
16-Hydroxycleroda-3,13-Dien-15,16-Olide Induces Apoptosis in Human Bladder Cancer Cells through Cell Cycle Arrest, Mitochondria ROS Overproduction, and Inactivation of EGFR-Related Signalling Pathways


Abstract
:1. Introduction
2. Results
2.1. CD Induces Apoptosis in T24 BC Cells
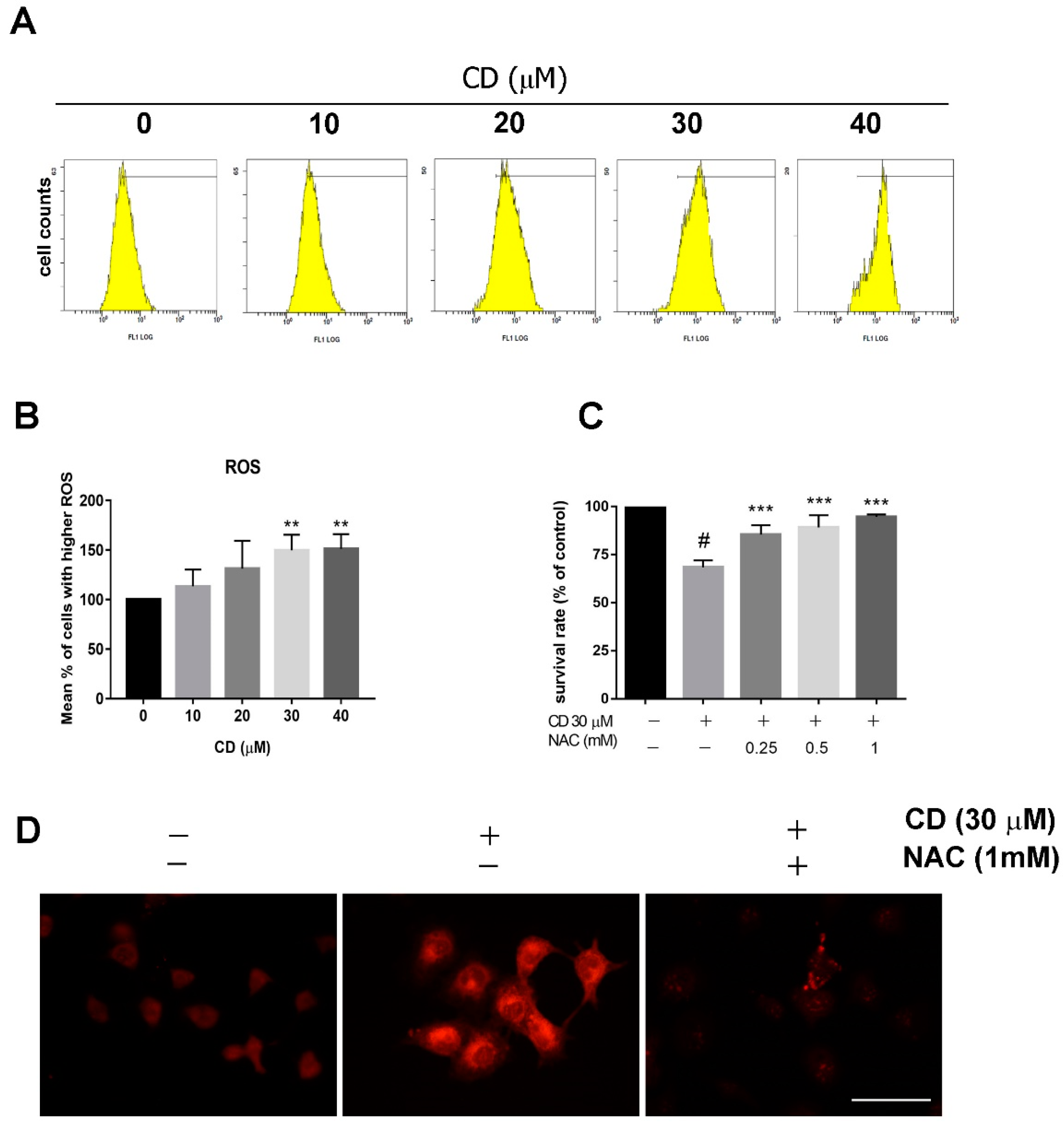
2.2. CD Suppresses MMP and Triggers ROS Production
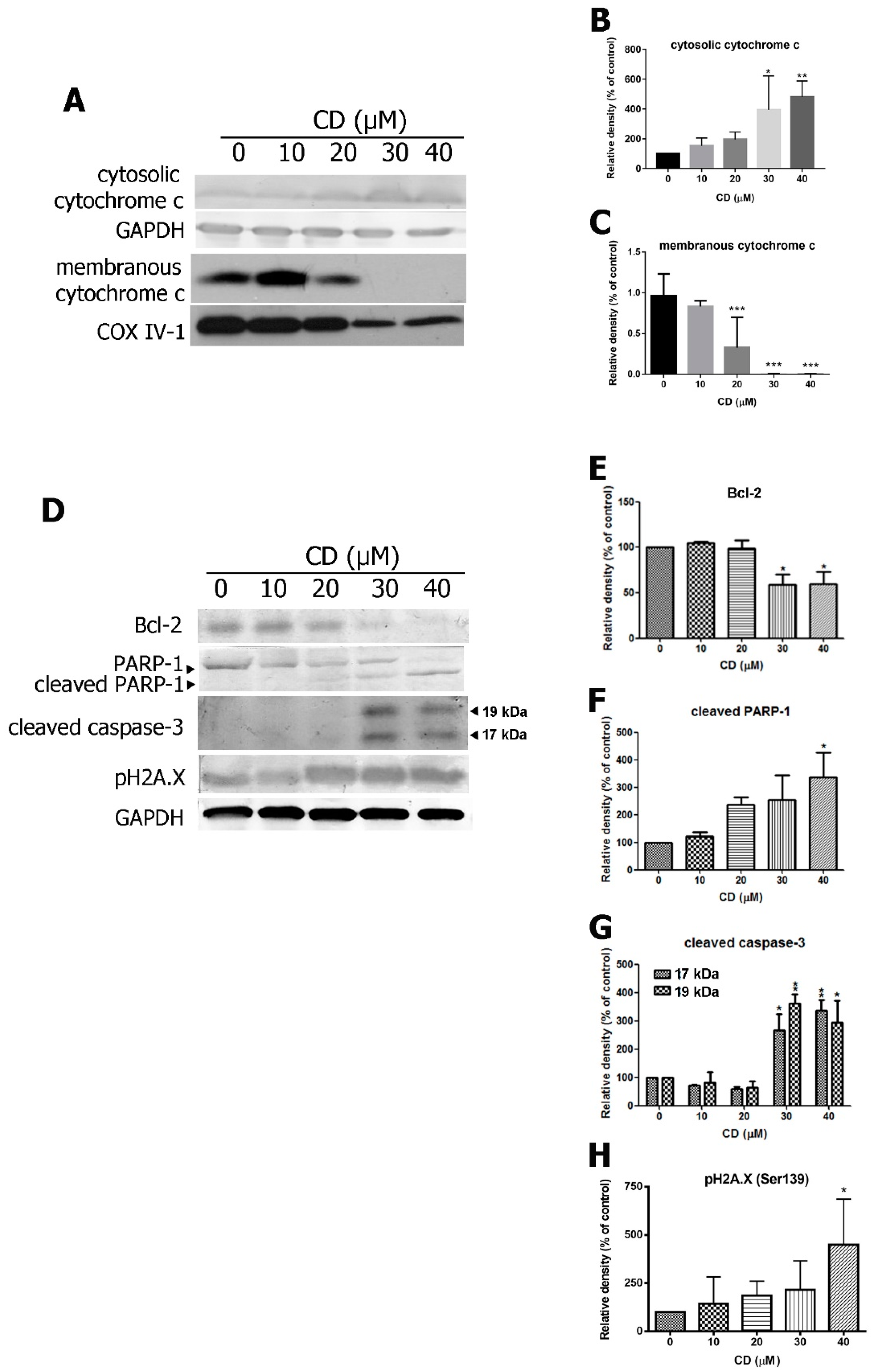
2.3. CD Triggers the Expression of Pro-Apoptotic Proteins and Inhibits Anti-Apoptotic Proteins
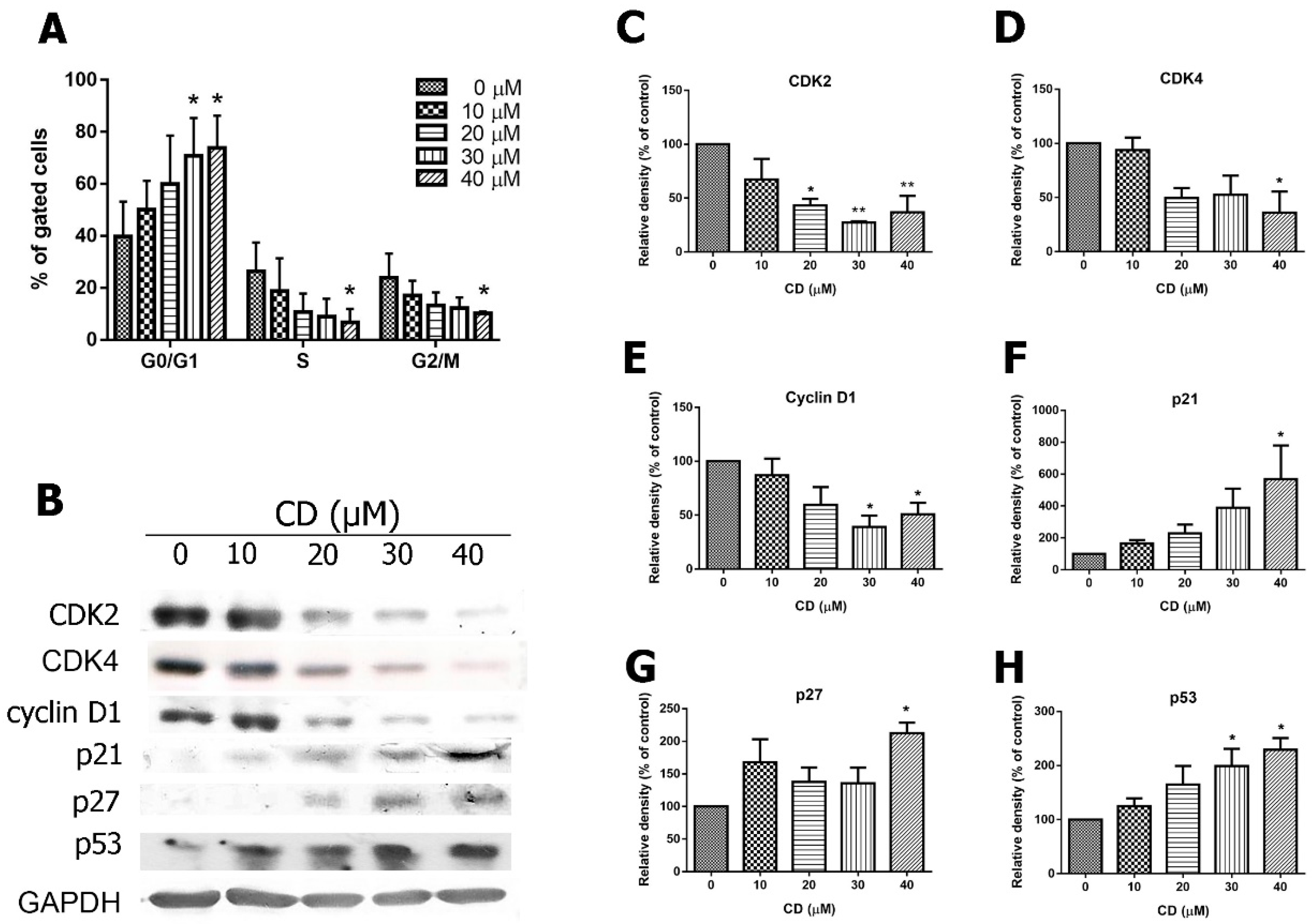
2.4. Effects of CD on Cell Cycle Progression, Cyclins and Cyclin-Dependent Kinases (CDKs)
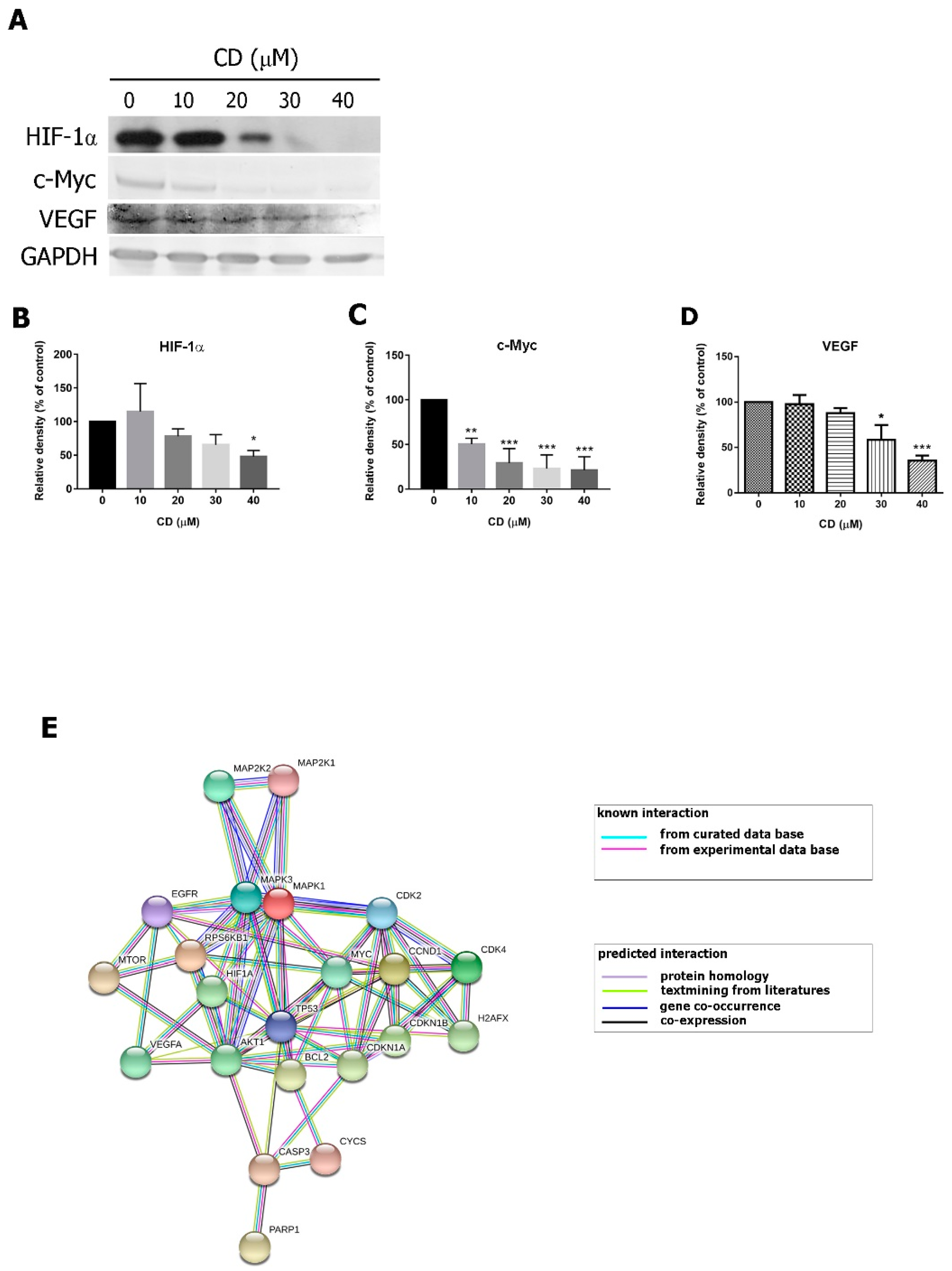
2.5. CD Modulates the Epidermal Growth Factor Receptor-Mediated Signalling Pathway
2.6. Expression Profiling of CD-Triggered Cell Death
3. Discussion
4. Experimental Procedures
4.1. Plant Authentication and Extraction
4.2. Cell Culture and Reagents
4.3. Morphology Observation and Cell Viability Assay
4.4. Acridine Orange and Ethidium Bromide (AO/EB) Double Staining
4.5. Flow Cytometry Analysis for Annexin V- Propidium Iodide (PI), Reactive Oxygen Species (ROS) and Mitochondrial Membrane Potential ΔΨM (MMP)
4.6. Fluorescence Staining for MMP and Mitochondrial ROS
4.7. Cell Cycle Analysis
4.8. Western Blot Analysis
4.9. RNA Extraction and Expression Profiling
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ΔΨM | mitochondrial membrane potential |
| BC | bladder cancer |
| CCCP | carbonyl cyanide 3-chlorophenylhydrazone |
| CD | 16-Hydroxycleroda-3,13-dien-15,16-olide |
| JC-1 | (5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-benzimidazolylcarbocyanine iodide) |
| MMP | mitochondrial membrane potential |
| MTT | [3-(4, 5-dimethylthiazole-2-yl)-2, 5-diphenyl tetrazolium bromide] |
| Rh123 | rhodamine 123 |
| ROS | reactive oxygen species |
References
- Sashidhara, K.V.; Singh, S.P.; Srivastava, A.; Puri, A.; Chhonker, Y.S.; Bhatta, R.S.; Shah, P.; Siddiqi, M.I. Discovery of a new class of HMG-CoA reductase inhibitor from Polyalthia longifolia as potential lipid lowering agent. Eur. J. Med. Chem. 2011, 46, 5206–5211. [Google Scholar] [CrossRef] [PubMed]
- Misra, P.; Sashidhara, K.V.; Singh, S.P.; Kumar, A.; Gupta, R.; Chaudhaery, S.S.; Gupta, S.S.; Majumder, H.K.; Saxena, A.K.; Dube, A. 16 alpha-Hydroxycleroda-3,13 (14)Z-dien-15,16-olide from Polyalthia longifolia: A safe and orally active antileishmanial agent. Br. J. Pharm. 2010, 159, 1143–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marthanda Murthy, M.; Subramanyam, M.; Hima Bindu, M.; Annapurna, J. Antimicrobial activity of clerodane diterpenoids from Polyalthia longifolia seeds. Fitoterapia 2005, 76, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.H.; Cheng, Y.Y.; Chen, C.J.; Ng, L.T.; Chou, L.C.; Huang, L.J.; Chen, Y.H.; Kuo, S.C.; El-Shazly, M.; Wu, Y.C.; et al. Three new clerodane diterpenes from Polyalthia longifolia var. pendula. Molecules 2014, 19, 2049–2060. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.F.; Lin, S.R.; Tseng, F.J.; Huang, Y.C.; Tsai, M.J.; Fu, Y.S.; Weng, C.F. The autophagic inhibition oral squamous cell carcinoma cancer growth of 16-hydroxy-cleroda-3,14-dine-15,16-olide. Oncotarget 2017, 8, 78379–78396. [Google Scholar] [CrossRef] [Green Version]
- Katkar, K.V.; Suthar, A.C.; Chauhan, V.S. The chemistry, pharmacologic, and therapeutic applications of Polyalthia longifolia. Pharm. Rev. 2010, 4, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Sari, D.P.; Ninomiya, M.; Efdi, M.; Santoni, A.; Ibrahim, S.; Tanaka, K.; Koketsu, M. Clerodane diterpenes isolated from Polyalthia longifolia induce apoptosis in human leukemia HL-60 cells. J. Oleo Sci. 2013, 62, 843–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Lee, W.C.; Huang, B.M.; Chia, Y.C.; Chen, Y.C.; Chen, Y.C. 16-Hydroxycleroda-3, 13-dien-15, 16-olide inhibits the proliferation and induces mitochondrial-dependent apoptosis through Akt, mTOR, and MEK-ERK pathways in human renal carcinoma cells. Phytomed. Int. J. Phytother. Phytopharm. 2017, 36, 95–107. [Google Scholar] [CrossRef]
- Antoni, S.; Ferlay, J.; Soerjomataram, I.; Znaor, A.; Jemal, A.; Bray, F. Bladder Cancer Incidence and Mortality: A Global Overview and Recent Trends. Eur. Urol. 2017, 71, 96–108. [Google Scholar] [CrossRef]
- Chiang, C.J.; Lo, W.C.; Yang, Y.W.; You, S.L.; Chen, C.J.; Lai, M.S. Incidence and survival of adult cancer patients in Taiwan, 2002-2012. J. Med. Assoc. 2016, 115, 1076–1088. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.S.; Bochner, B.H.; Chou, R.; Dreicer, R.; Kamat, A.M.; Lerner, S.P.; Lotan, Y.; Meeks, J.J.; Michalski, J.M.; Morgan, T.M.; et al. Treatment of Nonmetastatic Muscle-Invasive Bladder Cancer: American Urological Association/American Society of Clinical Oncology/American Society for Radiation Oncology/Society of Urologic Oncology Clinical Practice Guideline Summary. J. Oncol. Pract. 2017, 13, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Quan, Y.; Jeong, C.W.; Kwak, C.; Kim, H.H.; Kim, H.S.; Ku, J.H. Dose, duration and strain of bacillus Calmette-Guerin in the treatment of nonmuscle invasive bladder cancer: Meta-analysis of randomized clinical trials. Medicine 2017, 96, e8300. [Google Scholar] [CrossRef] [PubMed]
- Del Bene, G.; Sternberg, C.N. Systemic chemotherapy in muscle invasive and metastatic bladder cancer: Present and future. Urologia 2017, 84, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Voldborg, B.R.; Damstrup, L.; Spang-Thomsen, M.; Poulsen, H.S. Epidermal growth factor receptor (EGFR) and EGFR mutations, function and possible role in clinical trials. Ann. Oncol. Off. J. Eur. Soc. Med Oncol. 1997, 8, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Sigismund, S.; Avanzato, D.; Lanzetti, L. Emerging functions of the EGFR in cancer. Mol. Oncol. 2018, 12, 3–20. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [Green Version]
- Richters, A.; Aben, K.K.H.; Kiemeney, L. The global burden of urinary bladder cancer: An update. World J. Urol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.S. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Okon, I.S.; Zou, M.H. Mitochondrial ROS and cancer drug resistance: Implications for therapy. Pharm. Res. 2015, 100, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Marchi, S.; Giorgi, C.; Suski, J.M.; Agnoletto, C.; Bononi, A.; Bonora, M.; De Marchi, E.; Missiroli, S.; Patergnani, S.; Poletti, F.; et al. Mitochondria-ros crosstalk in the control of cell death and aging. J. Signal Transduct. 2012, 2012, 329635. [Google Scholar] [CrossRef] [Green Version]
- Su, L.J.; Zhang, J.H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.Y. Reactive Oxygen Species-Induced Lipid Peroxidation in Apoptosis, Autophagy, and Ferroptosis. Oxid. Med. Cell Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.F.; Liu, H.; Luo, X.J.; Zhao, Z.; Zou, Z.Y.; Li, J.; Lin, X.J.; Liang, Y. The roles of reactive oxygen species (ROS) and autophagy in the survival and death of leukemia cells. Crit. Rev. Oncol. Hematol. 2017, 112, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.J.; Ho, C.T.; Wang, Y.J. Pterostilbene induces autophagy and apoptosis in sensitive and chemoresistant human bladder cancer cells. Mol. Nutr. Food Res. 2010, 54, 1819–1832. [Google Scholar] [CrossRef]
- Hashmi, A.A.; Hussain, Z.F.; Irfan, M.; Khan, E.Y.; Faridi, N.; Naqvi, H.; Khan, A.; Edhi, M.M. Prognostic significance of epidermal growth factor receptor (EGFR) over expression in urothelial carcinoma of urinary bladder. BMC Urol. 2018, 18, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfost, B.; Seidl, C.; Autenrieth, M.; Saur, D.; Bruchertseifer, F.; Morgenstern, A.; Schwaiger, M.; Senekowitsch-Schmidtke, R. Intravesical alpha-radioimmunotherapy with 213Bi-anti-EGFR-mAb defeats human bladder carcinoma in xenografted nude mice. J. Nucl. Med. 2009, 50, 1700–1708. [Google Scholar] [CrossRef] [Green Version]
- Grivas, P.; Yu, E.Y. Role of Targeted Therapies in Management of Metastatic Urothelial Cancer in the Era of Immunotherapy. Curr. Treat. Opt. Oncol. 2019, 20, 67. [Google Scholar] [CrossRef]
- Sathe, A.; Nawroth, R. Targeting the PI3K/AKT/mTOR Pathway in Bladder Cancer. Methods Mol. Biol. 2018, 1655, 335–350. [Google Scholar] [CrossRef]
- Kasibhatla, S.; Amarante-Mendes, G.P.; Finucane, D.; Brunner, T.; Bossy-Wetzel, E.; Green, D.R. Acridine Orange/Ethidium Bromide (AO/EB) Staining to Detect Apoptosis. CSH Protoc. 2006, 2006. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, P.F.; Pattison, A.D.; Powell, D.R.; Beilharz, T.H. Topconfects: A package for confident effect sizes in differential expression analysis provides a more biologically useful ranked gene list. Genome Biol. 2019, 20, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Dilution | Brand |
|---|---|---|
| p27Kip1 (#3698), phospho-70 kDa ribosomal protein S6 kinase 1 (pP70S6K1, Thr389; #9234T), phospho-mitogen activated protein kinase kinase 1/2 (pMEK, Ser 217/221; #9121) | 1:1000 | Cell Signaling Technology Inc. (Danvers, MA, USA) |
| Caspase-3 (sc-56053), CDK2 (sc-163), CDK4 (sc-260), cyclin D1 (sc-8396), phospho-mammalian target of rapamycin (pmTOR, ser2448) (sc-293132), phospho-extracellular regulated kinase 1/2 (pERK1/2, Thr202/Tyr204; sc-7383) vascular endothelial growth factor (VEGF) (sc-152) | 1:1000 1:500 | Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA) |
| Cytochrome oxidase subunit IV isoform 1 (COX IV-1, 11242-1-AP) Glyceraldehyde 3 phosphate dehydrogenase (GAPDH, 60004-1-Ig) | 1:1000 1:2000 | Proteintech Group Inc. (Rosemont, IL, USA) |
| Hypoxia inducible factor 1 alpha (HIF-1α, 2015-S) | 1:1000 | Epitomics, Inc., a brand of Abcam (Burlingame, CA, USA) |
| Cytochrome c (#45-6100) | 1:1000 | Thermo Fisher Scientific Inc. |
| B-cell lymphoma 2 (Bcl-2) (B3170), phospho-histone H2A.X (Ser139) (pH2A.X), ZooMAb® (ZRB05636) | 1:1000 | Sigma |
| Poly [ADP-ribose] polymerase 1, (PARP-1, E12-173) | 1:500 | Enogene Biotech Co., Ltd. (New York, NY, USA) |
| p21 (GTX100444), p53 (GTX102965), c-Myc (GTX103436) | 1:1000 | GeneTex (Irvine, SC, USA) |
| pEGFR (Tyr1173) (ab32578), pAkt1 (Ser473) (ab81283) Akt1 (ab32505), mTOR (ab134903) | 1:1000 | Abcam Plc. (Cambridge, MA, USA) |
| P70S6K1 (ARG51221), EGFR (ARG66204) | 1:1000 | arigo Biolaboratories Corp. (Hsin-Chu, Taiwan) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-C.; Wang, P.-Y.; Huang, B.-M.; Chen, Y.-J.; Lee, W.-C.; Chen, Y.-C. 16-Hydroxycleroda-3,13-Dien-15,16-Olide Induces Apoptosis in Human Bladder Cancer Cells through Cell Cycle Arrest, Mitochondria ROS Overproduction, and Inactivation of EGFR-Related Signalling Pathways. Molecules 2020, 25, 3958. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173958
Chen Y-C, Wang P-Y, Huang B-M, Chen Y-J, Lee W-C, Chen Y-C. 16-Hydroxycleroda-3,13-Dien-15,16-Olide Induces Apoptosis in Human Bladder Cancer Cells through Cell Cycle Arrest, Mitochondria ROS Overproduction, and Inactivation of EGFR-Related Signalling Pathways. Molecules. 2020; 25(17):3958. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173958
Chicago/Turabian StyleChen, Yu-Chi, Po-Yu Wang, Bu-Miin Huang, Yu-Jen Chen, Wei-Chang Lee, and Yung-Chia Chen. 2020. "16-Hydroxycleroda-3,13-Dien-15,16-Olide Induces Apoptosis in Human Bladder Cancer Cells through Cell Cycle Arrest, Mitochondria ROS Overproduction, and Inactivation of EGFR-Related Signalling Pathways" Molecules 25, no. 17: 3958. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25173958