from Marine Algae; Stengel, D.B., Connan, S., Eds.; Humana Press: New York, NY, USA, 2015; pp. 159–172. [Google Scholar]
- Illijas, M.I.; Terasaki, M.; Nakamura, R.; Iijima, N.; Hara, A.; Fusetani, N.; Itabashi, Y. Purification and characterization of glycerolipid acyl-hydrolase from the red alga Gracilaria vermiculophylla. Fish. Sci. 2008, 74, 670–676. [Google Scholar] [CrossRef]
- Nakajima, I.; Suzaki, K.; Oba, K. Mechanism for prostaglandin synthesis in the marine alga, Gracilaria verrucosa. J. Jpn. Oil Chem. Soc. 1998, 47, 759–763. [Google Scholar] [CrossRef]
- Kanamoto, H.; Takemura, M.; Ohyama, K. Identification of a cyclooxygenase gene from the red alga Gracilaria vermiculophylla and bioconversion of arachidonic acid to PGF2α in engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2011, 91, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Ishimaru, T.; Itabashi, Y. Lipid classes, fatty acid composition, and glycerolipid molecular species of the red alga Gracilaria vermiculophylla, a prostaglandin-producing seaweed. J. Oleo Sci. 2016, 65, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Araki, S.; Sakurai, T.; Oohusa, T.; Kayama, M.; Nisizawa, K. Content of arachidonic and eicosapentaenoic acids in polar lipids from Gracilaria (Gracilariales, Rhodophyta). Hydrobiologia 1990, 204/205, 513–519. [Google Scholar] [CrossRef]
- Khotimchenko, S.V. Lipids from the marine alga Gracilaria verrucosa. Chem. Nat. Compd. 2005, 41, 285–288. [Google Scholar] [CrossRef]
- Da Costa, E.; Melo, T.; Moreira, A.S.P.; Bernardo, C.; Helguero, L.; Ferreira, I.; Cruz, M.T.; Rego, A.M.; Domingues, P.; Calado, R.; et al. Valorization of lipids from Gracilaria sp. through lipidomics and decoding of antiproliferative and anti-Inflammatory activity. Mar. Drugs 2017, 15, 62. [Google Scholar] [CrossRef]
- Cleaver, G.; Bullock, S. Development in ELSD technology to improve sensitivity and linearity of response over a wider dynamic range. Chromatogr. Today 2015, 8, 38–41. [Google Scholar]
- Surget, G.; Le Lann, K.; Delebecq, G.; Kervarec, N.; Donval, A.; Poullaouec, M.A.; Bihannic, I.; Poupart, N.; Stiger-Pouvreau, V. Seasonal phenology and metabolomics of the introduced red macroalga Gracilaria vermiculophylla, monitored in the Bay of Brest (France). J. Appl. Phycol. 2017, 29, 2651–2666. [Google Scholar] [CrossRef]
- Kumari, P.; Kumar, M.; Gupta, V.; Reddy, C.R.K.; Jha, B. Tropical marine macroalgae as potential sources of nutritionally important PUFAs. Food Chem. 2010, 120, 749–757. [Google Scholar] [CrossRef]
- Imbs, A.B.; Latyshev, N.A.; Svetashev, V.I.; Skriptsova, A.V.; Le, T.T.; Pham, M.Q.; Pham, L.Q. Distribution of polyunsaturated fatty acids in red algae of the genus Gracilaria, a promising source of prostaglandins. Russ. J. Mar. Biol. 2012, 38, 339–345. [Google Scholar] [CrossRef]
- Norziah, M.H.; Ching, C.Y. Nutritional composition of edible seaweed Gracilaria changgi. Food Chem. 2000, 68, 69–76. [Google Scholar] [CrossRef]
- Bhaskar, N.; Kinami, T.; Miyashita, K.; Park, S.B.; Endo, Y.; Fujimoto, K. Occurrence of conjugated polyenoic fatty acids in seaweeds from the Indian Ocean. Z. Naturforsch. C Biosci. 2004, 59, 310–314. [Google Scholar] [CrossRef]
- Ohlrogge, J.; Browse, J. Lipid biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Honda, M.; Kashima, A.; Takahashi, K.; Itabashi, Y. Molecular species of digalactosyldiacylglycerols in the brown alga Sargassum yezoense. Nippon Suisan Gakkaishi 2009, 75, 1061–1069. [Google Scholar] [CrossRef]
- Itabashi, Y.; Kuksis, A.; Marai, L.; Takagi, T. HPLC resolution of diacylglycerol moieties of natural triacylglycerols on a chiral phase consisting of bonded (R)-(+)-1-(1-naphthyl) ethylamine. J. Lipid Res. 1990, 31, 1711–1717. [Google Scholar] [PubMed]
- Itabashi, Y. Chiral-phase HPLC of glycerolipids. In HPLC in Acyl Lipids; Lin, J.-T., McKeon, T.A., Eds.; HNB Publishing: New York, NY, USA, 2005; pp. 167–198. [Google Scholar]
- Itabashi, Y. Reverse isomers of glycoglycerolipids in marine red algae. In Proceedings of the 3rd Japan-Korea Joint Seminar on Fisheries Sciences, Jinju/Tongyeong, Korea, 15–16 December 2003; pp. 161–163. [Google Scholar]
- Cho, S.H.; Thompson, G.A. On the metabolic relationships between monogalactosyldiacylglycerol and digalactosyldiacylglycerol molecular species in Dunaliella salina. J. Biol. Chem. 1987, 262, 7586–7593. [Google Scholar] [PubMed]
- Ohta, H.; Awai, K.; Takamiya, K. Glyceroglycolipids of photosynthetic organisms-Their biosynthesis and evolutionary origin. Trends Glycosci. Glycotechnol. 2000, 12, 241–253. [Google Scholar] [CrossRef]
- Somerville, C.; Browse, J. Plant lipids: Metabolism, mutants, and membranes. Science 1991, 252, 80–87. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Sugawara, T.; Miyazawa, T. Separation and determination of glycolipids from edible plant sources by high-performance liquid chromatography and evaporative light-scattering detection. Lipids 1999, 34, 1231–1237. [Google Scholar] [CrossRef]
- Yunoki, K.; Sato, M.; Seki, K.; Ohkubo, T.; Tanaka, Y.; Ohnishi, M. Simultaneous quantification of plant glyceroglycolipids including sulfoquinovosyldiacylglycerol by HPLC-ELSD with binary gradient elution. Lipids 2009, 44, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Christie, W.W.; Han, X. Preparation of derivatives of fatty acids. In Lipid Analysis, 4th ed.; The Oily Press: Bridgwater, UK, 2010; pp. 145–158. [Google Scholar]
- Takagi, T.; Asahi, M.; Itabashi, Y. Fatty acid composition of twelve algae from Japanese waters. J. Jpn. Oil Chem. Soc. 1985, 34, 1008–1012. [Google Scholar] [CrossRef]
- Heinze, F.J.; Linscheid, M.; Heinz, E. Release of diacylglycerol moieties from various glycosyl diacylglycerols. Anal. Biochem. 1984, 139, 126–133. [Google Scholar] [CrossRef]
- Hashidate, T.; Itabashi, Y. Electrospray ionization mass spectrometry of regioisomeric 1,2-diacylglycerols. Bunseki Kagaku 2005, 54, 807–816. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | MGDG | DGDG | SQDG | PC | Total |
|---|---|---|---|---|---|
| A. chilensis | 66.5 ± 2.7 d | 142.2 ± 2.9 a | 130.8 ± 6.1 b | 104.5 ± 3.6 c | 444.0 |
| A. vermiculophyllum * | 88.1 ± 1.2 | 123.7 ± 2.9 | 107.8 ± 5.8 | 110.7 ± 8.9 | 430.3 |
| Fatty Acid | A.Chilensis | A. Vermiculophyllum * | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Total Lipid | Lipid Class | Total Lipid | Lipid Class | |||||||
| MGDG | DGDG | SQDG | PC | MGDG | DGDG | SQDG | PC | |||
| 12:0 | 0.3 | tr | tr | 0.2 | nd | 0.3 | 0.1 | 0.2 | 1.1 | tr |
| 14:0 | 3.6 | 0.4 | 0.8 | 10.9 | 0.9 | 5.3 | 1.7 | 1.8 | 14.7 | 1.2 |
| 15:0 | 0.2 | 0.1 | 0.3 | 0.5 | 0.1 | 0.6 | 1.1 | 0.8 | 1.0 | 0.2 |
| 16:0 | 27.4 | 12.5 | 43.8 | 52.6 | 15.2 | 31.6 | 21.7 | 54.0 | 61.7 | 15.3 |
| iso 17:0 | 0.2 | nd | nd | 0.2 | nd | 0.6 | nd | nd | nd | nd |
| 17:0 | tr | nd | nd | nd | 0.2 | 0.4 | 1.1 | 0.3 | 0.2 | 0.2 |
| 18:0 | 0.5 | 0.4 | 0.6 | 0.8 | 0.7 | 0.7 | 0.5 | 0.4 | 0.4 | 0.7 |
| 22:0 | tr | 0.1 | 0.2 | 0.2 | nd | 0.1 | nd | nd | nd | nd |
| Σ Saturated | 32.2 | 13.5 | 45.7 | 65.4 | 17.1 | 39.6 | 26.2 | 57.5 | 79.1 | 17.6 |
| 16:1n-9 | 0.2 | tr | tr | 0.2 | nd | 3.7 | 1.5 | 0.7 | 1.3 | tr |
| 16:1n-7 | 0.1 | nd | nd | 0.9 | 0.2 | tr | nd | nd | nd | 0.7 |
| 18:1n-9 | 4.3 | 3.9 | 6.6 | 0.1 | 2.4 | 3.3 | 3.2 | 3.8 | 0.8 | 2.1 |
| 18:1n-7 | 1.5 | 0.3 | 0.2 | tr | 3.0 | 1.7 | 0.5 | 0.2 | 0.3 | 3.2 |
| 20:1n-9 | 0.1 | 0.1 | tr | nd | 0.1 | 0.2 | 0.2 | tr | tr | nd |
| 24:1n-9 | 0.2 | 0.2 | 0.1 | 0.2 | nd | tr | nd | nd | nd | nd |
| Σ Monounsaturated | 6.4 | 4.5 | 6.9 | 1.4 | 5.7 | 8.9 | 5.3 | 4.7 | 2.4 | 6.0 |
| 16:3n-3 | nd | nd | nd | nd | nd | 0.2 | 0.9 | tr | nd | nd |
| 16:4n-3 | tr | nd | nd | nd | nd | 0.2 | 0.7 | nd | nd | tr |
| 18:2n-6 | 0.4 | 0.2 | 0.3 | 0.1 | 1.2 | 0.8 | 0.4 | 0.5 | 0.2 | 0.9 |
| 18:3n-6 | 0.1 | tr | tr | tr | 0.8 | 0.4 | 0.1 | tr | tr | 1.1 |
| 18:3n-3 | tr | nd | nd | nd | 0.3 | 0.3 | 0.4 | 0.2 | 0.1 | 0.1 |
| 18:4n-3 | nd | nd | nd | nd | nd | 0.4 | 1.0 | 0.2 | nd | 0.2 |
| 20:2n-6 | 0.2 | 0.1 | 0.2 | tr | 0.1 | 0.2 | 0.2 | 0.2 | nd | 0.2 |
| 20:3n-6 | 0.5 | 0.2 | 0.2 | tr | 1.0 | 2.0 | 0.8 | 0.6 | 0.3 | 5.5 |
| 20:4n-6 | 58.9 | 80.5 | 46.0 | 32.3 | 72.7 | 44.6 | 63.4 | 35.2 | 17.2 | 63.8 |
| 20:5n-3 | 0.4 | 0.6 | 0.2 | tr | 0.4 | 1.7 | 1.6 | 0.5 | tr | 2.3 |
| 22:4n-6 | tr | nd | nd | nd | 0.2 | 0.2 | tr | nd | nd | 0.5 |
| Σ Polyunsaturated | 60.5 | 81.6 | 46.9 | 32.4 | 76.7 | 51.0 | 68.1 | 37.4 | 17.8 | 74.6 |
| Others | 0.9 | 0.4 | 0.5 | 0.8 | 0.5 | 0.5 | 0.4 | 0.4 | 0.7 | 1.8 |
| Peak No * | ECN ** | Molecular Species | A.Chilensis | A. Vermiculophyllum *** | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (sn-1/sn-2) | MGDG | DGDG | SQDG | PC | MGDG | DGDG | SQDG | PC | ||
| – | 18 | 20:5n-3/16:4n-3 | nd | nd | nd | nd | 2.0 | nd | nd | nd |
| – | 20 | 18:4n-3/18:4n-3 | nd | nd | nd | nd | 1.0 | nd | nd | nd |
| – | 20 | 20:5n-3/16:3n-3 | nd | nd | nd | nd | 0.8 | nd | nd | nd |
| – | 20 | 20:5n-3/18:4n-3 | nd | nd | nd | nd | 0.9 | 1.1 | nd | nd |
| – | 22 | 16:2n-6/16:3n-3 | nd | nd | nd | nd | 0.8 | nd | nd | nd |
| – | 22 | 20:4n-6/20:5n-3 | nd | nd | nd | nd | 1.0 | nd | nd | nd |
| – | 22 | 20:5n-3/16:2n-6 | nd | nd | nd | nd | nd | 0.5 | nd | nd |
| – | 24 | 14:0/18:4n-3 | nd | nd | nd | nd | 0.5 | nd | nd | nd |
| – | 24 | 16:2n-6/16:2n-6 | nd | nd | nd | nd | 0.8 | nd | nd | nd |
| 1 | 24 | 20:4n-6/18:3n-6 | nd | nd | nd | 1.0 | nd | nd | nd | 1.2 |
| 2 | 24 | 20:4n-6/20:4n-6 | 63.8 | 4.1 | nd | 48.2 | 56.5 | 7.8 | 0.8 | 40.0 |
| – | 26 | 20:5n-3/20:2n-6 | nd | nd | nd | nd | nd | nd | nd | 0.9 |
| – | 26 | 18:2n-6/20:4n-6 | nd | nd | nd | nd | nd | 0.2 | nd | nd |
| 3 | 26 | 20:4n-6/14:0 | nd | nd | 0.4 | nd | nd | 0.4 | 0.3 | nd |
| 4 | 26 | 14:0/20:4n-6 | 0.8 | nd | 0.2 | nd | 1.8 | 1.7 | 0.2 | nd |
| 5 | 26 | 20:4n-6/20:3n-6 | nd | nd | nd | 3.2 | nd | nd | nd | 10.2 |
| 6 | 26 | 20:5n-3/16:0 | nd | 0.8 | nd | nd | nd | 0.5 | 0.2 | nd |
| 7 | 27 | 20:4n-6/15:0 | nd | 0.5 | 0.4 | nd | nd | 1.0 | 0.7 | nd |
| – | 27 | 15:0/20:4n-6 | nd | nd | nd | nd | nd | 0.3 | nd | nd |
| 8 | 28 | 12:0/16:0 | nd | nd | 2.7 | nd | nd | nd | 3.2 | nd |
| 9 | 28 | 18:1n-9/20:4n-6 | nd | nd | nd | 7.6 | nd | nd | nd | 7.7 |
| 10 | 28 | 20:4n-6/16:0 | 16.2 | 71.1 | 29.4 | nd | 17.7 | 75.4 | 58.4 | nd |
| 11 | 28 | 16:0/20:4n-6 | 6.7 | 1.8 | 0.3 | 31.3 | 7.3 | 4.3 | 0.2 | 26.3 |
| 12 | 29 | 15:0/14:0 | nd | nd | 0.4 | nd | nd | nd | 0.7 | nd |
| – | 30 | 16:1n-9/16:0 | nd | nd | nd | nd | nd | nd | 1.2 | nd |
| 13 | 30 | 14:0/18:1n-9 | nd | 1.7 | nd | nd | 0.3 | 0.8 | nd | nd |
| – | 30 | 16:0/20:3n-6 | nd | nd | nd | nd | nd | nd | nd | 3.8 |
| 14 | 30 | 14:0/16:0 | nd | nd | 49.3 | nd | nd | nd | 29.4 | nd |
| – | 30 | 16:0/20:3n-6 | nd | nd | nd | nd | 0.3 | 0.6 | nd | nd |
| 15 | 31 | 15:0/16:0 | nd | nd | 0.3 | nd | nd | nd | nd | nd |
| 16 | 32 | 16:0/18:1n-9 | 5.6 | 13.4 | nd | nd | 2.5 | 3.5 | nd | nd |
| 17 | 32 | 16:0/16:0 | nd | nd | 11.7 | nd | nd | nd | 3.6 | nd |
| Others | 6.9 | 6.6 | 4.9 | 8.7 | 5.8 | 1.9 | 1.1 | 9.9 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Honda, M.; Ishimaru, T.; Itabashi, Y.; Vyssotski, M. Glycerolipid Composition of the Red Macroalga Agarophyton Chilensis and Comparison to the Closely Related Agarophyton Vermiculophyllum Producing Different Types of Eicosanoids. Mar. Drugs 2019, 17, 96. https://0-doi-org.brum.beds.ac.uk/10.3390/md17020096
Honda M, Ishimaru T, Itabashi Y, Vyssotski M. Glycerolipid Composition of the Red Macroalga Agarophyton Chilensis and Comparison to the Closely Related Agarophyton Vermiculophyllum Producing Different Types of Eicosanoids. Marine Drugs. 2019; 17(2):96. https://0-doi-org.brum.beds.ac.uk/10.3390/md17020096
Chicago/Turabian StyleHonda, Masaki, Takashi Ishimaru, Yutaka Itabashi, and Mikhail Vyssotski. 2019. "Glycerolipid Composition of the Red Macroalga Agarophyton Chilensis and Comparison to the Closely Related Agarophyton Vermiculophyllum Producing Different Types of Eicosanoids" Marine Drugs 17, no. 2: 96. https://0-doi-org.brum.beds.ac.uk/10.3390/md17020096