
Veronica Plants—Drifting from Farm to Traditional Healing, Food Application, and Phytopharmacology

 , ,
, , 
 ,
, 
 , , , ,
, , , ,  ,
,  and
and
Abstract
:1. Introduction
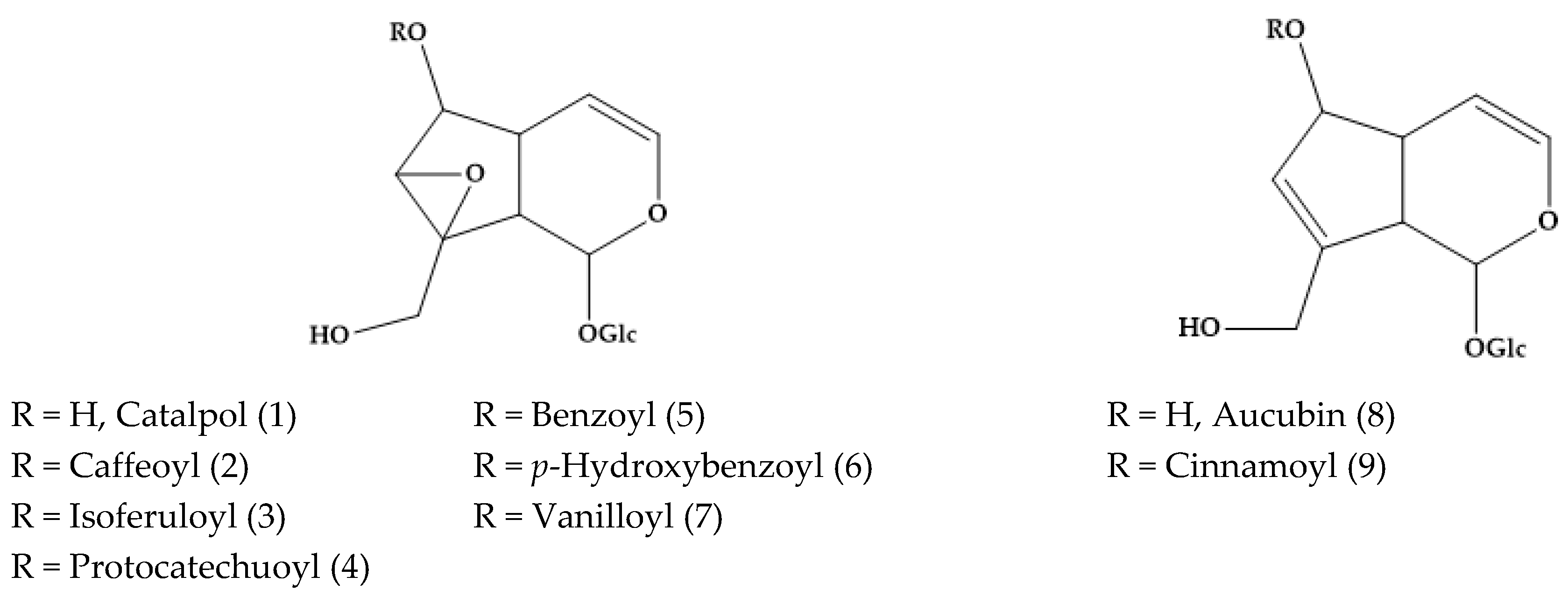
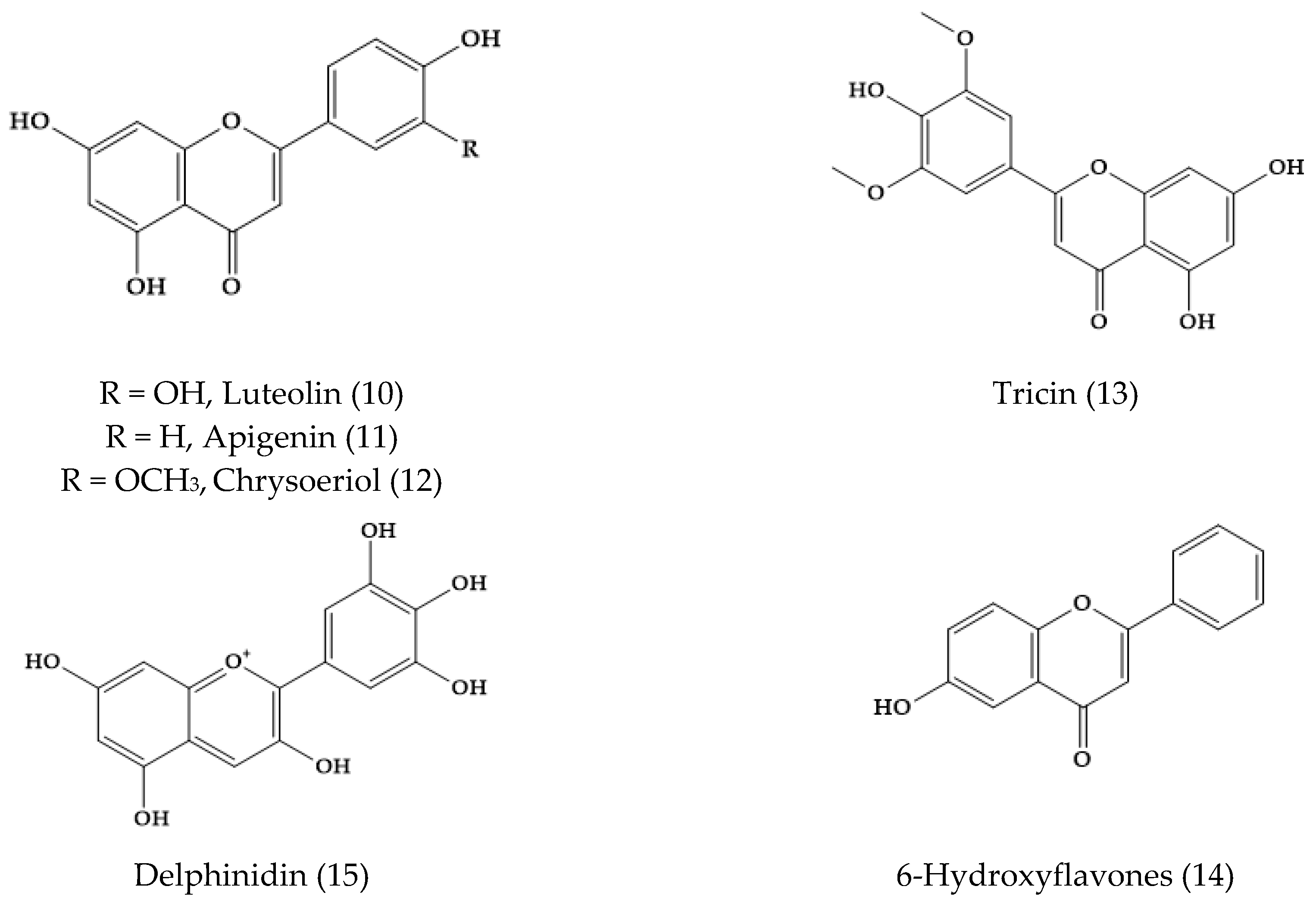
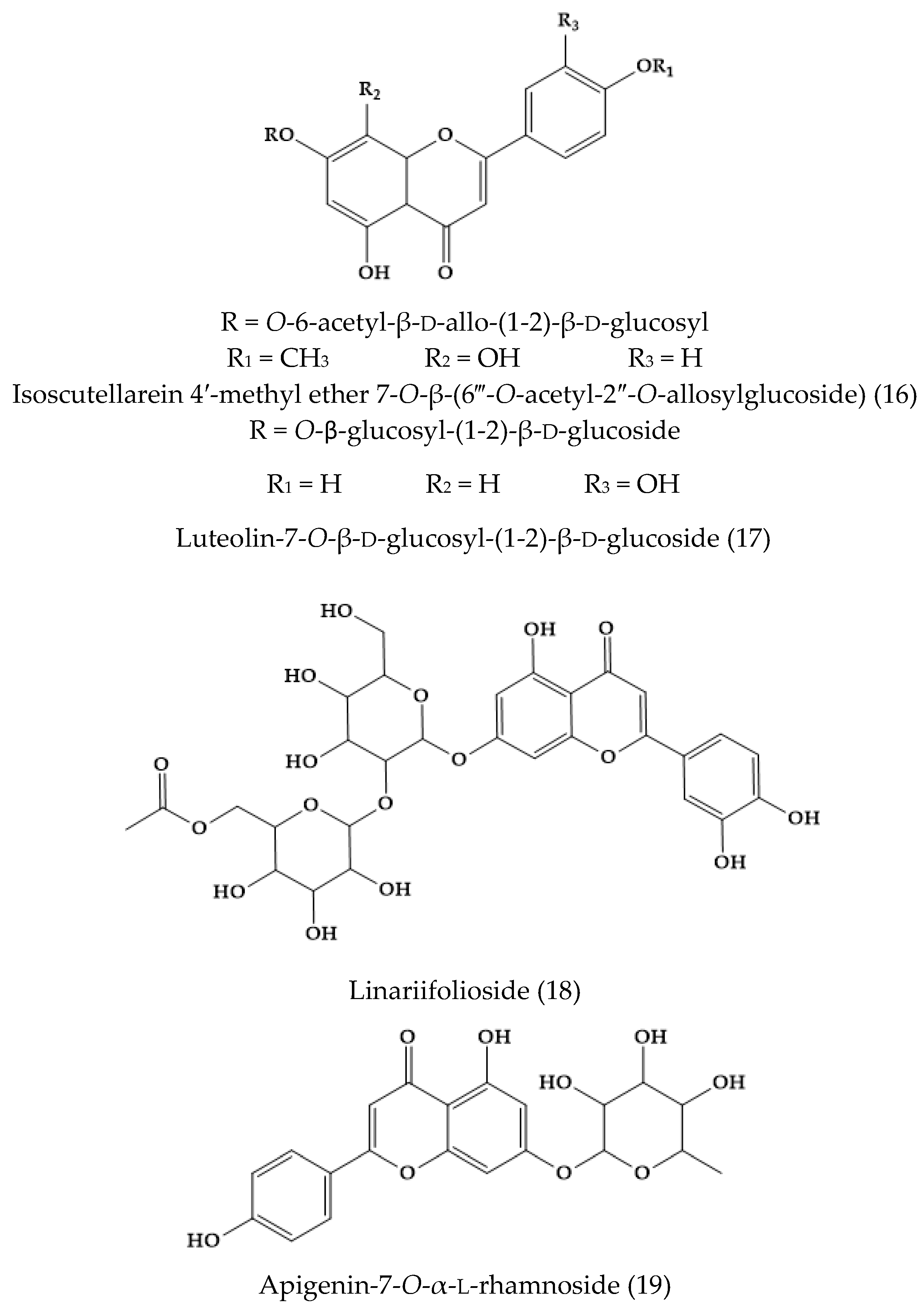
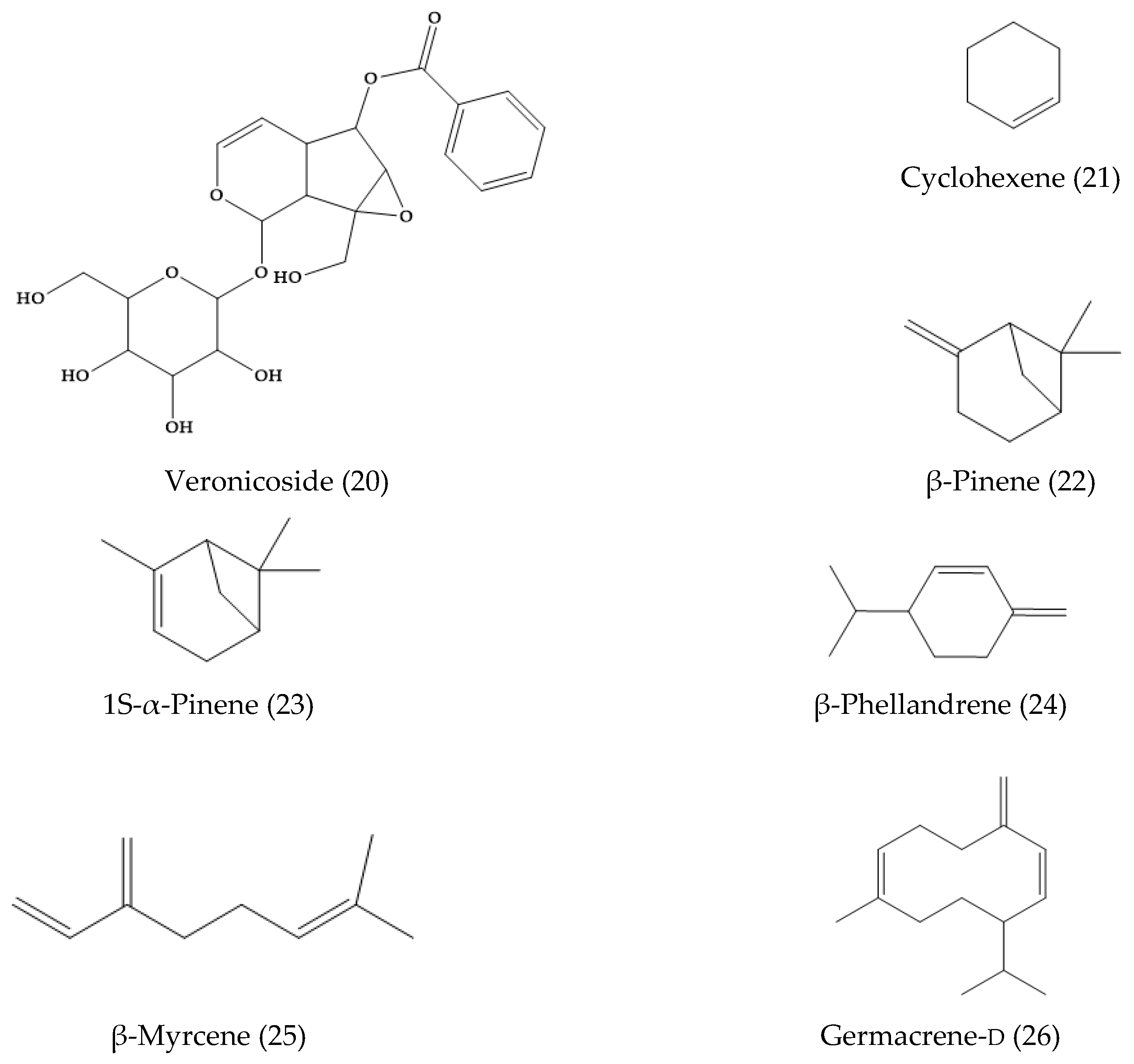
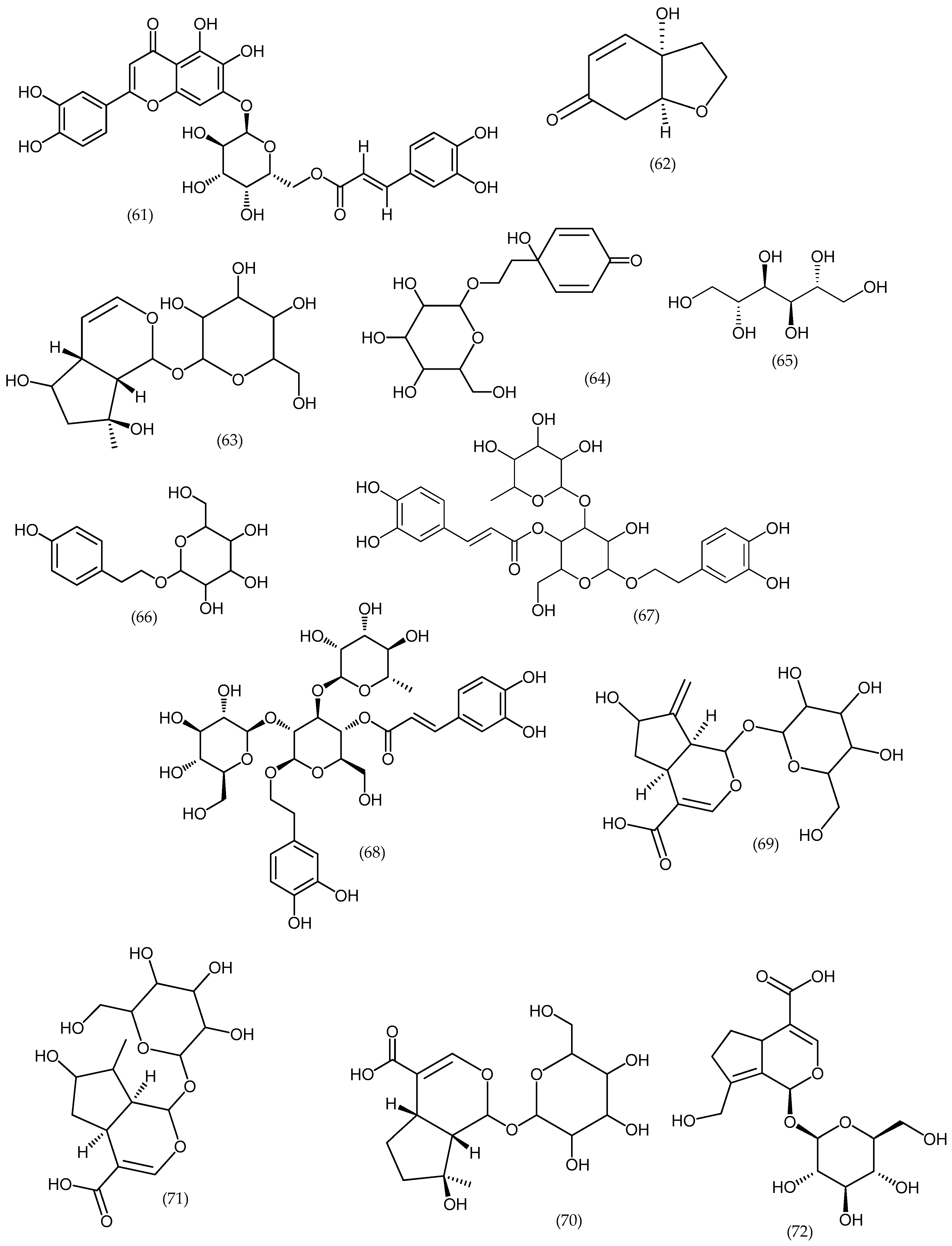
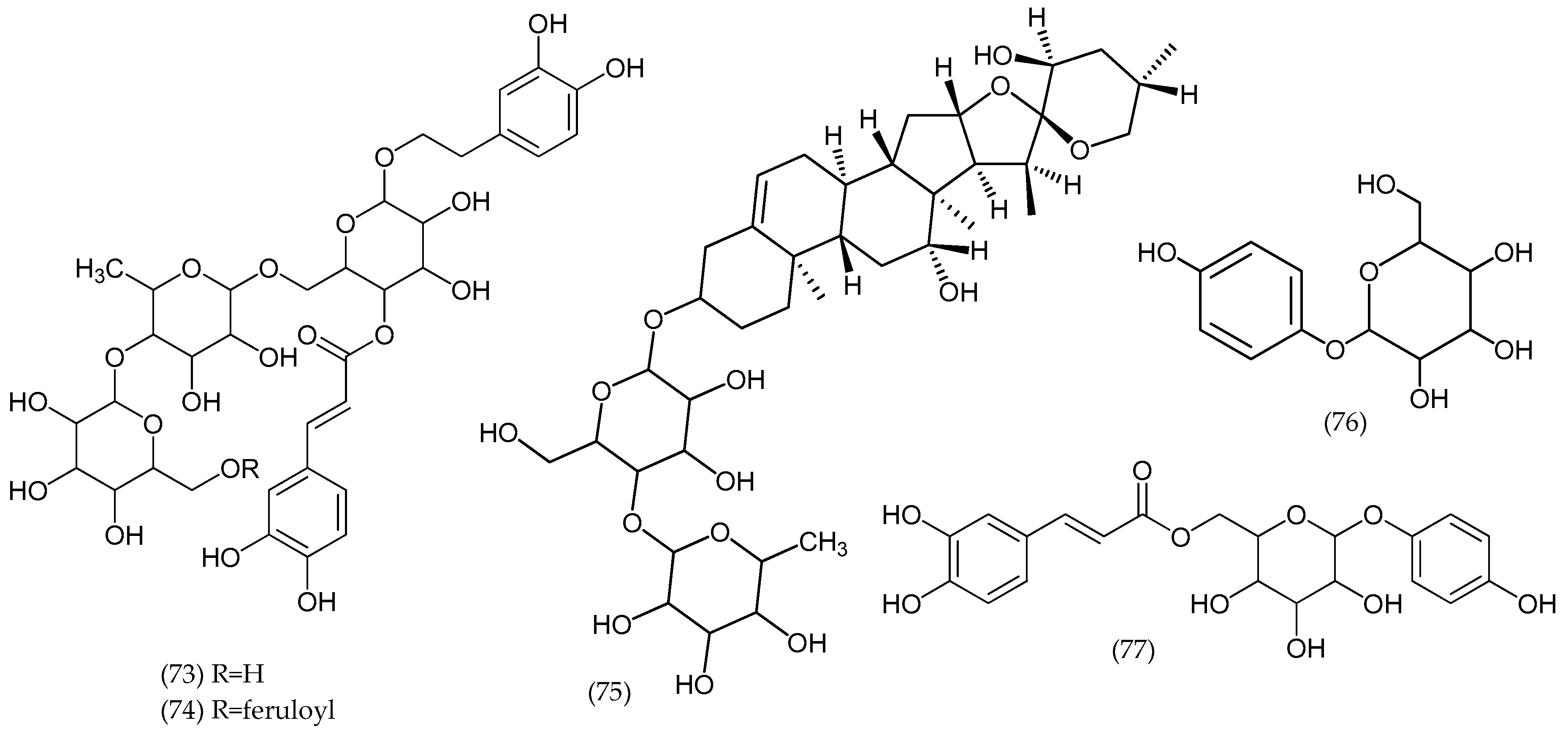
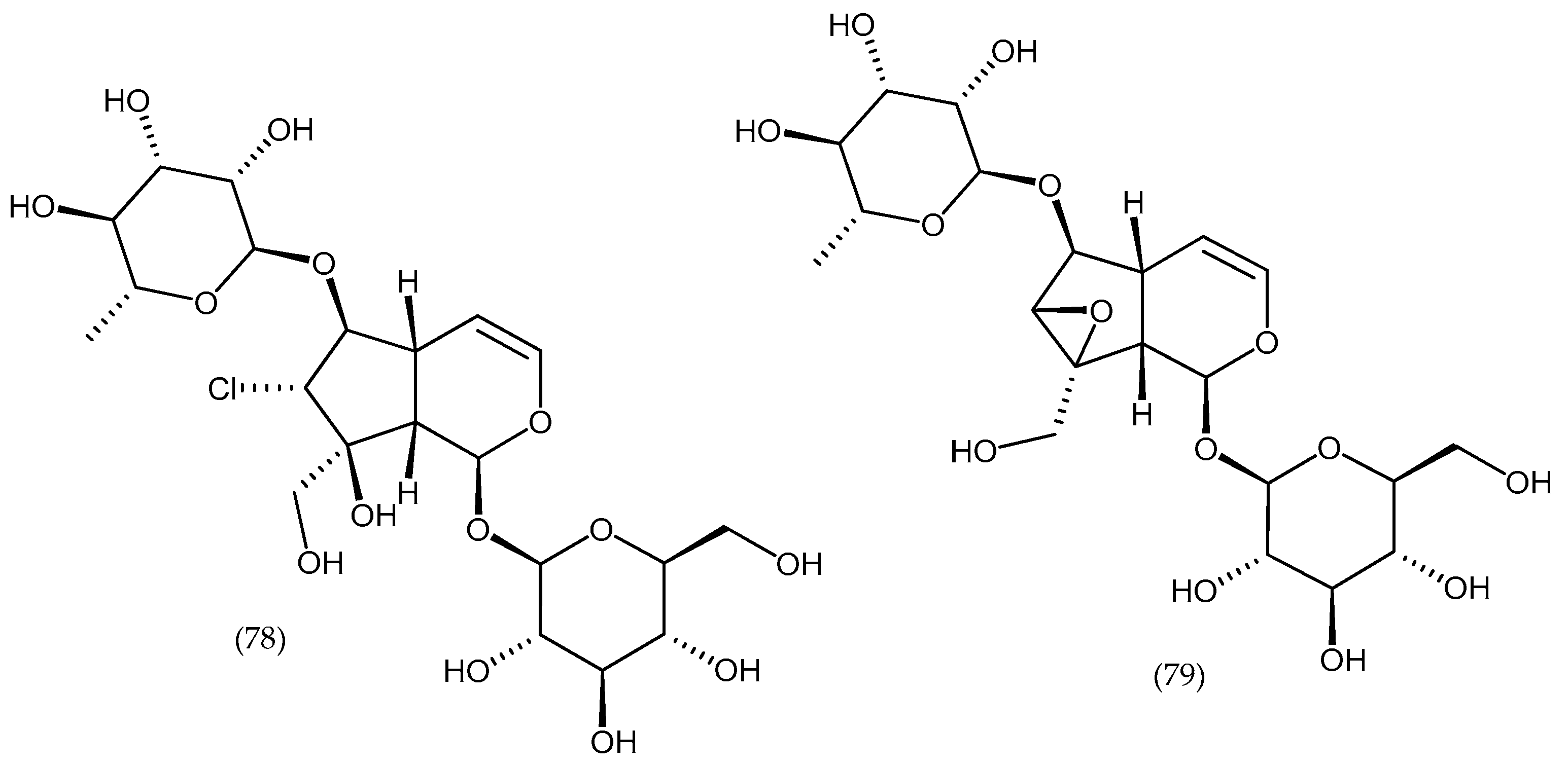
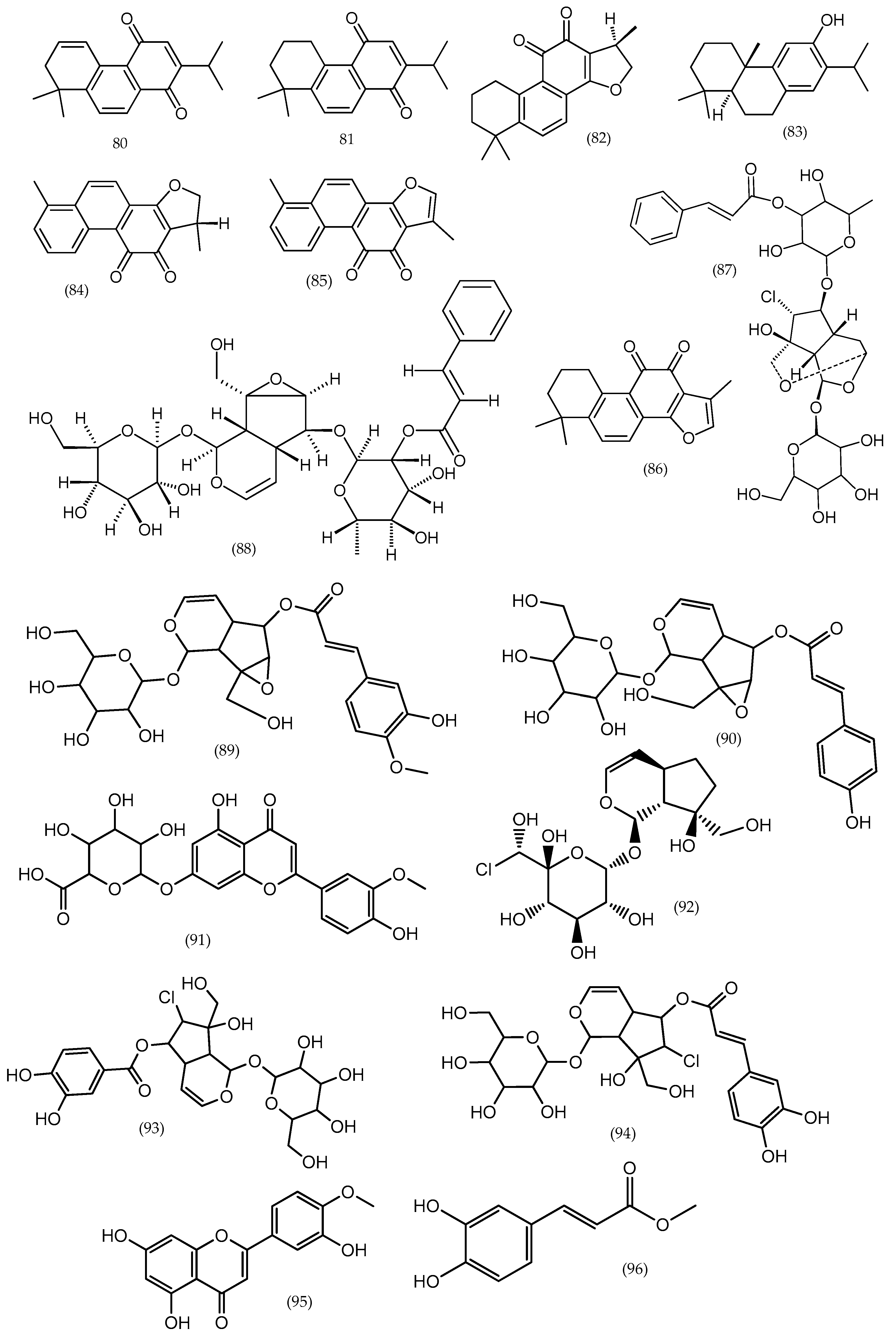
2. Phytochemical Characterization of Veronica Plants
2.1. Veronica filiformis
2.2. Veronica linariifolia Pall. ex Link
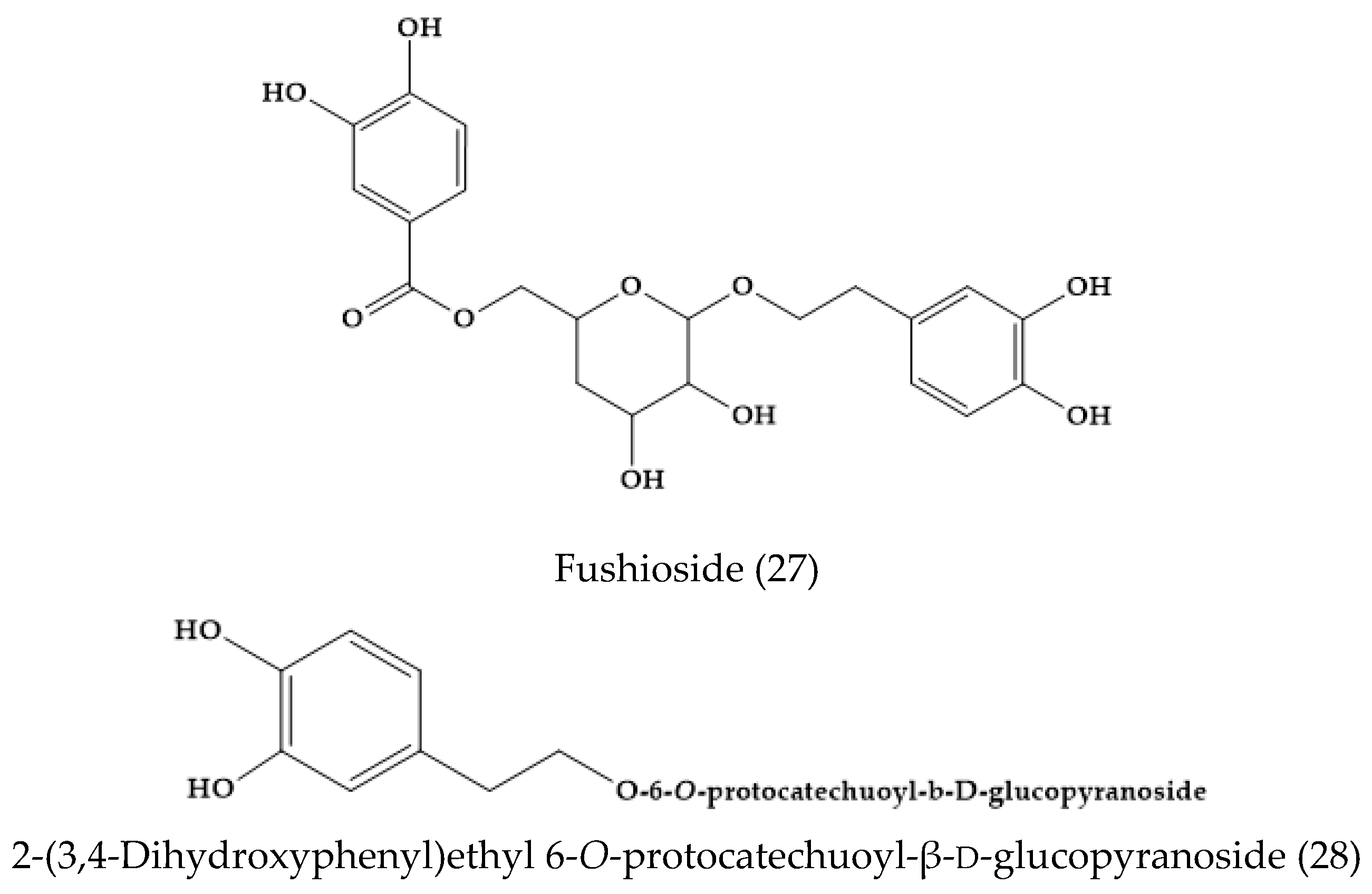
2.3. Veronica fushii
2.4. Veronica cymbalaria Bodard
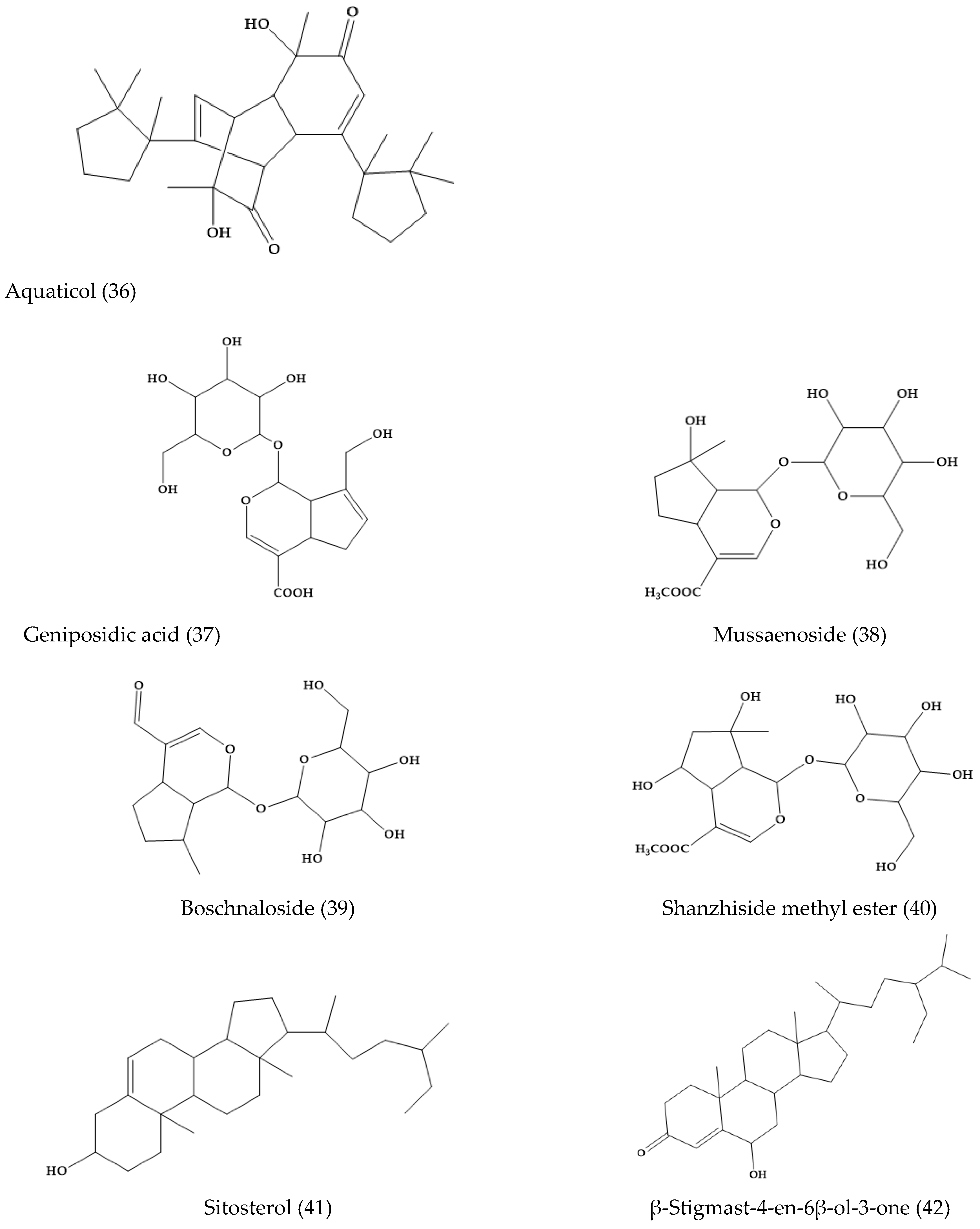
2.5. Veronica anagallis-aquatica L.
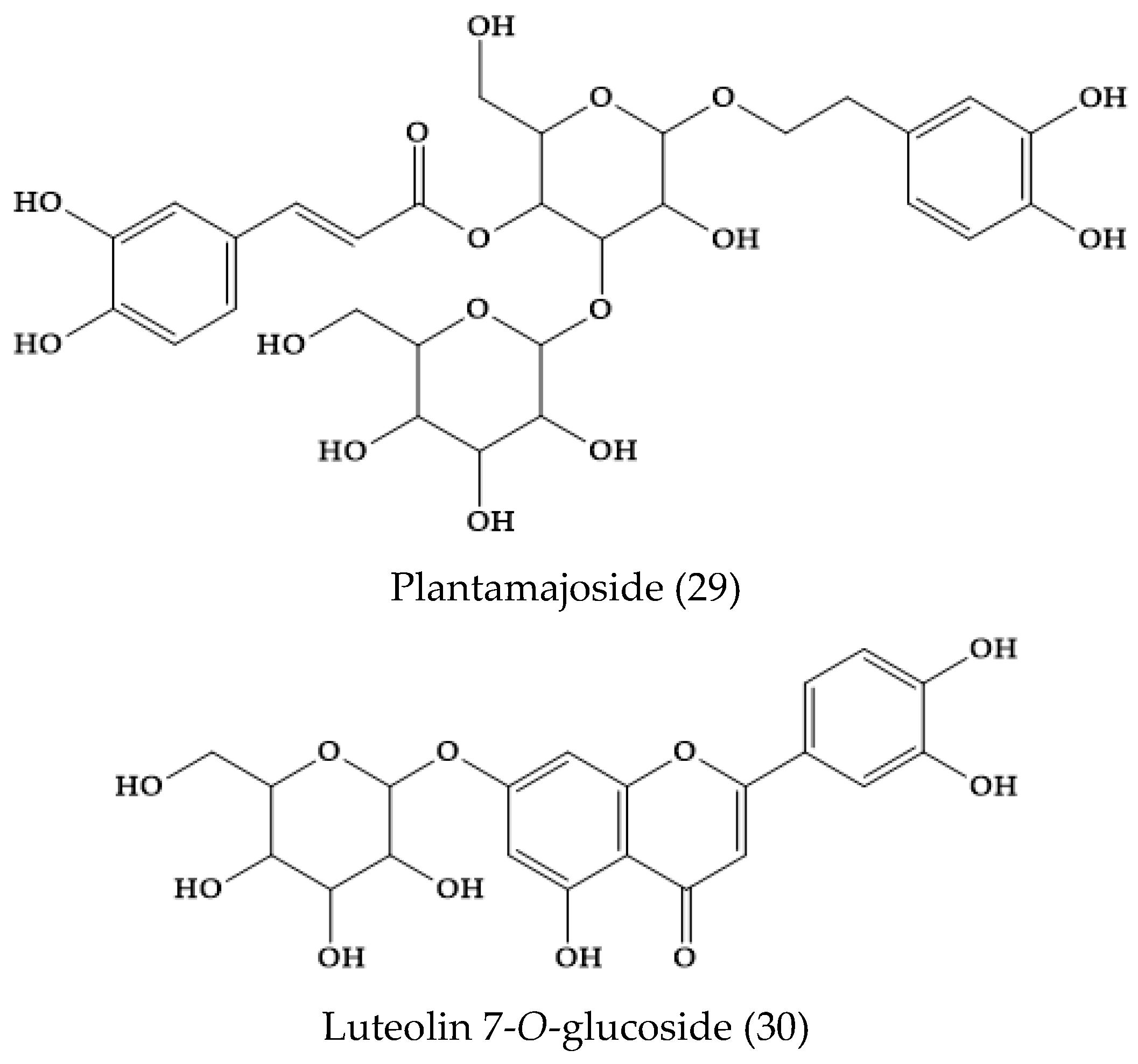
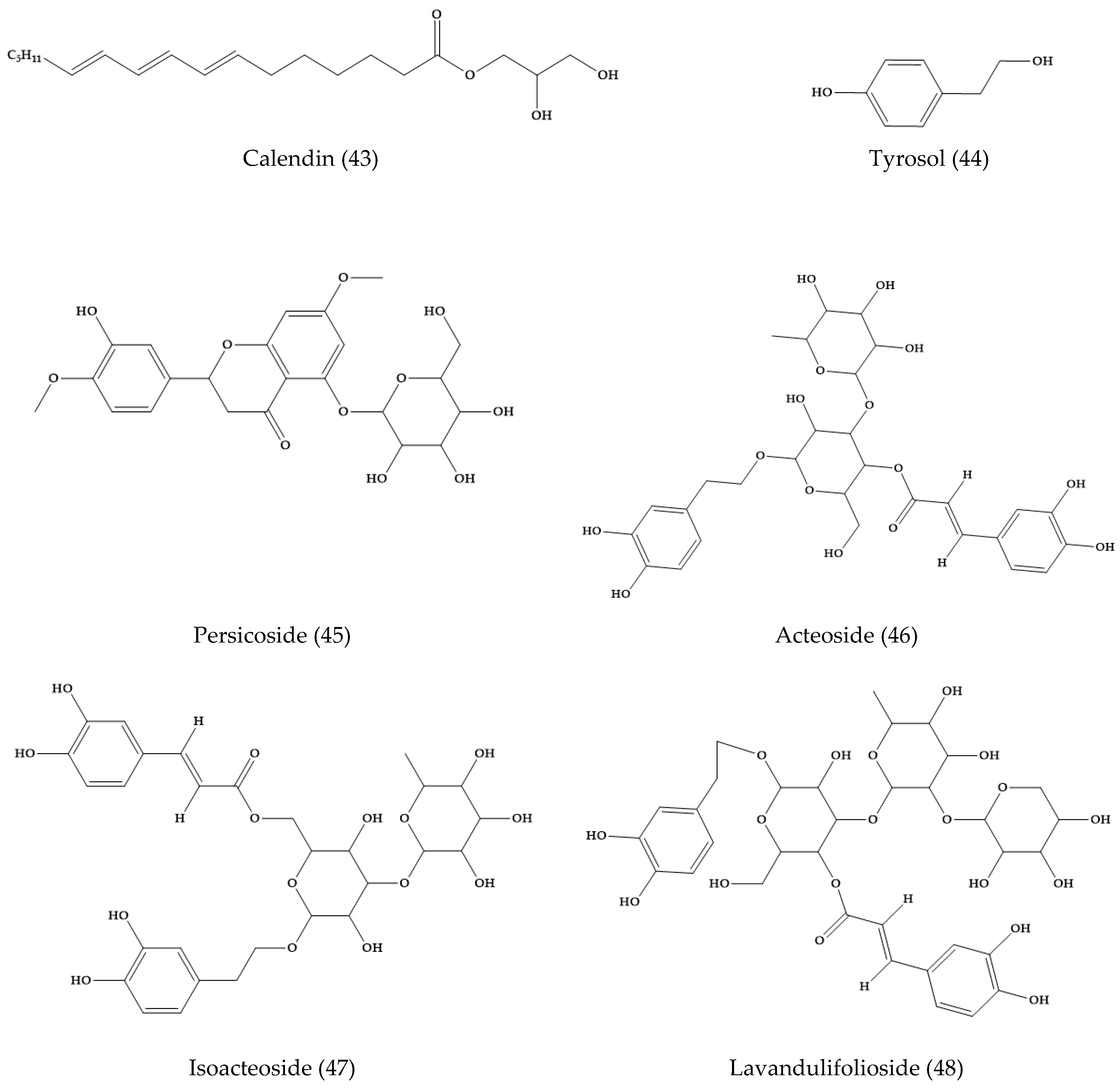
2.6. Veronica persica
2.7. Veronica longifolia L. and Veronica liwanensis K. Koch
2.8. Veronica orientalis Mill.
2.9. Veronica thymoides P. H. Davis
2.10. Veronica arvensis
2.11. Veronica turrilliana Stoj. & Stef.
2.12. Veronica cuneifolia D. Don
2.13. Veronica derwentiana Andrews and Veronica catarractae G. Forst.
2.14. Veronica sibirica L.
2.15. Veronica Peregrina L.
2.16. Veronica montana L., Veronica polita Fr., and Veronica spuria L.
2.17. Veronica spicata
2.18. Veronica officinalis
2.19. Veronica ciliata Fisch.
2.20. Veronica rosea Desf.
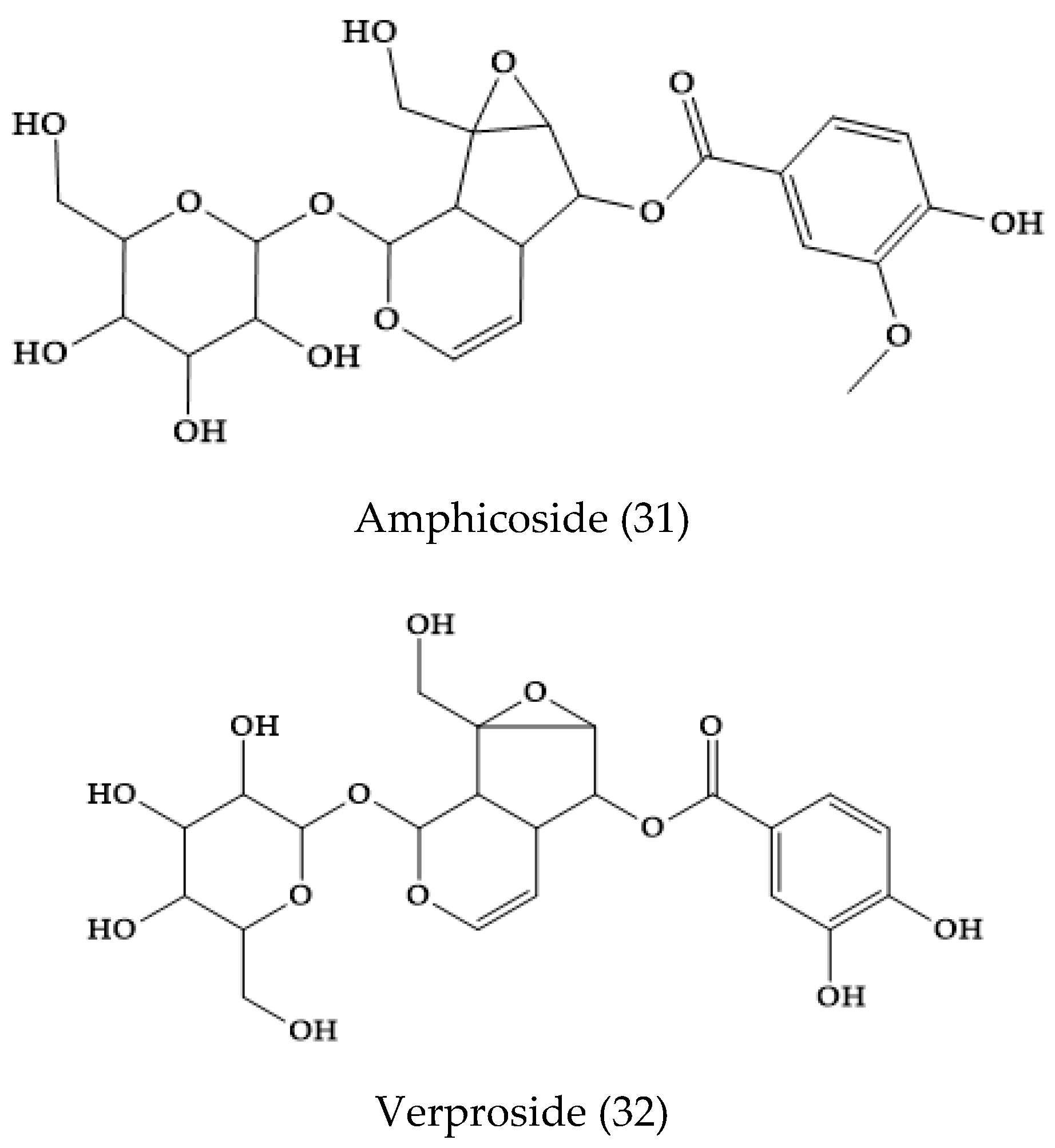
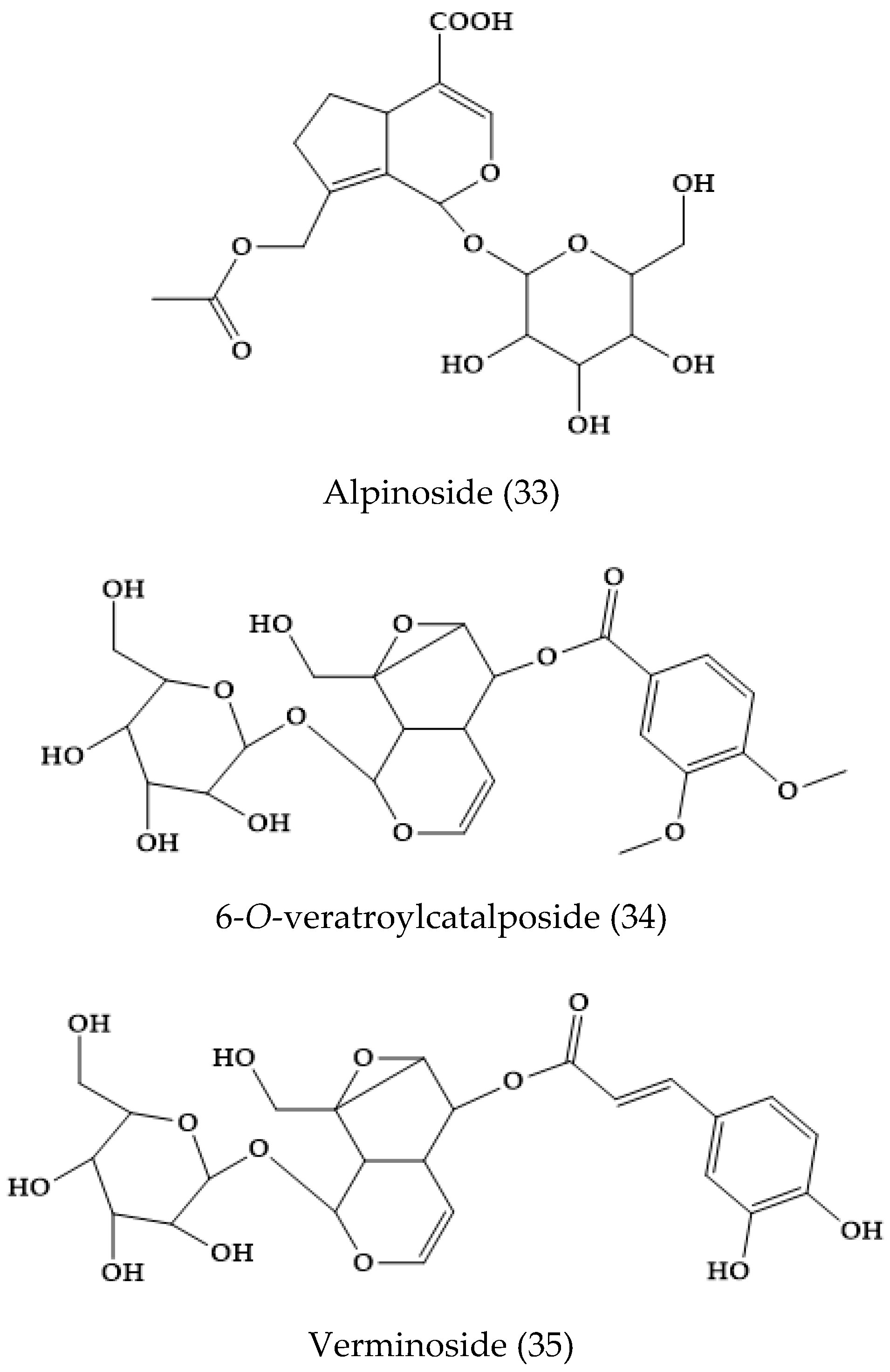
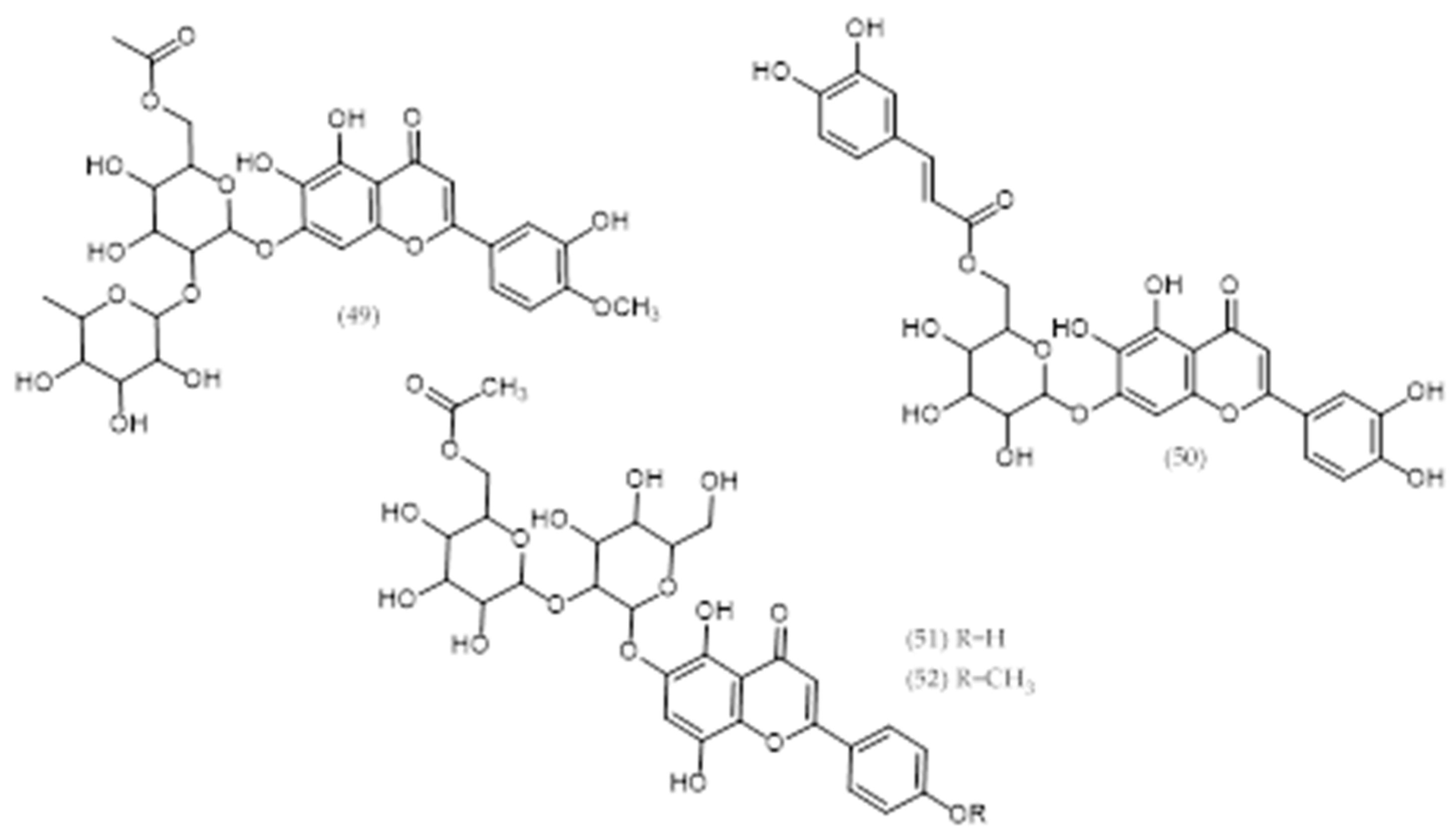
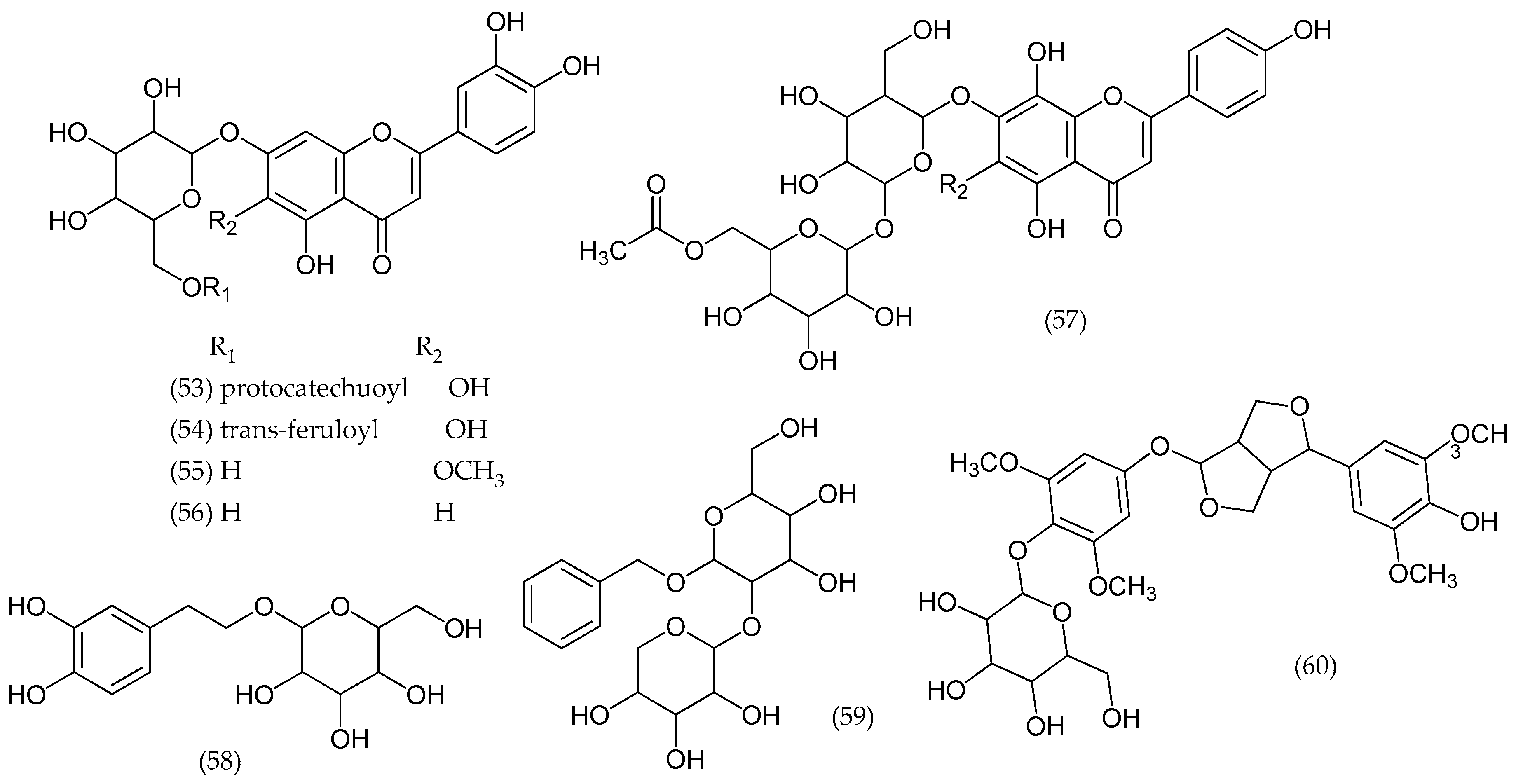
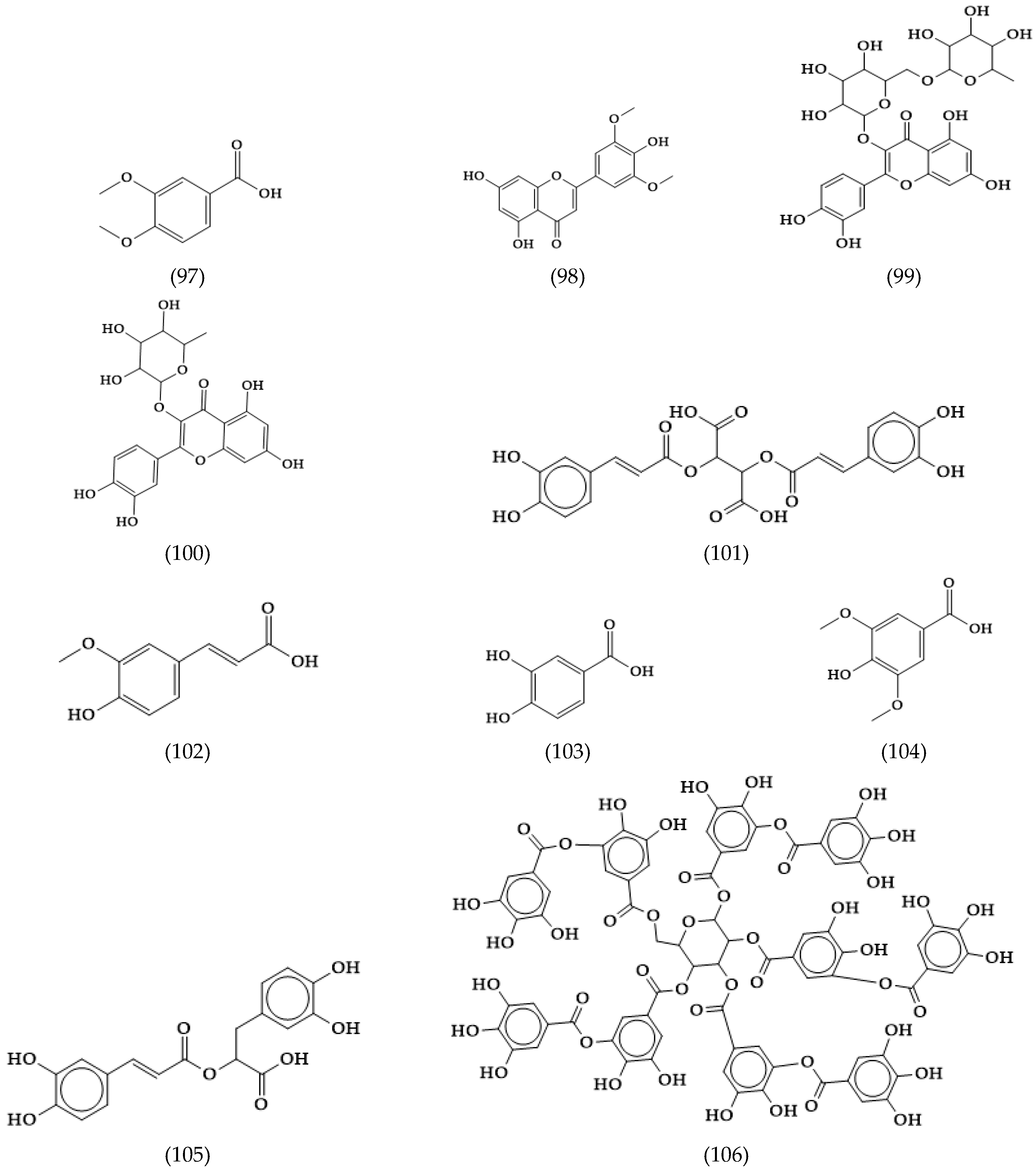
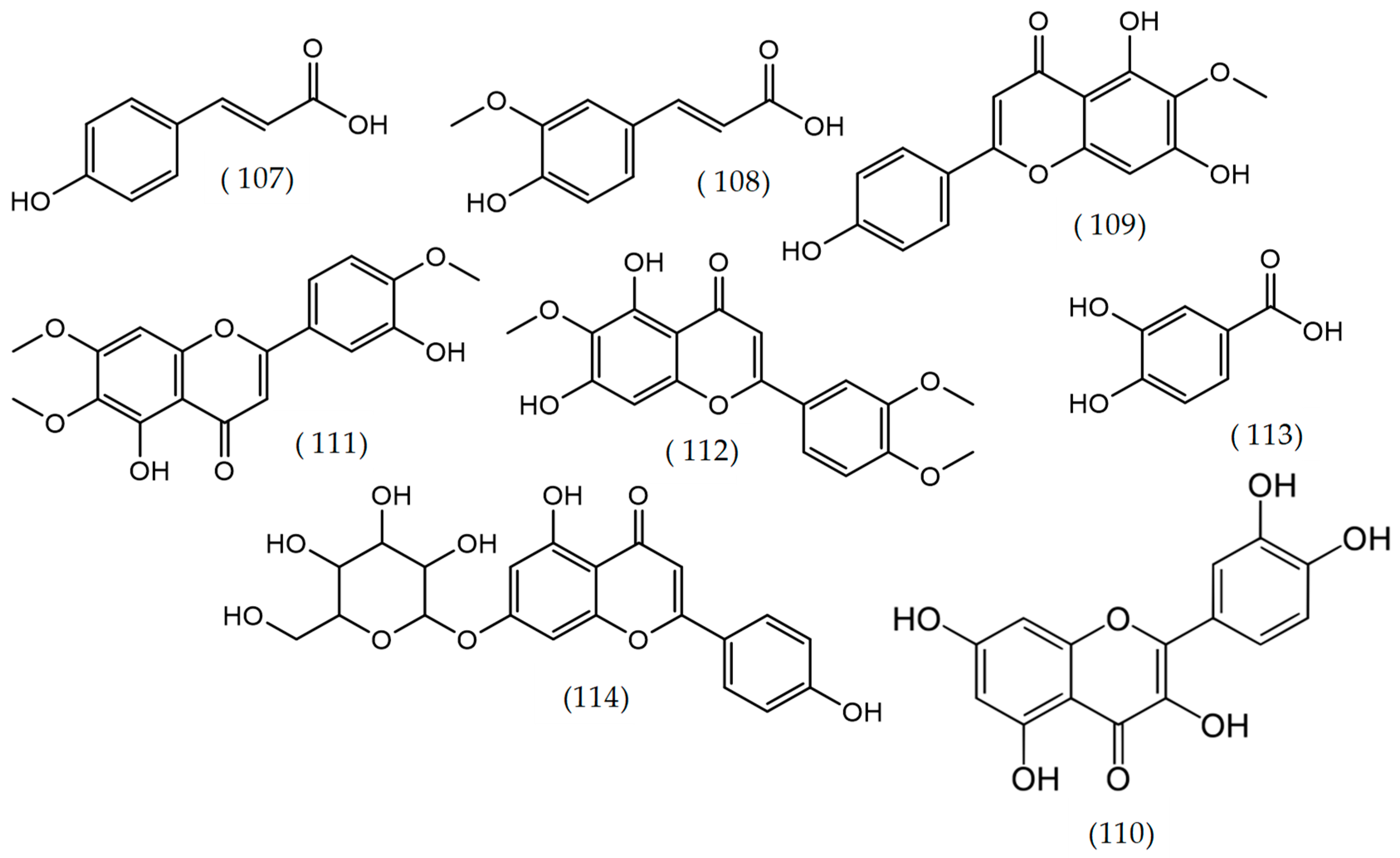
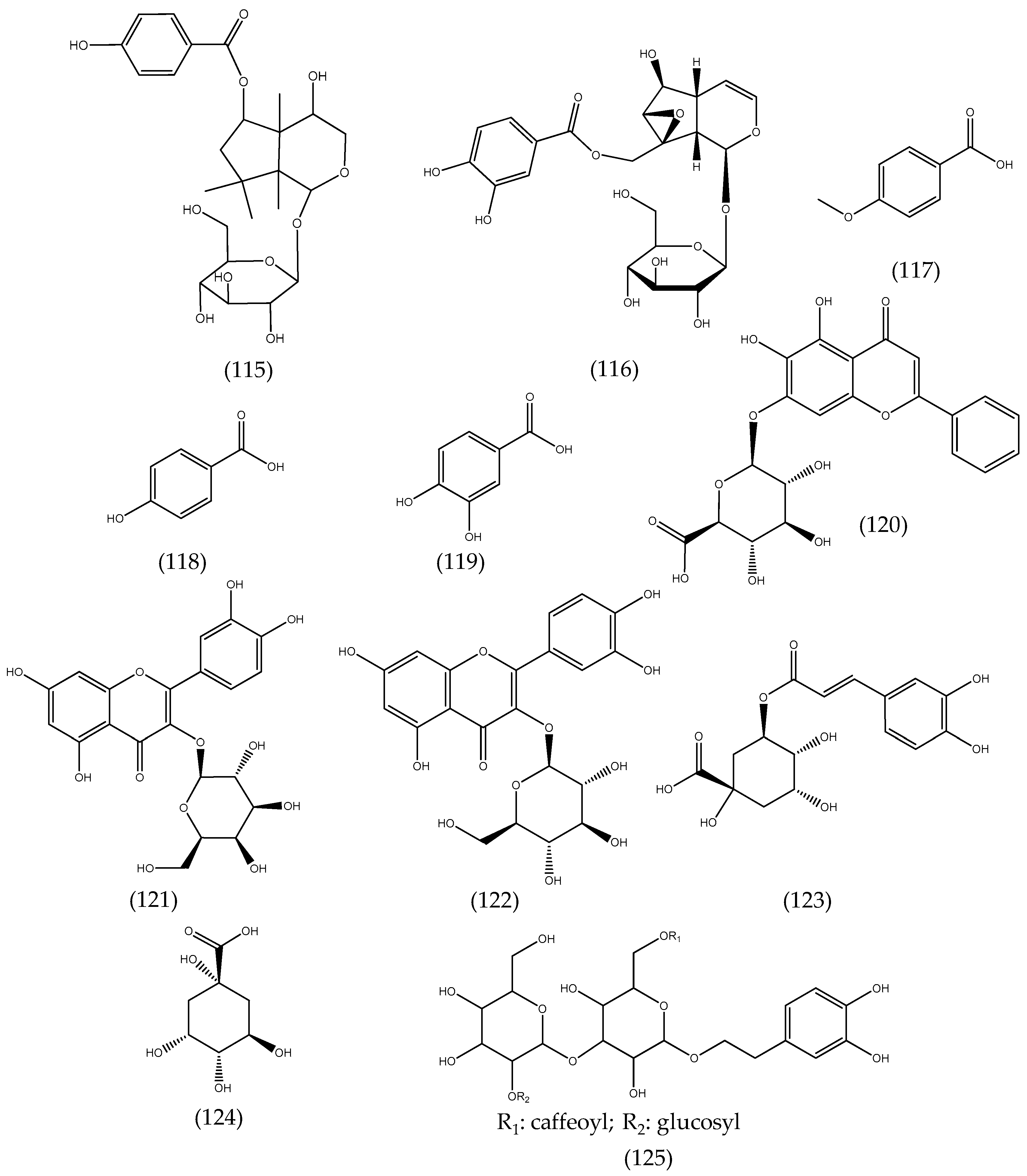
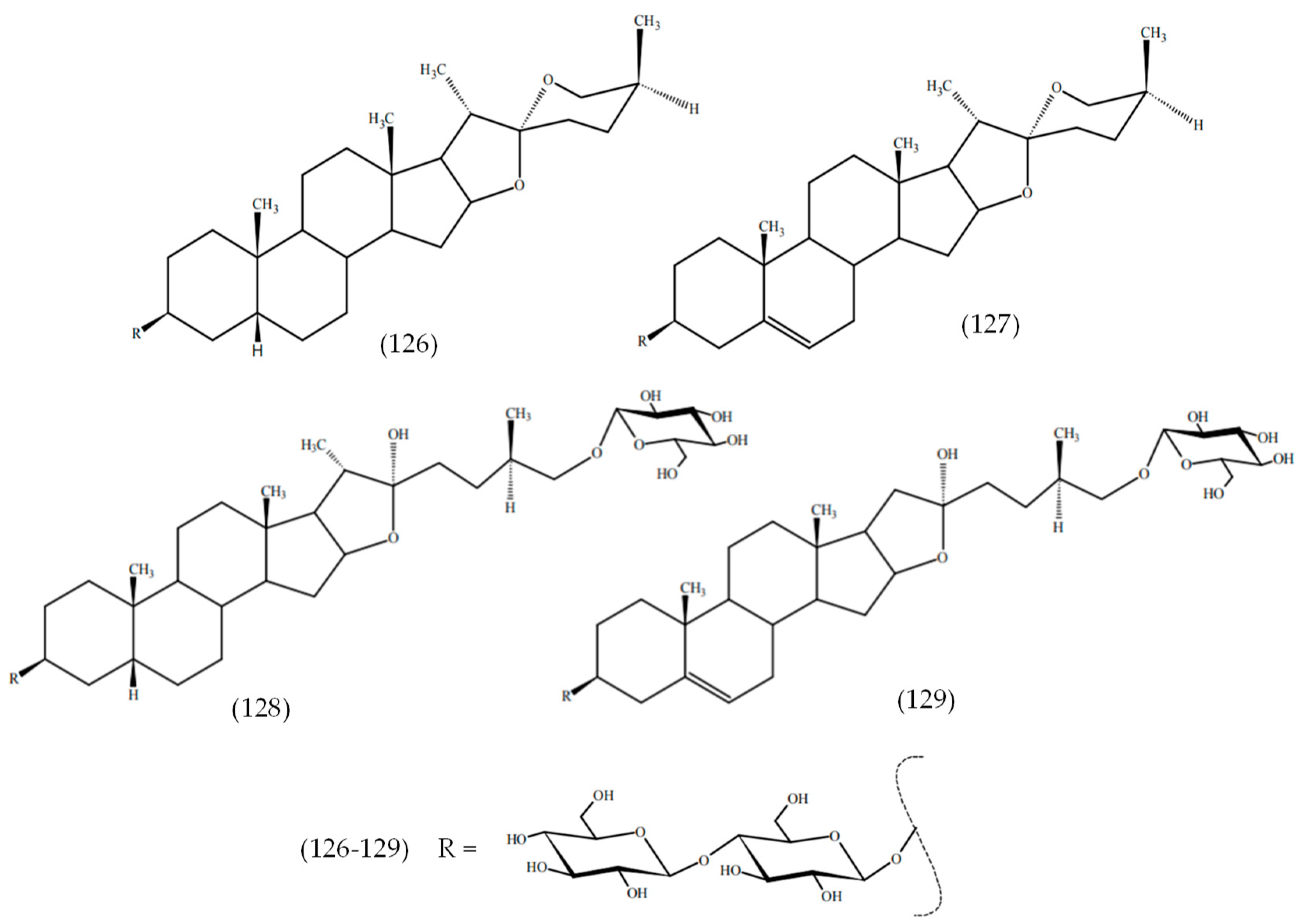
2.21. Others Phytochemicals and Species
3. Antimicrobial Activities of Veronica Plants
3.1. Antibacterial Activity
3.2. Antifungal, Antiviral, and Antiparasitic Activity
4. Antioxidant Activities of Veronica Plants
4.1. In Vitro Studies
4.2. In Vivo Studies
5. Anticancer Activities of Veronica Species
5.1. In Vitro Studies
5.2. In Vivo Studies
6. Anti-Inflammatory Activity
6.1. In Vitro Studies
6.2. In Vivo Studies
7. Other Properties
8. Food Applications of Veronica Plants and Other Uses
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Olmstead, R.G. Whatever happened to scrophulariaceae. Fremontia 2002, 30, 13–22. [Google Scholar]
- Albach, D.C.; Martınez-Ortega, M.M.; Fischer, M.A.; Chase, M.W. A new classification of the tribe veroniceae-problems and a possible solution. Taxon 2004, 53, 429–452. [Google Scholar] [CrossRef]
- Jensen, S.R.; Albach, D.C.; Ohno, T.; Grayer, R.J. Veronica: Iridoids and cornoside as chemosystematic markers. Biochem. Syst. Ecol. 2005, 33, 1031–1047. [Google Scholar] [CrossRef]
- The Plant List. Version 1.1. Available online: http://www.theplantlist.org/ (accessed on 1 January 2019).
- Ellmouni, F.Y.; Karam, M.A.; Ali, R.M.; Albach, D.C. Molecular and morphometric analysis of Veronica l. Section Beccabunga (hill) dumor. Aquat. Bot. 2017, 136, 95–111. [Google Scholar] [CrossRef]
- Plants for a Future. Available online: https://pfaf.org/user/DatabaseSearhResult.aspx (accessed on 1 January 2019).
- Wangkheirakpam, S. Traditional and folk medicine as a target for drug discovery. In Natural Products and Drug Discovery: An Integrated Approach; Mandal, S.C., Mandal, V., Konishi, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 29–56. [Google Scholar]
- World Health Organization. WHO Global Report on Traditional and Complementary Medicine 2019; World Health Organization: Geneva, Switzerland, 2019; pp. 10–80. [Google Scholar]
- Vural, C.; Özcan, S.; Akbulut, M. New combination in Veronica (Scrophulariaceae s.l.) based on morphological characters and the seed storage protein polymorphism. J. Syst. Evol. 2009, 47, 168–172. [Google Scholar] [CrossRef]
- Jeon, H. Anti-inflammatory activity of Veronica peregrina. Nat. Prod. Sci. 2012, 18, 141–146. [Google Scholar]
- Dunkic, V.; Kosalec, I.; Kosir, I.J.; Potocnik, T.; Cerenak, A.; Koncic, M.Z.; Vitali, D.; Muller, I.D.; Kopricanec, M.; Bezic, N.; et al. Antioxidant and antimicrobial properties of Veronica spicata L. (plantaginaceae). Curr. Drug Targets 2015, 16, 1660–1670. [Google Scholar] [CrossRef] [PubMed]
- Kwak, J.H.; Kim, H.J.; Lee, K.H.; Kang, S.C.; Zee, O.P. Antioxidative iridoid glycosides and phenolic compounds from Veronica peregrina. Arch. Pharm. Res. 2009, 32, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Beara, I.; Zivkovic, J.; Lesjak, M.; Ristic, J.; Savikin, K.; Maksimovic, Z.; Jankovic, T. Phenolic profile and anti-inflammatory activity of three veronica species. Ind. Crop. Prod. 2015, 63, 276–280. [Google Scholar] [CrossRef]
- Su, B.; Zhu, Q.; Jia, Z. Aquaticol, a novel bis-sesquiterpene from Veronica anagallis-aquatica. Tetrahedron Lett. 1999, 40, 357–358. [Google Scholar] [CrossRef]
- Grundemann, C.; Garcia-Kaufer, M.; Sauer, B.; Stangenberg, E.; Konczol, M.; Merfort, I.; Zehl, M.; Huber, R. Traditionally used Veronica officinalis inhibits proinflammatory mediators via the nf-kb signalling pathway in a human lung cell line. J. Ethnopharmacol. 2013, 145, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Kupeli, E.; Harput, U.S.; Varel, M.; Yesilada, E.; Saracoglu, I. Bioassay-guided isolation of iridoid glucosides with antinociceptive and anti-inflammatory activities from Veronica anagallis-aquatica L. J. Ethnopharmacol. 2005, 102, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Vogl, S.; Picker, P.; Mihaly-Bison, J.; Fakhrudin, N.; Atanasov, A.G.; Heiss, E.H.; Wawrosch, C.; Reznicek, G.; Dirsch, V.M.; Saukel, J.; et al. Ethnopharmacological in vitro studies on austria’s folk medicine-an unexplored lore in vitro anti-inflammatory activities of 71 austrian traditional herbal drugs. J. Ethnopharmacol. 2013, 149, 750–771. [Google Scholar] [CrossRef] [PubMed]
- Raclariu, A.C.; Mocan, A.; Popa, M.O.; Vlase, L.; Ichim, M.C.; Crisan, G.; Brysting, A.; Boer, H. Veronica officinalis product authentication using DNA metabarcoding and hplc-ms reveals widespread adulteration with Veronica chamaedrys. Front. Pharmacol. 2017, 8, 378. [Google Scholar] [CrossRef] [PubMed]
- Harput-Hudaverdi, U.S.; Oztunca, F.H.; Saracoglu, I. Comparative phytochemical and biological studies on Veronica cuneifolia subsp. Cuneifolia and v. Cymbalaria. Planta Med. 2008, 74, PC88. [Google Scholar]
- Sharifi-Rad, J.; Tayeboon, G.S.; Niknam, F.; Sharifi-Rad, M.; Mohajeri, M.; Salehi, B.; Iriti, M.; Sharifi-Rad, M. Veronica persica poir. Extract–antibacterial, antifungal and scolicidal activities, and inhibitory potential on acetylcholinesterase, tyrosinase, lipoxygenase and xanthine oxidase. Cell. Mol. Biol. 2018, 64, 50–56. [Google Scholar] [CrossRef]
- Grayer-Barkmeijer, R.J. A chemosystematic study of Veronica: Iridoid glucosides. Biochem. Syst. Ecol. 1973, 1, 101–110. [Google Scholar] [CrossRef]
- Afifi-Yazar, F.U.; Sticher, O. Verproside, a new iridoid glucoside from Veronica officinalis L. (scrophulariaceae). Helivetica Chim. Acta 1980, 63, 1905–1907. [Google Scholar] [CrossRef]
- Johansen, M.; Larsen, T.S.; Mattebjerg, M.A.; Gotfredsen, C.H.; Jensen, S.R. Chemical markers in Veronica sect. Hebe. Biochem. Syst. Ecol. 2007, 35, 614–620. [Google Scholar] [CrossRef]
- Grayer-Barkmeijer, R.J. Flavonoids in Parahebe and Veronica: A chemosystematic study. Biochem. Syst. Ecol. 1978, 6, 131–137. [Google Scholar] [CrossRef]
- Nikolova, M.T.; Taskova, R.M.; Peev, D.R. Exudate flavonoid aglycones of Veronica: Ecological and systematic implications. Biochem. Syst. Ecol. 2005, 33, 1258–1268. [Google Scholar] [CrossRef]
- Chari, V.M.; Grayer-Barkmeijer, R.J.; Harborne, J.B.; Österdahl, B.G. An acylated allose-containing 8-hydroxyflavone glycoside from Veronica filiformis. Phytochemistry 1981, 20, 1977–1979. [Google Scholar] [CrossRef]
- Ma, C.Y.; Zhu, K.X.; Yang, D.M.; Yang, J.S.; Yu, D.Q. Studies on chemical constituents of Veronica linariifolia pall. Ex link. Sub. Dilatata (nakai et kitagawa) hong. Yao Xue Xue Bao = Acta Pharm. Sin. 1991, 26, 203–208. [Google Scholar]
- Hong, J.L. Phenolic Constituents of Veronica linariifolia. Chin. J. Nat. Med. 2008, 6, 126–129. [Google Scholar] [CrossRef]
- Li, F. Analysis of chemical constituents of essential oil in Veronica linariifolia by gas chromatography-mass spectrometry. Chin. J. Anal. Chem. 2002, 30, 822–825. [Google Scholar]
- Ozipek, M.; Saracoglu, I.; Kojima, K.; Ogihara, Y.; Calis, I. Fuhsioside, a new phenylethanoid glucoside from Veronica fuhsii. Chem. Pharm. Bull. 1999, 47, 561–562. [Google Scholar] [CrossRef]
- Taskova, R.; Handjieva, N.; Evstatieva, L.; Popov, S. Iridoid glucosides from Plantago cornuti, Plantago major and Veronica cymbalaria. Phytochemistry 1999, 52, 1443–1445. [Google Scholar] [CrossRef]
- Handan Öztunca, F.; Saracoglu, I.; Harput, U. Comparative hplc determination of iridoid contents in Veronica cuneifolia subsp. Cuneifolia and V. cymbalaria. Turk. J. Pharm. Sci. 2011, 8, 63–70. [Google Scholar]
- Su, B.N.; Yang, L.; Gao, K.; Jia, Z.J. Aquaticol, a bis-sesquiterpene and iridoid glucosides from Veronica anagallis-aquatica. Planta Med. 2000, 66, 281–283. [Google Scholar] [CrossRef]
- Sarker, S.D.; Bright, C.; Bartholomew, B.; Watson, A.A.; Nash, R.J. Calendin, tyrosol and two benzoic acid derivatives from Veronica persica (scrophulariaceae). Biochem. Syst. Ecol. 2000, 28, 799–801. [Google Scholar] [CrossRef]
- Harput, U.S.; Saracoglu, I.; Inoue, M.; Ogihara, Y. Phenylethanoid and iridoid glycosides from Veronica persica. Chem. Pharm. Bull. 2002, 50, 869–871. [Google Scholar] [CrossRef] [PubMed]
- Albach, D.C.; Grayer, R.J.; Jensen, S.R.; Ozgokce, F.; Veitch, N.C. Acylated flavone glycosides from Veronica. Phytochemistry 2003, 64, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.R.; Gotfredsen, C.H.; Harput, U.S.; Saracoglu, I. Chlorinated iridoid glucosides from Veronica longifolia and their antioxidant activity. J. Nat. Prod. 2010, 73, 1593–1596. [Google Scholar] [CrossRef] [PubMed]
- Saracoglu, I.; Varel, M.; Harput, U.S.; Nagatsu, A. Acylated flavonoids and phenol glycosides from Veronica thymoides subsp. Pseudocinerea. Phytochemistry 2004, 65, 2379–2385. [Google Scholar] [CrossRef] [PubMed]
- Albach, D.C.; Grayer, R.J.; Kite, G.C.; Jensen, S.R. Veronica: Acylated flavone glycosides as chemosystematic markers. Biochem. Syst. Ecol. 2005, 33, 1167–1177. [Google Scholar] [CrossRef]
- Kostadinova, E.P.; Alipieva, K.I.; Kokubun, T.; Taskova, R.M.; Handjieva, N.V. Phenylethanoids, iridoids and a spirostanol saponin from Veronica turrilliana. Phytochemistry 2007, 68, 1321–1326. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.; Gotfredsen, C.; Grayer, R. Unusual iridoid glycosides in Veronica sects. Hebe and labiatoides. Biochem. Syst. Ecol. 2008, 36, 207–215. [Google Scholar] [CrossRef]
- Teng, J.; Li, H.Q.; Yao, Z.; Zhang, Y.W.; Zhang, F.G.; Duan, H.Q. Anticancer activity of diterpenes from Veronica sibirica in vitro. Chin. Tradit. Herb. Drugs 2008, 39, 967–970. [Google Scholar]
- Teng, J.; Zhang, F.G.; Zhang, Y.W.; Takaishi, Y.; Duan, H.Q. A new iridoid glycoside from Veronica sibirica. Chin. Chem. Lett. 2008, 19, 450–452. [Google Scholar] [CrossRef]
- Ahn, D.; Lee, S.I.; Yang, J.H.; Cho, C.H.; Hwang, Y.H.; Park, J.H.; Kim, D.K. Superoxide radical scavengers from the whole plant of Veronica peregrina. Nat. Prod. Sci. 2011, 17, 142–146. [Google Scholar]
- Kim, D.K.; Jeon, H.; Cha, D.S. 4-hydroxybenzoic acid-mediated lifespan extension in Caenorhabditis elegans. J. Funct. Foods 2014, 7, 630–640. [Google Scholar] [CrossRef]
- Barreira, J.C.M.; Dias, M.I.; Živković, J.; Stojković, D.; Soković, M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Phenolic profiling of Veronica spp. Grown in mountain, urban and sandy soil environments. Food Chem. 2014, 163, 275–283. [Google Scholar] [CrossRef]
- Mocan, A.; Vodnar, D.C.; Vlase, L.; Crisan, O.; Gheldiu, A.M.; Crisan, G. Phytochemical characterization of Veronica officinalis l., v. Teucrium l. And v. Orchidea crantz from romania and their antioxidant and antimicrobial properties. Int. J. Mol. Sci. 2015, 16, 21109–21127. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Sun, Y.; Shu, Y.; Tan, S.; Yin, L.; Guo, Y.; Tang, L. Hsccc separation of the two iridoid glycosides and three phenolic compounds from Veronica ciliata and their in vitro antioxidant and anti-hepatocarcinoma activities. Molecules 2016, 21, 1234. [Google Scholar] [CrossRef] [PubMed]
- Ouache, R.; Harkat, H.; Pale, P.; Oulmi, K. Phytochemical compounds and anti-corrosion activity of Veronica rosea. Nat. Prod. Res. 2018, 16, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Escobar, J.A.; Alvarez, L.; Rodríguez-López, V.; Bahena, S.M. Cytotoxic glucosydic iridoids from Veronica americana. Phytochem. Lett. 2013, 6, 610–613. [Google Scholar] [CrossRef]
- Kroll-Møller, P.; Pedersen, K.; Gousiadou, C.; Kokubun, T.; Albach, D.; Taskova, R.; Garnock-Jones, P.J.; Gotfredsen, C.H.; Jensen, S.R. Iridoid glucosides in the genus Veronica (plantaginaceae) from new zealand. Phytochemistry 2017, 140, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Taskova, R.M.; Kokubun, T.; Ryan, K.G.; Garnock-Jones, P.J.; Jensen, S.R. Phenylethanoid and iridoid glycosides in the new zealand snow hebes (Veronica, plantaginaceae). Chem. Pharm. Bull. 2010, 58, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Maggi, A.; Taskova, R.; Gotfredsen, C.H.; Bianco, A.; Jensen, S.R. Chemical markers in Veronica sect. Hebe. Iiiq. Biochem. Syst. Ecol. 2009, 37, 731–736. [Google Scholar] [CrossRef]
- Jensen, S.R.; Opitz, S.E.W.; Gotfredsen, C.H. A new phenylethanoid triglycoside in Veronica beccabunga L. Biochem. Syst. Ecol. 2011, 39, 193–197. [Google Scholar] [CrossRef]
- Marchenko, A.; Kintya, P.; Wyrzykiewicz, B.; Gorincioi, E. Steroidal glycosides from Veronica chamaedrys l. Part i. The structures of chamaedrosides c, c1, c2, e, e1 and e2. Nat. Prod. Commun. 2012, 7, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Tayeboon, G.S.; Sharifi-Rad, J.; Iriti, M.; Varoni, E.M.; Razazi, S. Inhibitory activity on type 2 diabetes and hypertension key-enzymes, and antioxidant capacity of Veronica persica phenolic-rich extracts. Cell. Mol. Biol. 2016, 62, 80–85. [Google Scholar] [PubMed]
- Stojkovic, D.S.; Zivkovic, J.; Sokovic, M.; Glamoclija, J.; Ferreira, I.C.F.R.; Jankovic, T.; Maksimovic, Z. Antibacterial activity of Veronica montana L. Extract and of protocatechuic acid incorporated in a food system. Food Chem. Toxicol. 2013, 55, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Dunki, V.; Kosalec, I.; Kosir, J.I.; Potonik, T.; Gerenak, A.; Koncik, M.Z.; Vitali, D.; Müller, I.D.; Koprianec, M.; Bezi, N.; et al. Antioxidant and antimicrobial properties of Veronica spicata L. (plantaginaceae). Curr. Drug Targets 2014, 15, 1–12. [Google Scholar] [CrossRef]
- Zivkovic, J.; Barreira, J.C.M.; Stojkovic, D.; Cebovic, T.; Santos-Buelga, C.; Maksimovic, Z.; Ferreira, I.C.F.R. Phenolic profile, antibacterial, antimutagenic and antitumour evaluation of Veronica urticifolia jacq. J. Funct. Foods 2014, 9, 192–201. [Google Scholar] [CrossRef]
- Dulger, B.; Ugurlu, E. Evaluation of antimicrobial activity of some endemic scrophulariaceae members from turkey. Pharm. Biol. 2005, 43, 275–279. [Google Scholar] [CrossRef]
- Ginovyan, M.M.; Trchounian, A.H. Screening of some plant materials used in armenian traditional medicine for their antimicrobial activity. Chem. Biol. 2017, 51, 44–53. [Google Scholar]
- Mocan, A.; Vlase, L.; Arsene, A.L.; Vodnar, D.; Bischin, C.; Silaghi-Dumitrescu, R.; Crişan, G. Hplc/ms analysis of caffeic and chlorogenic acids from three romanian Veronica species and their antioxidant and antimicrobial properties. Farmacia 2015, 63, 890–896. [Google Scholar]
- Sharifi-Rad, J.; Iriti, M.; Setzer, W.N.; Sharifi-Rad, M.; Roointan, A.; Salehi, B. Antiviral activity of Veronica persica poir. On herpes virus infection. Cell. Mol. Biol. 2018, 64, 11–17. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Roointan, A.; Setzer, W.N.; Sharifi-Rad, M.; Iriti, M.; Salehi, B. Susceptibility of Leishmania major to Veronica persica poir. Extracts-in vitro and in vivo assays. Cell. Mol. Biol. 2018, 64, 44–49. [Google Scholar] [CrossRef]
- Fierascu, R.C.; Georgiev, M.I.; Fierascu, I.; Ungureanu, C.; Avramescu, S.M.; Ortan, A.; Georgescu, M.I.; Sutan, A.N.; Zanfirescu, A.; Dinu-Pirvu, C.E.; et al. Mitodepressive, antioxidant, antifungal and anti-inflammatory effects of wild-growing romanian native Arctium lappa L. (Asteraceae) and Veronica persica poiret (plantaginaceae). Food Chem. Toxicol. 2018, 111, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Saracoglu, I.; Harput, U.S.; Inoue, M.; Ogihara, Y. New phenylethanoid glycosides from Veronica pectinata var. Glandulosa and their free radical scavenging activities. Chem. Pharm. Bull. 2002, 50, 665–668. [Google Scholar] [PubMed]
- Valyova, M.; Hadjimitova, V.; Stoyanov, S.; Ganeva, Y.; Petrov, T.; Petrov, I. Free radicals scavenging activity of extracts from bulgarian Veronica officinalis L. and GC-MS analysis of ethanol extract. Int. J. Aesthet. Antiaging Med. 2009, 2, 1–5. [Google Scholar]
- Harput, U.S.; Genc, Y.; Khan, N.; Saracoglu, I. Radical scavenging effects of different Veronica species. Rec. Nat. Prod. 2011, 5, 100–107. [Google Scholar]
- Sharifi-Rad, J.; Sharifi-Rad, M.; Salehi, B.; Iriti, M.; Roointan, A.; Mnayer, D.; Afshari, A. In vitro and in vivo assessment of free radical scavenging and antioxidant activities of Veronica persica poir. Cell. Mol. Biol. 2018, 64, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, J.; Cebovic, T.; Maksimovic, Z. In vivo and in vitro antioxidant effects of three Veronica species. Cent. Eur. J. Biol. 2012, 7, 559–568. [Google Scholar]
- Sun, Y.; Lu, Q.; He, L.; Shu, Y.; Zhang, S.; Tan, S.; Tang, L. Active fragment of Veronica ciliata fisch. Attenuates t-bhp-induced oxidative stress injury in hepg2 cells through antioxidant and antiapoptosis activities. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef]
- Lee, H.; Ghimeray, A.; Yim, J.; Chang, M. Antioxidant, collagen synthesis activity in vitro and clinical test on anti-wrinkle activity of formulated cream containing Veronica officinalis extract. J. Cosmet. Dermatol. Sci. Appl. 2015, 5, 45–51. [Google Scholar]
- Shin, S.A.; Moon, S.Y.; Kim, W.Y.; Paek, S.M.; Park, H.H.; Lee, C.S. Structure-based classification and anti-cancer effects of plant metabolites. Int. J. Mol. Sci. 2018, 19, 2651. [Google Scholar] [CrossRef]
- Harput, U.S.; Iclal, S.; Makoto, I.; Yukio, O. Anti-inflammatory and cytotoxic activities of five Veronica species. Biol. Pharm. Bull. 2002, 25, 483–486. [Google Scholar] [CrossRef]
- Saracoglu, I.; Oztunca, F.H.; Nagatsu, A.; Harput, U.S. Iridoid content and biological activities of Veronica cuneifolia subsp. Cuneifolia and v. Cymbalaria. Pharm. Biol. 2011, 49, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Escobar, J.A.; Bazaldúa, S.; Villarreal, M.L.; Bonilla-Barbosa, J.R.; Mendoza, S.; Rodríguez-López, V. Cytotoxic and antioxidant activities of selected lamiales species from mexico. Pharm. Biol. 2011, 49, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Qiuxia, L.; Shancai, T.; Lisheng, D.; Yiran, G.; Fang, C.; Lin, T. Bioactivity-guided isolation of antioxidant and anti-hepatocarcinoma constituents from Veronica ciliata. Chem. Cent. J. 2016, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Saracoglu, I.; Harput, U.S. In vitro cytotoxic activity and structure activity relationships of iridoid glucosides derived from veronica species: Bioactivities of iridoid glucosides derived from Veronica species. Phytother. Res. 2012, 26, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Ignjatović, D.; Živković, J.; Tovilović, G.; Šavikin, K.; Tomić, M.; Maksimović, Z.; Janković, T. Evaluation of angiogenic and neuroprotective potential of different extracts from three Veronica species. Front. Life Sci. 2015, 8, 107–116. [Google Scholar] [CrossRef]
- Gomes, N.M.; Rezende, C.M.; Fontes, S.P.; Hovell, A.M.C.; Landgraf, R.G.; Matheus, M.E.; Pinto, A.C.; Fernandes, P.D. Antineoplasic activity of Copaifera multijuga oil and fractions against ascitic and solid ehrlich tumor. J. Ethnopharmacol. 2008, 119, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Ohno, T.; Inoue, M.; Ogihara, Y.; Saracoglu, I. Antimetastatic activity of acteoside, a phenylethanoid glycoside. Biol. Pharm. Bull. 2002, 25, 666–668. [Google Scholar] [CrossRef] [PubMed]
- Rang, N.Y.; Sungmo, J.; Seung, H.S. Metabolic features of macrophages innflammatory diseases and cancer. Cancer Lett. 2018, 413, 46–58. [Google Scholar]
- Park, E.J.; Lee, H.S.; Oh, S.R.; Lee, H.K.; Lee, H.S. Pharmacokinetics of verproside after intravenous and oral administration in rats. Arch. Pharm. Res. 2009, 32, 559–564. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Carding, S.R. Inflammatory bowel disease: Cause and immunobiology. Lancet 2007, 369, 1627–1640. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Z. Inflammatory bowel disease related innate immunity and adaptive immunity. Am. J. Transl. Res. 2016, 8, 2490–2497. [Google Scholar] [PubMed]
- Boland, B.S.; Sandborn, W.J.; Chang, J.T. Update on janus kinase antagonists in inflammatory bowel disease. Gastroenterol. Clin. N. Am. 2014, 43, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Coskun, M.; Salem, M.; Pedersen, J.; Nielsen, O.H. Involvement of jak/stat signaling in the pathogenesis of inflammatory bowel disease. Pharmacol. Res. 2013, 76, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pearl, D.S.; Shah, K.; Whittaker, M.A.; Nitch-Smith, H.; Brown, J.F.; Shute, J.K.; Trebble, T.M. Cytokine mucosal expression in ulcerative colitis, the relationship between cytokine release and disease activity. J. Crohn’s Colitis 2013, 7, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soufli, I.; Toumi, R.; Rafa, H.; Touil-Boukoffa, C. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World J. Gastrointest. Pharmacol. 2016, 7, 353–360. [Google Scholar]
- Akanda, M.R.; Nam, H.W.; Wishun, T.; Islam, A.; Choo, B.K.; Park, B.Y. Regulation of jak2/stat3 and nf-κb signal transduction pathways; Veronica polita alleviates dextran sulfate sodium-induced murine colitis. Biomed. Pharmacother. 2018, 100, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Albarracin, H.W.; Alfonso, A.C.; Sanchez, B. Application of essential oils as a preservative to improve the shelf life of Nile tilapia. Vitae Rev. Fac. Quim. Farm. 2012, 19, 34–40. [Google Scholar]
- Zahran, S.E.; El Shehedi, M.A. Efficacy of antimicrobial activity of some herbs on improving quality of some chicken products. Anim. Health Res. J. 2017, 5, 181–194. [Google Scholar]
- Fernandes, D.B.R.V.; Guimaraes, C.I.; Ferreira, R.C.L.; Botrel, A.D.; Borges, V.S.; De Souza, U.A. Microencapsulated rosemary (Rosmarinus officinalis) essential oil as a biopreservative in minas frescal cheese. J. Food Process. Preserv. 2017, 41, e12759. [Google Scholar] [CrossRef]
- Hintz, T.; Matthews, K.K.; Di, R. The use of plant antimicrobial compound for food preservation. Biomed. Res. Int. 2015. [Google Scholar] [CrossRef]
- Tzima, K.; Makris, D.; Nikiforidis, C.V.; Mourtzinos, I. Potential use of rosemary, propolis and thyme as natural food preservatives. J. Nutr. Health 2015, 1, 6. [Google Scholar]
- Antolak, H.; Kregiel, D. Food preservatives from plants, food additives, desiree nedra karunaratne and geethi pamunuwa. IntechOpen 2017. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA). Food Additive Status List. Available online: https://www.fda.gov/food/food-additives-petitions/food-additive-status-list (accessed on 27 November 2018).
- Çelik, E.; Çelik, Y.G.; Meysun, A. Essential oil composition and antibacterial activity of some plant species. J. Appl. Biol. Sci. 2010, 4, 45–48. [Google Scholar]
- Kovaleva, A.M.; Osmachko, A.P.; Kashpur, N.V.; Grudko, I.V. The antibacterial activity of complexes of Veronica longifolia herb. Ukr. Biopharm. J. 2016, 1, 58–62. [Google Scholar] [CrossRef]
- Doğan, A.; Otlu, S.; Çelebi, Ö.; Aksu Kılıçle, P.; Gülmez Sağlam, A.; Doğan, A.N.C.; Mutlu, N. An investigation of antibacterial effects of steroids. Turk. J. Vet. Anim. Sci. 2017, 41, 302–305. [Google Scholar] [CrossRef]
- Blanco-Salas, J.; Gutiérrez-García, L.; Labrador-Moreno, J.; Ruiz-Téllez, T. Wild plants potentially used in human food in the protected area “Sierra Grande de Hornachos” of Extremadura (Spain). Sustainability 2019, 11, 456. [Google Scholar] [CrossRef]
Sample Availability: Samples of compounds are not available from authors. |





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | Common Name | Edibility | Medicinal Use |
|---|---|---|---|
| Veronica agrestis L. | Field speedwell, green field speedwell | Yes | Yes |
| Veronica americana Schwein. ex Benth. | American brooklime, American speedwell | Yes | Yes |
| Veronica anagallis-aquatica L. | Water speedwell | Yes | Yes |
| Veronica arvensis L. | Corn speedwell | No | Yes |
| Veronica beccabunga L. | Brooklime, European speedwell | Yes | Yes |
| Veronica catenata Pennell | Yes | No | |
| Veronica chamaedrys L. | Germander speedwell | Yes | Yes |
| Veronica hederifolia L. | Ivy-leaf speedwell | No | Yes |
| Veronica longifolia L. | Garden speedwell, long-leaf speedwell | Yes | No |
| Veronica officinalis L. | Common speedwell | Yes | Yes |
| Veronica peregrina L. | Necklace weed, neckweed, hairy purslane speedwell | No | Yes |
| Veronica polita Fr. | Gray field speedwell | Yes | Yes |
| Veronica scutellata L. | Marsh speedwell, skullcap speedwell | Yes | No |
| Veronica spuria L. | Bastard speedwell | Yes | No |
| Veronica undulata Wall. | Undulate speedwell | Yes | Yes |
| Veronica strum virginicum (L.) Farw. | Beaumont’s root, Culver’s root, Bowman’s root, Culver’s root, Black root | No | Yes |
| Species | Extract | Compounds |
|---|---|---|
| Veronica argute-serrata | Ethanol | Mannitol (65), catalpol (1), aucubin (8), gardoside (69), ajugol (63), mussaenosidic acid (70), epiloganica acid (71), arborescosidic acid (72), verbascoside-like compounds, acetyl-flavone glycoside |
| Veronica arvensis L. | Ethanol | Mannitol (65), cornoside (64), ajugol (63), salidroside (66), verbascoside-like compounds |
| Veronica biloba schreb. ex L. | Ethanol | Catalpol (1), aucubin (8), ajugol (63), epiloganic acid (71), alpinoside (33) |
| Veronica campylopoda Boiss. | Ethanol | Mannitol (65), catalpol (1), aucubin (8), ajugol (63), verminoside (35), acetyl-flavone glycoside |
| Veronica chamaedryoides Engl. | Ethanol | Verbascoside-like compounds; some iridoid |
| Veronica dillenii Crantz | Ethanol | Verbascoside (67) and cornoside (64) |
| Veronica longifolia L. | Ethanol | Mannitol (65), catalpol (1), aucubin (8), verposide, catalposide (6), verminoside (35), catalpol ester, flavones |
| Veronica magna M.A.Fisch. | Ethanol | Verbascoside-like compounds |
| Veronica micans (M.A.Fisch.) Landolt | Ethanol | Verbascoside (67) and cornoside |
| Veronica micrantha Hoffmanns. & Link | Ethanol | Mannitol (65), aucubin (8), verpectoside B (68), triterpene glycosides |
| Veronica orbelica | Ethanol | Verbascoside-like compounds |
| Veronica vindobonensis (M.A.Fisch.) M.A.Fisch. | Ethanol | Verbascoside (67) and cornoside (64) |
| Species | Plant Part | Extract | Effect | Reference |
|---|---|---|---|---|
| Veronica spicata L. | Flowers and stem | Methanol and ethyl-acetate extracts | MIC values were between 1.25 and 5 mg/mL against Staphylococcus aureus, Microccocus flavus, Listeria monocytogenes, Enterobacter cloacae, Escherichia coli, Bacillus cereus, and Pseudomonas aeruginosa | [11,58] |
| Veronica urticifolia Jacq. | The aerial parts | Methanol extract | The most sensitive germ was Staphylococcus aureus (MIC and MBC = 7.5 mg/mL) | [59] |
| Veronica lycica E. Lehm. | The aerial parts | Methanol extract | The antimicrobial activity was determined against E. coli, S. aureus, Klebsiella pneumoniae, P. aeruginosa, Proteus vulgaris, B. cereus, Mycobacterium smegmatis, L. monocytogenes, Micrococcus luteus, Candida albicans, Rhodotorula rubra, and Kluyveromyces fragilis. The weak antimicrobial effect was observed against the tested microorganisms | [60] |
| Veronica anagallis-aquatica L. | The aerial parts | Methanol extract | The extracts were tested against five bacterial and two yeast strains. They showed significant inhibition compared to the positive control (gentamicin) | [61] |
| Veronica officinalis L., Veronica teucrium L., Veronica orchidea Crantz | The aerial parts | 70% ethanol extract | Two anaerobic bacterial strains were used: Peptostreptococcus anaerobius and Fusobacterium nucleatum. V. teucrium and V. orchidea presented a higher activity (MIC = 31.25 mg/mL and MBC = 62.5 mg/mL) than V. officinalis (MIC and MBC of 62.5 mg/mL), with the most sensitive strain being Peptostreptococcus anaerobius | [62] |
| V. officinalis, V. teucrium, V. orchidea | The aerial parts | 70% ethanol extract | Eight bacterial strains were used: Staphylococcus aureus, Bacillus cereus, Listeria monocytogenes, Listeria ivanovii, Pseudomonas aeruginosa, Enterococcus faecalis, Salmonella typhimurium, and Escherichia coli. The most sensitive strains were Staphylococcus aureus, Listeria monocytogenes, and Listeria ivanovii with MIC values between 3.9 and 15.62 mg/mL | [47] |
| Veronica persica Poir | The aerial parts | 70% methanol extract | V. persica extract demonstrated an antifungal effect against Candida albicans and Aspergillus niger at a concentration 300 μg/mL of extract | [20] |
| Type of Studies | Primary Outcomes | Active Compounds | Veronica spp. | References | |
|---|---|---|---|---|---|
| In vitro | Human neuroblastoma cell line SH-SY5Y | Neuroprotective against H2O2 induced cytotoxicity | Iridoid glucosides acteoside, and aucubin (only in V. urticifolia) | Veronica urticifolia Jacq. Veronica teucrium L. Veronica jacquinii Baumg. | [79] |
| Human endothelial cells EA.hy 926 | Angiogenic | Phenylpropanoids and flavonoids | V. jacquinii V. teucrium V. urticifolia | ||
| Human lung epithelial cells A549 | Anti-inflammatory in lung diseases (anti-asthmatic) | Iridoid glycosides (verminoside, verproside) | Veronicaofficinalis L. | [15] | |
| Human cancer cell lines HF-6 (colon), PC-3 (prostate) human normal MRC-5 cells (fetal lung fibroblast) | Cytotoxic | Iridoids | Veronica americana Schwein. ex Benth. | [50] | |
| SK-N-SH human neuroblastoma cell line, BEL-7402 human hepatoma cell line | Cytotoxic | Diterpenes | Veronica sibirica L. | [42] | |
| In vivo | Phenyl-p-benzoquinone writhing test and carrageenan induced hind paw edema model in mice | Antinociceptive and anti-inflammatory | Iridoid glucosides, catalposide and verproside | Veronica anagallis-aquatica L. | [16] |
| Rats′ paw edema induced by dextran | Anti-inflammatory | Phenolic compounds and iridoids | Veronica persica Poir | [65] | |
| Clinical | Study design: randomized, placebo controlled for 58 days | Anti-wrinkles, antiaging of skin | Verbascoside | V. officinalis | [72] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salehi, B.; Shivaprasad Shetty, M.; V. Anil Kumar, N.; Živković, J.; Calina, D.; Oana Docea, A.; Emamzadeh-Yazdi, S.; Sibel Kılıç, C.; Goloshvili, T.; Nicola, S.; et al. Veronica Plants—Drifting from Farm to Traditional Healing, Food Application, and Phytopharmacology. Molecules 2019, 24, 2454. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132454
Salehi B, Shivaprasad Shetty M, V. Anil Kumar N, Živković J, Calina D, Oana Docea A, Emamzadeh-Yazdi S, Sibel Kılıç C, Goloshvili T, Nicola S, et al. Veronica Plants—Drifting from Farm to Traditional Healing, Food Application, and Phytopharmacology. Molecules. 2019; 24(13):2454. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132454
Chicago/Turabian StyleSalehi, Bahare, Mangalpady Shivaprasad Shetty, Nanjangud V. Anil Kumar, Jelena Živković, Daniela Calina, Anca Oana Docea, Simin Emamzadeh-Yazdi, Ceyda Sibel Kılıç, Tamar Goloshvili, Silvana Nicola, and et al. 2019. "Veronica Plants—Drifting from Farm to Traditional Healing, Food Application, and Phytopharmacology" Molecules 24, no. 13: 2454. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132454