Mycobacterium tuberculosis Infection Up-Regulates Sialyl Lewis X Expression in the Lung Epithelium

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals
2.3. Bacteria and Bacterial Growth
2.4. M. tuberculosis Infection
2.5. Tissue Samples and Immunohistochemistry
2.6. RNA Extraction, cDNA Synthesis and Real-Time PCR Analysis
2.7. Statistical Analysis
3. Results
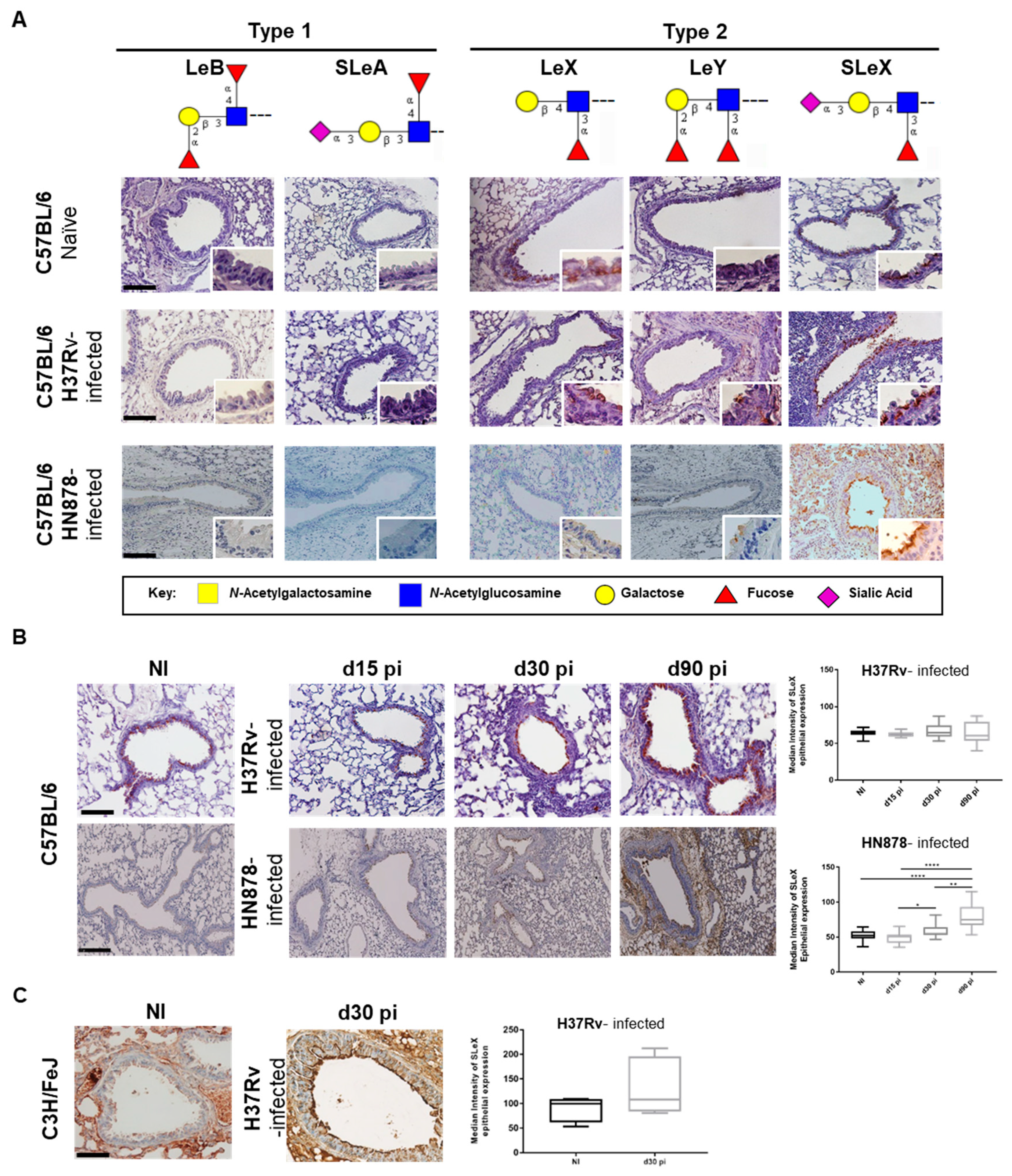
3.1. M. tuberculosis Infected Lungs Display Increased Sialylation
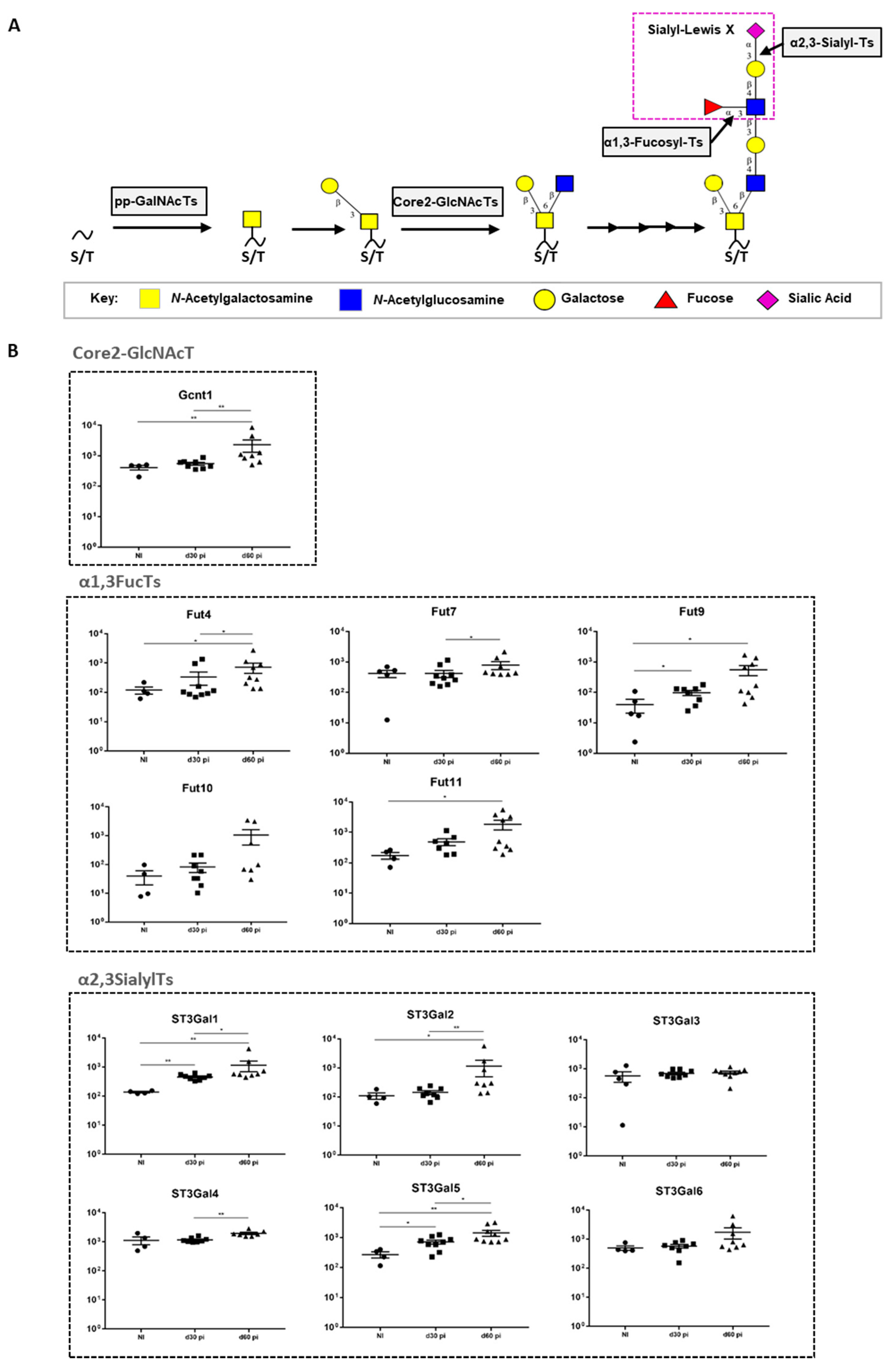
3.2. M. tuberculosis Infection Up-Regulates the Transcription of Enzymes Controlling the Biosynthesis of SLeX on Core-2 O-Glycans
3.3. Lack of Gcnt1 Does not Preclude Increased Lung Sialylation upon M. tuberculosis Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cummings, R.D. Stuck on sugars-how carbohydrates regulate cell adhesion, recognition, and signaling. Glycoconj. J. 2019, 36, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Mereiter, S.; Balmana, M.; Campos, D.; Gomes, J.; Reis, C.A. Glycosylation in the Era of Cancer-Targeted Therapy: Where Are We Heading? Cancer Cell 2019, 36, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A.V. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schjoldager, K.T.; Narimatsu, Y.; Joshi, H.J.; Clausen, H. Global view of human protein glycosylation pathways and functions. Nat. Rev. Mol. Cell Biol. 2020. [Google Scholar] [CrossRef]
- Lubbers, J.; Rodriguez, E.; van Kooyk, Y. Modulation of Immune Tolerance via Siglec-Sialic Acid Interactions. Front. Immunol. 2018, 9, 2807. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.S.; Alves, I.; Vicente, M.; Campar, A.; Silva, M.C.; Padrao, N.A.; Pinto, V.; Fernandes, A.; Dias, A.M.; Pinho, S.S. Glycans as Key Checkp. of T Cell Activity and Function. Front. Immunol. 2018, 9, 2754. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; van Kooyk, Y.; Cobb, B.A. Glycobiology of immune responses. Ann. N. Y. Acad. Sci. 2012, 1253, 1–15. [Google Scholar] [CrossRef]
- Sophie Groux-Degroote, S.C.; Uchimura, K.; Allain, F.; Delannoy, P. Chapter Four-Glycosylation changes in inflammatory diseases. In Advances in Protein Chemistry and Structural Biology; Donev, R., Ed.; Elsevier Academic Press: Cambridge, MA, USA, 2020; Volume 119, pp. 111–156. [Google Scholar]
- Kreisman, L.S.; Cobb, B.A. Infection, inflammation and host carbohydrates: A Glyco-Evasion Hypothesis. Glycobiology 2012, 22, 1019–1030. [Google Scholar] [CrossRef] [Green Version]
- Poole, J.; Day, C.J.; von Itzstein, M.; Paton, J.C.; Jennings, M.P. Glycointeractions in bacterial pathogenesis. Nat. Rev. Microbiol. 2018, 16, 440–452. [Google Scholar] [CrossRef]
- Magalhaes, A.; Marcos-Pinto, R.; Nairn, A.V.; Dela Rosa, M.; Ferreira, R.M.; Junqueira-Neto, S.; Freitas, D.; Gomes, J.; Oliveira, P.; Santos, M.R.; et al. Helicobacter pylori chronic infection and mucosal inflammation switches the human gastric glycosylation pathways. Biochim. Biophys. Acta 2015, 1852, 1928–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos, N.T.; Magalhaes, A.; Ferreira, B.; Oliveira, M.J.; Carvalho, A.S.; Mendes, N.; Gilmartin, T.; Head, S.R.; Figueiredo, C.; David, L.; et al. Helicobacter pylori induces beta3GnT5 in human gastric cell lines, modulating expression of the SabA ligand sialyl-Lewis x. J. Clin. Investig. 2008, 118, 2325–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, E. WHO global progress report on tuberculosis elimination. Lancet Respir. Med. 2020, 8, 19. [Google Scholar] [CrossRef]
- Fonseca, K.L.; Maceiras, A.R.; Matos, R.; Simoes-Costa, L.; Sousa, J.; Ca, B.; Barros, L.; Fernandes, A.I.; Mereiter, S.; Reis, R.; et al. Deficiency in the glycosyltransferase Gcnt1 increases susceptibility to tuberculosis through a mechanism involving neutrophils. Mucosal. Immunol. 2020, 13, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, S.; Schreiber, T.; Dunzendorfer, A.; Lowe, J.B.; Holscher, C. Fucosyltransferase IV and VII-directed selectin ligand function determines long-term survival in experimental tuberculosis. Immunobiology 2009, 214, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, T.; Ehlers, S.; Aly, S.; Holscher, A.; Hartmann, S.; Lipp, M.; Lowe, J.B.; Holscher, C. Selectin ligand-independent priming and maintenance of T cell immunity during airborne tuberculosis. J. Immunol. 2006, 176, 1131–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nairn, A.V.; Aoki, K.; dela Rosa, M.; Porterfield, M.; Lim, J.M.; Kulik, M.; Pierce, J.M.; Wells, L.; Dalton, S.; Tiemeyer, M.; et al. Regulation of glycan structures in murine embryonic stem cells: Combined transcript profiling of glycan-related genes and glycan structural analysis. J. Biol. Chem. 2012, 287, 37835–37856. [Google Scholar] [CrossRef] [Green Version]
- Delannoy, C.; Huang, C.; Coddeville, B.; Chen, J.Y.; Mouajjah, D.; Groux-Degroote, S.; Harduin-Lepers, A.; Khoo, K.H.; Guerardel, Y.; Elass-Rochard, E. Mycobacterium bovis BCG infection alters the macrophage N-glycome. Mol. Omics 2020, 16, 345–354. [Google Scholar] [CrossRef]
- Magalhaes, A.; Reis, C.A. Glycosyltransferases and Gastric Cancer. In Glycosignals in Cancer: Mechanims of Malignat Phenotypes; Springer: Berlin, Germany, 2016. [Google Scholar]
- Delmotte, P.; Degroote, S.; Lafitte, J.J.; Lamblin, G.; Perini, J.M.; Roussel, P. Tumor necrosis factor alpha increases the expression of glycosyltransferases and sulfotransferases responsible for the biosynthesis of sialylated and/or sulfated Lewis x epitopes in the human bronchial mucosa. J. Biol. Chem. 2002, 277, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Mahdavi, J.; Sonden, B.; Hurtig, M.; Olfat, F.O.; Forsberg, L.; Roche, N.; Angstrom, J.; Larsson, T.; Teneberg, S.; Karlsson, K.A.; et al. Helicobacter pylori SabA adhesin in persistent infection and chronic inflammation. Science 2002, 297, 573–578. [Google Scholar] [CrossRef] [Green Version]
- Ellies, L.G.; Tsuboi, S.; Petryniak, B.; Lowe, J.B.; Fukuda, M.; Marth, J.D. Core 2 oligosaccharide biosynthesis distinguishes between selectin ligands essential for leukocyte homing and inflammation. Immunity 1998, 9, 881–890. [Google Scholar] [CrossRef] [Green Version]
- Kroesen, V.M.; Rodriguez-Martinez, P.; Garcia, E.; Rosales, Y.; Diaz, J.; Martin-Cespedes, M.; Tapia, G.; Sarrias, M.R.; Cardona, P.J.; Vilaplana, C. A Beneficial Effect of Low-Dose Aspirin in a Murine Model of Active Tuberculosis. Front. Immunol. 2018, 9, 798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatt, K.; Machado, H.; Osório, N.S.; Sousa, J.; Cardoso, F.; Magalhães, C.; Chen, B.; Chen, M.; Kim, J.; Ferreira, C.M.; et al. A Nonribosomal Peptide Synthase gen Driving Virulence in Mycobacterium tuberculosis. mSphere 2018, 3, e00352-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira-Teixeira, L.; Sousa, J.; McNab, F.W.; Torrado, E.; Cardoso, F.; Machado, H.; Castro, F.; Cardoso, V.; Gaifem, J.; Wu, X.; et al. Type I IFN Inhibits Alternative Macrophage Activation during Mycobacterium tuberculosis Infection and Leads to Enhanced Protection in the Absence of IFN-gamma Signaling. J. Immunol. 2016, 197, 4714–4726. [Google Scholar] [CrossRef] [Green Version]
- Magalhaes, A.; Gomes, J.; Ismail, M.N.; Haslam, S.M.; Mendes, N.; Osorio, H.; David, L.; Le Pendu, J.; Haas, R.; Dell, A.; et al. Fut2-null mice display an altered glycosylation profile and impaired BabA-mediated Helicobacter pylori adhesion to gastric mucosa. Glycobiology 2009, 19, 1525–1536. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Fukushi, Y.; Hakomori, S.; Nudelman, E.; Cochran, N. Novel Fucolipids Accumulating in Human Adenocarcinoma. J. Biol. Chem. 1984, 259, 4681–4685. [Google Scholar]
- Abe, K.; McKibbin, J.M.; Hakomori, S. The Monoclonal Antibody Directed to Difucosylated Type 2 Chain (Fucα1-2Galβ1-4[Fucα1-3]GlcNAc; Y Determinant). J. Biol. Chem. 1993, 238, 11793–11797. [Google Scholar]
- Morozov, V.; Borkowski, J.; Hanisch, F.G. The Double Face of Mucin-Type O-Glycans in Lectin-Mediated Infection and Immunity. Molecules 2018, 23, 1151. [Google Scholar] [CrossRef] [Green Version]
- Fukushima, K.; Hirota, M.; Terasaki, P.I.; Wasisaka, A.; Togashi, H.; Chia, D.; Suyama, N.; Fukushi, Y.; Nudelman, E.; Hakomorf, S. Characterization of Sialosylated Lewis X as a new tumor-associated antigen. Cancer Res. 1984, 44, 5279–528528. [Google Scholar]
- Matsumura, R.; Hirakawa, J.; Sato, K.; Ikeda, T.; Nagai, M.; Fukuda, M.; Imai, Y.; Kawashima, H. Novel Antibodies Reactive with Sialyl Lewis X in Both Humans and Mice Define Its Critical Role in Leukocyte Trafficking and Contact Hypersensitivity Responses. J. Biol. Chem. 2015, 290, 15313–15326. [Google Scholar] [CrossRef] [Green Version]
- Mitoma, J.; Miyazaki, T.; Sutton-Smith, M.; Suzuki, M.; Saito, H.; Yeh, J.C.; Kawano, T.; Hindsgaul, O.; Seeberger, P.H.; Panico, M.; et al. The N-glycolyl form of mouse sialyl Lewis X is recognized by selectins but not by HECA-452 and FH6 antibodies that were raised against human cells. Glycoconj. J. 2009, 26, 511–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manca, C.; Tsenova, L.; Freeman, S.; Barczak, A.K.; Tovey, M.; Murray, P.J.; BarryIII, C.; Kaplan, G. Hypervirulent M. tuberculosis W/Beijing Strains Upregulate Type I IFNs and Increase Expression of Negative Regulators of the Jak-Stat Pathway. J. Interferon Cytokine Res. 2005, 25, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Teixeira, L.; Tabone, O.; Graham, C.M.; Singhania, A.; Stavropoulos, E.; Redford, P.S.; Chakravarty, P.; Priestnall, S.L.; Suarez-Bonnet, A.; Herbert, E.; et al. Mouse transcriptome reveals potential signatures of protection and pathogenesis in human tuberculosis. Nat. Immunol. 2020, 21, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.; Skerry, C.; Davis, S.L.; Tasneen, R.; Weir, M.; Kramnik, I.; Bishai, W.R.; Pomper, M.G.; Nuermberger, E.L.; Jain, S.K. Mouse model of necrotic tuberculosis granulomas develops hypoxic lesions. J. Infect. Dis. 2012, 205, 595–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neelamegham, S.; Aoki-Kinoshita, K.; Bolton, E.; Frank, M.; Lisacek, F.; Lutteke, T.; O’Boyle, N.; Packer, N.H.; Stanley, P.; Toukach, P.; et al. Updates to the Symbol Nomenclature for Glycans guidelines. Glycobiology 2019, 29, 620–624. [Google Scholar] [CrossRef]
- Sperandio, M.; Gleissner, C.A.; Ley, K. Glycosylation in immune cell trafficking. Immunol. Rev. 2009, 230, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.L.; Jones, M.B.; Ryan, S.O.; Cobb, B.A. The regulatory power of glycans and their binding partners in immunity. Trends Immunol. 2013, 34, 290–298. [Google Scholar] [CrossRef] [Green Version]
- van Kooyk, Y.; Rabinovich, G.A. Protein-glycan interactions in the control of innate and adaptive immune responses. Nat. Immunol. 2008, 9, 593–601. [Google Scholar] [CrossRef]
- Kumagai, T.; Palacios, A.; Casadevall, A.; Garcia, M.J.; Toro, C.; Tiemeyer, M.; Prados-Rosales, R. Serum IgM Glycosylation Associated with Tuberculosis Infection in Mice. mSphere 2019, 4, e00684-18. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.L.; Das, J.; Grace, P.S.; Fortune, S.M.; Restrepo, B.I.; Alter, G. Antibody Fc Glycosylation Discriminates between Latent and Active Tuberculosis. J. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, M.N.; Stone, E.L.; Panico, M.; Lee, S.H.; Luu, Y.; Ramirez, K.; Ho, S.B.; Fukuda, M.; Marth, J.D.; Haslam, S.M.; et al. High-sensitivity O-glycomic analysis of mice deficient in core 2 {beta}1,6-N-acetylglucosaminyltransferases. Glycobiology 2011, 21, 82–98. [Google Scholar] [CrossRef] [Green Version]
- Ohtsubo, K.; Marth, J.D. Glycosylation in cellular mechanisms of health and disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isa, F.; Collins, S.; Lee, M.H.; Decome, D.; Dorvil, N.; Joseph, P.; Smith, L.; Salerno, S.; Wells, M.T.; Fischer, S.; et al. Mass Spectrometric Identification of Urinary Biomarkers of Pulmonary Tuberculosis. EBioMedicine 2018, 31, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Magnani, J.L. The discovery, biology, and drug development of sialyl Lea and sialyl Lex. Arch. Biochem. Biophys. 2004, 426, 122–131. [Google Scholar] [CrossRef]
- Colomb, F.; Krzewinski-Recchi, M.A.; Steenackers, A.; Vincent, A.; Harduin-Lepers, A.; Delannoy, P.; Groux-Degroote, S. TNF up-regulates ST3GAL4 and sialyl-Lewisx expression in lung epithelial cells through an intronic ATF2-responsive element. Biochem. J. 2017, 474, 65–78. [Google Scholar] [CrossRef]
- Schroeter, M.F.; Ratsch, B.A.; Lehmann, J.; Baumgrass, R.; Hamann, A.; Syrbe, U. Differential regulation and impact of fucosyltransferase VII and core 2 beta1,6-N-acetyl-glycosaminyltransferase for generation of E-selectin and P-selectin ligands in murine CD4+ T cells. Immunology 2012, 137, 294–304. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Antigen | Antibody Clone | Dilution (IHC) | Company/Reference |
|---|---|---|---|
| Lewis B (LeB) | BG-6 (T218) | 1:50 | Signet |
| Sialyl-Lewis A (SLeA) | CA19-9 | 1:500 | Santa Cruz Biotechnology |
| Lewis X (LeX) | SH1 | 1:5 | [29] |
| Lewis Y (LeY) | AH6 | 1:10 | [30] |
| Sialyl-Lewis X (SLeX) | CSLEX-1 | 1:80 | BD Biosciences |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Gcnt1 | 5′- CGAAGGCCATGTTTCCAACGG -3′ | 5′- TCCGAAGACGCACACAGAGC -3′ |
| Fut4 | 5′- ACCAGGAGGGAGCAGTGACG -3′ | 5′- TCCACACCCACCTCTGCCC -3′ |
| Fut7 | 5´- GGACGACTTCAGCTCTGCCC -3′ | 5′- CGCCAAGCAAAGAAGCCACG -3′ |
| Fut9 | 5′- TCCCATGCGGTCCTGATTCAC -3′ | 5′- TTCTGAAAGGGTGGCCTGGC -3′ |
| Fut10 | 5′- GGGTGTGCAGGACATTAACC -3′ | 5′- AGCCTACTGTTTGCCCACAC -3′ |
| Fut11 | 5′- GGGTGCTCAGTGTCTGTTCGG -3′ | 5′- CCCACGGCTCCTCCCTCTC -3′ |
| St3gal1 | 5′- TCCTACAACTGCACAGCGTCG -3′ | 5′- TGTTTCGCCTGGTGCCTGG -3′ |
| St3gal2 | 5′- GCTCTCTTCGGGTGTGGTTCC -3′ | 5′- ATGCTGTGGTGCGAGTAGGTG -3′ |
| St3gal3 | 5′- CAGCAAGAAACCCAGACCAT -3′ | 5′- ATGAATGGCTCCGTCCATAG -3′ |
| St3gal4 | 5′- GCTCCTGTGGCTGGCTACG -3′ | 5′- GGGTCAAAGTGGGCCGACTC -3′ |
| St3gal5 | 5′- AGCCTCTTGGATATGCTGCCC -3′ | 5′- CGTTCCCAACAACCACACAGC -3′ |
| St3gal6 | 5′- ATGGTGGCATTCCCGTAGTA -3′ | 5′- AAGTGCACCTCGCTGGTTT -3′ |
| Ubiquitin | 5′- TGGCTATTAATTATTCGGTCTGCA -3′ | 5′- GCAAGTGGCTAGAGTGCAGAGTAA -3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matos, R.; Fonseca, K.L.; Mereiter, S.; Maceiras, A.R.; Gomes, J.; Vilaplana, C.; Gartner, F.; Rodrigues, P.N.S.; Reis, C.A.; Saraiva, M.; et al. Mycobacterium tuberculosis Infection Up-Regulates Sialyl Lewis X Expression in the Lung Epithelium. Microorganisms 2021, 9, 99. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010099
Matos R, Fonseca KL, Mereiter S, Maceiras AR, Gomes J, Vilaplana C, Gartner F, Rodrigues PNS, Reis CA, Saraiva M, et al. Mycobacterium tuberculosis Infection Up-Regulates Sialyl Lewis X Expression in the Lung Epithelium. Microorganisms. 2021; 9(1):99. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010099
Chicago/Turabian StyleMatos, Rita, Kaori L. Fonseca, Stefan Mereiter, Ana Raquel Maceiras, Joana Gomes, Cristina Vilaplana, Fátima Gartner, Pedro N. S. Rodrigues, Celso A. Reis, Margarida Saraiva, and et al. 2021. "Mycobacterium tuberculosis Infection Up-Regulates Sialyl Lewis X Expression in the Lung Epithelium" Microorganisms 9, no. 1: 99. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9010099