
Seasonal and Diurnal Variation in Leaf Phenolics of Three Medicinal Mediterranean Wild Species: What Is the Best Harvesting Moment to Obtain the Richest and the Most Antioxidant Extracts?


 and
and 
Abstract
:1. Introduction
- (i)
- evaluate the monthly and daily changes of polyphenols in leaves of P. latifolia, C. incanus, and P. lentiscus collected in their natural environment in order to identify the best harvesting moment (balsamic time) for each species;
- (ii)
- evaluate the possible correlation between phenolic content and temperature and irradiation to know how these environmental factors modulate leaf phenolic production; and,
- (iii)
- verify the monthly variations in antioxidant capacity of the richest-polyphenol extracts.
2. Results and Discussion
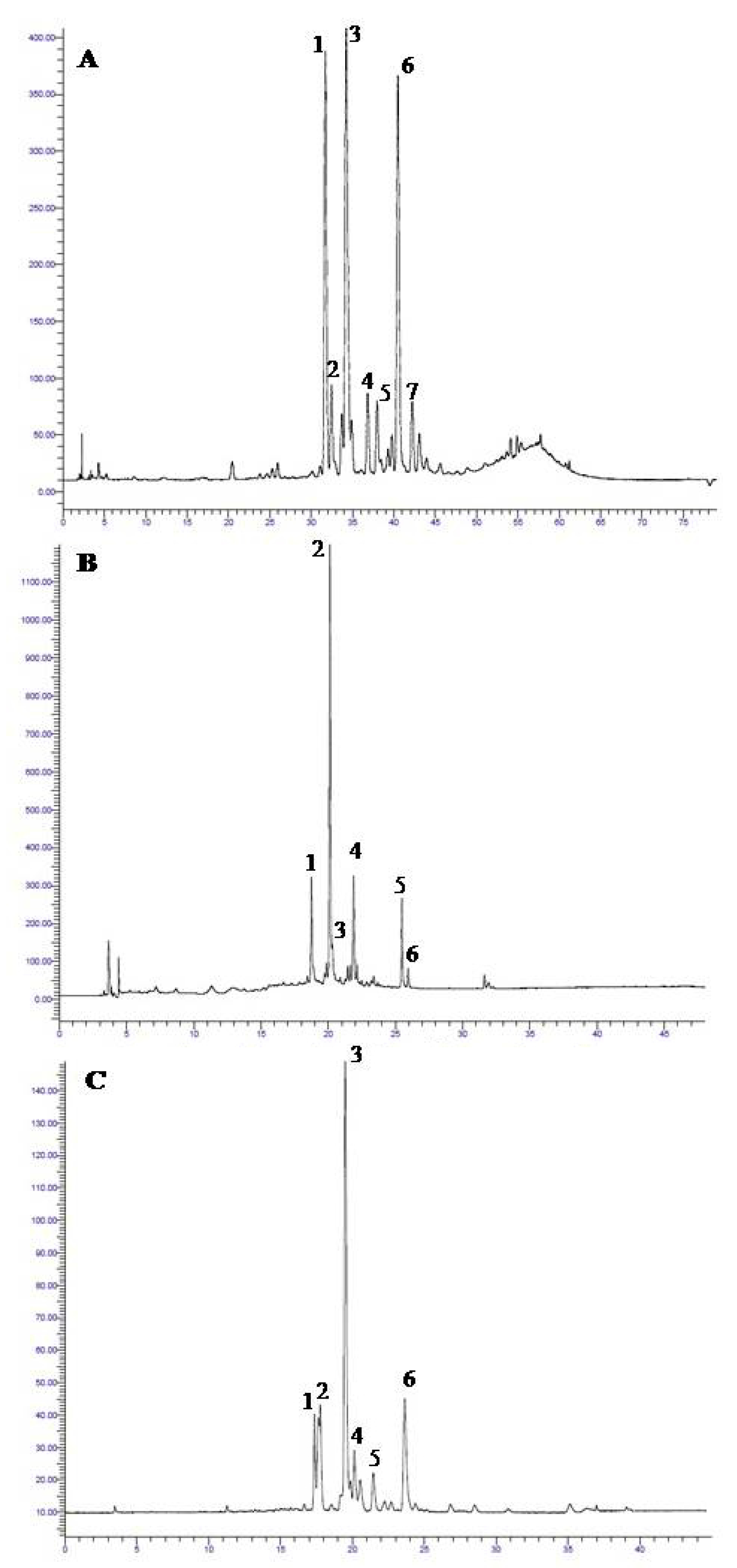
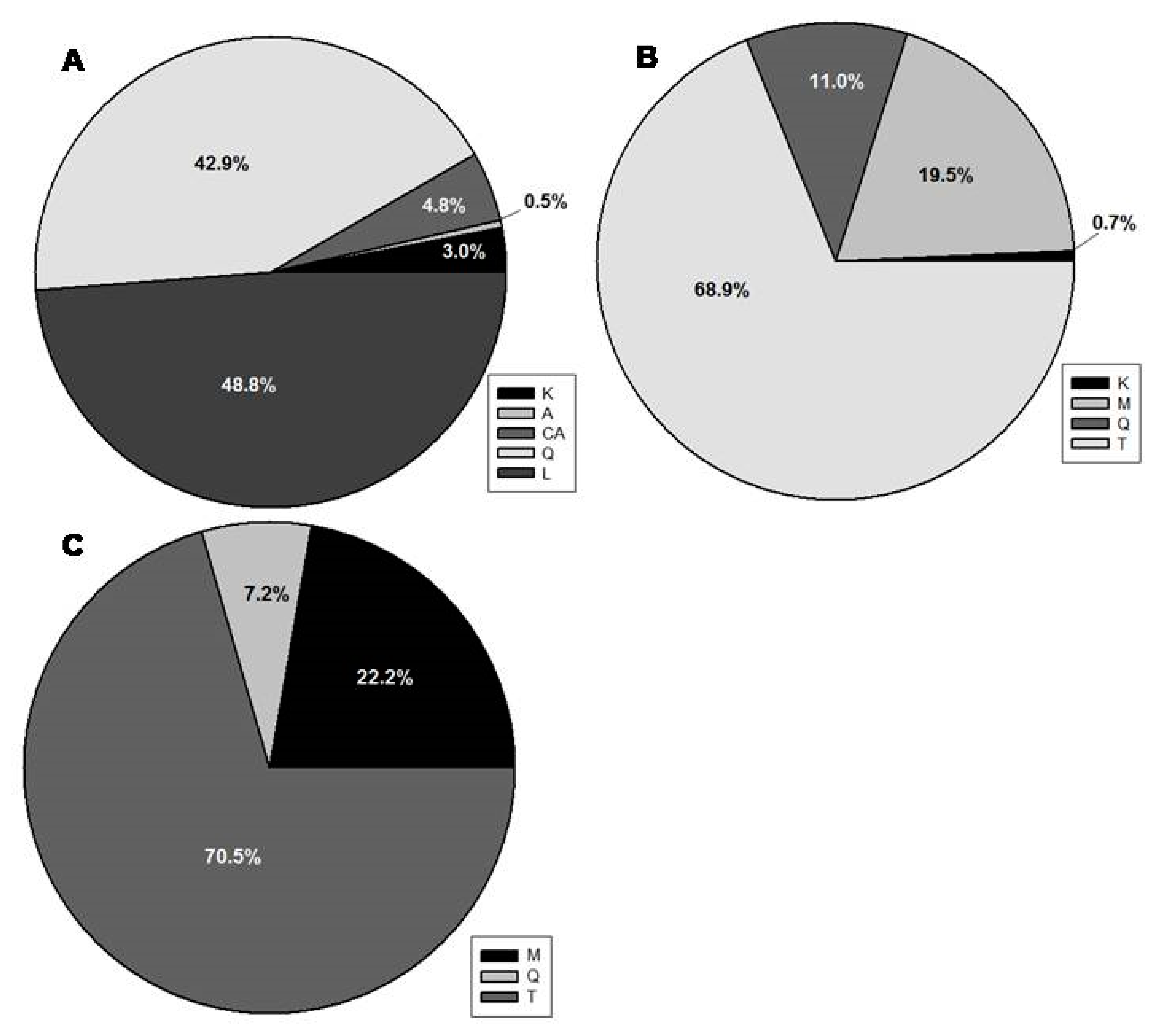
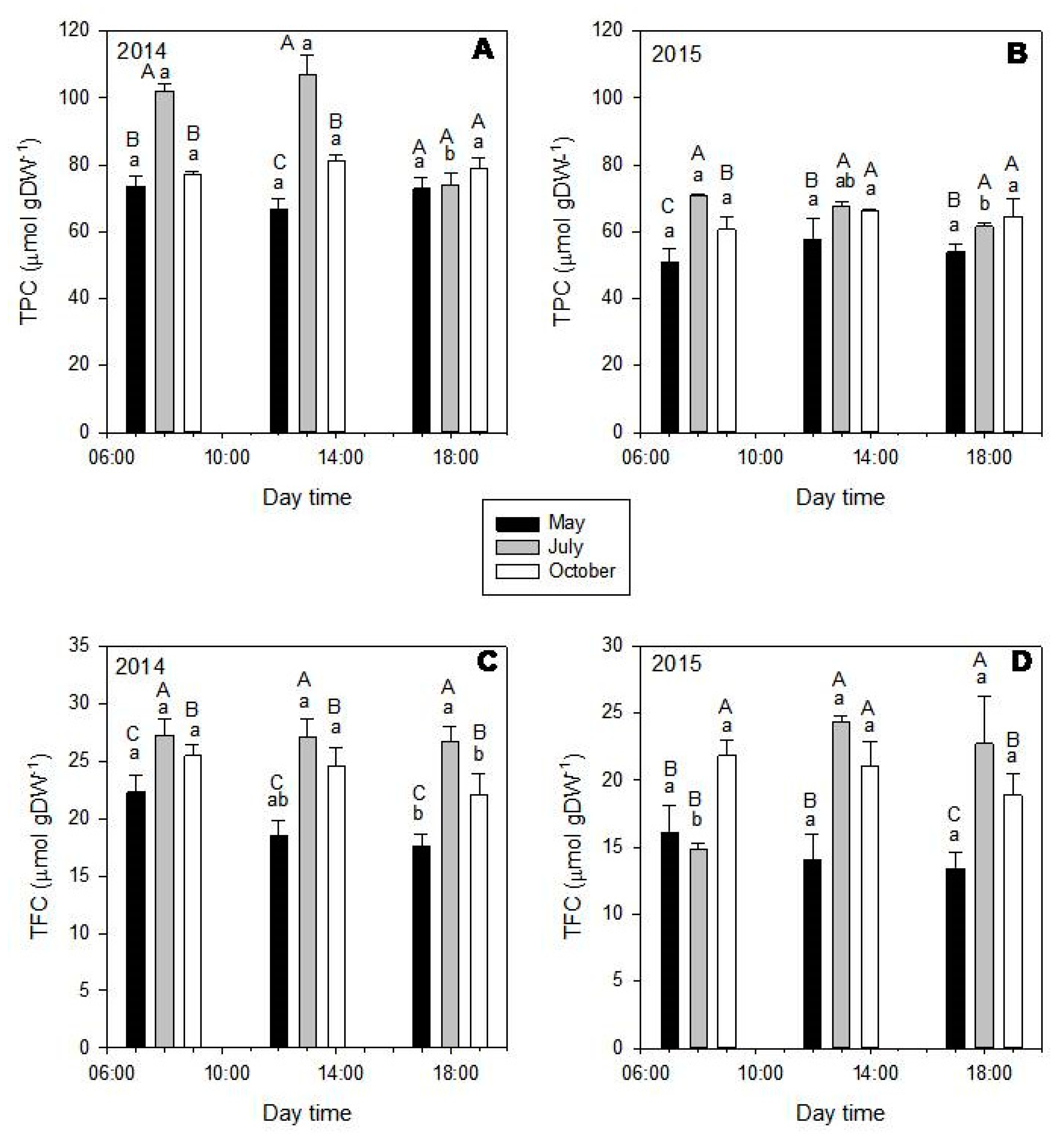
2.1. General Phenolic Profile of the Leaf Extracts
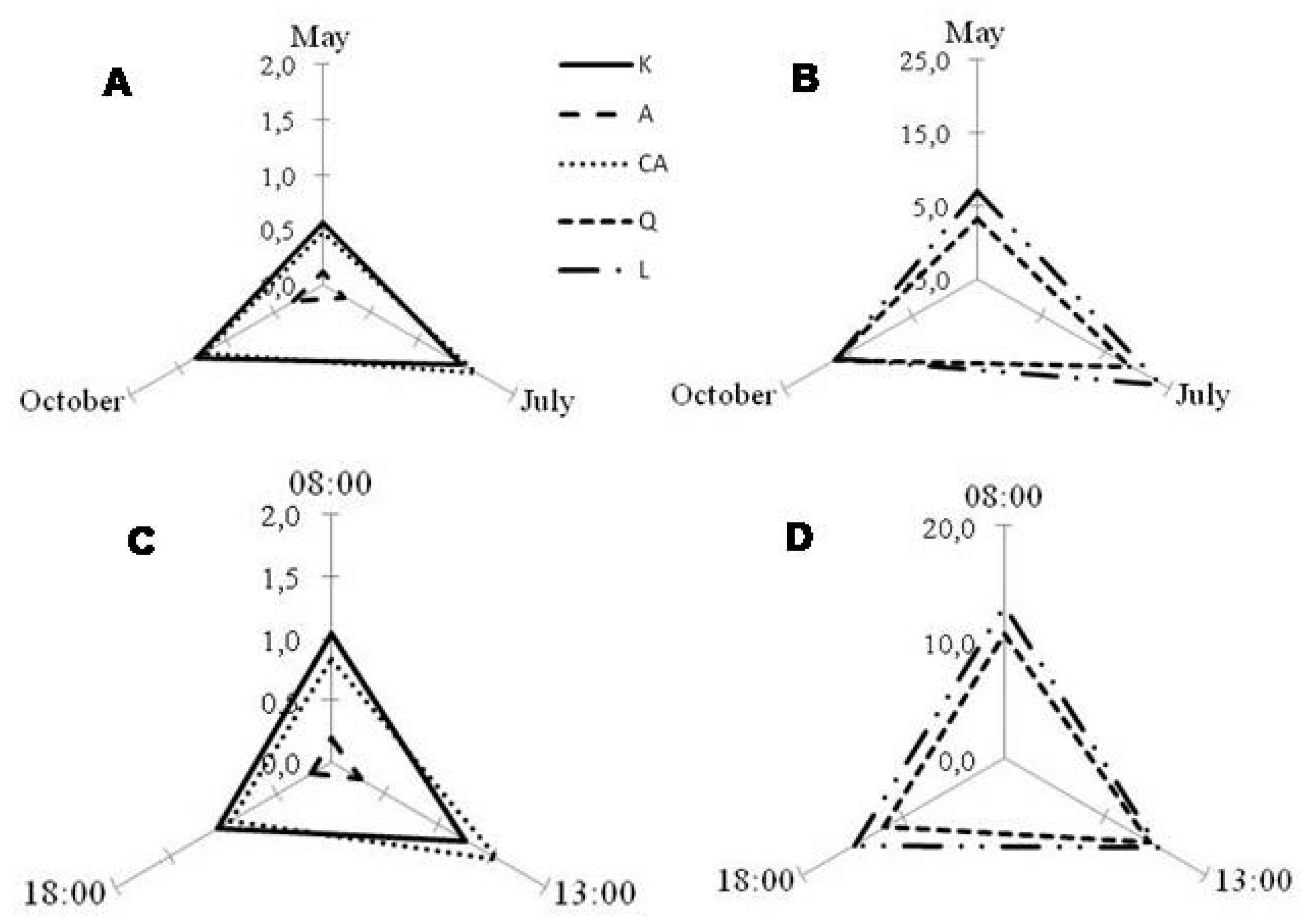
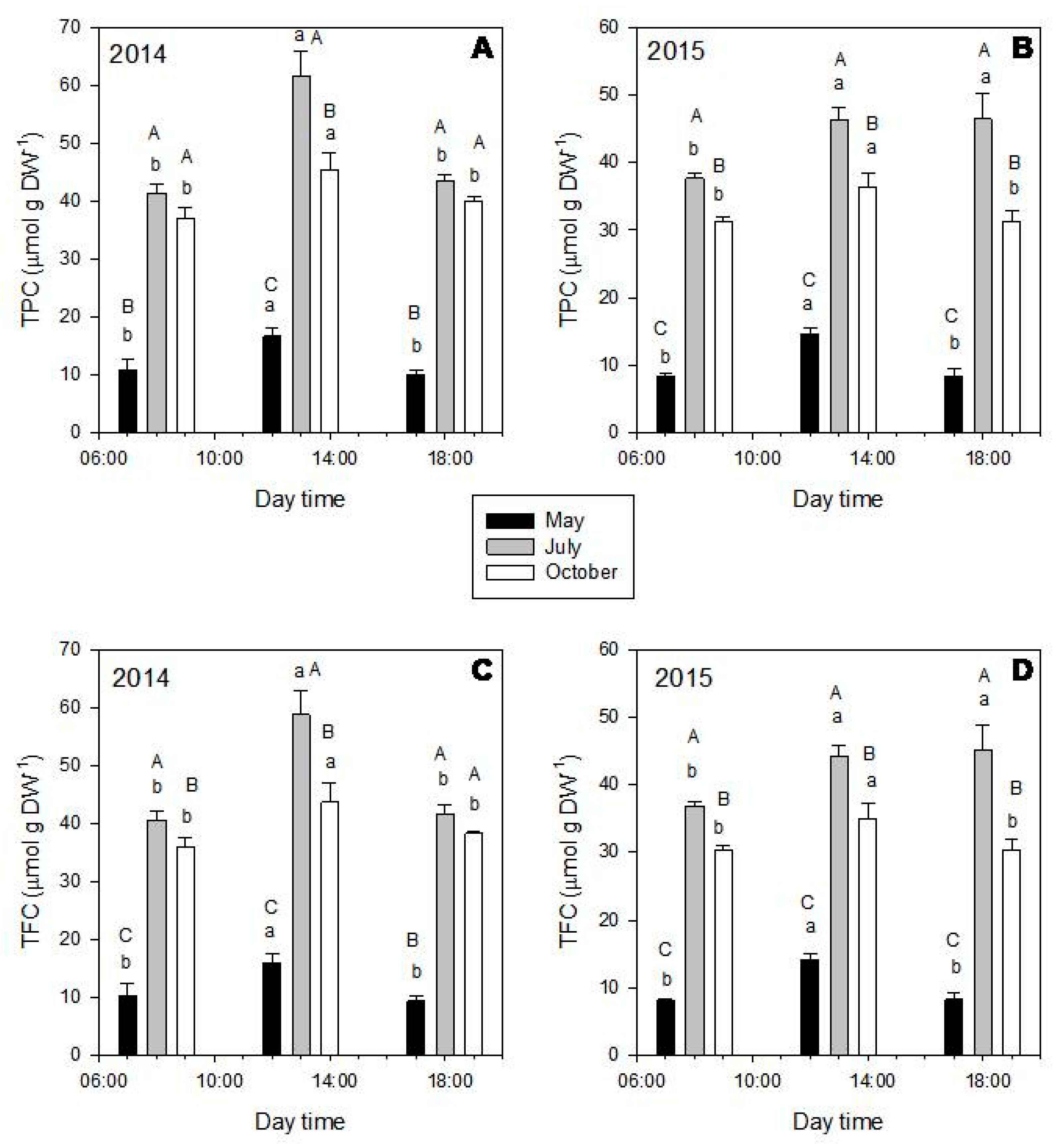
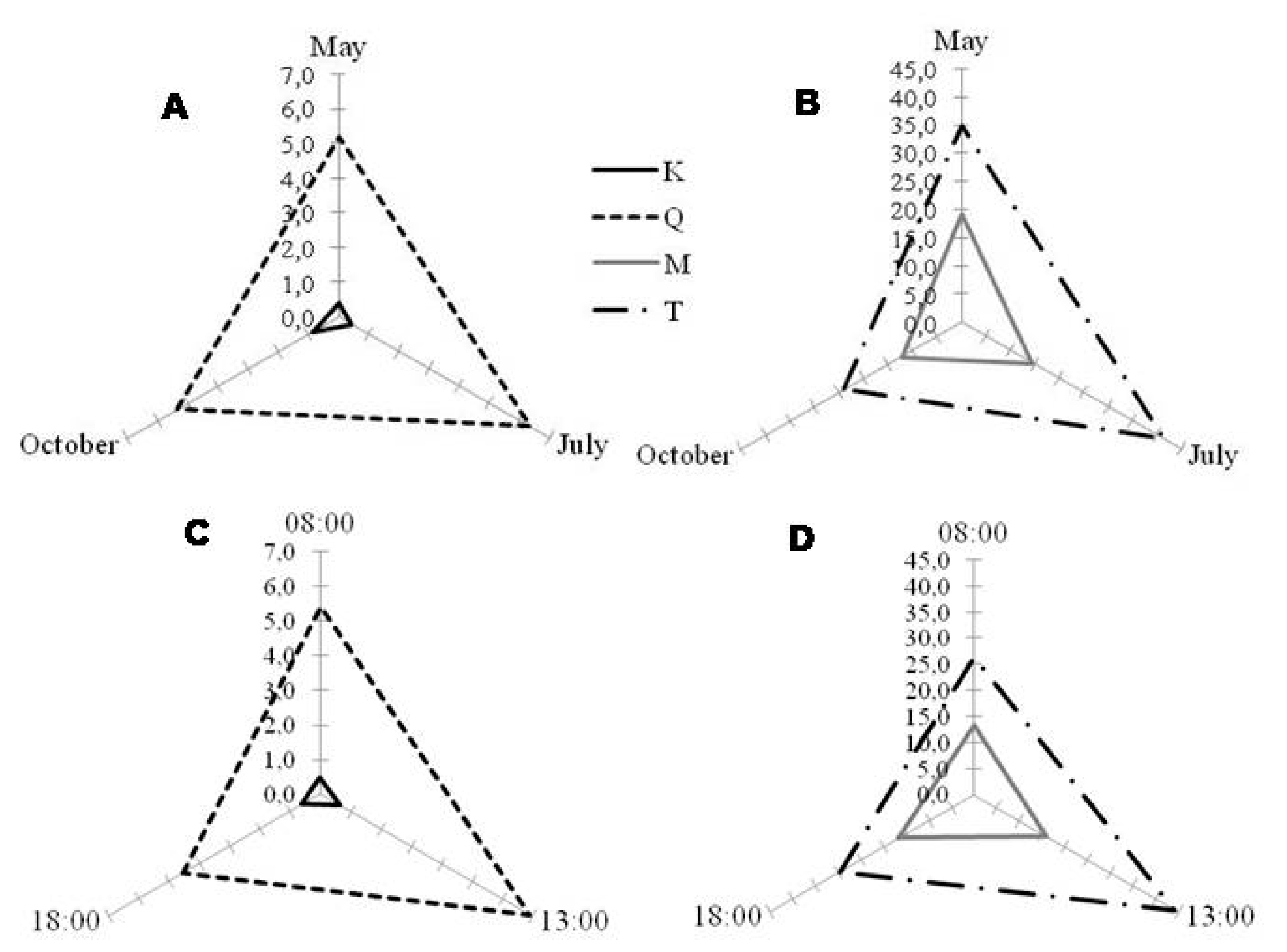
2.2. Seasonal and Diurnal Variation in Polyphenolic Composition
2.3. Correlation with Environmental Parameters
2.4. Antioxidant Capacity of the Extracts
3. Material and Methods
3.1. Plant Material, Study Area and Meteorological Data
3.2. Sampling and Biochemical Analyses
3.3. Antioxidant Capacity Assays
3.4. Statistical Analysis and Correlation with Meteorological Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853. [Google Scholar] [CrossRef] [PubMed]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Galli, C. Bioactive components in Mediterranean diets. Nutrafoods 2012, 11, 11–17. [Google Scholar] [CrossRef]
- Gurib-Fakim, A. Novel Plant Bioresources: Applications in Food, Medicine and Cosmetics. John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Soni, U.; Brar, S.; Gauttam, V.K. Effect of seasonal variation on secondary metabolites of medicinal plants. Int. J. Pharm. Sci. Res. 2015, 6, 3654–3662. [Google Scholar]
- Gori, A.; Tattini, M.; Centritto, M.; Ferrini, F.; Marino, G.; Mori, J.; Guidi, L.; Brunetti, C. Seasonal and daily variations in primary and secondary metabolism of three maquis shrubs unveil different adaptive responses to Mediterranean climate. Conserv. Physiol. 2019, 7. [Google Scholar] [CrossRef]
- Baskar, V.; Venkatesh, R.; Ramalingam, S. Flavonoids (antioxidant systems) in higher plants and their response to stresses. In Antioxidants and Antioxidant Enzymes in Higher Plants; Springer: Cham, Switzerland, 2018; pp. 253–268. [Google Scholar]
- Winkel-Shirley, B. Flavonoid biosynthesis: “New” functions for an “old” pathway. Trends Plant. Sci. 1996, 1, 377–382. [Google Scholar]
- Brunetti, C.; Di Ferdinando, M.; Fini, A.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants and developmental regulators: Relative significance in plants and humans. Int. J. Mol. Sci. 2013, 14, 3540–3555. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P. Polyphenols and human health: Prevention of disease and mechanisms of action. Nutrients 2010, 2, 1106–1131. [Google Scholar] [CrossRef] [Green Version]
- Fraga, C.G.; Croft, K.D.; Kennedy, D.O.; Tomás-Barberán, F.A. The effects of polyphenols and other bioactives on human health. Food Funct. 2019, 10, 514–528. [Google Scholar] [CrossRef] [Green Version]
- Lahlou, M. Screening of natural products for drug discovery. Expert Opin. Drug. Discov. 2007, 2, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Gori, A.; Ferrini, F.; Marzano, M.C.; Tattini, M.; Centritto, M.; Baratto, M.C.; Pogni, R.; Brunetti, C. Characterization and antioxidant activity of crude extract and polyphenolic rich fractions from C. incanus leaves. Int J. Mol. Sci. 2016, 17, 1344. [Google Scholar] [CrossRef] [PubMed]
- Piccolella, S.; Crescente, G.; Pacifico, F.; Pacifico, S. Wild aromatic plants bioactivity: A function of their (poly) phenol seasonality? A case study from Mediterranean area. Phytochem. Rev. 2018, 17, 785–799. [Google Scholar] [CrossRef]
- Raimundo, J.R.; Frazão, D.F.; Domingues, J.L.; Quintela-Sabaris, C.; Dentinho, T.P.; Anjos, O.; Delgado, F. Neglected Mediterranean plant species are valuable resources: The example of Cistus ladanifer. Planta 2018, 248, 1351–1364. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Rollinger, J.M. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceccanti, C.; Landi, M.; Benvenuti, S.; Pardossi, A.; Guidi, L. Mediterranean wild edible plants: Weeds or “new functional crops”? Molecules 2018, 23, 2299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayes, S.; Massawe, F.J.; Alderson, P.G.; Roberts, J.A.; Azam-Ali, S.N.; Hermann, M. The potential for underutilized crops to improve security of food production. J. Exp. Bot. 2011, 63, 1075–1079. [Google Scholar] [CrossRef] [Green Version]
- Bruni, R.; Sacchetti, G. Factors affecting polyphenol biosynthesis in wild and field grown St. John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae). Molecules 2009, 14, 682–725. [Google Scholar] [CrossRef] [Green Version]
- Szakiel, A.; Pączkowski, C.; Henry, M. Influence of environmental abiotic factors on the content of saponins in plants. Phytochem. Rev. 2011, 10, 471–491. [Google Scholar] [CrossRef]
- Borges, C.V.; Minatel, I.O.; Gomez-Gomez, H.A.; Lima, G.P.P. Medicinal plants: Influence of environmental factors on the content of secondary metabolites. In Medicinal Plants and Environmental Challenges; Springer: Cham, Switzerland, 2017; pp. 259–277. [Google Scholar]
- Carretero, C.R.; Lanza, A.M.; Matellano, L.F.; Sánchezb, A.R.; Castillo, L.V. Phytochemical analysis of Phillyrea latifolia L., a new source of oleuropeoside. J. Biosci. 2001, 56, 353356. [Google Scholar] [CrossRef]
- Riehle, P.; Rusche, N.; Saake, B.; Rohn, S. Influence of the leaf content and herbal particle size on the presence and extractability of quantitated phenolic compounds in Cistus incanus herbal teas. J. Agric. Food Chem. 2014, 62, 10978–10988. [Google Scholar] [CrossRef] [PubMed]
- Remila, S.; Atmani-Kilani, D.; Delemasure, S.; Connat, J.L.; Azib, L.; Richard, T.; Atmani, D. Antioxidant, cytoprotective, anti-inflammatory and anticancer activities of Pistacia lentiscus (Anacardiaceae) leaf and fruit extracts. Eur. J. Integr. Med. 2015, 7, 274–286. [Google Scholar] [CrossRef]
- Agnieszka Stępień, A.; David Aebisher, D.; Dorota Bartusik-Aebisher, D. Biological properties of Cistus species. Eur. J. Clin. Exp. Med. 2018, 2, 27–132. [Google Scholar]
- Rebensburg, S.; Helfer, M.; Schneider, M.; Koppensteiner, H.; Eberle, J.; Schindler, M.; Brack-Werner, R. Potent in vitro antiviral activity of Cistus incanus extract against HIV and Filoviruses targets viral envelope proteins. Sci. Rep. 2016, 6, 20394. [Google Scholar] [CrossRef]
- Kalus, U.; Grigorov, A.; Kadecki, O.; Jansen, J.P.; Kiesewetter, H.; Radtke, H. Cistus incanus (CYSTUS052) for treating patients with infection of the upper respiratory tract: A prospective, randomised, placebo-controlled clinical study. Antivir. Res. 2009, 84, 267–271. [Google Scholar] [CrossRef]
- Kuchta, A.; Konopacka, A.; Waleron, K.; Viapiana, A.; Wesołowski, M.; Dąbkowski, K.; Gliwińska, A. The effect of Cistus incanus herbal tea supplementation on oxidative stress markers and lipid profile in healthy adults. Cardiol. J. 2019. [Google Scholar] [CrossRef] [Green Version]
- Cacak-Pietrzak, G.; Różyło, R.; Dziki, D.; Gawlik-Dziki, U.; Sułek, A.; Biernacka, B. Cistus incanus L. as an innovative functional additive to wheat bread. Foods 2019, 8, 349. [Google Scholar] [CrossRef] [Green Version]
- Hadjimbei, E.; Botsaris, G.; Goulas, V.; Gekas, V. Health-promoting effects of Pistacia resins: Recent advances, challenges, and potential applications in the food industry. Food Rev. Int. 2015, 31, 1–12. [Google Scholar] [CrossRef]
- Paraskevopoulou, A.; Kiosseoglou, V. Chios mastic gum and its food applications. In Functional Properties of Traditional Foods; Springer: Boston, MA, USA, 2016; pp. 271–287. [Google Scholar]
- Romani, A.; Pinelli, P.; Galardi, C.; Mulinacci, N.; Tattini, M. Identification and quantification of galloyl derivatives, flavonoid glycosides and anthocyanins in leaves of Pistacia lentiscus L. Phytochem. Anal. 2002, 13, 79–86. [Google Scholar] [CrossRef]
- Azaizeh, H.; Halahleh, F.; Abbas, N.; Markovics, A.; Muklada, H.; Ungar, E.D.; Landau, S.Y. Polyphenols from Pistacia lentiscus and Phillyrea latifolia impair the exsheathment of gastro-intestinal nematode larvae. Vet. Parasitol. 2013, 191, 44–50. [Google Scholar] [CrossRef]
- Debbabi, H.; Nemri, K.; Riahi, H. Antimicrobial effects of Pistacia lentiscus L. foliar extracts on fresh turkey breast cutlets. J. New Sci. 2017, 40, 2144–2152. [Google Scholar]
- Azib, L.; Debbache-Benaida, N.; Da Costa, G.; Atmani-Kilani, D.; Saidene, N.; Ayouni, K.; Atmani, D. Pistacia lentiscus L. leaves extract and its major phenolic compounds reverse aluminium-induced neurotoxicity in mice. Ind. Crop. Prod. 2019, 137, 576–584. [Google Scholar] [CrossRef]
- Moeini, R.; Memariani, Z.; Asadi, F.; Bozorgi, M.; Gorji, N. Pistacia genus as a potential source of neuroprotective natural products. Planta Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Janakat, S.; Al-Merie, H. Evaluation of hepatoprotective effect of Pistacia lentiscus, Phillyrea latifolia and Nicotiana glauca. J. Ethnopharmacol 2009, 83, 135–138. [Google Scholar] [CrossRef]
- Yazici-Tutunis, S.; Gurel-Gurevin, E.; Ustunova, S.; Demirci-Tansel, C.; Mericli, F. Possible effects of Phillyrea latifolia on weight loss in rats fed a high-energy diet. Pharm. Biol. 2016, 54, 1991–1997. [Google Scholar] [CrossRef] [Green Version]
- Lanza, A.M.D.; Martínez, M.J.; Matellano, L.F.; Carretero, C.R.; Castillo, L.V.; Sen, A.M.S.; Benito, P.B. Lignan and phenylpropanoid glycosides from Phillyrea latifolia and their in vitro anti-inflammatory activity. Planta Med. 2001, 67, 219–223. [Google Scholar] [CrossRef]
- Danne, A.; Petereit, F.; Nahrstedt, A. Proanthocyanidins from Cistus incanus. Phytochemistry 1993, 34, 1129–1133. [Google Scholar] [CrossRef]
- Petereit, F.; Kolodziej, H.; Nahrstedt, A. Flavan-3-ols and proanthocyanidins from the herb of Cistus incanus. Planta Med. 1990, 56, 576. [Google Scholar] [CrossRef]
- Djidel, S.; Bouaziz, A.; Bentehar, A.; Khennouf, S.; Baghiani, A.; Dahamna, S.; Amira, S. Effect of methanol extract prepared from leaf of Pistacia lentiscus on plasma antioxidant activity and biomarkers of oxidative stress in liver tissue of healthy rats. Ann. Res. Rev. Biol. 2018, 1–10. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, C.; Quirantes-Piné, R.; Amessis-Ouchemoukh, N.; Madani, K.; Segura-Carretero, A.; Fernández-Gutierrez, A. A metabolite-profiling approach allows the identification of new compounds from Pistacia lentiscus leaves. J. Pharm. Biomed. Anal. 2013, 77, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant. Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Barrajón-Catalán, E.; Fernández-Arroyo, S.; Roldán, C.; Guillén, E.; Saura, D.; Segura-Carretero, A.; Micol, V. A systematic study of the polyphenolic composition of aqueous extracts deriving from several Cistus genus species: Evolutionary relationship. Phytochem. Anal. 2011, 22, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Santagati, N.A.; Salerno, L.; Attaguile, G.; Savoca, F.; Ronsisvalle, G. Simultaneous determination of catechins, rutin, and gallic acid in Cistus species extracts by HPLC with diode array detection. J. Chrom. Sci. 2008, 46, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.W.; Tobler, M.A.; Keefover-Ring, K.; Flint, S.D.; Barkley, A.E.; Ryel, R.J.; Lindroth, R.L. Rapid modulation of ultraviolet shielding in plants is influenced by solar ultraviolet radiation and linked to alterations in flavonoids. Plant. Cell Env. 2016, 39, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Tattini, M.; Gravano, E.; Pinelli, P.; Mulinacci, N.; Romani, A. Flavonoids accumulate in leaves and glandular trichomes of Phillyrea latifolia exposed to excess solar radiation. New Phytol. 2000, 148, 69–77. [Google Scholar] [CrossRef]
- Agati, G.; Galardi, C.; Gravano, E.; Romani, A.; Tattini, M. Flavonoid distribution in tissues of Phillyrea latifolia L. leaves as estimated by microspectrofluorometry and multispectral fluorescence microimaging. Photochem. Photobiol. 2002, 76, 350–360. [Google Scholar] [CrossRef]
- Di Ferdinando, M.; Brunetti, C.; Fini, A.; Tattini, M. Flavonoids as antioxidants in plants under abiotic stresses. In Abiotic stress responses in plants; Springer: New York, NY, USA, 2012; pp. 159–179. [Google Scholar]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus x acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef]
- Agati, G.; Tattini, M. Multiple functional roles of flavonoids in photoprotection. New Phytol. 2010, 186, 786–793. [Google Scholar] [CrossRef]
- Hättenschwiler, S.; Vitousek, P.M. The role of polyphenols in terrestrial ecosystem nutrient cycling. Trends Ec. 2000, 15, 238–243. [Google Scholar] [CrossRef]
- Chomel, M.; Guittonny-Larchevêque, M.; Fernandez, C.; Gallet, C.; DesRochers, A.; Paré, D.; Baldy, V. Plant secondary metabolites: A key driver of litter decomposition and soil nutrient cycling. J. Ecol 2016, 104, 1527–1541. [Google Scholar] [CrossRef]
- Castells, E.; Peñuelas, J.; Valentine, D.W. Are phenolic compounds released from the Mediterranean shrub Cistus albidus responsible for changes in N cycling in siliceous and calcareous soils? New Phytol. 2004, 162, 187–195. [Google Scholar] [CrossRef]
- Di Ferdinando, M.; Brunetti, C.; Agati, G.; Tattini, M. Multiple functions of polyphenols in plants inhabiting unfavorable Mediterranean areas. Env. Exp. Bot. 2014, 103, 107–116. [Google Scholar] [CrossRef]
- Bautista, I.; Boscaiu, M.; Lidón, A.; Llinares, J.V.; Lull, C.; Donat, M.P.; Vicente, O. Environmentally induced changes in antioxidant phenolic compounds levels in wild plants. Acta Physiol. Plant. 2016, 38, 9. [Google Scholar] [CrossRef]
- Ahmed, B.Z.; Yousfi, M.; Viaene, J.; Dejaegher, B.; Demeyer, K.; Mangelings, D.; Vander Heyden, Y. Seasonal, gender and regional variations in total phenolic, flavonoid, and condensed tannins contents and in antioxidant properties from Pistacia atlantica ssp. leaves. Pharm. Biol. 2017, 55, 1185–1194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Yang, M.; Gao, J.; Jin, S.; Wu, Z.; Wu, L.; Zhang, X. Seasonal variation and gender pattern of phenolic and flavonoid contents in Pistacia chinensis Bunge inflorescences and leaves. J. Plant. Physiol. 2016, 191, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Pandey, M.M.; Khatoon, S.; Rastogi, S.; Rawat, A.K.S. Determination of flavonoids, polyphenols and antioxidant activity of Tephrosia purpurea: A seasonal study. J. Integr. Med. 2016, 14, 447–455. [Google Scholar] [CrossRef]
- Schulze, E.D.; Beck, E.; Müller-Hohenstein, K. Plant. Ecology; Springer: New York, NY, USA, 2002. [Google Scholar]
- Katerova, Z.; Todorova, D.; Sergiev, I. Plant secondary metabolites and some plant growth regulators elicited by UV irradiation, light and/or shade. In Medicinal Plants and Environmental Challenges; Springer: Cham, Switzerland, 2017; pp. 97–121. [Google Scholar]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant. Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant. Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental stress and secondary metabolites in plants: An overview. In Plant Metabolites and Regulation Under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 153–167. [Google Scholar]
- Verma, N.; Shukla, S. Impact of various factors responsible for fluctuation in plant secondary metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Jain, R.; Urwin, P.E. The response of plants to simultaneous biotic and abiotic stress. In Combined stresses in plants; Springer: Cham, Switzerland, 2015; pp. 181–201. [Google Scholar]
- Musa, K.H.; Abdullah, A.; Al-Haiqi, A. Determination of DPPH free radical scavenging activity: Application of artificial neural networks. Food Chem. 2016, 194, 705–711. [Google Scholar] [CrossRef]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Musa, K.H.; Abdullah, A.; Kuswandi, B.; Hidayat, M.A. A novel high throughput method based on the DPPH dry reagent array for determination of antioxidant activity. Food Chem. 2013, 141, 4102–4106. [Google Scholar] [CrossRef] [PubMed]
- Jacobo-Velázquez, D.A.; Cisneros-Zevallos, L. Correlations of antioxidant activity against phenolic content revisited: A new approach in data analysis for food and medicinal plants. J. Food Sci. 2009, 74, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. Rsc. Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Grajeda-Iglesias, C.; Salas, E.; Barouh, N.; Baréa, B.; Panya, A.; Figueroa-Espinoza, M.C. Antioxidant activity of protocatechuates evaluated by DPPH, ORAC, and CAT methods. Food Chem. 2016, 194, 749–757. [Google Scholar] [CrossRef]
Sample Availability: Samples of the ethanolic extracts of the three species are available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Composition | Temperature | Solar Irradiance |
|---|---|---|
| P. latifolia | ||
| K | 0.26 | 0.10 |
| A | 0.12 | 0.20 |
| CA | 0.54* | 0.35 |
| Q | 0.52* | 0.05 |
| L | 0.65** | 0.14 |
| TFC | 0.59** | 0.11 |
| TPC | 0.59** | 0.12 |
| C. incanus | ||
| K | -0.28 | -0.17 |
| M | -0.02 | 0.13 |
| Q | 0.46* | 0.67** |
| T | 0.45* | 0.49* |
| TFC | 0.12 | 0.32 |
| TPC | 0.43 | 0.51* |
| P. lentiscus | ||
| M | 0.18 | -0.07 |
| Q | 0.09 | 0.45 |
| T | 0.17 | 0.25 |
| TFC | 0.17 | 0.05 |
| TPC | 0.19 | 0.21 |
| May | July | October | ||||
|---|---|---|---|---|---|---|
| DPPH | HRS | DPPH | HRS | DPPH | HRS | |
| P. latifolia | 9.6 ± 1.0 a | 0.47 ± 0.05 a | 2.2 ± 0.4 b | 0.12 ± 0.02 b | 1.2 ± 0.2 c | 0.23 ± 0.03 c |
| C. incanus | 0.74 ± 0.12 a | 0.22 ± 0.03 a | 0.53 ± 0.08 a | 0.24 ± 0.04 a | 0.53 ± 0.05 a | 0.24 ± 0.03 a |
| P. lentiscus | 2.7 ± 0.4 a | 0.85 ± 0.11 a | 2.8 ± 0.3 a | 0.82 ± 0.13 a | 2.8 ± 0.4 a | 0.81 ± 0.10 a |
| Month and Year of Harvesting (season) | Phenological Stage | Meteorological Conditions | ||||
|---|---|---|---|---|---|---|
| Cistus incanus | Phillyrea latifolia | Pistacia lentiscus | Monthly Cumulative Rainfall (mm) | Daily Temperature—Mean Values (°C) | Daily Global Solar Irradiance—Mean Values (W m-2) | |
| 2014 | ||||||
| May (Spring) | Blooming | Blooming | Blooming | 48.0 | 17.8 ± 3.0 | 919.4 ± 75.6 |
| July (Summer) | Fructification (End) | Fructification (Beginning) | Fructification (Beginning) | 84.0 | 24.2 ± 2.8 | 847.1 ± 73.2 |
| October (Autumn) | Vegetative | Fructification (End) | Fructification | 173.8 | 19.3 ± 3.6 | 611.4 ± 91.7 |
| 2015 | ||||||
| May (Spring) | Blooming | Blooming | Blooming | 53.4 | 20.8 ± 3.6 | 803.8 ± 146.6 |
| July (Summer) | Fructification (End) | Fructification (Beginning) | Fructification (Beginning) | 29.2 | 27.3 ± 2.9 | 857.2 ± 50.9 |
| October (Autumn) | Vegetative | Fructification (End) | Fructification | 70.6 | 20.9 ± 2.0 | 562.7 ± 118.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gori, A.; Nascimento, L.B.; Ferrini, F.; Centritto, M.; Brunetti, C. Seasonal and Diurnal Variation in Leaf Phenolics of Three Medicinal Mediterranean Wild Species: What Is the Best Harvesting Moment to Obtain the Richest and the Most Antioxidant Extracts? Molecules 2020, 25, 956. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040956
Gori A, Nascimento LB, Ferrini F, Centritto M, Brunetti C. Seasonal and Diurnal Variation in Leaf Phenolics of Three Medicinal Mediterranean Wild Species: What Is the Best Harvesting Moment to Obtain the Richest and the Most Antioxidant Extracts? Molecules. 2020; 25(4):956. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040956
Chicago/Turabian StyleGori, Antonella, Luana Beatriz Nascimento, Francesco Ferrini, Mauro Centritto, and Cecilia Brunetti. 2020. "Seasonal and Diurnal Variation in Leaf Phenolics of Three Medicinal Mediterranean Wild Species: What Is the Best Harvesting Moment to Obtain the Richest and the Most Antioxidant Extracts?" Molecules 25, no. 4: 956. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040956