Characterization of a New Intracellular Alginate Lyase with Metal Ions-Tolerant and pH-Stable Properties

 and
and
Abstract
:1. Introduction
2. Results and Discussion
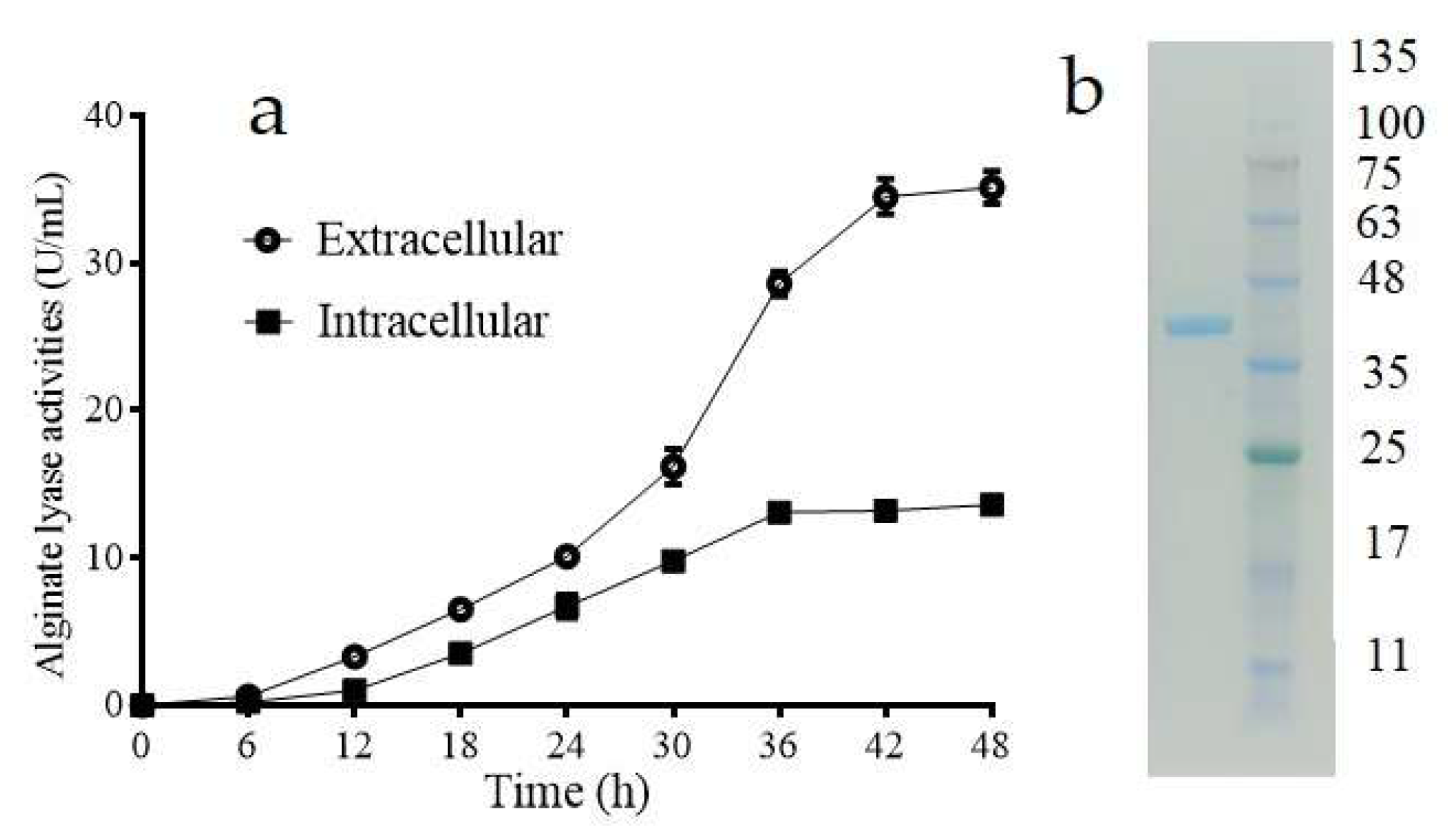
2.1. Purification of the Intracellular Alginate Lyase in Vibrio sp. W2
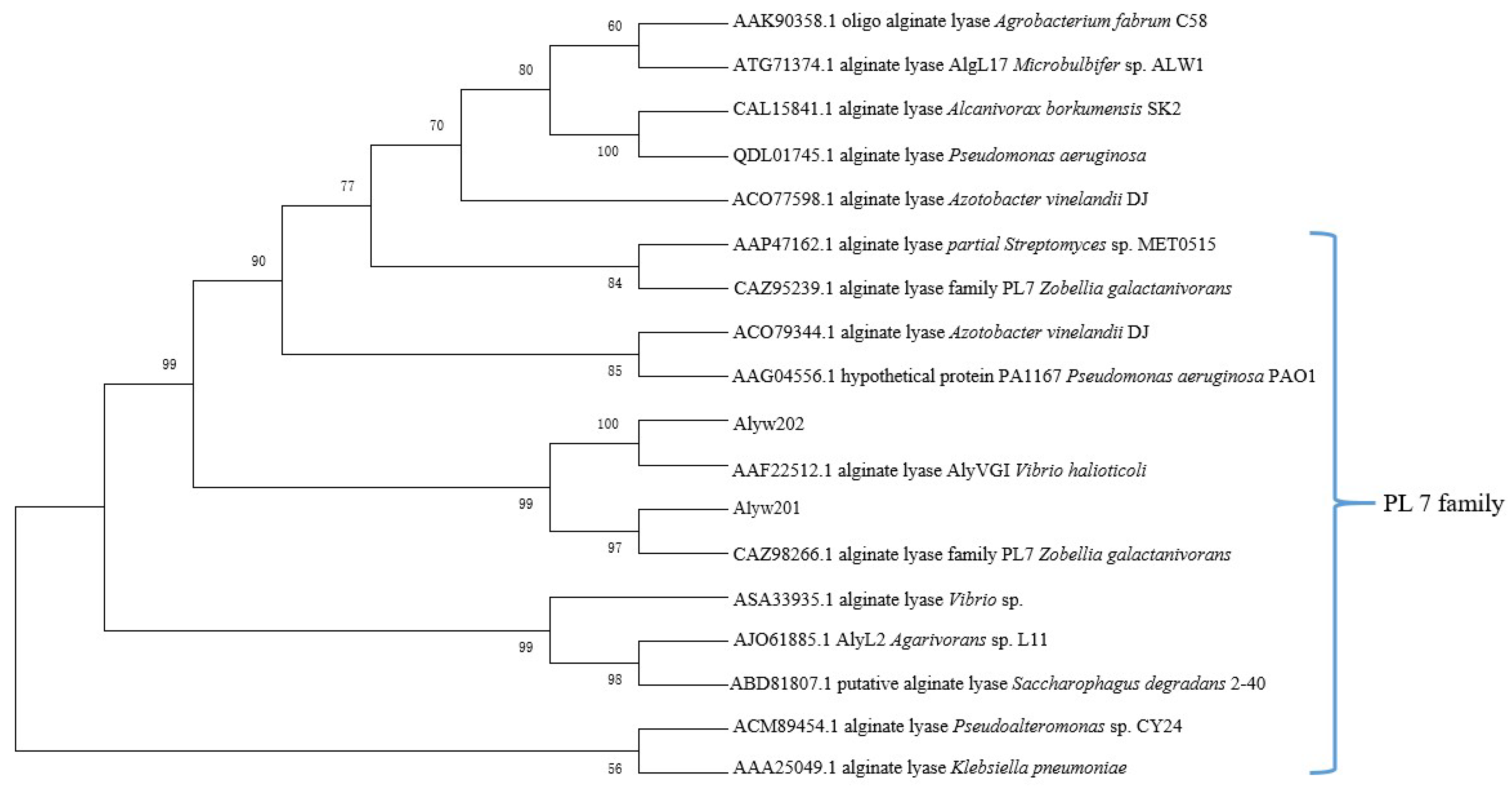
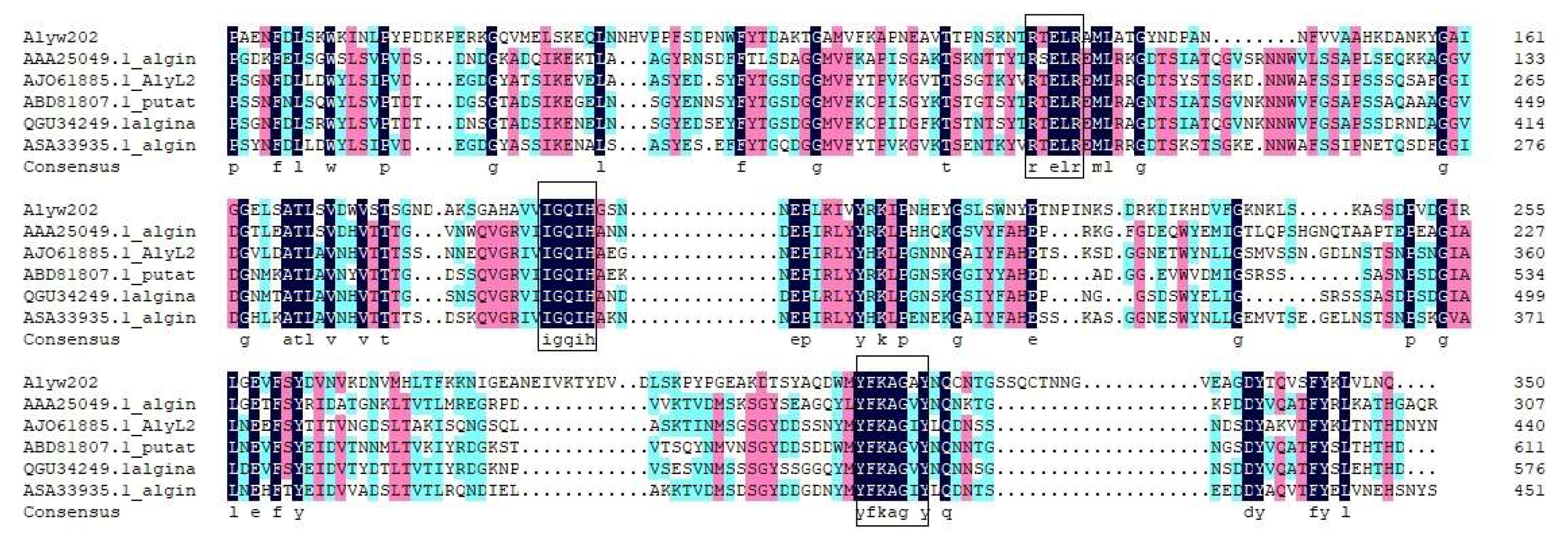
2.2. Bioinformatics Analysis of the Alginate Lyase Alyw202
2.3. Expression of Alyw202
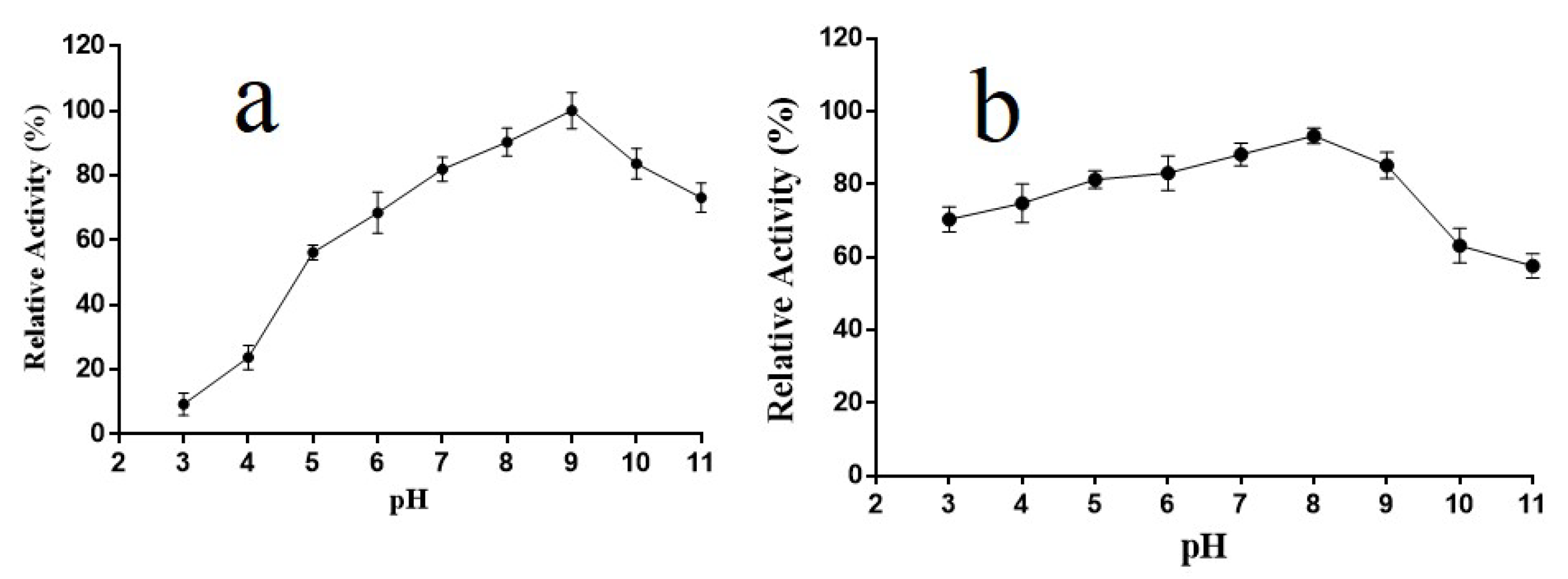
2.4. pH Properties of Alyw202
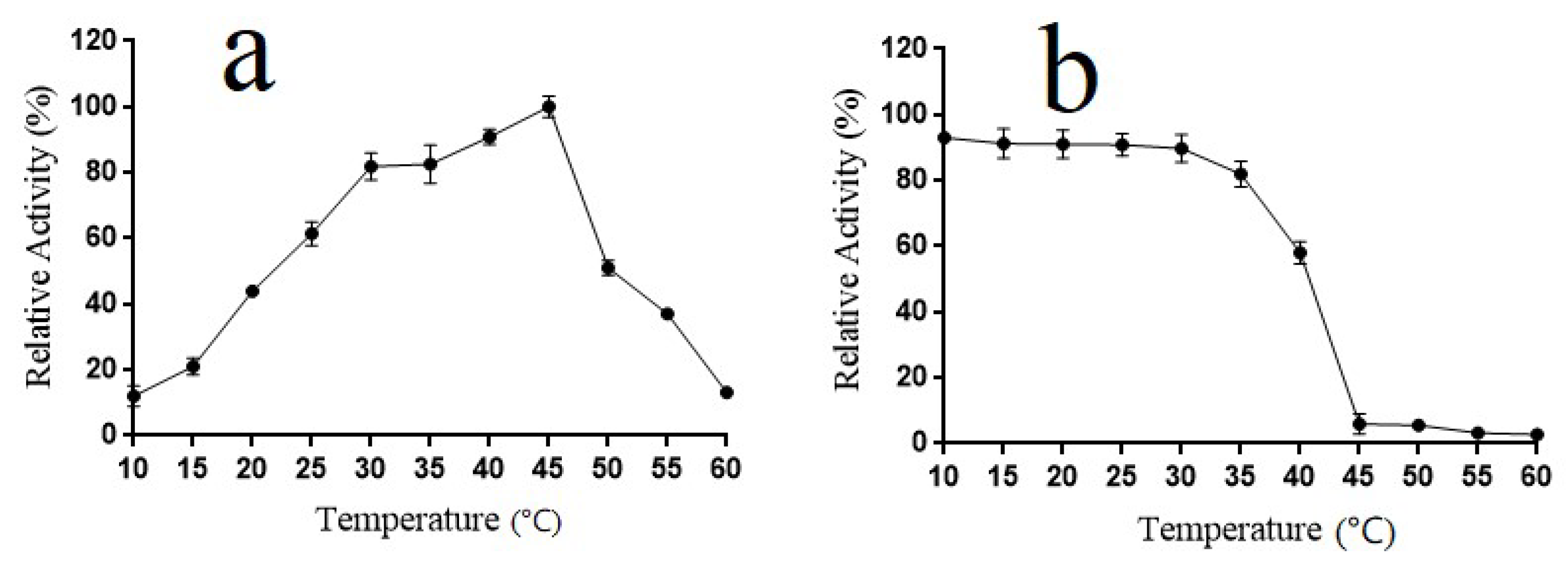
2.5. Temperature Properties of Alyw202
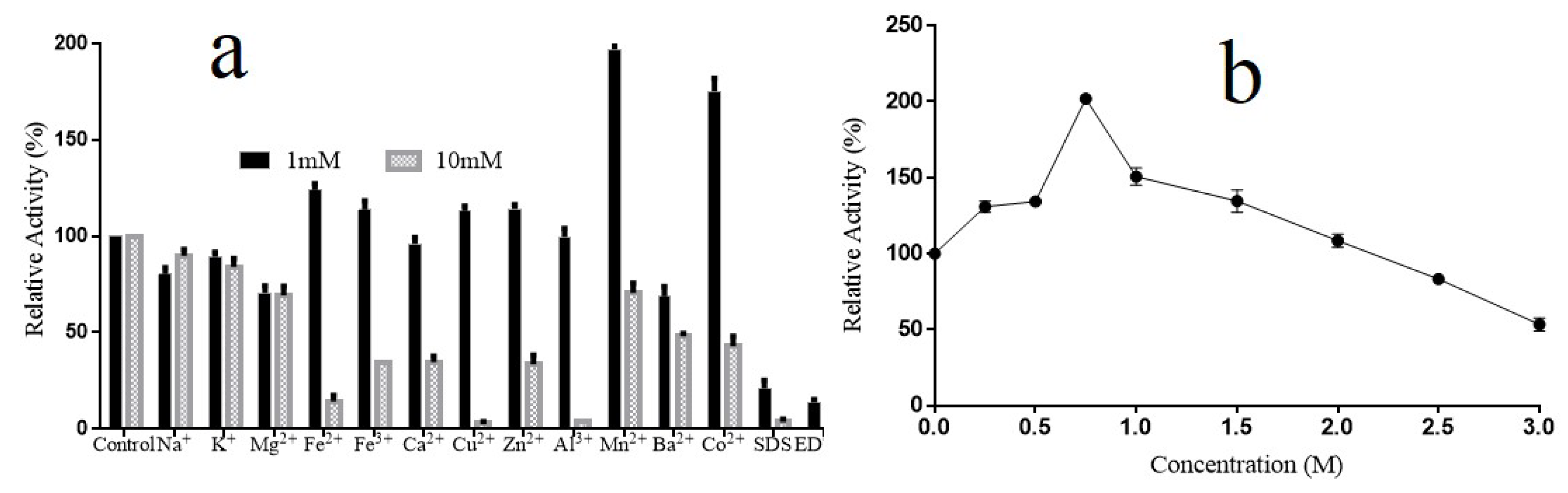
2.6. Effects of Ions on the Activity of Alyw202
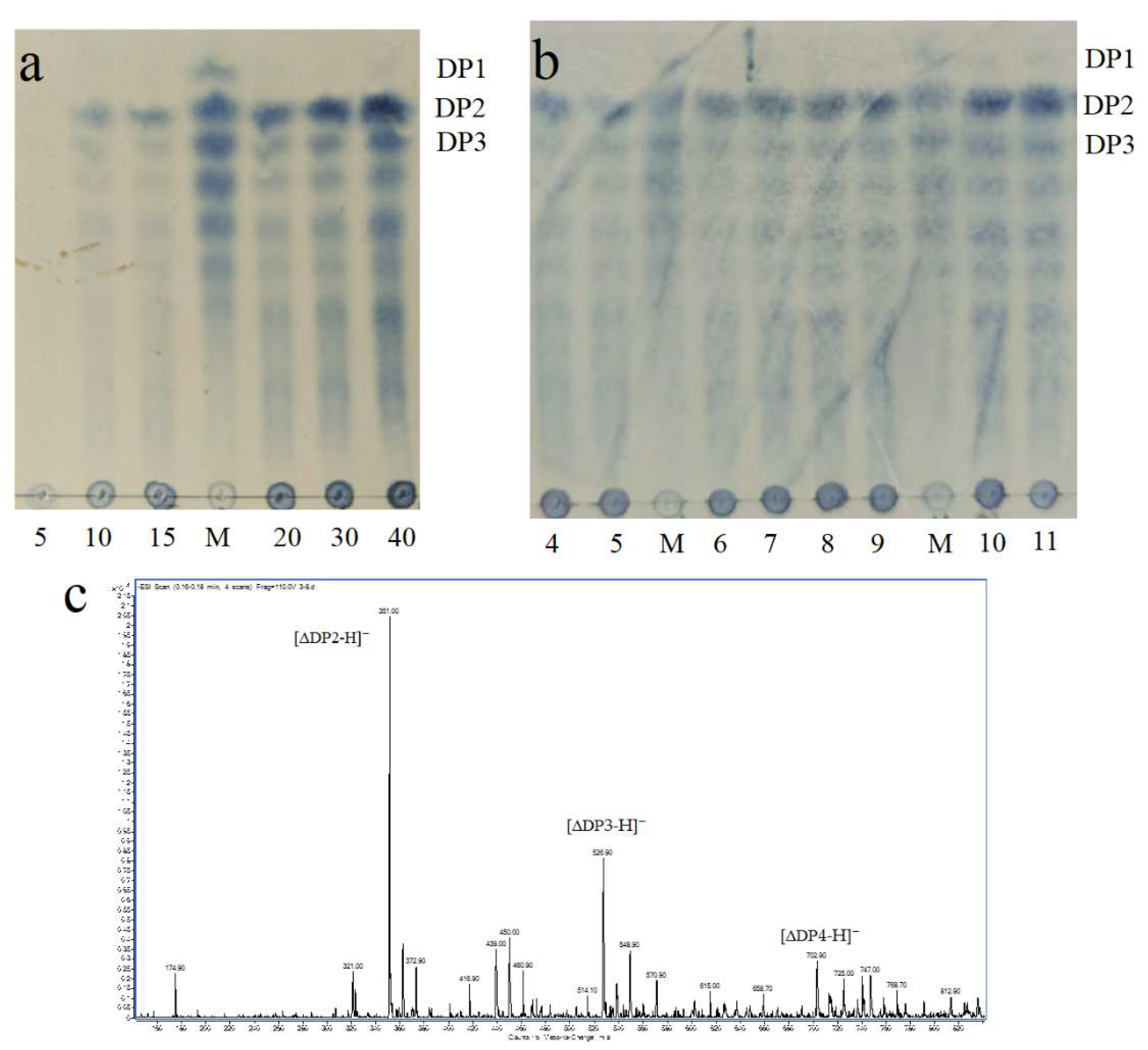
2.7. ESI-MS Analysis of Degradation Products
3. Materials and Methods
3.1. Materials, Strains and Mediums
3.2. Purification of Intracellular Alginate Lyase
3.3. Enzyme Activity Assay
3.4. Bioinformatics Analysis of Alyw202
3.5. Secretory Expression and Purification of Alyw202
3.6. Effects of pH and Temperature on Alyw202 Activity and Stability
3.7. Effects of Metal Ions, NaCl and Chemical Compounds on Alyw201 Activity
3.8. Analysis of Alyw202 Reaction Products
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gacesa, P. Enzymic degradation of alginates. Int. J. Biochem. 1992, 24, 545–552. [Google Scholar] [CrossRef]
- Inoue, A. Characterization of PL-7 Family Alginate Lyases from Marine Organisms. Methods Enzymol. 2018, 605, 499–524. [Google Scholar] [PubMed]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertesvåg, H. Alginate-modifying enzymes: Biological roles and biotechnological uses. Front. Microbiol. 2015, 6, 523. [Google Scholar]
- Tusi, S.K.; Khalaj, L.; Ashabi, G.; Kiaei, M.; Khodagholi, F. Alginate oligosaccharide protects against endoplasmic reticulum- and mitochondrial-mediated apoptotic cell death and oxidative stress. Biomaterials 2011, 32, 5438–5458. [Google Scholar] [CrossRef]
- Liu, S.; Liu, G.; Yi, Y. Novel vanadyl complexes of alginate saccharides: Synthesis, characterization, and biological activities. Carbohydr. Polym. 2015, 121, 86–91. [Google Scholar] [CrossRef]
- Uno, T.; Hattori, M.T.; Yoshida, T. Oral administration of alginic acid oligosaccharide suppresses IgE production and inhibits the induction of oral tolerance. Biosci. Biotechnol. Biochem. 2006, 70, 3054–3057. [Google Scholar] [CrossRef]
- Xu, X.; Iwamoto, Y.; Kitamura, Y.; Oda, T.; Muramatsu, T. Root growth-promoting activity of unsaturated oligomeric uronates from alginate on carrot and rice plants. Biosci. Biotechnol. Biochem. 2003, 67, 2022–2025. [Google Scholar] [CrossRef] [Green Version]
- Schiller, N.L.; Monday, S.R.; Boyd, C.M.; Keen, N.T.; Ohman, D.E. Characterization of the Pseudomonas aeruginosa alginate lyase gene (algL): Cloning, sequencing, and expression in Escherichia coli. J. Bacteriol. 1993, 175, 4780–4789. [Google Scholar] [CrossRef] [Green Version]
- Thomas, F.; Lundqvist, L.C.; Jam, M.; Jeudy, A.; Barbeyron, T.; Sandström, C.; Michel, G.; Czjzek, M. Comparative characterization of two marine alginate lyases from Zobellia galactanivorans reveals distinct modes of action and exquisite adaptation to their natural substrate. J. Biol. Chem. 2013, 288, 23021–23037. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Nishiyama, R.; Ojima, T. The alginate lyases FlAlyA, FlAlyB, FlAlyC, and FlAlex from Flavobacterium sp. UMI-01 have distinct roles in the complete degradation of alginate. Algal Res. 2016, 19, 355–362. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Hehemann, J.H.; Polz, M.F.; Lee, J.K.; Zhao, H. Comparative biochemical characterization of three exolytic oligoalginate lyases from Vibrio splendidus reveals complementary substrate scope, temperature, and pH adaptations. Appl. Environ. Microbiol. 2014, 80, 4207–4214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.P.; Cao, M.; Li, B.; Ji, X.F.; Zhang, X.Y.; Zhang, Y.Q.; Wang, H.Y. Cloning, Secretory Expression and Characterization of a Unique pH-Stable and Cold-Adapted Alginate Lyase. Mar. Drugs 2020, 18, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Zhu, Y.; Men, Y.; Zeng, Y.; Sun, Y. Purification and Characterization of a Novel Alginate Lyase from the Marine Bacterium Bacillus sp. Alg07. Mar. Drugs 2018, 16, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, X.; Bi, X.; Ren, Y.; Han, Q.; Zhou, Y.; Han, Y.; Yao, R.; Li, S. Characterization of an Alkaline Alginate Lyase with pH-Stable and Thermo-Tolerance Property. Mar. Drugs 2019, 17, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterner, M.; Edlund, U. Multicomponent Fractionation Of Saccharina Latissimabrown Algae Using Chelating Salt Solutions. J. Appl. Phycol. 2016, 28, 2561–2574. [Google Scholar] [CrossRef] [Green Version]
- Ravanal, M.C.; Pezoa-Conte, R.; von Schoultz, S.; Hemming, J.; Salazar, O.; Anugwom, I.; Jogunola, O.; Mäki-Arvela, P.; Willför, S.; Mikkola, J.P.; et al. Comparison of Different Types of Pretreatment and Enzymatic Saccharification of Macrocystis Pyrifera for the Production of Biofuel. Algal Res. 2016, 13, 141–147. [Google Scholar] [CrossRef]
- Zhuang, J.; Zhang, K.; Liu, X.; Liu, W.; Lyu, Q.; Ji, A. Characterization of a novel polyM-preferred alginate lyase from Marine Vibrio splendidus OU02. Mar. Drugs 2018, 16, 295. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Guo, E.W.; Yu, W.G.; Han, F. Purification and characterization of a new alginate lyase from a marine bacterium Vibrio sp. Biotechnol. Lett. 2013, 35, 703–708. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Shao, Y.; Jiao, C.; Yang, Q.M.; Weng, H.F.; Xiao, A.F. Characterization and Application of an Alginate Lyase, Aly1281 from Marine Bacterium Pseudoalteromonas carrageenovora ASY5. Mar. Drugs 2020, 18, 95. [Google Scholar] [CrossRef] [Green Version]
- Sugimura, I.; Sawabe, T.; Ezura, Y. Cloning and sequence analysis of Vibrio halioticoli genes encoding three types of polyguluronate lyase. Mar. Biotechnol. 2000, 2, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, X.; Bao, M.; Wu, Y.; Yu, W.; Han, F. Family 13 carbohydrate-binding module of alginate lyase from Agarivorans sp. L11 enhances its catalytic efficiency and thermostability, and alters its substrate preference and product distribution. FEMS Microbiol. Lett. 2015, 362, fnv054. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.T.; Ko, H.J.; Kim, N.; Kim, D.; Lee, D.; Choi, I.G.; Woo, H.C.; Kim, M.D.; Kim, K.H. Characterization of a recombinant endo-type alginate lyase (Alg7D) from Saccharophagus degradans. Biotechnol. Lett. 2012, 34, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Chen, P.; Zeng, Y.; Men, Y.; Mu, S.; Zhu, Y.; Chen, Y.; Sun, Y. The Characterization and Modification of a Novel Bifunctional and Robust Alginate Lyase Derived from Marinimicrobium Sp. H1. Mar. Drugs 2019, 17, 545. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Sun, Y.; Ni, F.; Ning, L.; Yao, Z. Characterization of a new endo-type alginate lyase from Vibrio sp. NJU-03. Int. J. Biol. Macromol. 2018, 108, 1140–1147. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, Y.; Kawada, R.; Iwasaki, K.I.; Kimura, Y.; Oda, T.; Muramatsu, T. Cloning and sequence analysis of a gene (aly PG) encoding poly(alpha-L-guluronate) lyase from Corynebacterium sp. strain ALY-1. J. Biosci. Bioeng. 2000, 89, 199. [Google Scholar] [CrossRef]
- Yoon, H.J.; Hashimoto, W.; Miyake, O.; Okamoto, M.; Mikami, B.; Murata, K. Overexpression in Escherichia coli, purification, and characterization of Sphingomonas sp. A1 alginate lyases. Protein Expr. Purif. 2000, 19, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Madzak, C. Yarrowia lipolytica: Recent achievements in heterologous protein expression and pathway engineering. Appl. Microbiol. Biotechnol. 2015, 99, 4559–4577. [Google Scholar] [CrossRef]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef]
- Zhu, B.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Li, J.W.; Dong, S.; Song, J.; Li, C.B.; Chen, X.L.; Xie, B.B.; Zhang, Y.Z. Purification and characterization of a bifunctional alginate lyase from Pseudoalteromonas sp. SM0524. Mar. Drugs 2011, 9, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Dou, W.F.; Wei, D.; Li, H.; Li, H.; Rahman, M.M.; Shi, J.S.; Xu, Z.H.; Ma, Y.H. Purification and characterisation of a bifunctional alginate lyase from novel Isoptericola halotolerans CGMCC 5336. Carbohydr. Polym. 2013, 98, 1476–1482. [Google Scholar] [CrossRef]
- Zhu, B.; Ni, F.; Ning, L.; Sun, Y.; Yao, Z. Cloning and characterization of a new pH-stable alginate lyase with high salt tolerance from marine Vibrio sp. NJ-04. Int. J. Biol. Macromol. 2018, 115, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Jiang, X.; Hwang, H.M. Purification and characterization of an alginate lyase from marine bacterium Vibrio sp. Mutant Strain 510-64. Curr. Microbiol. 2006, 53, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhang, Z.; Li, S.; Su, H.; Tang, L.; Tan, Y.; Yu, W.; Han, F. Characterization of a new endo-type polysaccharide lyase (PL) family 6 alginate lyase with cold-adapted and metal ions-resisted property. Int. J. Biol. Macromol. 2018, 120, 729–735. [Google Scholar] [CrossRef]
- Zhang, Z.; Tang, L.; Bao, M.; Liu, Z.; Yu, W.; Han, F. Functional Characterization of Carbohydrate-Binding Modules in a New Alginate Lyase, TsAly7B, From Thalassomonas Sp. LD5. Mar. Drugs 2019, 18, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Tan, H.; Qin, Y.; Xu, Q.; Du, Y.; Yin, H. Characterization of a new endo-type alginate lyase from Vibrio sp. W13. Int. J. Biol. Macromol. 2015, 75, 330–337. [Google Scholar] [CrossRef]
- Xu, F.; Dong, F.; Wang, P.; Cao, H.Y.; Li, C.Y.; Li, P.Y.; Pang, X.H.; Zhang, Y.Z.; Chen, X.L. Novel molecular insights into the catalytic mechanism of marine bacterial alginate lyase AlyGC from polysaccharide lyase family 6. J. Biol. Chem. 2017, 292, 4457–4468. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.L.; Sheng, D.; Fei, X.; Fang, D.; Li, P.Y.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Xie, B.B. Characterization of a new cold-adapted and salt-activated polysaccharide lyase family 7 alginate lyase from Pseudoalteromonas sp. SM0524. Front. Microbiol. 2016, 7, 1120. [Google Scholar] [CrossRef]
- Li, H.; Wang, S.; Zhang, Y.; Chen, L. High-level expression of a thermally stable alginate lyase using pichia pastoris, characterization and application in producing brown alginate oligosaccharide. Mar. Drugs 2018, 16, 158. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wang, Z.P.; Sheng, J.; Zheng, Y.; Ji, X.F.; Zhou, H.X.; Liu, X.Y.; Chi, Z.M. High and efficient isomaltulose production using an engineered Yarrowia lipolytica strain. Bioresour. Technol. 2018, 265, 577–580. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Purification Fold | Yield (%) |
|---|---|---|---|---|---|
| Crude enzyme | 6802.3 ± 0.5 | 86.4 ± 0.1 | 78.7 ± 0.6 | 1 | 100 |
| DEAE-Fast Flow | 5641.7 ± 0.6 | 12.3 ± 0.1 | 458.7 ± 0.4 | 5.8 | 82.9 |
| Sephadex G-75 | 4611.3 ± 0.7 | 2.4 ± 0.1 | 1926.4 ± 0.4 | 24.5 | 67.8 |
| Name | Source | Molecular Weights (kDa) | Specific Activity | pH-Stable Range |
|---|---|---|---|---|
| Alyw202 | This study | 38.3 | 1926.4 U/mg | 3.0–10.0 |
| Alyw201 | Vibrio sp. W2 [13] | 38.0 | 876.4 U/mg | 3.0–10.0 |
| TsAly6A | Thalassomonas sp. [34] | 83.9 | 15,960 U/μmol | 6.6–8.95 |
| TsAly7B | Thalassomonas sp. [35] | 65 | 488.8 U/mg | 7.3–8.6 |
| ZH0-IV | Sphingomonas sp. [27] | 113 | 12.3 U/mg | 6.0–9.0 |
| Algb | Vibrio sp. W13 [36] | 55.05 | 457 U/mg | 4.0–10.0 |
| AlgNJ-04 | Vibrio sp. NJU-04 [32] | 50 | 2416 U/mg | 4.0–10.0 |
| rSAGL | Flavobacterium sp. [37] | 33 | 4044 U/mg | - |
| AlgNJU-03 | Vibrio sp. NJU-03 [25] | 55.05 | 457 U/mg | 6.0–9.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Li, J.; Zhang, X.-Y.; Ni, H.-D.; Wang, F.-B.; Wang, H.-Y.; Wang, Z.-P. Characterization of a New Intracellular Alginate Lyase with Metal Ions-Tolerant and pH-Stable Properties. Mar. Drugs 2020, 18, 416. https://0-doi-org.brum.beds.ac.uk/10.3390/md18080416
Ma Y, Li J, Zhang X-Y, Ni H-D, Wang F-B, Wang H-Y, Wang Z-P. Characterization of a New Intracellular Alginate Lyase with Metal Ions-Tolerant and pH-Stable Properties. Marine Drugs. 2020; 18(8):416. https://0-doi-org.brum.beds.ac.uk/10.3390/md18080416
Chicago/Turabian StyleMa, Yan, Jie Li, Xin-Yue Zhang, Hao-Dong Ni, Feng-Biao Wang, Hai-Ying Wang, and Zhi-Peng Wang. 2020. "Characterization of a New Intracellular Alginate Lyase with Metal Ions-Tolerant and pH-Stable Properties" Marine Drugs 18, no. 8: 416. https://0-doi-org.brum.beds.ac.uk/10.3390/md18080416