
Relationship between the Chromosome Structural Dynamics and Gene Expression—A Chicken and Egg Dilemma?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
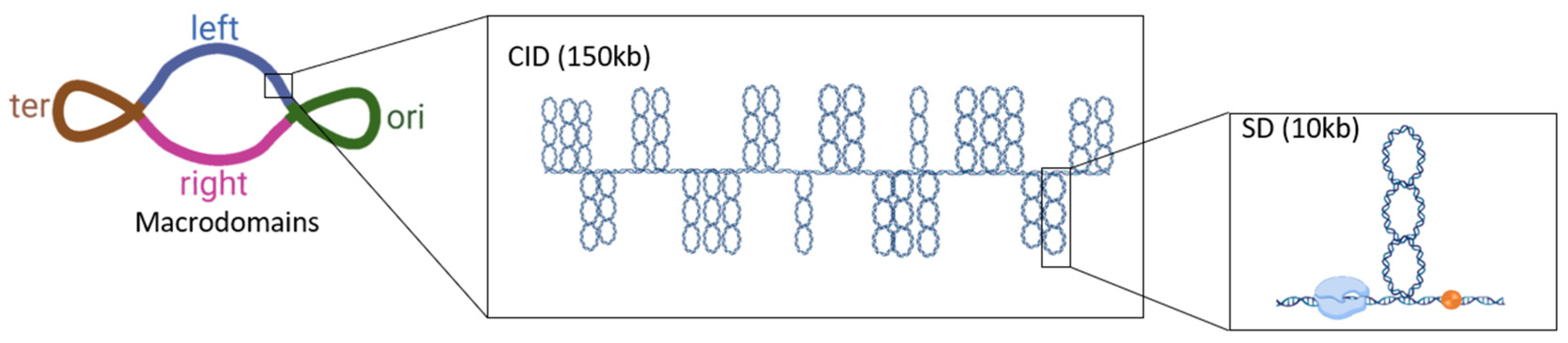{kind=link}
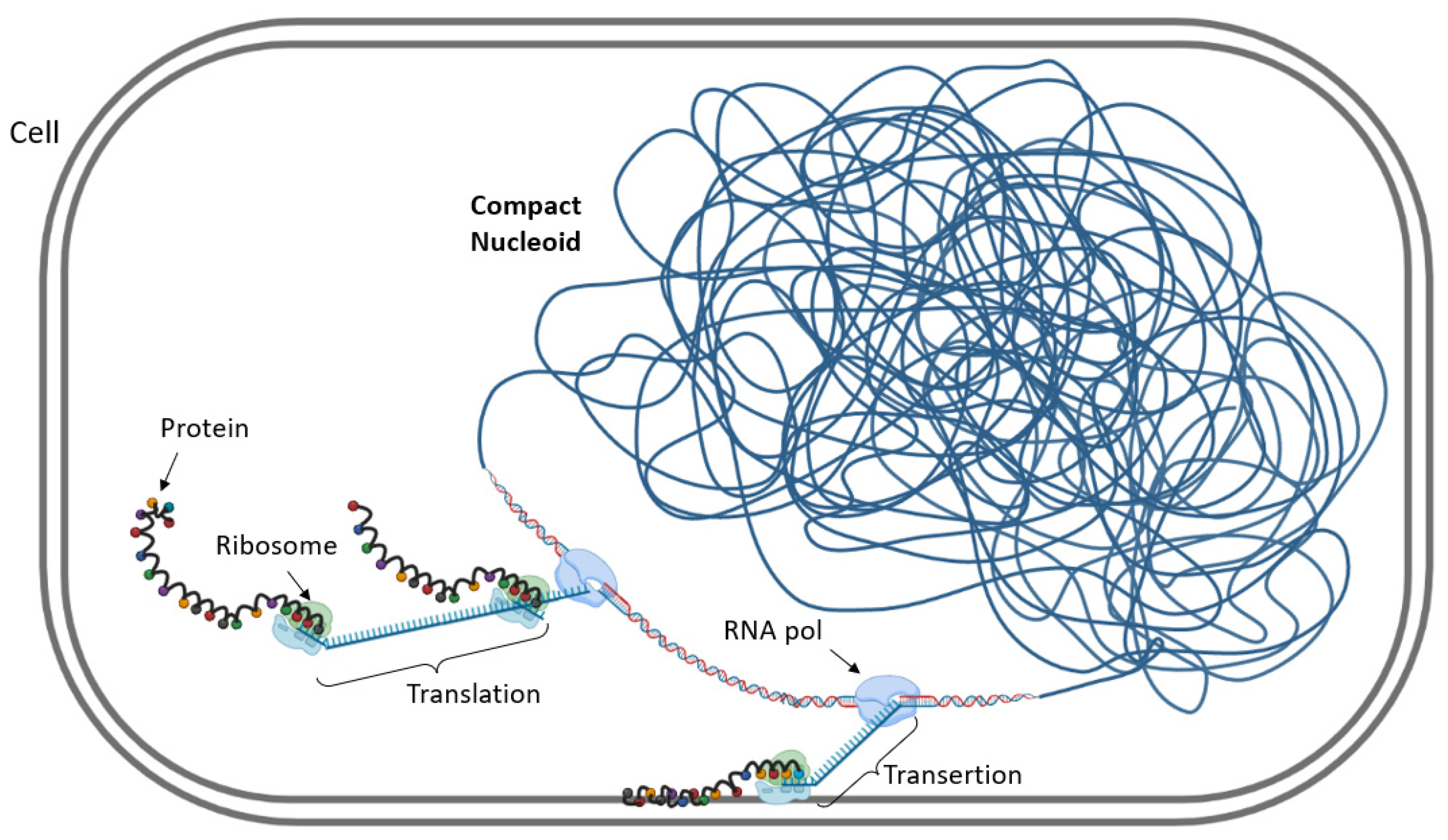{kind=link}
Abstract
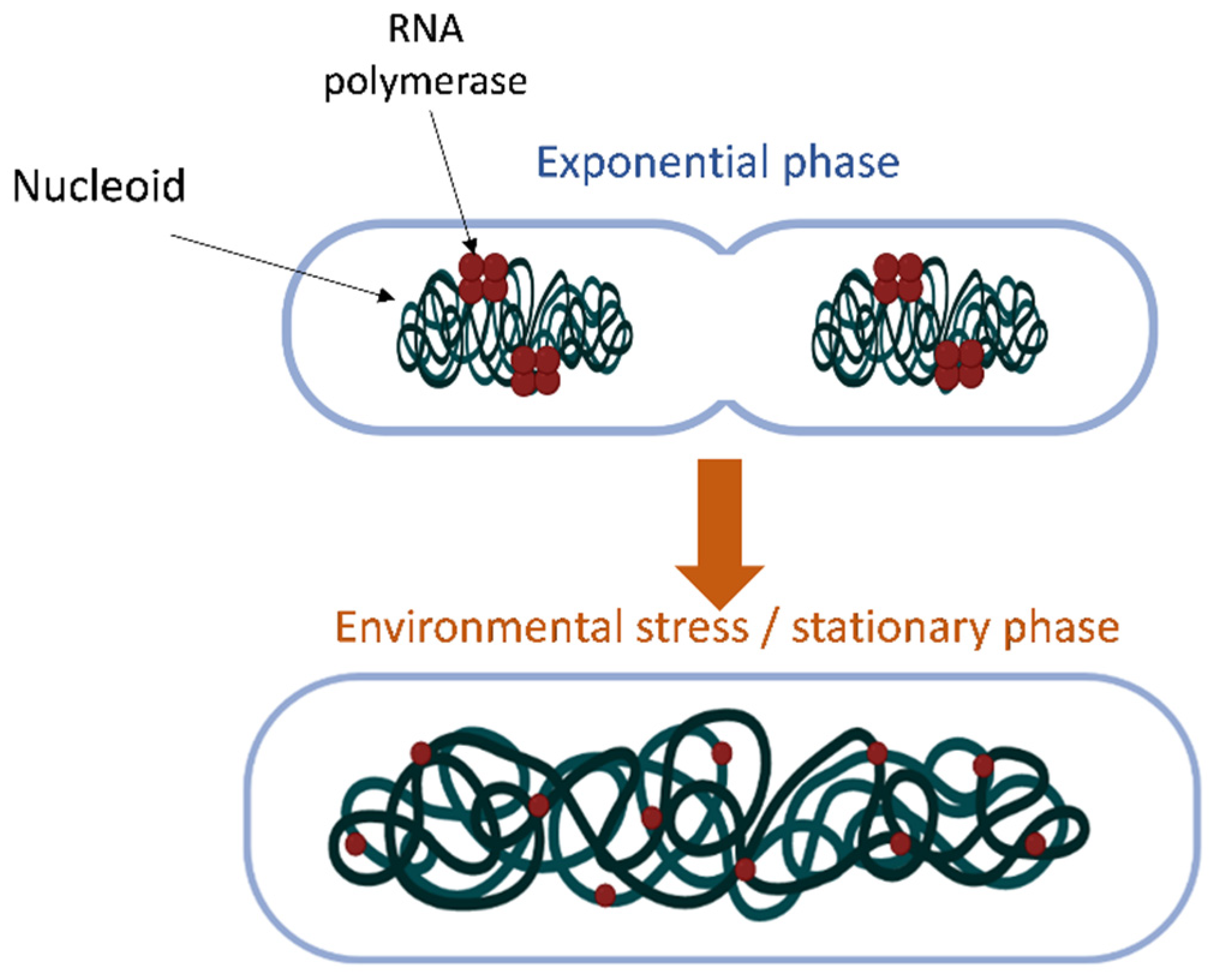
:1. Introduction
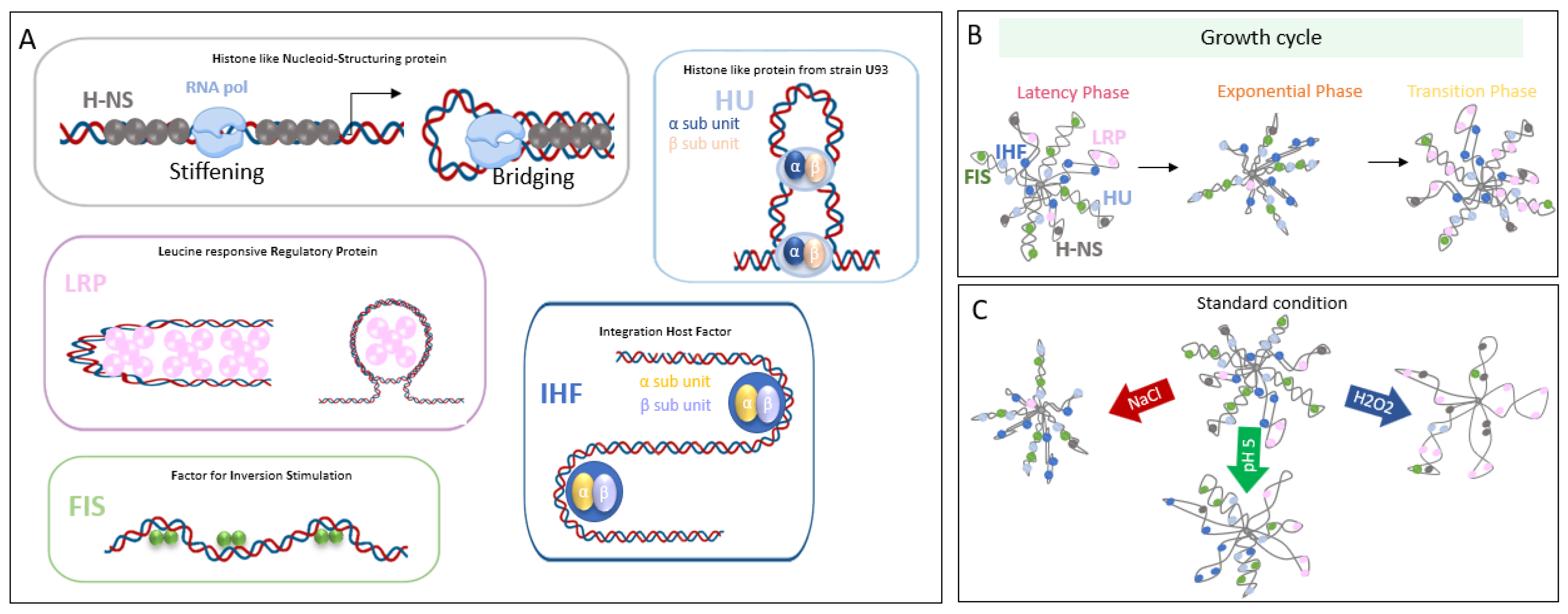
2. NAPs Are Versatile and Flexible Actors in Coupling Nucleoid Organization and Transcription Request
2.1. H-NS
2.2. FIS
2.3. IHF
2.4. HU
2.5. LRP
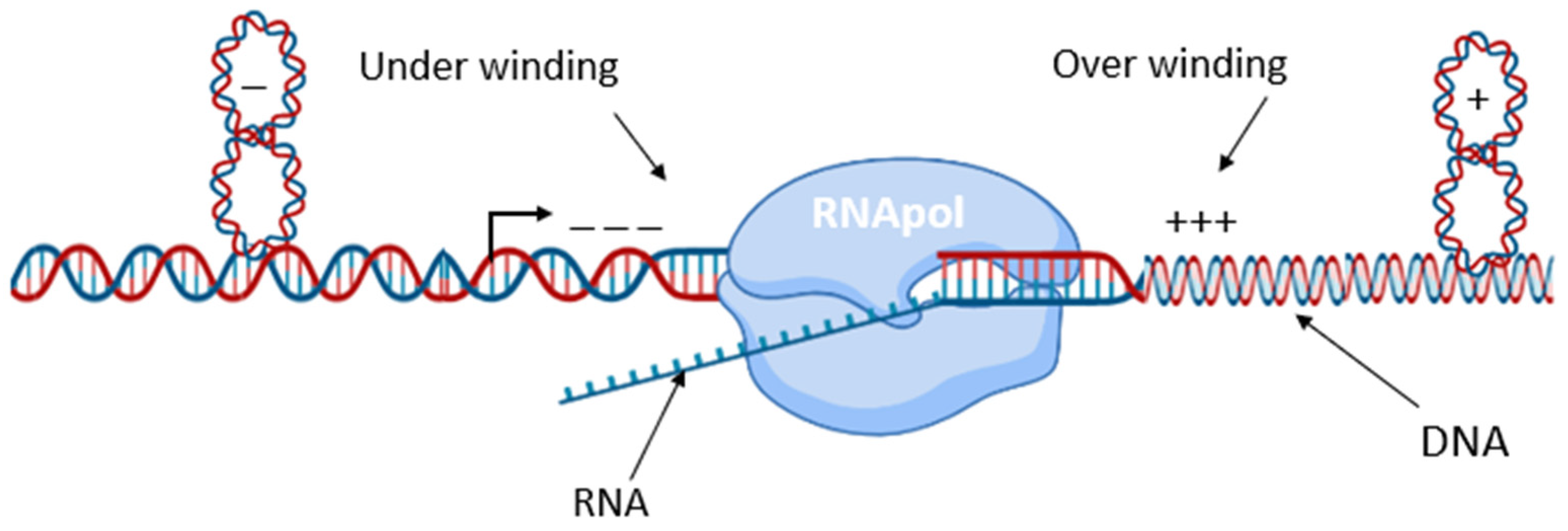
3. Transcription, a Global Regulator of DNA Supercoiling State and Local Organization of the Nucleoid
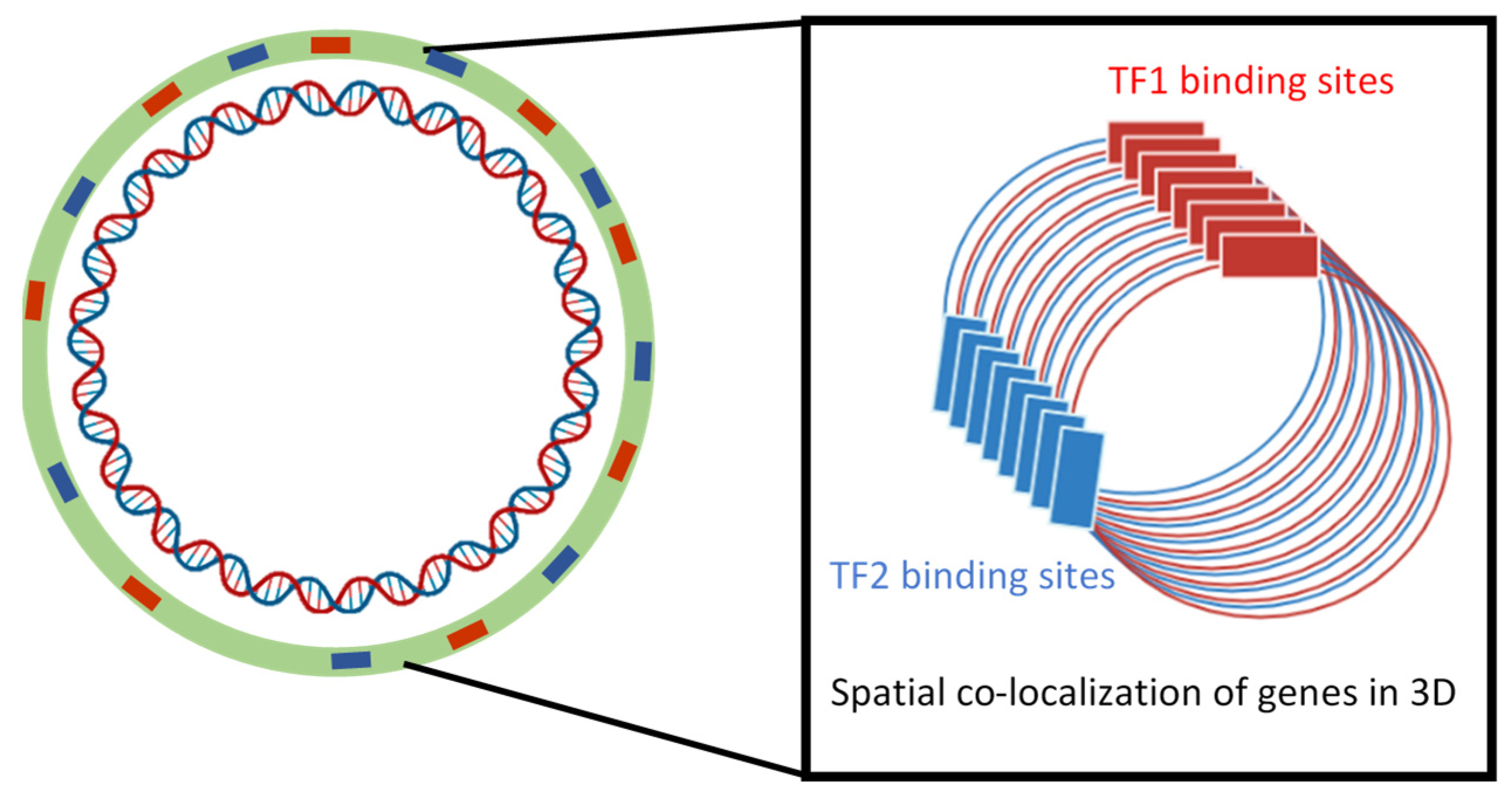
4. Chromosomal Organization of an “Archipelago” of Functionally Related Genes Facilitates Coordinated Gene Transcription
5. Impact of Genomic Sequence Organization and Physical Locations of Genes on Transcription
6. Possible Impact of Transcription on Subcellular Localization of Genes
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Dame, R.T.; Rashid, F.-Z.M.; Grainger, D.C. Chromosome organization in bacteria: Mechanistic insights into genome structure and function. Nat. Rev. Genet. 2020, 21, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Umbarger, M.A.; Toro, E.; Wright, M.A.; Porreca, G.J.; Baù, D.; Hong, S.-H.; Fero, M.J.; Zhu, L.J.; Marti-Renom, M.A.; McAdams, H.H.; et al. The three-dimensional architecture of a bacterial genome and its alteration by genetic perturbation. Mol. Cell 2011, 44, 252–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiggins, P.A.; Cheveralls, K.C.; Martin, J.S.; Lintner, R.; Kondev, J. Strong intranucleoid interactions organize the Escherichia coli chromosome into a nucleoid filament. Proc. Natl. Acad. Sci. USA 2010, 107, 4991–4995. [Google Scholar] [CrossRef] [Green Version]
- Wray, N.R.; Ripke, S.; Mattheisen, M.; Trzaskowski, M.; Byrne, E.M.; Abdellaoui, A.; Adams, M.J.; Agerbo, E.; Air, T.M.; Andlauer, T.M.F.; et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat. Genet. 2018, 50, 668–681. [Google Scholar] [CrossRef] [Green Version]
- Sobetzko, P.; Travers, A.; Muskhelishvili, G. Gene order and chromosome dynamics coordinate spatiotemporal gene expression during the bacterial growth cycle. Proc. Natl. Acad. Sci. USA 2012, 109, E42–E50. [Google Scholar] [CrossRef] [Green Version]
- Kuhlman, T.E.; Cox, E.C. Gene location and DNA density determine transcription factor distributions in Escherichia coli. Mol. Syst. Biol. 2012, 8, 610. [Google Scholar] [CrossRef] [PubMed]
- Montero Llopis, P.; Jackson, A.F.; Sliusarenko, O.; Surovtsev, I.; Heinritz, J.; Emonet, T.; Jacobs-Wagner, C. Spatial organization of the flow of genetic information in bacteria. Nature 2010, 466, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Cabrera, N.; Casaña, P.; Cid, A.R.; Haya, S.; Moret, A.; Aznar, J.A. Novel missense mutation c.2685G>C (p.Q895H) in VWF gene associated with very low levels of VWF mRNA. Ann. Hematol. 2009, 88, 245–247. [Google Scholar] [CrossRef]
- Shen, B.A.; Landick, R. Transcription of Bacterial Chromatin. J. Mol. Biol. 2019, 431, 4040–4066. [Google Scholar] [CrossRef]
- Sobetzko, P. Transcription-coupled DNA supercoiling dictates the chromosomal arrangement of bacterial genes. Nucleic Acids Res. 2016, 44, 1514–1524. [Google Scholar] [CrossRef]
- Weng, X.; Bohrer, C.H.; Bettridge, K.; Lagda, A.C.; Cagliero, C.; Jin, D.J.; Xiao, J. Spatial organization of RNA polymerase and its relationship with transcription in Escherichia coli. Proc. Natl. Acad. Sci. USA 2019, 116, 20115–20123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Landick, R. A Two-Way Street: Regulatory Interplay between RNA Polymerase and Nascent RNA Structure. Trends Biochem. Sci. 2016, 41, 293–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorman, C.J. DNA supercoiling and transcription in bacteria: A two-way street. BMC Mol. Cell Biol. 2019, 20, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hołówka, J.; Zakrzewska-Czerwińska, J. Nucleoid Associated Proteins: The Small Organizers That Help to Cope with Stress. Front. Microbiol. 2020, 11, 590. [Google Scholar] [CrossRef]
- Dillon, S.C.; Dorman, C.J. Bacterial nucleoid-associated proteins, nucleoid structure and gene expression. Nat. Rev. Microbiol. 2010, 8, 185–195. [Google Scholar] [CrossRef]
- Dorman, C.J. Function of nucleoid-associated proteins in chromosome structuring and transcriptional regulation. J. Mol. Microbiol. Biotechnol. 2014, 24, 316–331. [Google Scholar] [CrossRef] [Green Version]
- Rimsky, S.; Travers, A. Pervasive regulation of nucleoid structure and function by nucleoid-associated proteins. Curr. Opin. Microbiol. 2011, 14, 136–141. [Google Scholar] [CrossRef]
- Taniguchi, S.; Kasho, K.; Ozaki, S.; Katayama, T. Escherichia coli CrfC Protein, a Nucleoid Partition Factor, Localizes to Nucleoid Poles via the Activities of Specific Nucleoid-Associated Proteins. Front. Microbiol. 2019, 10, 72. [Google Scholar] [CrossRef]
- Blot, N.; Mavathur, R.; Geertz, M.; Travers, A.; Muskhelishvili, G. Homeostatic regulation of supercoiling sensitivity coordinates transcription of the bacterial genome. EMBO Rep. 2006, 7, 710–715. [Google Scholar] [CrossRef] [Green Version]
- Ali Azam, T.; Iwata, A.; Nishimura, A.; Ueda, S.; Ishihama, A. Growth phase-dependent variation in protein composition of the Escherichia coli nucleoid. J. Bacteriol. 1999, 181, 6361–6370. [Google Scholar] [CrossRef] [Green Version]
- Balke, V.L.; Gralla, J.D. Changes in the linking number of supercoiled DNA accompany growth transitions in Escherichia coli. J. Bacteriol. 1987, 169, 4499–4506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnenschein, N.; Geertz, M.; Muskhelishvili, G.; Hütt, M.-T. Analog regulation of metabolic demand. BMC Syst. Biol. 2011, 5, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Travers, A.; Muskhelishvili, G. DNA supercoiling—A global transcriptional regulator for enterobacterial growth? Nat. Rev. Microbiol. 2005, 3, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, L.S.; Burger, R.M.; Drlica, K. Bacterial DNA supercoiling and [ATP]/[ADP]. Changes associated with a transition to anaerobic growth. J. Mol. Biol. 1991, 219, 443–450. [Google Scholar] [CrossRef]
- Martis, B.S.; Forquet, R.; Reverchon, S.; Nasser, W.; Meyer, S. DNA Supercoiling: An Ancestral Regulator of Gene Expression in Pathogenic Bacteria? Comput. Struct. Biotechnol. J. 2019, 17, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Maurer, S.; Fritz, J.; Muskhelishvili, G. A systematic in vitro study of nucleoprotein complexes formed by bacterial nucleoid-associated proteins revealing novel types of DNA organization. J. Mol. Biol. 2009, 387, 1261–1276. [Google Scholar] [CrossRef]
- Prieto, A.I.; Kahramanoglou, C.; Ali, R.M.; Fraser, G.M.; Seshasayee, A.S.N.; Luscombe, N.M. Genomic analysis of DNA binding and gene regulation by homologous nucleoid-associated proteins IHF and HU in Escherichia coli K12. Nucleic Acids Res. 2012, 40, 3524–3537. [Google Scholar] [CrossRef]
- Cameron, A.D.S.; Stoebel, D.M.; Dorman, C.J. DNA supercoiling is differentially regulated by environmental factors and FIS in Escherichia coli and Salmonella enterica. Mol. Microbiol. 2011, 80, 85–101. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Sobetzko, P.; Nasser, W.; Reverchon, S.; Muskhelishvili, G. Chromosomal “stress-response” domains govern the spatiotemporal expression of the bacterial virulence program. MBio 2015, 6, e00353-15. [Google Scholar] [CrossRef] [Green Version]
- Ouafa, Z.-A.; Reverchon, S.; Lautier, T.; Muskhelishvili, G.; Nasser, W. The nucleoid-associated proteins H-NS and FIS modulate the DNA supercoiling response of the pel genes, the major virulence factors in the plant pathogen bacterium Dickeya dadantii. Nucleic Acids Res. 2012, 40, 4306–4319. [Google Scholar] [CrossRef]
- Reverchon, S.; Meyer, S.; Forquet, R.; Hommais, F.; Muskhelishvili, G.; Nasser, W. The nucleoid-associated protein IHF acts as a “transcriptional domainin” protein coordinating the bacterial virulence traits with global transcription. Nucleic Acids Res. 2021, 49, 776–790. [Google Scholar] [CrossRef] [PubMed]
- Atlung, T.; Ingmer, H. H-NS: A modulator of environmentally regulated gene expression. Mol. Microbiol. 1997, 24, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Datta, C.; Jha, R.K.; Ahmed, W.; Ganguly, S.; Ghosh, S.; Nagaraja, V. Physical and functional interaction between nucleoid-associated proteins HU and Lsr2 of Mycobacterium tuberculosis: Altered DNA binding and gene regulation. Mol. Microbiol. 2019, 111, 981–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangan, M.W.; Lucchini, S.; Cróinín, T.Ó.; Fitzgerald, S.; Hinton, J.C.D.; Dorman, C.J. Nucleoid-associated protein HU controls three regulons that coordinate virulence, response to stress and general physiology in Salmonella enterica serovar Typhimurium. Microbiology 2011, 157, 1075–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Shahul Hameed, U.F.; Kharchenko, V.; Liao, C.; Huser, F.; Remington, J.M.; Radhakrishnan, A.K.; Jaremko, M.; Jaremko, Ł.; Arold, S.T.; et al. Molecular basis for the adaptive evolution of environment-sensing by H-NS proteins. Elife 2021, 10, e57467. [Google Scholar] [CrossRef]
- Bouffartigues, E.; Buckle, M.; Badaut, C.; Travers, A.; Rimsky, S. H-NS cooperative binding to high-affinity sites in a regulatory element results in transcriptional silencing. Nat. Struct. Mol. Biol. 2007, 14, 441–448. [Google Scholar] [CrossRef]
- Lang, B.; Blot, N.; Bouffartigues, E.; Buckle, M.; Geertz, M.; Gualerzi, C.O.; Mavathur, R.; Muskhelishvili, G.; Pon, C.L.; Rimsky, S.; et al. High-affinity DNA binding sites for H-NS provide a molecular basis for selective silencing within proteobacterial genomes. Nucleic Acids Res. 2007, 35, 6330–6337. [Google Scholar] [CrossRef]
- Castang, S.; McManus, H.R.; Turner, K.H.; Dove, S.L. H-NS family members function coordinately in an opportunistic pathogen. Proc. Natl. Acad. Sci. USA 2008, 105, 18947–18952. [Google Scholar] [CrossRef] [Green Version]
- Falconi, M.; Colonna, B.; Prosseda, G.; Micheli, G.; Gualerzi, C.O. Thermoregulation of Shigella and Escherichia coli EIEC pathogenicity. A temperature-dependent structural transition of DNA modulates accessibility of virF promoter to transcriptional repressor H-NS. EMBO J. 1998, 17, 7033–7043. [Google Scholar] [CrossRef] [Green Version]
- Nasser, W.; Faelen, M.; Hugouvieux-Cotte-Pattat, N.; Reverchon, S. Role of the nucleoid-associated protein H-NS in the synthesis of virulence factors in the phytopathogenic bacterium Erwinia chrysanthemi. Mol. Plant-Microbe Interact. 2001, 14, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Prosseda, G.; Fradiani, P.A.; Di Lorenzo, M.; Falconi, M.; Micheli, G.; Casalino, M.; Nicoletti, M.; Colonna, B. A role for H-NS in the regulation of the virF gene of Shigella and enteroinvasive Escherichia coli. Res. Microbiol. 1998, 149, 15–25. [Google Scholar] [CrossRef]
- Tran, C.N.; Giangrossi, M.; Prosseda, G.; Brandi, A.; Di Martino, M.L.; Colonna, B.; Falconi, M. A multifactor regulatory circuit involving H-NS, VirF and an antisense RNA modulates transcription of the virulence gene icsA of Shigella flexneri. Nucleic Acids Res. 2011, 39, 8122–8134. [Google Scholar] [CrossRef]
- Song, D.; Loparo, J.J. Building bridges within the bacterial chromosome. Trends Genet. 2015, 31, 164–173. [Google Scholar] [CrossRef] [PubMed]
- van der Valk, R.A.; Vreede, J.; Qin, L.; Moolenaar, G.F.; Hofmann, A.; Goosen, N.; Dame, R.T. Mechanism of environmentally driven conformational changes that modulate H-NS DNA-bridging activity. Elife 2017, 6, e27369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahul Hameed, U.F.; Liao, C.; Radhakrishnan, A.K.; Huser, F.; Aljedani, S.S.; Zhao, X.; Momin, A.A.; Melo, F.A.; Guo, X.; Brooks, C.; et al. H-NS uses an autoinhibitory conformational switch for environment-controlled gene silencing. Nucleic Acids Res. 2019, 47, 2666–2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, W.; Thompson, J.F.; Newlands, J.T.; Gourse, R.L.E. coli Fis protein activates ribosomal RNA transcription in vitro and in vivo. EMBO J. 1990, 9, 3733–3742. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.; Travers, A.; Kutateladze, T.; Muskhelishvili, G. A DNA architectural protein couples cellular physiology and DNA topology in Escherichia coli. Mol. Microbiol. 1999, 34, 953–964. [Google Scholar] [CrossRef]
- Weinstein-Fischer, D.; Elgrably-Weiss, M.; Altuvia, S. Escherichia coli response to hydrogen peroxide: A role for DNA supercoiling, topoisomerase I and Fis. Mol. Microbiol. 2000, 35, 1413–1420. [Google Scholar] [CrossRef]
- Schneider, R.; Travers, A.; Muskhelishvili, G. FIS modulates growth phase-dependent topological transitions of DNA in Escherichia coli. Mol. Microbiol. 1997, 26, 519–530. [Google Scholar] [CrossRef]
- Ball, C.A.; Osuna, R.; Ferguson, K.C.; Johnson, R.C. Dramatic changes in Fis levels upon nutrient upshift in Escherichia coli. J. Bacteriol. 1992, 174, 8043–8056. [Google Scholar] [CrossRef] [Green Version]
- Browning, D.F.; Grainger, D.C.; Busby, S.J. Effects of nucleoid-associated proteins on bacterial chromosome structure and gene expression. Curr. Opin. Microbiol. 2010, 13, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.D.; Johnson, M.; Hinton, J.C.; Williams, P.H. Role of the nucleoid-associated protein Fis in the regulation of virulence properties of enteropathogenic Escherichia coli. Mol. Microbiol. 2001, 41, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Falconi, M.; Prosseda, G.; Giangrossi, M.; Beghetto, E.; Colonna, B. Involvement of FIS in the H-NS-mediated regulation of virF gene of Shigella and enteroinvasive Escherichia coli. Mol. Microbiol. 2001, 42, 439–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cróinín, T.O.; Carroll, R.K.; Kelly, A.; Dorman, C.J. Roles for DNA supercoiling and the Fis protein in modulating expression of virulence genes during intracellular growth of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2006, 62, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Schechter, L.M.; Jain, S.; Akbar, S.; Lee, C.A. The small nucleoid-binding proteins H-NS, HU, and Fis affect hilA expression in Salmonella enterica serovar Typhimurium. Infect. Immun. 2003, 71, 5432–5435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenz, D.H.; Bassler, B.L. The small nucleoid protein Fis is involved in Vibrio cholerae quorum sensing. Mol. Microbiol. 2007, 63, 859–871. [Google Scholar] [CrossRef]
- Lautier, T.; Nasser, W. The DNA nucleoid-associated protein Fis co-ordinates the expression of the main virulence genes in the phytopathogenic bacterium Erwinia chrysanthemi. Mol. Microbiol. 2007, 66, 1474–1490. [Google Scholar] [CrossRef]
- Sheridan, S.D.; Benham, C.J.; Hatfield, G.W. Activation of gene expression by a novel DNA structural transmission mechanism that requires supercoiling-induced DNA duplex destabilization in an upstream activating sequence. J. Biol. Chem. 1998, 273, 21298–21308. [Google Scholar] [CrossRef] [Green Version]
- Mangan, M.W.; Lucchini, S.; Danino, V.; Cróinín, T.O.; Hinton, J.C.D.; Dorman, C.J. The integration host factor (IHF) integrates stationary-phase and virulence gene expression in Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2006, 59, 1831–1847. [Google Scholar] [CrossRef]
- Stonehouse, E.; Kovacikova, G.; Taylor, R.K.; Skorupski, K. Integration host factor positively regulates virulence gene expression in Vibrio cholerae. J. Bacteriol. 2008, 190, 4736–4748. [Google Scholar] [CrossRef] [Green Version]
- Remesh, S.G.; Verma, S.C.; Chen, J.-H.; Ekman, A.A.; Larabell, C.A.; Adhya, S.; Hammel, M. Nucleoid remodeling during environmental adaptation is regulated by HU-dependent DNA bundling. Nat. Commun. 2020, 11, 2905. [Google Scholar] [CrossRef] [PubMed]
- Oberto, J.; Nabti, S.; Jooste, V.; Mignot, H.; Rouviere-Yaniv, J. The HU regulon is composed of genes responding to anaerobiosis, acid stress, high osmolarity and SOS induction. PLoS ONE 2009, 4, e4367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, M.; Farcas, A.; Geertz, M.; Zhelyazkova, P.; Brix, K.; Travers, A.; Muskhelishvili, G. Coordination of genomic structure and transcription by the main bacterial nucleoid-associated protein HU. EMBO Rep. 2010, 11, 59–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroner, G.M.; Wolfe, M.B.; Freddolino, P.L. Escherichia coli Lrp Regulates One-Third of the Genome via Direct, Cooperative, and Indirect Routes. J. Bacteriol. 2019, 201, e00411-18. [Google Scholar] [CrossRef] [Green Version]
- Tani, T.H.; Khodursky, A.; Blumenthal, R.M.; Brown, P.O.; Matthews, R.G. Adaptation to famine: A family of stationary-phase genes revealed by microarray analysis. Proc. Natl. Acad. Sci. USA 2002, 99, 13471–13476. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, C.P.; Dorman, C.J. DNA relaxation-dependent phase biasing of the fim genetic switch in Escherichia coli depends on the interplay of H-NS, IHF and LRP. Mol. Microbiol. 2009, 74, 1071–1082. [Google Scholar] [CrossRef]
- Engstrom, M.D.; Mobley, H.L.T. Regulation of Expression of Uropathogenic Escherichia coli Nonfimbrial Adhesin TosA by PapB Homolog TosR in Conjunction with H-NS and Lrp. Infect. Immun. 2016, 84, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Foer, D.; Zhu, M.; Cardone, R.L.; Simpson, C.; Sullivan, R.; Nemiroff, S.; Lee, G.; Kibbey, R.G.; Petersen, K.F.; Insogna, K.L. Impact of gain-of-function mutations in the low-density lipoprotein receptor-related protein 5 (LRP5) on glucose and lipid homeostasis. Osteoporos. Int. 2017, 28, 2011–2017. [Google Scholar] [CrossRef]
- Browning, D.F.; Busby, S.J.W. Local and global regulation of transcription initiation in bacteria. Nat. Rev. Microbiol. 2016, 14, 638–650. [Google Scholar] [CrossRef] [Green Version]
- Kröger, C.; MacKenzie, K.D.; Alshabib, E.Y.; Kirzinger, M.W.B.; Suchan, D.M.; Chao, T.-C.; Akulova, V.; Miranda-CasoLuengo, A.A.; Monzon, V.A.; Conway, T.; et al. The primary transcriptome, small RNAs and regulation of antimicrobial resistance in Acinetobacter baumannii ATCC 17978. Nucleic Acids Res. 2018, 46, 9684–9698. [Google Scholar] [CrossRef] [Green Version]
- Meyer, S.; Reverchon, S.; Nasser, W.; Muskhelishvili, G. Chromosomal organization of transcription: In a nutshell. Curr. Genet. 2018, 64, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Kusano, S.; Ding, Q.; Fujita, N.; Ishihama, A. Promoter selectivity of Escherichia coli RNA polymerase E sigma 70 and E sigma 38 holoenzymes. Effect of DNA supercoiling. J. Biol. Chem. 1996, 271, 1998–2004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.F.; Wang, J.C. Supercoiling of the DNA template during transcription. Proc. Natl. Acad. Sci. USA 1987, 84, 7024–7027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhi, X.; Dages, S.; Dages, K.; Liu, Y.; Hua, Z.-C.; Makemson, J.; Leng, F. Transient and dynamic DNA supercoiling potently stimulates the leu-500 promoter in Escherichia coli. J. Biol. Chem. 2017, 292, 14566–14575. [Google Scholar] [CrossRef] [Green Version]
- Conway, T.; Creecy, J.P.; Maddox, S.M.; Grissom, J.E.; Conkle, T.L.; Shadid, T.M.; Teramoto, J.; San Miguel, P.; Shimada, T.; Ishihama, A.; et al. Unprecedented high-resolution view of bacterial operon architecture revealed by RNA sequencing. MBio 2014, 5, e01442-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harden, T.T.; Herlambang, K.S.; Chamberlain, M.; Lalanne, J.-B.; Wells, C.D.; Li, G.-W.; Landick, R.; Hochschild, A.; Kondev, J.; Gelles, J. Alternative transcription cycle for bacterial RNA polymerase. Nat. Commun. 2020, 11, 448. [Google Scholar] [CrossRef] [Green Version]
- Cevost, J.; Vaillant, C.; Meyer, S.; Rost, B. ThreaDNA: Predicting DNA mechanics’ contribution to sequence selectivity of proteins along whole genomes. Bioinformatics 2018, 34, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Junier, I.; Rivoire, O. Conserved Units of Co-Expression in Bacterial Genomes: An Evolutionary Insight into Transcriptional Regulation. PLoS ONE 2016, 11, e0155740. [Google Scholar] [CrossRef] [Green Version]
- Deato, M.D.E.; Marr, M.T.; Sottero, T.; Inouye, C.; Hu, P.; Tjian, R. MyoD targets TAF3/TRF3 to activate myogenin transcription. Mol. Cell 2008, 32, 96–105. [Google Scholar] [CrossRef] [Green Version]
- Jeong, K.-H.; Chin, W.W.; Kaiser, U.B. Essential role of the homeodomain for pituitary homeobox 1 activation of mouse gonadotropin-releasing hormone receptor gene expression through interactions with c-Jun and DNA. Mol. Cell. Biol. 2004, 24, 6127–6139. [Google Scholar] [CrossRef] [Green Version]
- Sonnenschein, N.; Hütt, M.-T.; Stoyan, H.; Stoyan, D. Ranges of control in the transcriptional regulation of Escherichia coli. BMC Syst. Biol. 2009, 3, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muskhelishvili, G.; Travers, A. Integration of syntactic and semantic properties of the DNA code reveals chromosomes as thermodynamic machines converting energy into information. Cell. Mol. Life Sci. 2013, 70, 4555–4567. [Google Scholar] [CrossRef] [PubMed]
- Postow, L.; Hardy, C.D.; Arsuaga, J.; Cozzarelli, N.R. Topological domain structure of the Escherichia coli chromosome. Genes Dev. 2004, 18, 1766–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesterlin, C.; Gigant, E.; Boccard, F.; Espéli, O. Sister chromatid interactions in bacteria revealed by a site-specific recombination assay. EMBO J. 2012, 31, 3468–3479. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.M. Notch signaling and Notch signaling modifiers. Int. J. Biochem. Cell Biol. 2011, 43, 1550–1562. [Google Scholar] [CrossRef] [Green Version]
- Yazdi, S.; Ardekani, A.M. Bacterial aggregation and biofilm formation in a vortical flow. Biomicrofluidics 2012, 6, 44114. [Google Scholar] [CrossRef] [Green Version]
- Bouyioukos, C.; Reverchon, S.; Képès, F. From multiple pathogenicity islands to a unique organized pathogenicity archipelago. Sci. Rep. 2016, 6, 27978. [Google Scholar] [CrossRef] [Green Version]
- Junier, P.; Molina, V.; Dorador, C.; Hadas, O.; Kim, O.-S.; Junier, T.; Witzel, J.-P.; Imhoff, J.F. Phylogenetic and functional marker genes to study ammonia-oxidizing microorganisms (AOM) in the environment. Appl. Microbiol. Biotechnol. 2010, 85, 425–440. [Google Scholar] [CrossRef] [Green Version]
- Képès, F. Periodic transcriptional organization of the E.coli genome. J. Mol. Biol. 2004, 340, 957–964. [Google Scholar] [CrossRef]
- Képès, F.; Jester, B.C.; Lepage, T.; Rafiei, N.; Rosu, B.; Junier, I. The layout of a bacterial genome. FEBS Lett. 2012, 586, 2043–2048. [Google Scholar] [CrossRef]
- Korotkov, K.V.; Sandkvist, M.; Hol, W.G.J. The type II secretion system: Biogenesis, molecular architecture and mechanism. Nat. Rev. Microbiol. 2012, 10, 336–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magaud, D.; Grandjean, C.; Doutheau, A.; Anker, D.; Shevchik, V.; Cotte-Pattat, N.; Robert-Baudouy, J. Synthesis of the two monomethyl esters of the disaccharide 4-O-alpha-D-galacturonosyl-D-galacturonic acid and of precursors for the preparation of higher oligomers methyl uronated in definite sequences. Carbohydr. Res. 1998, 314, 189–199. [Google Scholar] [CrossRef]
- Nasser, W.; Reverchon, S.; Condemine, G.; Robert-Baudouy, J. Specific interactions of Erwinia chrysanthemi KdgR repressor with different operators of genes involved in pectinolysis. J. Mol. Biol. 1994, 236, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Reverchon, S.; Nasser, W.; Robert-Baudouy, J. Characterization of kdgR, a gene of Erwinia chrysanthemi that regulates pectin degradation. Mol. Microbiol. 1991, 5, 2203–2216. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, E.; Sclavi, B. Gene regulation by H-NS as a function of growth conditions depends on chromosomal position in Escherichia coli. G3 Genes Genomes Genet. 2015, 5, 605–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerganova, V.; Berger, M.; Zaldastanishvili, E.; Sobetzko, P.; Lafon, C.; Mourez, M.; Travers, A.; Muskhelishvili, G. Chromosomal position shift of a regulatory gene alters the bacterial phenotype. Nucleic Acids Res. 2015, 43, 8215–8226. [Google Scholar] [CrossRef] [Green Version]
- Fritsche, M.; Li, S.; Heermann, D.W.; Wiggins, P.A. A model for Escherichia coli chromosome packaging supports transcription factor-induced DNA domain formation. Nucleic Acids Res. 2012, 40, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Japaridze, A.; Yang, W.; Dekker, C.; Nasser, W.; Muskhelishvili, G. DNA sequence-directed cooperation between nucleoid-associated proteins. Iscience 2021, 24, 102408. [Google Scholar] [CrossRef]
- Lioy, V.S.; Boccard, F. Conformational Studies of Bacterial Chromosomes by High-Throughput Sequencing Methods. Methods Enzymol. 2018, 612, 25–45. [Google Scholar] [CrossRef]
- Krogh, T.J.; Franke, A.; Møller-Jensen, J.; Kaleta, C. Elucidating the Influence of Chromosomal Architecture on Transcriptional Regulation in Prokaryotes—Observing Strong Local Effects of Nucleoid Structure on Gene Regulation. Front. Microbiol. 2020, 11, 2002. [Google Scholar] [CrossRef]
- Booker, B.M.; Deng, S.; Higgins, N.P. DNA topology of highly transcribed operons in Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2010, 78, 1348–1364. [Google Scholar] [CrossRef] [PubMed]
- Higgins, N.P. RNA polymerase: Chromosome domain boundary maker and regulator of supercoil density. Curr. Opin. Microbiol. 2014, 22, 138–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Dame, R.T.; Kalmykowa, O.J.; Grainger, D.C. Chromosomal macrodomains and associated proteins: Implications for DNA organization and replication in gram negative bacteria. PLoS Genet. 2011, 7, e1002123. [Google Scholar] [CrossRef] [Green Version]
- Scolari, V.F.; Bassetti, B.; Sclavi, B.; Lagomarsino, M.C. Gene clusters reflecting macrodomain structure respond to nucleoid perturbations. Mol. Biosyst. 2011, 7, 878–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrándiz, M.-J.; Martín-Galiano, A.J.; Schvartzman, J.B.; de la Campa, A.G. The genome of Streptococcus pneumoniae is organized in topology-reacting gene clusters. Nucleic Acids Res. 2010, 38, 3570–3581. [Google Scholar] [CrossRef] [Green Version]
- Muskhelishvili, G.; Forquet, R.; Reverchon, S.; Meyer, S.; Nasser, W. Coherent Domains of Transcription Coordinate Gene Expression During Bacterial Growth and Adaptation. Microorganisms 2019, 7, 694. [Google Scholar] [CrossRef] [Green Version]
- Wasim, A.; Gupta, A.; Mondal, J. A Hi-C data-integrated model elucidates E. coli chromosome’s multiscale organization at various replication stages. Nucleic Acids Res. 2021, 49, 3077–3091. [Google Scholar] [CrossRef]
- Lioy, V.S.; Lorenzi, J.-N.; Najah, S.; Poinsignon, T.; Leh, H.; Saulnier, C.; Aigle, B.; Lautru, S.; Thibessard, A.; Lespinet, O.; et al. Dynamics of the compartmentalized Streptomyces chromosome during metabolic differentiation. Nat. Commun. 2021, 12, 5221. [Google Scholar] [CrossRef]
- Le, T.B.; Laub, M.T. Transcription rate and transcript length drive formation of chromosomal interaction domain boundaries. EMBO J. 2016, 35, 1582–1595. [Google Scholar] [CrossRef] [Green Version]
- Chereji, R.V.; Bharatula, V.; Elfving, N.; Blomberg, J.; Larsson, M.; Morozov, A.V.; Broach, J.R.; Björklund, S. Mediator binds to boundaries of chromosomal interaction domains and to proteins involved in DNA looping, RNA metabolism, chromatin remodeling, and actin assembly. Nucleic Acids Res. 2017, 45, 8806–8821. [Google Scholar] [CrossRef] [PubMed]
- Marbouty, M.; Koszul, R. Metagenome Analysis Exploiting High-Throughput Chromosome Conformation Capture (3C) Data. Trends Genet. 2015, 31, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Trussart, M.; Yus, E.; Martinez, S.; Baù, D.; Tahara, Y.O.; Pengo, T.; Widjaja, M.; Kretschmer, S.; Swoger, J.; Djordjevic, S.; et al. Defined chromosome structure in the genome-reduced bacterium Mycoplasma pneumoniae. Nat. Commun. 2017, 8, 14665. [Google Scholar] [CrossRef] [PubMed]
- Dugar, G.; Hofmann, A.; Heermann, D.W.; Hamoen, L.W. A chromosomal loop anchor mediates bacterial genome organization. Nat. Genet. 2022, 54, 194–201. [Google Scholar] [CrossRef]
- Cagliero, C.; Grand, R.S.; Jones, M.B.; Jin, D.J.; O’Sullivan, J.M. Genome conformation capture reveals that the Escherichia coli chromosome is organized by replication and transcription. Nucleic Acids Res. 2013, 41, 6058–6071. [Google Scholar] [CrossRef] [Green Version]
- Espeli, O.; Mercier, R.; Boccard, F. DNA dynamics vary according to macrodomain topography in the E. coli chromosome. Mol. Microbiol. 2008, 68, 1418–1427. [Google Scholar] [CrossRef]
- Badrinarayanan, A.; Le, T.B.K.; Laub, M.T. Bacterial chromosome organization and segregation. Annu. Rev. Cell Dev. Biol. 2015, 31, 171–199. [Google Scholar] [CrossRef] [Green Version]
- Jin, D.J.; Cagliero, C.; Martin, C.M.; Izard, J.; Zhou, Y.N. The dynamic nature and territory of transcriptional machinery in the bacterial chromosome. Front. Microbiol. 2015, 6, 497. [Google Scholar] [CrossRef] [Green Version]
- Scholz, S.A.; Diao, R.; Wolfe, M.B.; Fivenson, E.M.; Lin, X.N.; Freddolino, P.L. High-Resolution Mapping of the Escherichia coli Chromosome Reveals Positions of High and Low Transcription. Cell Syst. 2019, 8, 212–225.e9. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Bai, L.; Wang, M.D. Transcription under torsion. Science 2013, 340, 1580–1583. [Google Scholar] [CrossRef] [Green Version]
- Sobetzko, P.; Glinkowska, M.; Travers, A.; Muskhelishvili, G. DNA thermodynamic stability and supercoil dynamics determine the gene expression program during the bacterial growth cycle. Mol. BioSyst. 2013, 9, 1643–1651. [Google Scholar] [CrossRef]
- Meyer, S.; Beslon, G. Torsion-mediated interaction between adjacent genes. PLoS Comput. Biol. 2014, 10, e1003785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bremer, H.; Dennis, P.; Ehrenberg, M. Free RNA polymerase and modeling global transcription in Escherichia coli. Biochimie 2003, 85, 597–609. [Google Scholar] [CrossRef]
- Jin, D.J.; Cabrera, J.E. Coupling the distribution of RNA polymerase to global gene regulation and the dynamic structure of the bacterial nucleoid in Escherichia coli. J. Struct. Biol. 2006, 156, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.J.; Mata Martin, C.; Sun, Z.; Cagliero, C.; Zhou, Y.N. Nucleolus-like compartmentalization of the transcription machinery in fast-growing bacterial cells. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stracy, M.; Lesterlin, C.; Garza de Leon, F.; Uphoff, S.; Zawadzki, P.; Kapanidis, A.N. Live-cell superresolution microscopy reveals the organization of RNA polymerase in the bacterial nucleoid. Proc. Natl. Acad. Sci. USA 2015, 112, E4390–E4399. [Google Scholar] [CrossRef] [Green Version]
- Bakshi, S.; Choi, H.; Mondal, J.; Weisshaar, J.C. Time-dependent effects of transcription- and translation-halting drugs on the spatial distributions of the Escherichia coli chromosome and ribosomes. Mol. Microbiol. 2014, 94, 871–887. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Kim, S.; Kim, D.-K.; Jeon An, H.; Bae Son, J.; Hedén Gynnå, A.; Ki Lee, N. Transcription and translation contribute to gene locus relocation to the nucleoid periphery in E. coli. Nat. Commun. 2019, 10, 5131. [Google Scholar] [CrossRef] [Green Version]
- Ferrándiz, M.-J.; Carreño, D.; Ayora, S.; de la Campa, A. HU of Streptococcus pneumoniae Is Essential for the Preservation of DNA Supercoiling. Front. Microbiol. 2018, 9, 493. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Bensaid, A.; Rouviere-Yaniv, J.; Drlica, K. Histone-like protein HU and bacterial DNA topology: Suppression of an HU deficiency by gyrase mutations. J. Mol. Biol. 1996, 256, 66–76. [Google Scholar] [CrossRef]
- Cannon, W.; Bordes, P.; Wigneshweraraj, S.R.; Buck, M. Nucleotide-dependent triggering of RNA polymerase-DNA interactions by an AAA regulator of transcription. J. Biol. Chem. 2003, 278, 19815–19825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geertz, M.; Travers, A.; Mehandziska, S.; Sobetzko, P.; Chandra-Janga, S.; Shimamoto, N.; Muskhelishvili, G. Structural coupling between RNA polymerase composition and DNA supercoiling in coordinating transcription: A global role for the omega subunit? MBio 2011, 2, e00034-11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohniwa, R.L.; Ushijima, Y.; Saito, S.; Morikawa, K. Proteomic analyses of nucleoid-associated proteins in Escherichia coli, Pseudomonas aeruginosa, Bacillus subtilis, and Staphylococcus aureus. PLoS ONE 2011, 6, e19172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafiei, N.; Cordova, M.; Navarre, W.W.; Milstein, J.N. Growth Phase-Dependent Chromosome Condensation and Heat-Stable Nucleoid-Structuring Protein Redistribution in Escherichia coli under Osmotic Stress. J. Bacteriol. 2019, 201, e00469-19. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Berre, D.; Reverchon, S.; Muskhelishvili, G.; Nasser, W. Relationship between the Chromosome Structural Dynamics and Gene Expression—A Chicken and Egg Dilemma? Microorganisms 2022, 10, 846. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050846
Le Berre D, Reverchon S, Muskhelishvili G, Nasser W. Relationship between the Chromosome Structural Dynamics and Gene Expression—A Chicken and Egg Dilemma? Microorganisms. 2022; 10(5):846. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050846
Chicago/Turabian StyleLe Berre, Diana, Sylvie Reverchon, Georgi Muskhelishvili, and William Nasser. 2022. "Relationship between the Chromosome Structural Dynamics and Gene Expression—A Chicken and Egg Dilemma?" Microorganisms 10, no. 5: 846. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10050846