
Effects of Aloe vera Flower Extract and Its Active Constituent Isoorientin on Skin Moisturization via Regulating Involucrin Expression: In Vitro and Molecular Docking Studies


Abstract
:1. Introduction
2. Results
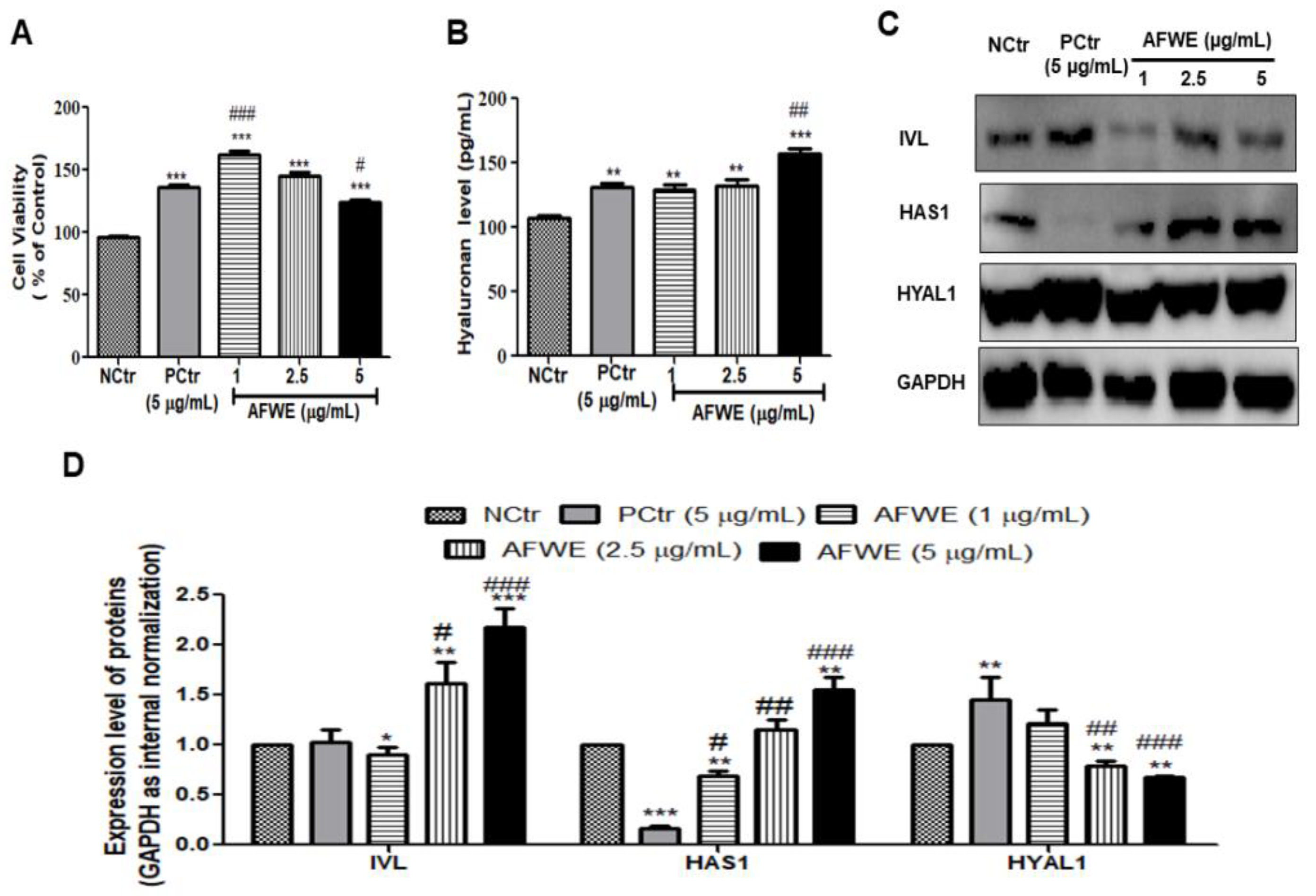
2.1. AFWE Increases IVL Expression as Well as Hyaluronan Secretion by Upregulating HAS 1 and Downregulating HYAL 1 Expression
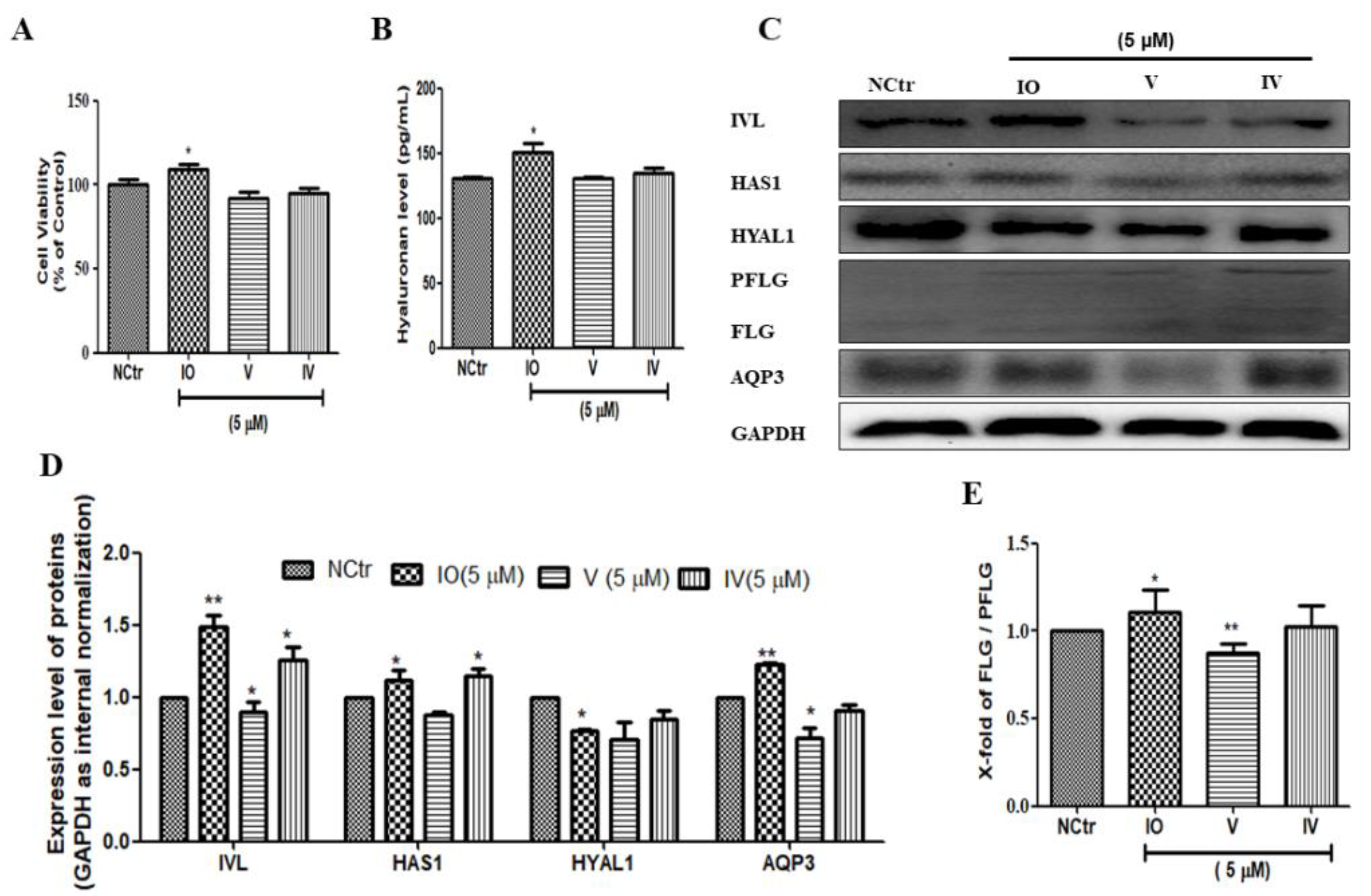
2.2. AFWE Induces FLG Formation and AQP3 Expression
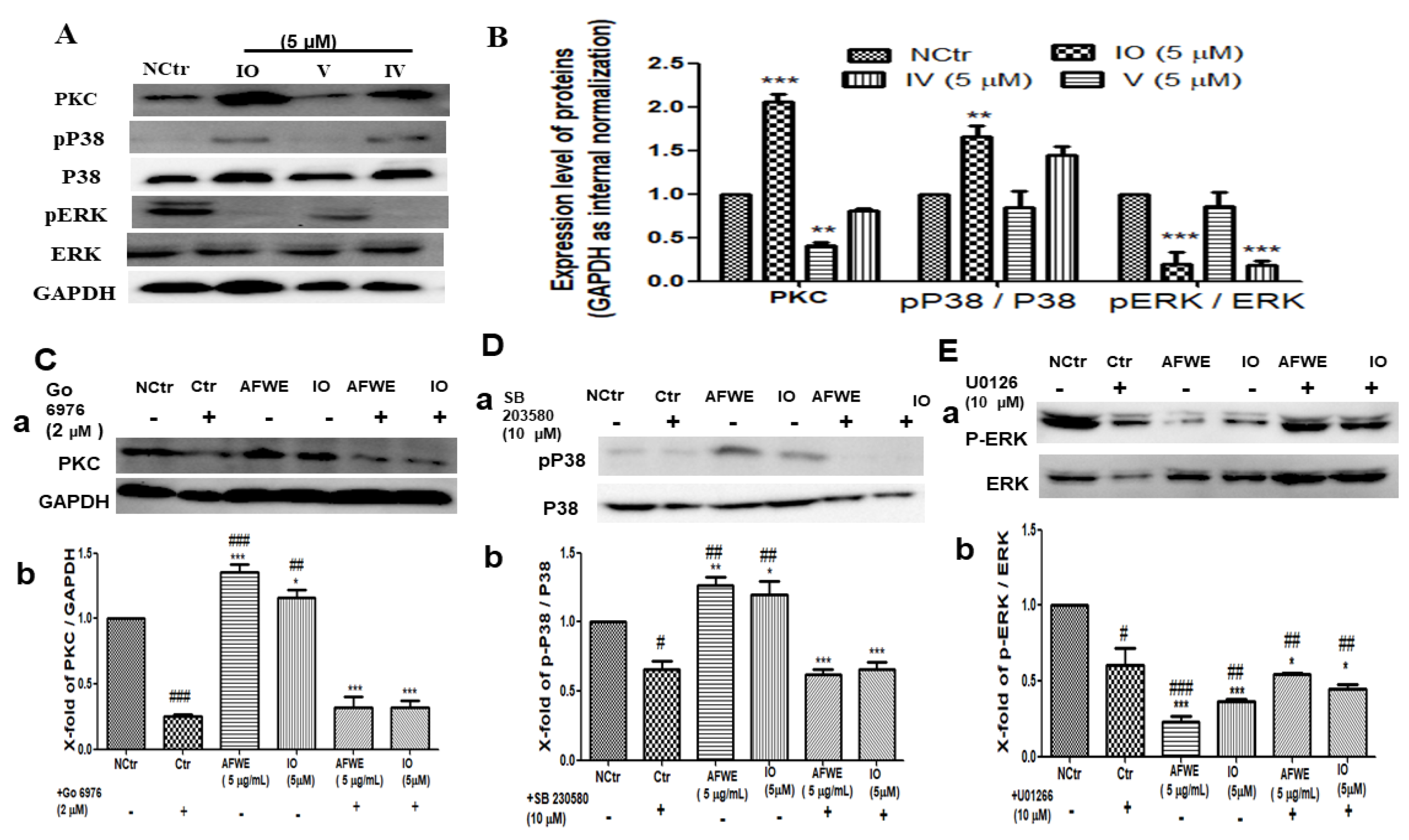
2.3. AFWE Regulates IVL Expression via Activation of PKC and MAPK Signaling Pathway
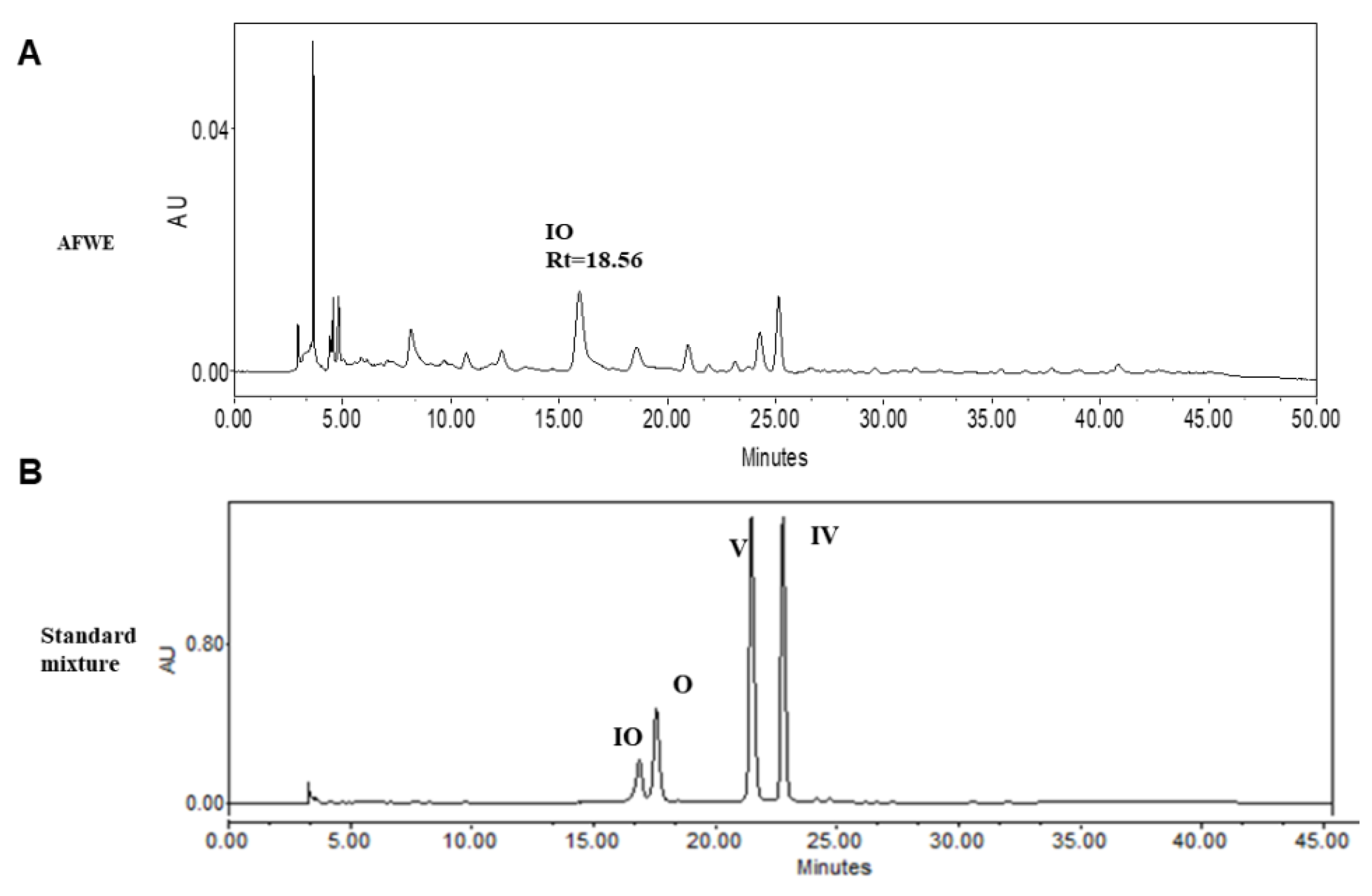
2.4. HPLC Analysis Confirms the Presence of Active Constituents of AFWE Extract
2.5. Isoorietin (IO), an Active Constituent of AFWE Demonstrates Moisturizing Effects
2.6. Isoorietin (IO) Also Activates PKC and MAPK Signaling Pathway
2.7. AFWE and IO Both Confirm the Regulatory Role of PKC and MAPK Signaling in IVL Expression
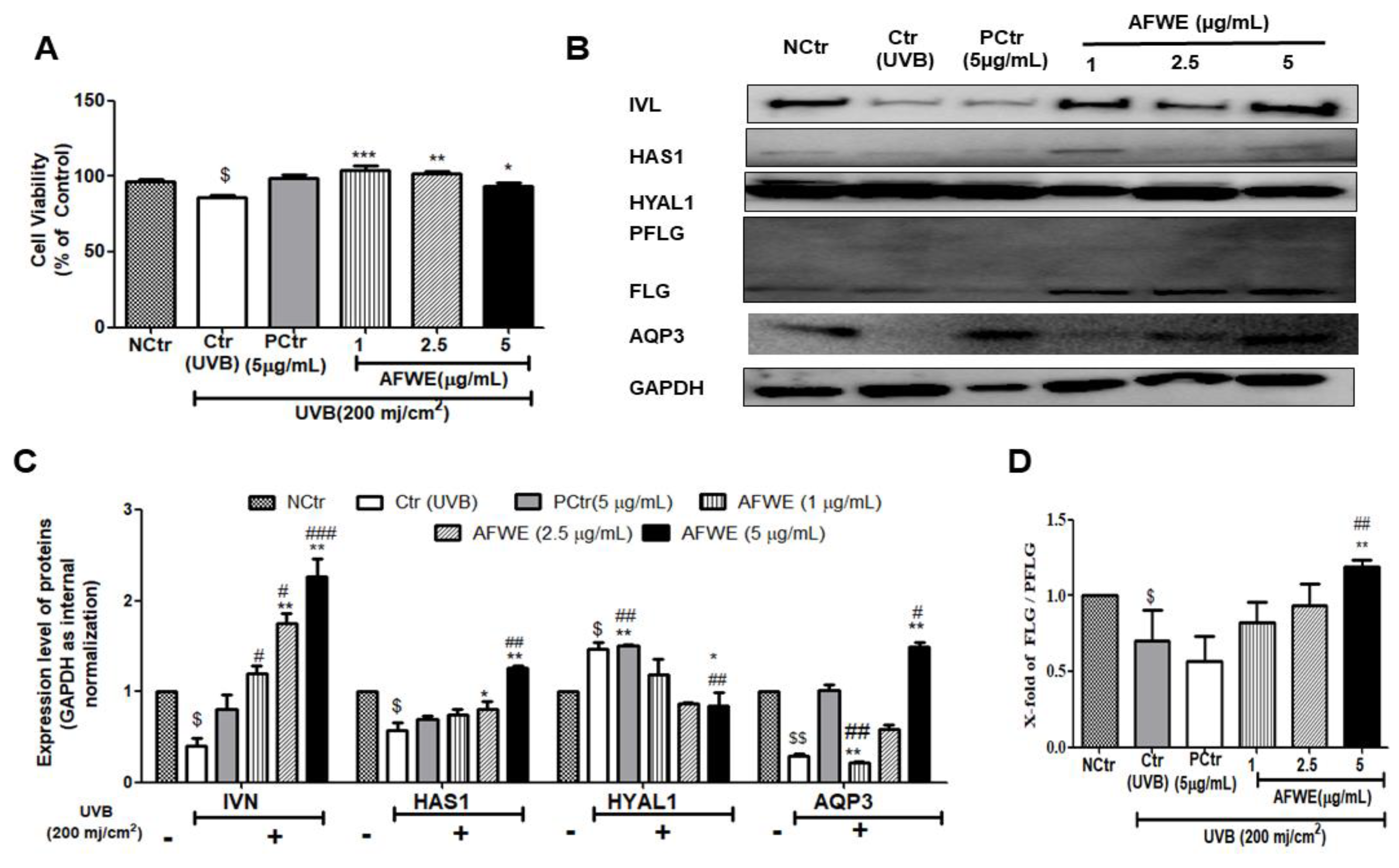
2.8. AFWE Protects UVB-Induced Photodamage in a Pattern Similar to That Observed in Normal Condition
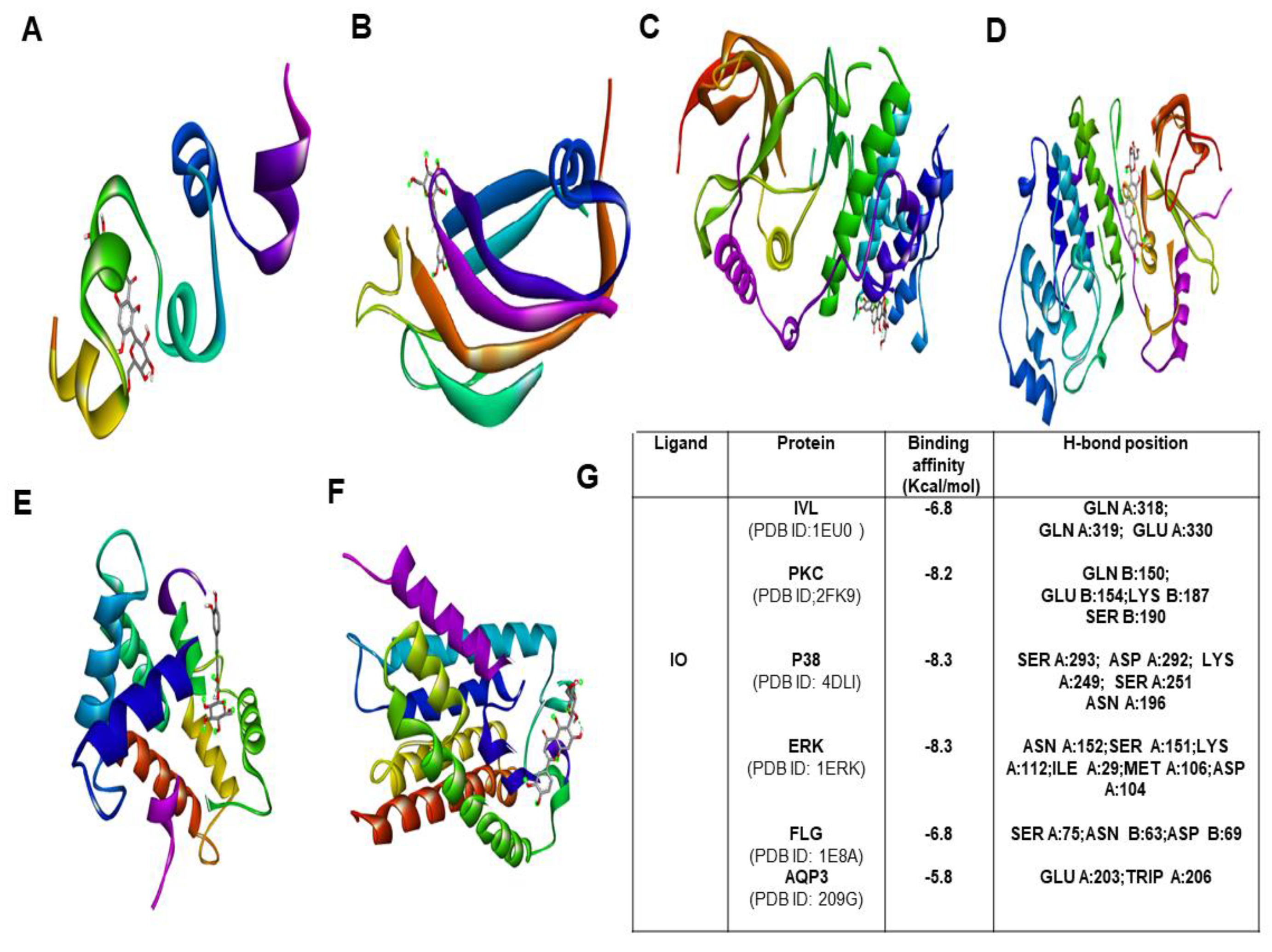
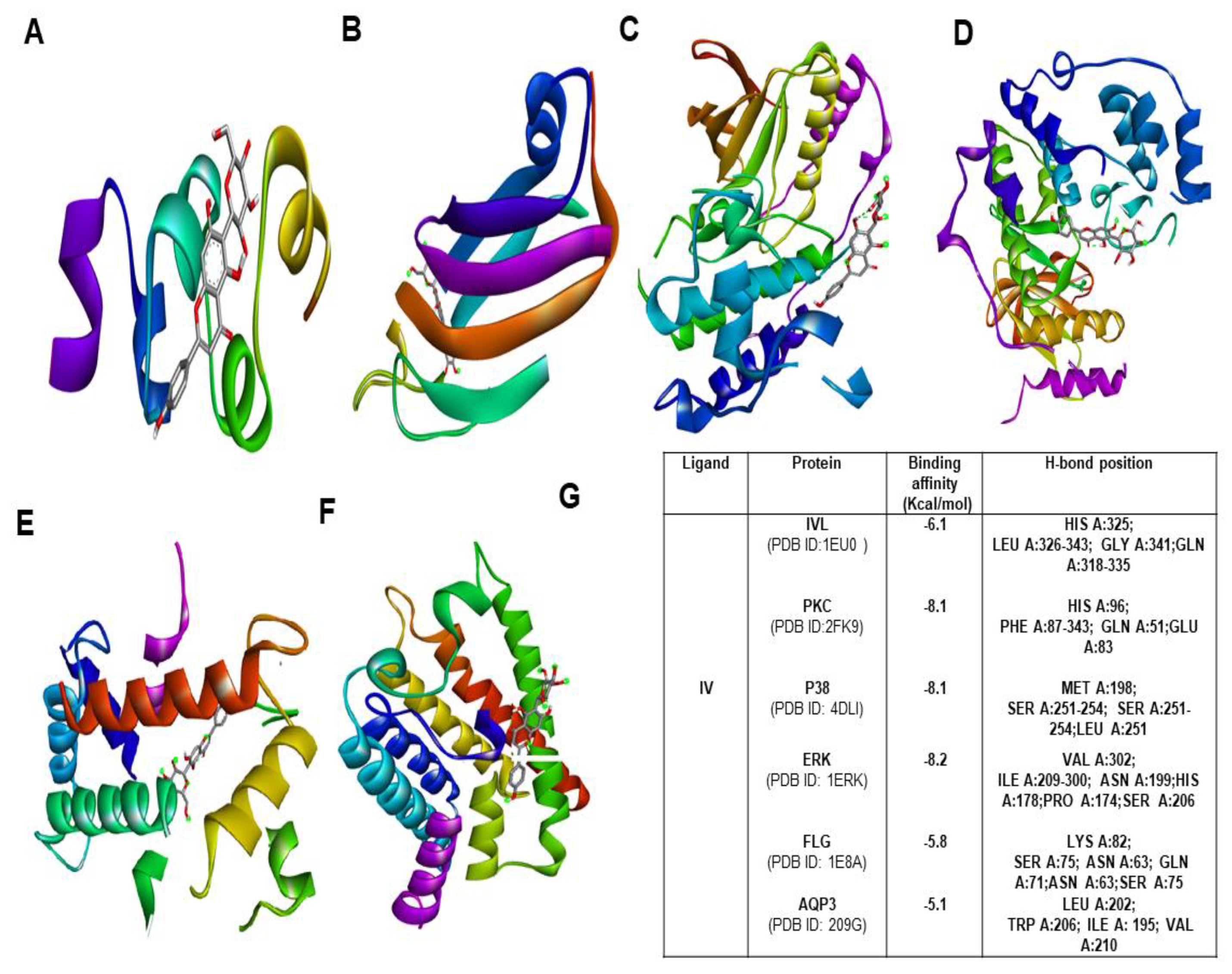
2.9. Molecular Docking Studies
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Extract Preparation
4.3. Cell Culture
4.4. Sample Preparation and Treatment of Cells
4.5. Cell Viability Assay
4.6. Enzyme Linked-Immuno Sorbent Assay (ELISA)
4.7. Western Blotting
4.8. Quantitative Real Time-Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
4.9. Thin-Layer Chromatography (TLC) and HPLC Analysis of Aloe vera Flower Extracts
4.10. Molecular Docking Studies
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| AFWE | A. vera flower water extracts |
| HaCaT cells | human epidermal keratinocytes |
| IO | Isoorientin |
| HPLC | High-performance liquid chromatography |
| IVL | Involucrin |
| Aquaporins | AQP3 |
| HA | Hyaluronic acid |
| PKC | Protein kinase C |
| UVB | Ultraviolet B |
| ELISAs | Enzyme-Linked Immunosorbent Assays |
References
- Yousef, H.; Alhajj, M.; Sharma, S. Anatomy, Skin (Integument), Epidermis. StatPearls Publ. 2020. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/books/NBK470464/ (accessed on 15 January 2021).
- Lee, A.Y. Molecular mechanism of epidermal barrier dysfunction as primary abnormalities. Int. J. Mol. Sci. 2020, 21, 1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdier-Sévrain, S.; Bonté, F. Skin hydration: A review on its molecular mechanisms. J. Cosmet. Dermatol. 2007, 6, 75–82. [Google Scholar] [CrossRef]
- Brettmann, E.A.; de Guzman Strong, C. Recent evolution of the human skin barrier. Exp. Dermatol. 2018, 27, 859–866. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; Visscher, M.; Laruffa, A.; Wickett, R. Natural moisturizing factors (NMF) in the stratum corneum (SC). II. regeneration of NMF over time after soaking. J. Cosmet. Sci. 2010, 61, 23–29. [Google Scholar]
- Debacq-Chainiaux, F.; Leduc, C.; Verbeke, A.; Toussaint, O. UV, stress and aging. Dermatoendocrinology 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röck, K.; Fischer, J.W. Rolle der extrazellulären Matrix bei der extrinsischen HautalterungRole of the extracellular matrix in extrinsic skin aging. Der. Hautarzt. 2011. [Google Scholar] [CrossRef]
- Caon, I.; Parnigoni, A.; Viola, M.; Karousou, E.; Passi, A.; Vigetti, D. Cell energy metabolism and hyaluronan synthesis. J. Histochem. Cytochem. 2020. [Google Scholar] [CrossRef] [PubMed]
- Verkman, A.S.; Anderson, M.O.; Papadopoulos, M.C. Aquaporins: Important but elusive drug targets. Nat. Rev. Drug Discov. 2014, 13, 259–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckert, R.L.; Crish, J.F.; Efimova, T.; Dashti, S.R.; Deucher, A.; Bone, F.; Adhikary, G.; Huang, G.; Gopalakrishnan, R.; Balasubramanian, S. Regulation of involucrin gene expression. J. Investig. Dermatol. 2004, 123, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Purnamawati, S.; Indrastuti, N.; Danarti, R.; Saefudin, T. The role of moisturizers in addressing various kinds of dermatitis: A review. Clin. Med. Res. 2017, 15, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Thibane, V.S.; Ndhlala, A.R.; Abdelgadir, H.A.; Finnie, J.F.; Van Staden, J. The cosmetic potential of plants from the eastern cape province traditionally used for skincare and beauty. S. Afr. J. Bot. 2019, 122, 475–483. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, A.K.; Gupta, A.; Bishayee, A.; Pandey, A.K. Therapeutic potential of Aloe vera—A miracle gift of nature. Phytomedicine 2019, 60, 152996. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, M.; González-Burgos, E.; Iglesias, I.; Gómez-Serranillos, M.P. Pharmacological Update Properties of Aloe Vera and its Major Active Constituents. Molecules 2020, 25, 1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyhanian, S.; Stahl-Biskup, E. Flavonoid and polyphenol content of Aloe vera (Aloe barbadensis Mill.) flowers and their in vitro antioxidative capacity. Planta Med. 2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, T.L.; Van Der Westhuizen, F.H.; Botes, L. Aloe ferox leaf gel phytochemical content, antioxidant capacity, and possible health benefits. J. Agric. Food Chem. 2007, 55, 6891–6896. [Google Scholar] [CrossRef]
- Añibarro-Ortega, M.; Pinela, J.; Barros, L.; Ćirić, A.; Silva, S.P.; Coelho, E.; Mocan, A.; Calhelha, R.C.; Soković, M.; Coimbra, M.A.; et al. Compositional features and bioactive properties of aloe vera leaf (Fillet, mucilage, and rind) and flower. Antioxidants 2019, 8, 444. [Google Scholar] [CrossRef] [Green Version]
- López, A.; De Tangil, M.S.; Vega-Orellana, O.; Ramírez, A.S.; Rico, M. Phenolic constituents, antioxidant and preliminary antimycoplasmic activities of leaf skin and flowers of Aloe vera (L.) Burm. f. (syn. A. barbadensis Mill.) from the Canary Islands (Spain). Molecules 2013, 18, 4942–4954. [Google Scholar] [CrossRef] [Green Version]
- Efimova, T.; Eckert, R.L. Regulation of human involucrin promoter activity by novel protein kinase C isoforms. J. Biol. Chem. 2000. [Google Scholar] [CrossRef] [Green Version]
- Tan, Q.; Yang, H.; Liu, E.; Wang, H. P38/ERK MAPK signaling pathways are involved in the regulation of filaggrin and involucrin by IL-17. Mol. Med. Rep. 2017. [Google Scholar] [CrossRef] [Green Version]
- Efimova, T.; Broome, A.M.; Eckert, R.L. A regulatory role for p38δ MAPK in keratinocyte differentiation: Evidence for p38δ-ERK1/2 complex formation. J. Biol. Chem. 2003. [Google Scholar] [CrossRef] [Green Version]
- Tsatsou, F.; Trakatelli, M.; Patsatsi, A.; Kalokasidis, K.; Sotiriadis, D. Extrinsic aging: UV-mediated skin carcinogenesis. Dermatoendocrinol. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratz-Lyko, A.; Arct, J.; Pytkowska, K. Moisturizing and antiinflammatory properties of cosmetic formulations containing Centella asiatica extract. Indian J. Pharm. Sci. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Guo, M.; Yu, H.; Liu, D.; Lu, Z.; Lu, Y. Propionibacterium acnes related anti-inflammation and skin hydration activities of madecassoside, a pentacyclic triterpene saponin from Centella asiatica. Biosci. Biotechnol. Biochem. 2019, 83, 561–568. [Google Scholar] [CrossRef]
- Lee, W.J.; Park, K.H.; Cha, H.W.; Sohn, M.Y.; Park, K.D.; Lee, S.J.; Kim, D.W. The expression of involucrin, loricrin, filaggrin in cultured sebocytes. Ann. Dermatol. 2014, 26, 134–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efimova, T.; LaCelle, P.; Welter, J.F.; Eckert, R.L. Regulation of human involucrin promoter activity by a protein kinase C, Ras, MEKK1, MEK3, p38/RK, AP1 signal transduction pathway. J. Biol. Chem. 1998. [CrossRef] [PubMed] [Green Version]
- Denning, M.F.; Dlugosz, A.A.; Williams, E.K.; Szallasi, Z.; Blumberg, P.M.; Yuspa, S.H. Specific protein kinase C isozymes mediate the induction of keratinocyte differentiation markers by calcium. Cell Growth Differ. 1995, 6, 149–158. [Google Scholar]
- Adhikary, G.; Chew, Y.C.; Reece, E.A.; Eckert, R.L. PKC-δ and-η, MEKK-1, MEK-6, MEK-3, and p38-δ Are essential mediators of the response of normal human epidermal keratinocytes to differentiating agents. J. Investig. Dermatol. 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Zhang, M.; Luo, H.; Li, H. Isoorientin alleviates UVB-induced skin injury by regulating mitochondrial ROS and cellular autophagy. Biochem. Biophys. Res. Commun. 2019, 514, 1133–1139. [Google Scholar] [CrossRef]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Zhou, Y.; Li, L.; Shen, X.; Chen, G.; Wang, X.; Liang, X.; Tan, M.; Huang, Z. Computational approaches in preclinical studies on drug discovery and development. Front. Chem. 2020, 8, 726. [Google Scholar] [CrossRef]
- Odontuya, G.; Hoult, J.R.S.; Houghton, P.J. Structure-activity relationship for antiinflammatory effect of luteolin and its derived glycosides. Phyther. Res. 2005. [Google Scholar] [CrossRef]
- Bylka, W.; Znajdek-Awizeń, P.; Studzińska-Sroka, E.; Brzezińska, M. Centella asiatica in cosmetology. Postep. Dermatol. I Alergol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Milani, M.; Sparavigna, A. The 24-hour skin hydration and barrier function effects of a hyaluronic 1%, glycerin 5%, and Centella asiatica stem cells extract moisturizing fluid: An intra-subject, randomized, assessor-blinded study. Clin. Cosmet. Investig. Dermatol. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, K.; Choi, H.S.; Hong, Y.H.; Jung, E.Y.; Suh, H.J. Cactus cladodes (Opuntia humifusa) extract minimizes the effects of UV irradiation on keratinocytes and hairless mice. Pharm. Biol. 2017, 55, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Wahedi, H.M.; Jeong, M.; Chae, J.K.; Do, S.G.; Yoon, H.; Kim, S.Y. Aloesin from Aloe vera accelerates skin wound healing by modulating MAPK/Rho and Smad signaling pathways in vitro and in vivo. Phytomedicine 2017, 28, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Radaszkiewicz, K.A.; Beckerová, D.; Woloszczuková, L.; Radaszkiewicz, T.W.; Lesáková, P.; Blanářová, O.V.; Kubala, L.; Humpolíček, P.; Pachernik, J. 12-O-Tetradecanoylphorbol-13-acetate increases cardiomyogenesis through PKC/ERK signaling. Sci. Rep. 2020. [Google Scholar] [CrossRef]
- Chelyn, J.L.; Omar, H.; Akmal, S.; Yousof, M.; Ranggasamy, R.; Wasiman, I.; Ismail, Z. Analysis of flavone c-glycosides in the leaves of clinacanthus nutans (burm. f.) lindau by HPTLC and HPLC-UV/DAD. Sci. World J. 2014. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.A.M.; Yariwake, J.H.; McCullagh, M. Distinction of the C-glycosylfavone isomer pairs orientin/isoorientin and vitexin/isovitexin using HPLC-MS exact mass measurement and in-source CID. Phytochem. Anal. 2005. [Google Scholar] [CrossRef]
- Haque, A.K.M.M.; Leong, K.H.; Lo, Y.L.; Awang, K.; Nagoor, N.H. In vitro inhibitory mechanisms and molecular docking of 1′-S-1′-acetoxychavicol acetate on human cytochrome P450 enzymes. Phytomedicine 2017, 31, 1–9. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | IO | V | IV |
|---|---|---|---|
| Retention time | 18.56 | 23.64 | 25.004 |
| Contents in AFWE | 518.7 ± 0.06 | 153.2 ± 0.9 | 264.8 ± 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razia, S.; Park, H.; Shin, E.; Shim, K.-S.; Cho, E.; Kim, S.-Y. Effects of Aloe vera Flower Extract and Its Active Constituent Isoorientin on Skin Moisturization via Regulating Involucrin Expression: In Vitro and Molecular Docking Studies. Molecules 2021, 26, 2626. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092626
Razia S, Park H, Shin E, Shim K-S, Cho E, Kim S-Y. Effects of Aloe vera Flower Extract and Its Active Constituent Isoorientin on Skin Moisturization via Regulating Involucrin Expression: In Vitro and Molecular Docking Studies. Molecules. 2021; 26(9):2626. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092626
Chicago/Turabian StyleRazia, Sultana, Hyunsung Park, Eunju Shin, Kyu-Suk Shim, Eunae Cho, and Sun-Yeou Kim. 2021. "Effects of Aloe vera Flower Extract and Its Active Constituent Isoorientin on Skin Moisturization via Regulating Involucrin Expression: In Vitro and Molecular Docking Studies" Molecules 26, no. 9: 2626. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092626