
Soil-Associated Bacillus Species: A Reservoir of Bioactive Compounds with Potential Therapeutic Activity against Human Pathogens

 , ,
, ,  , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collections and Isolation of Bacterial Isolates
2.2. Biochemical Characterization of Bacterial Isolates
2.2.1. Determination of the Amylolytic Activity
2.2.2. Determination of the Esterase Activity
2.2.3. Determination of the Proteolytic Activity
2.2.4. Determination of the Hemolytic Activity
2.3. PCR Amplification and Sequencing
2.4. Antimicrobial Susceptibility
2.5. Determination of the Antimicrobial Producing Activity
2.5.1. Media and Culture Conditions
2.5.2. Screening for Antimicrobial Activity
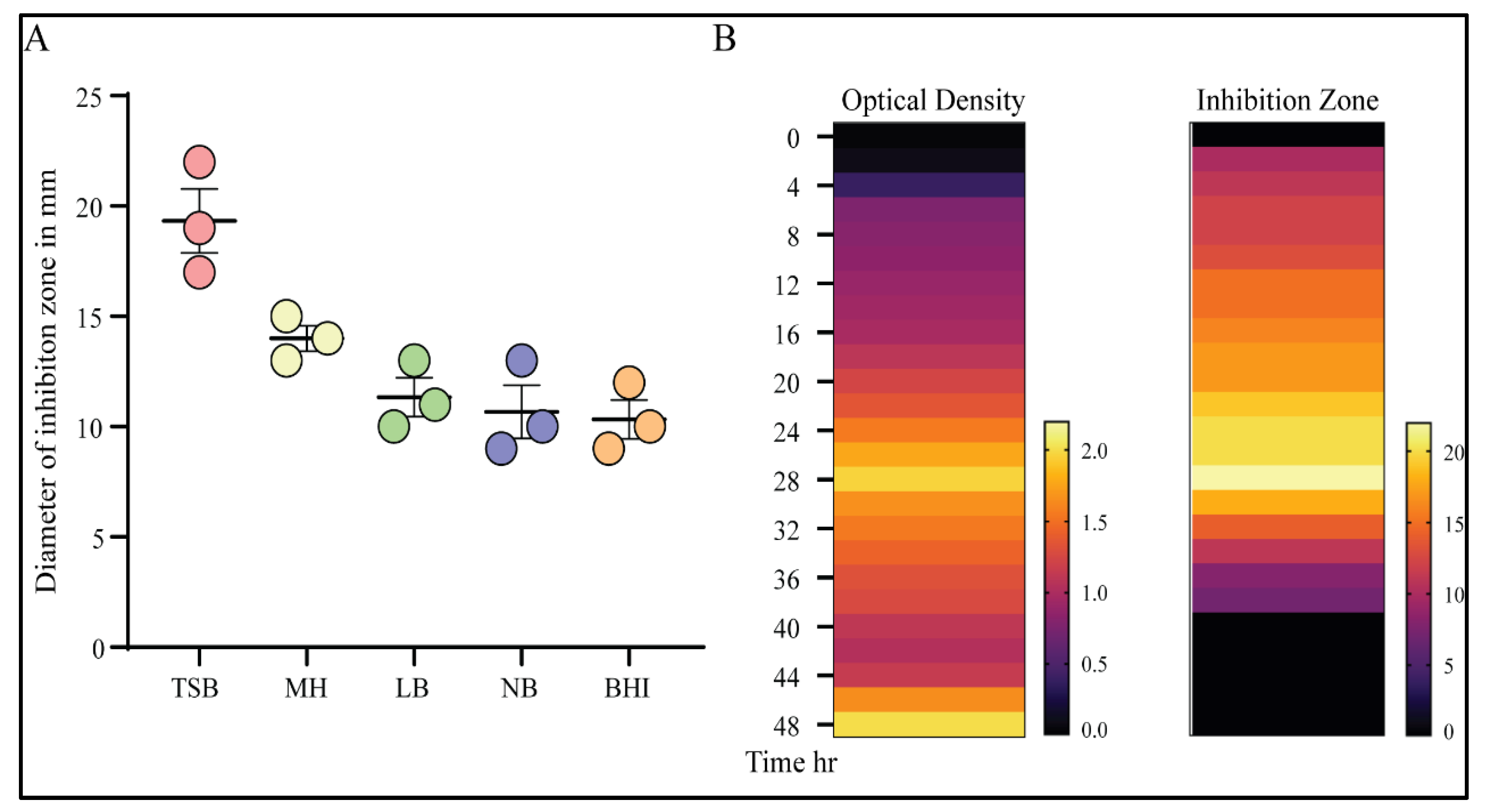
2.5.3. Effect of Medium Type for Optimum Antimicrobial Activity Production
2.5.4. Kinetic Production of Antimicrobial Activity
2.6. Purification and Biocompatibility Studies of the Isolated Antimicrobial Molecule
2.6.1. Partial Purification of the Antimicrobial Molecule
2.6.2. Effect of pH and Temperature
2.6.3. Effect of Surfactants
2.6.4. Effect of Metal Salts
2.7. Chemical Characterization
2.7.1. Ninhydrin Test: (Detection of Amino Acids)
2.7.2. Sudan III Test: (Detection of Lipid)
2.7.3. Molisch Test: (Detection of Carbohydrates)
2.8. In Vivo Study for the Antimicrobial Efficacy of Bacillus Extract
2.8.1. Animals
2.8.2. Induction of Keratitis
2.8.3. Experimental Design
2.9. Statistical Analysis
2.10. Phylogenetic Tree Construction
3. Results
3.1. Strain Biochemical Characterization and Antimicrobial Spectrum
3.2. 16S rRNA Gene Sequencing and Phylogenetic Analysis
3.3. Optimization of the Antimicrobial Productivity
3.4. Effect of pH and Temperature on the Efficacy of the Antimicrobial Compound
3.5. Effect of Surfactant and Metal Salts
3.6. Chemical Characterization and Qualitative Tests for Identification
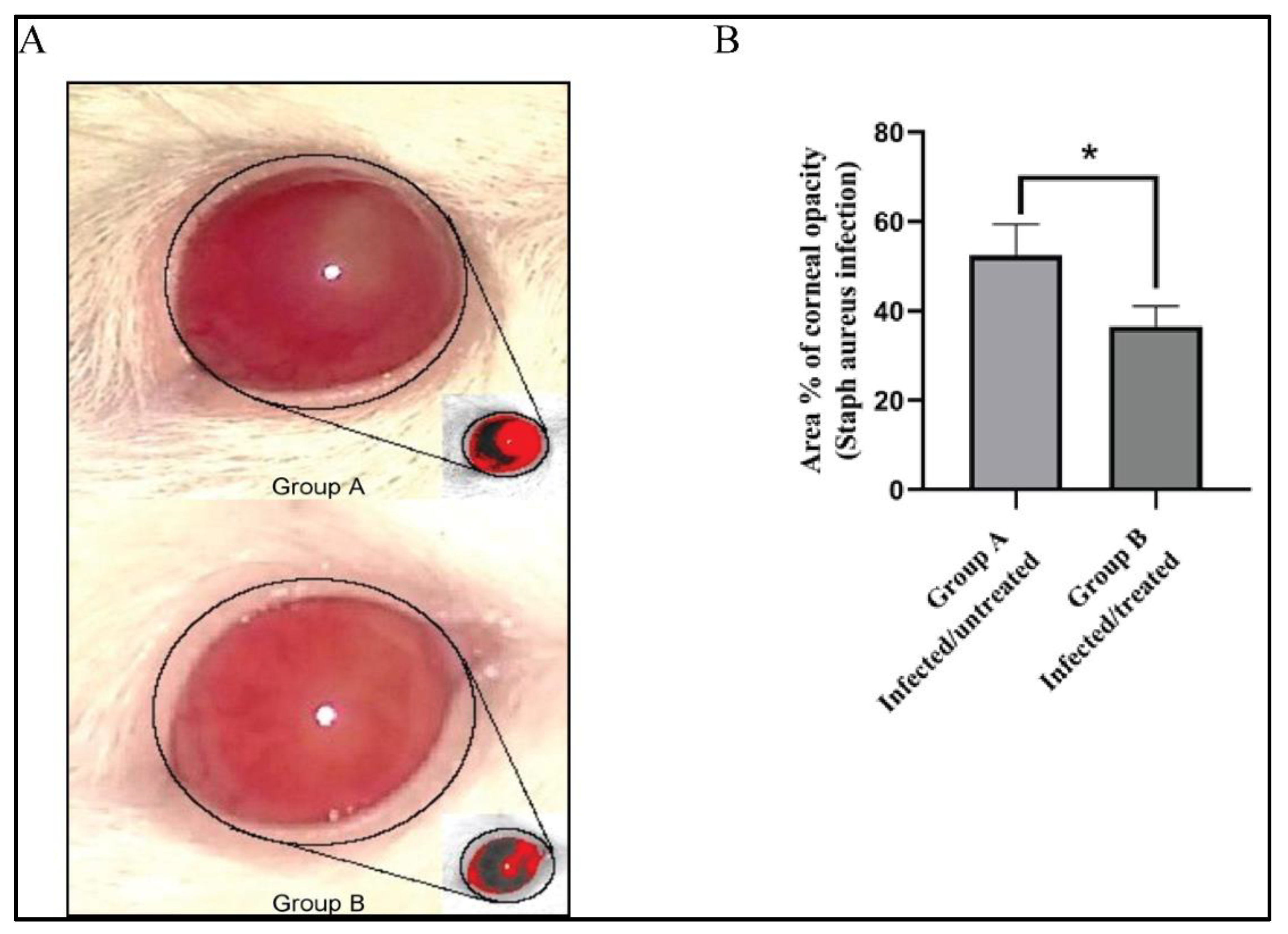
3.7. Effect of Bacillus subtilis Extract on the % Area of Corneal Opacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Maestre, F.T.; Singh, B.K.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Srividya, A.R.; Saritha, G.S.; Suresh, B. Study of the soil isolates for antimicrobial activity. Indian J. Pharm. Sci. 2008, 70, 812. [Google Scholar] [CrossRef] [Green Version]
- Chandra, N.; Kumar, S. Antibiotics producing soil microorganisms. In Antibiotics and Antibiotics Resistance Genes in Soils; Hashmi, M.Z., Strezov, V., Varma, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–18. ISBN 978-3-319-66259-6. [Google Scholar]
- Zinsser, H.; Joklik, W.K.; Willett, H.P.; Wilfert, C.M.; Amos, D.B. Zinsser Microbiology, 20th ed.; Appleton & Lange; Prentice Hall International: London, UK, 1992; ISBN 0838599834. [Google Scholar]
- Amin, A.; Khan, M.A.; Ehsanullah, M.; Haroon, U.; Azam, S.M.F.; Hameed, A. Production of peptide antibiotics by Bacillus sp. GU 057 indigenously isolated from saline soil. Braz. J. Microbiol. 2012, 43, 1340–1346. [Google Scholar] [CrossRef] [Green Version]
- Tabbene, O.; Ben Slimene, I.; Bouabdallah, F.; Mangoni, M.-L.; Urdaci, M.-C.; Limam, F. Production of anti-methicillin-resistant Staphylococcus activity from Bacillus subtilis sp. strain B38 newly isolated from soil. Appl. Biochem. Biotechnol. 2009, 157, 407–419. [Google Scholar] [CrossRef]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Stoica, R.-M.; Moscovici, M.; Tomulescu, C.; Cășărică, A.; Băbeanu, N.; Popa, O.; Kahraman, H.A. Antimicrobial compounds of the genus Bacillus: A review. Rom. Biotechnol. Lett. 2019, 24, 1111–1119. [Google Scholar] [CrossRef]
- Abriouel, H.; Franz, C.M.A.P.; Omar, N.B.; Gálvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, H.; Zhang, Y.; Duan, X.; Liu, J. Characterization of a bacteriocin-like substance produced from a novel isolated strain of Bacillus subtilis SLYY-3. J. Ocean Univ. China 2014, 13, 995–999. [Google Scholar] [CrossRef]
- Hover, B.M.; Kim, S.-H.; Katz, M.; Charlop-Powers, Z.; Owen, J.G.; Ternei, M.A.; Maniko, J.; Estrela, A.B.; Molina, H.; Park, S.; et al. Culture-independent discovery of the malacidins as calcium-dependent antibiotics with activity against multidrug-resistant Gram-positive pathogens. Nat. Microbiol. 2018, 3, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Chalasani, A.G.; Dhanarajan, G.; Nema, S.; Sen, R.; Roy, U. An antimicrobial metabolite from Bacillus sp.: Significant activity against pathogenic bacteria including multidrug-resistant clinical strains. Front. Microbiol. 2015, 6, 1335. [Google Scholar] [CrossRef] [Green Version]
- Valaperta, R.; Tejada, M.R.; Frigerio, M.; Moroni, A.; Ciulla, E.; Cioffi, S.; Capelli, P.; Costa, E. Staphylococcus aureus nosocomial infections: The role of a rapid and low-cost characterization for the establishment of a surveillance system. New Microbiol. 2010, 33, 223–232. [Google Scholar]
- Aqil, F.; Ahmad, I.; Owais, M. Evaluation of anti-methicillin-resistant Staphylococcus aureus (MRSA) activity and synergy of some bioactive plant extracts. Biotechnol. J. 2006, 1, 1093–1102. [Google Scholar] [CrossRef]
- Ida, T.; Okamoto, R.; Shimauchi, C.; Okubo, T.; Kuga, A.; Inoue, M. Identification of aminoglycoside-modifying enzymes by susceptibility testing: Epidemiology of methicillin-resistant Staphylococcus aureus in Japan. J. Clin. Microbiol. 2001, 39, 3115–3121. [Google Scholar] [CrossRef] [Green Version]
- Hashem, R.A.; Yassin, A.S.; Zedan, H.H.; Amin, M.A. Fluoroquinolone resistant mechanisms in methicillin-resistant Staphylococcus aureus clinical isolates in Cairo, Egypt. J. Infect. Dev. Ctries. 2013, 7, 796–803. [Google Scholar] [CrossRef] [Green Version]
- Guignard, B.; Entenza, J.M.; Moreillon, P. Beta-lactams against methicillin-resistant Staphylococcus aureus. Curr. Opin. Pharmacol. 2005, 5, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Gardete, S.; Tomasz, A. Mechanisms of vancomycin resistance in Staphylococcus aureus. J. Clin. Investig. 2014, 124, 2836–2840. [Google Scholar] [CrossRef]
- Siberry, G.K.; Tekle, T.; Carroll, K.; Dick, J. Failure of clindamycin treatment of methicillin-resistant Staphylococcus aureus expressing inducible clindamycin resistance in vitro. Clin. Infect. Dis. 2003, 37, 1257–1260. [Google Scholar] [CrossRef]
- Hsieh, Y.-C.; Lin, Y.-C.; Huang, Y.-C. Vancomycin, teicoplanin, daptomycin, and linezolid MIC creep in methicillin-resistant Staphylococcus aureus is associated with clonality. Medicine 2016, 95, e5060. [Google Scholar] [CrossRef] [PubMed]
- Jian, Y.; Lv, H.; Liu, J.; Huang, Q.; Liu, Y.; Liu, Q.; Li, M. Dynamic changes of Staphylococcus aureus susceptibility to vancomycin, teicoplanin, and linezolid in a Central Teaching Hospital in Shanghai, China, 2008–2018. Front. Microbiol. 2020, 11, 908. [Google Scholar] [CrossRef] [PubMed]
- Tarai, B.; Das, P.; Kumar, D. Recurrent challenges for clinicians: Emergence of methicillin-resistant Staphylococcus aureus, vancomycin resistance, and current treatment options. J. Lab. Physicians 2013, 5, 71–78. [Google Scholar] [CrossRef]
- Vaudaux, P.; Francois, P.; Berger-Bächi, B.; Lew, D.P. In vivo emergence of subpopulations expressing teicoplanin or vancomycin resistance phenotypes in a glycopeptide-susceptible, methicillin-resistant strain of Staphylococcus aureus. J. Antimicrob. Chemother. 2001, 47, 163–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.M.; Seo, C.; Ji, M.; Paik, M.-J.; Myung, S.-W.; Kim, J. Effective soil extraction method for cultivating previously uncultured soil bacteria. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chilcott, C.N.; Wigley, P.J. Isolation and toxicity of Bacillus thuringiensis from soil and insect habitats in New Zealand. J. Invertebr. Pathol. 1993, 61, 244–247. [Google Scholar] [CrossRef]
- Marinho, S.A.; Teixeira, A.B.; Santos, O.S.; Cazanova, R.F.; Ferreira, C.A.S.; Cherubini, K.; de Oliveira, S.D. Identification of Candida spp. by phenotypic tests and PCR. Braz. J. Microbiol. 2010, 41, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powthong, P.; Suntornthiticharoen, P. Antimicrobial and enzyme activity produced by Bacillus SPP. isolated from soil. Int. J. Pharm. Pharm. Sci. 2017, 9, 205. [Google Scholar] [CrossRef] [Green Version]
- Moshafi, M.H.; Forootanfar, H.; Ameri, A.; Shakibaie, M.; Dehghan-Noudeh, G.; Razavi, M. Antimicrobial activity of Bacillus sp. strain FAS1 isolated from soil. Pak. J. Pharm. Sci. 2011, 24, 269–275. [Google Scholar]
- Gerhardt, P. Methods for General and Molecular Bacteriology; American Society for Microbiology: Washington, DC, USA, 1994; ISBN 1555810489. [Google Scholar]
- Sun, J.; Yang, M.; Sreevatsan, S.; Davies, P.R. Prevalence and characterization of Staphylococcus aureus in growing pigs in the USA. PLoS ONE 2015, 10, e0143670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.-Y.; Zou, B.; Yan, Y.-W.; Jeon, C.O.; Li, M.; Cai, M.; Quan, Z.-X. Design of targeted primers based on 16S rRNA sequences in meta-transcriptomic datasets and identification of a novel taxonomic group in the Asgard archaea. BMC Microbiol. 2020, 20. [Google Scholar] [CrossRef]
- Yilmaz, M.; Soran, H.; Beyatli, Y. Antimicrobial activities of some Bacillus spp. strains isolated from the soil. Microbiol. Res. 2006, 161, 127–131. [Google Scholar] [CrossRef]
- Yu, G.Y.; Sinclair, J.B.; Hartman, G.L.; Bertagnolli, B.L. Production of iturin A by Bacillus amyloliquefaciens suppressing Rhizoctonia solani. Soil Biol. Biochem. 2002, 34, 955–963. [Google Scholar] [CrossRef]
- Hernández, D.; Cardell, E.; Zárate, V. Antimicrobial activity of lactic acid bacteria isolated from Tenerife cheese: Initial characterization of plantaricin TF711, a bacteriocin-like substance produced by Lactobacillus plantarum TF711. J. Appl. Microbiol. 2005, 99, 77–84. [Google Scholar] [CrossRef]
- Ramachandran, R.; Chalasani, A.G.; Lal, R.; Roy, U. A broad-spectrum antimicrobial activity of Bacillus subtilis RLID 12.1. Sci. World J. 2014, 2014, 968487. [Google Scholar] [CrossRef] [Green Version]
- Kayalvizhi, N.; Gunasekaran, P. Purification and characterization of a novel broad-spectrum bacteriocin from Bacillus licheniformis MKU3. Biotechnol. Bioproc. E 2010, 15, 365–370. [Google Scholar] [CrossRef]
- Das, S. Recent applications of ninhydrin in multicomponent reactions. RSC Adv. 2020, 10, 18875–18906. [Google Scholar] [CrossRef]
- Masamune, O.; Takahashi, T.; Nagasaki, A.; Iwabuchi, J.; Ishikawa, M. Diagnostic significance of the sudan III staining for fecal fat. Tohoku J. Exp. Med. 1977, 122, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Foulger, J.H. The use of the Molisch (α-naphthol) reactions in the study of sugars in biological fluids. J. Biol. Chem. 1931, 92, 345–353. [Google Scholar] [CrossRef]
- Saber, S.; Ghanim, A.M.H.; El-Ahwany, E.; El-Kader, E.M.A. Novel complementary antitumour effects of celastrol and metformin by targeting IκBκB, apoptosis and NLRP3 inflammasome activation in diethylnitrosamine-induced murine hepatocarcinogenesis. Cancer Chemother. Pharmacol. 2020, 85, 331–343. [Google Scholar] [CrossRef]
- Saber, S.; Khalil, R.M.; Abdo, W.S.; Nassif, D.; El-Ahwany, E. Olmesartan ameliorates chemically-induced ulcerative colitis in rats via modulating NFκB and Nrf-2/HO-1 signaling crosstalk. Toxicol. Appl. Pharmacol. 2019, 364, 120–132. [Google Scholar] [CrossRef] [PubMed]
- Desper, R. Theoretical foundation of the balanced minimum evolution method of phylogenetic inference and its relationship to weighted least-squares tree fitting. Mol. Biol. Evol. 2003, 21, 587–598. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.-H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Al-Ansari, M.; Kalaiyarasi, M.; Almalki, M.A.; Vijayaraghavan, P. Optimization of medium components for the production of antimicrobial and anticancer secondary metabolites from Streptomyces sp. AS11 isolated from the marine environment. J. King Saud Univ. Sci. 2020, 32, 1993–1998. [Google Scholar] [CrossRef]
- Friedman, M. Applications of the ninhydrin reaction for analysis of amino acids, peptides, and proteins to agricultural and biomedical sciences. J. Agric. Food Chem. 2004, 52, 385–406. [Google Scholar] [CrossRef] [PubMed]
- Devor, A.W. Carbohydrate tests using sulfonated α-naphthol. J. Am. Chem. Soc. 1950, 72, 2008–2012. [Google Scholar] [CrossRef]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with sudan red 7B or fluoral yellow 088 in polyethylene glycol-glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Nasr, M.; Teiama, M.; Ismail, A.; Ebada, A.; Saber, S. In vitro and in vivo evaluation of cubosomal nanoparticles as an ocular delivery system for fluconazole in treatment of keratomycosis. Drug Deliv. Transl. Res. 2020, 10, 1841–1852. [Google Scholar] [CrossRef]
- El-Baz, A.M.; Yahya, G.; Mansour, B.; El-Sokkary, M.M.A.; Alshaman, R.; Alattar, A.; El-Ganiny, A.M. The link between oc-currence of class I integron and acquired aminoglycoside resistance in clinical MRSA isolates. Antibiotics 2021, 10, 488. [Google Scholar] [CrossRef]
- Yang, L.; Wang, K.; Li, H.; Denstedt, J.D.; Cadieux, P.A. The influence of urinary pH on antibiotic efficacy against bacterial uropathogens. Urology 2014, 84, 731.e1–731.e7. [Google Scholar] [CrossRef]
- Serafin, A.; Stańczak, A. The complexes of metal ions with fluoroquinolones. Russ. J. Coord. Chem. 2009, 35, 81–95. [Google Scholar] [CrossRef]
- Berthon, G.; Brion, M.; Lambs, L. Metal ion-tetracycline interactions in biological fluids. 2. potentiometric study of magnesium complexes with tetracycline, oxytetracycline, doxycycline, and minocycline, and discussion of their possible influence on the bioavailability of these antibiotics in blood plasma. J. Inorg. Biochem. 1983, 19, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Schwameis, R.; Erdogan-Yildirim, Z.; Manafi, M.; Zeitlinger, M.A.; Strommer, S.; Sauermann, R. Effect of pulmonary surfactant on antimicrobial activity in vitro. Antimicrob. Agents Chemother. 2013, 57, 5151–5154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannanov, R.N.; Sattarova, R.K. Antibiotics produced by Bacillus bacteria. Chem. Nat. Compd. 2001, 37, 117–123. [Google Scholar] [CrossRef]
- Khalaf, E.M.; Raizada, M.N. Draft genome sequence of Bacillus sp. strain EKM601B (phylum firmicutes), living inside the seeds of Luffa acutangula (chinese okra). Microbiol. Resour. Announc. 2020, 9, e00180-20. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, E.M.; Raizada, M.N. Draft genome sequences of Bacillus and Paenibacillus species isolated from seeds of Citrullus lanata (watermelon), Cucurbita moschata (butternut squash), and Cucurbita pepo L. var. pepo L. (pumpkin). Microbiol. Resour. Announc. 2020, 9, e00727-20. [Google Scholar] [CrossRef]
- Binda, E.; Marinelli, F.; Marcone, G. Old and new glycopeptide antibiotics: Action and resistance. Antibiotics 2014, 3, 572–594. [Google Scholar] [CrossRef] [Green Version]
- Garau, J.; Bouza, E.; Chastre, J.; Gudiol, F.; Harbarth, S. Management of methicillin-resistant Staphylococcus aureus infections. Clin. Microbiol. Infect. 2009, 15, 125–136. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Amplicon Size | Target | Annealing Temperature | Ref. |
|---|---|---|---|---|---|
| mecA-F | GTAGAAATGACTGAACGTCCGATAA | 310 bp | mecA gene | 52 °C | [31] |
| mecA-R | CCAATTCCACATTGTTCGGTCTAA | ||||
| 16S rRNA-F | AAGTCGAACGGACACGCAT | 1492 bp | 16S-rRNA | 52 °C | [32] |
| 16S rRNA-R | TACGGATACCTTGTTACGACTT |
| Isolate No. | Indicator Organisms | Media | Zone of Inhibition (Diameter in mm) |
|---|---|---|---|
| 1 | Staphylococcus aureus ATCC 29312 | MH | 17 |
| 2 | Staphylococcus epidermidis * | MH | 14 |
| 3 | Methicillin resistant Staphylococcus aureus 1 * | MH | 17 |
| 4 | Methicillin resistant Staphylococcus aureus 2 * | MH | 16 |
| 5 | Methicillin resistant Staphylococcus aureus 3 * | MH | 15 |
| 6 | Methicillin resistant Staphylococcus aureus 4 * | MH | 12 |
| 7 | Candida albicans * | SDA | 30 |
| 8 | Escherichia coli * | MH | No zone |
| 9 | Escherichia coli ATCC 12435 | MH | No zone |
| 10 | Klebsiella pneumonia * | MH | No zone |
| 11 | Pseudomonas aeruginosa * | MH | No zone |
| 12 | Proteus mirabilis * | MH | No zone |
| Group | Ketamine/Xylazine | Methyl Prednisolone | MRSA Infection | Bacillus subtilis Extract |
|---|---|---|---|---|
| Group A | + | + | + | - |
| Group B | + | + | + | + |
| Treatment | Antimicrobial Activity% of Control Value |
|---|---|
| Untreated (Control) | 100 ± 4.26 |
| pH | |
| 2.0 | 77.203 ± 6.5 * |
| 3.0 | 77.31 ± 5.34 * |
| 4.0 | 77.42 ± 5,66 * |
| 5.0 | 77.1 ± 6.43 * |
| 6.0 | 77.22 ± 4.38 * |
| 7.0 | 100.36 ± 5.32 |
| 8.0 | 72.41 ± 6.2 * |
| 9.0 | 66.33 ± 5.87 * |
| 10.0 | 0 * |
| 11.0 | 0 * |
| 12.0 | 0 * |
| Temperature | |
| 40 °C | 100 ± 3.23 |
| 50 °C | 100.36 ± 6.32 |
| 60 °C | 100.38 ± 4.7 |
| 70 °C | 100.43 ± 5.32 |
| 80 °C | 100.4 ± 6.47 |
| 90 °C | 94.44 ± 5.2 |
| 100 °C | 94.22 ± 4.29 |
| Autoclaving 121 °C for 20 min | 94.34 ± 4.82 |
| Treatment | Antimicrobial Activity |
|---|---|
| Surfactants | |
| Sodium dodecyl sulfate (SDS) | + |
| Tween 80 | + |
| Metal Salts | |
| FeSO4 | + |
| MgSO4 | + |
| AgNO3 | + |
| ZnSO4 | + |
| CdCl2 | + |
| CuSO4 | + |
| CaCl2 | + |
| NiCl2 | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yahya, G.; Ebada, A.; Khalaf, E.M.; Mansour, B.; Nouh, N.A.; Mosbah, R.A.; Saber, S.; Moustafa, M.; Negm, S.; El-Sokkary, M.M.A.; et al. Soil-Associated Bacillus Species: A Reservoir of Bioactive Compounds with Potential Therapeutic Activity against Human Pathogens. Microorganisms 2021, 9, 1131. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061131
Yahya G, Ebada A, Khalaf EM, Mansour B, Nouh NA, Mosbah RA, Saber S, Moustafa M, Negm S, El-Sokkary MMA, et al. Soil-Associated Bacillus Species: A Reservoir of Bioactive Compounds with Potential Therapeutic Activity against Human Pathogens. Microorganisms. 2021; 9(6):1131. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061131
Chicago/Turabian StyleYahya, Galal, Asmaa Ebada, Eman M. Khalaf, Basem Mansour, Nehal A. Nouh, Rasha A. Mosbah, Sameh Saber, Mahmoud Moustafa, Sally Negm, Mohamed M. A. El-Sokkary, and et al. 2021. "Soil-Associated Bacillus Species: A Reservoir of Bioactive Compounds with Potential Therapeutic Activity against Human Pathogens" Microorganisms 9, no. 6: 1131. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9061131