4.1. Assessing the Iron Bioavailability of African Yellow Beans
Three yellow beans were selected from market classes that would be recognized by consumers in Sub-Saharan Africa [
23,
24,
25]. Two non-yellow bean varieties were also included in this study, each representing a white and red kidney bean produced for commercial food manufacturing. Red beans, such as the dark red kidney, are preferred seed types in many regions of Sub-Saharan Africa and are often marketed alongside yellow beans [
23,
24]. The white kidney, Snowdon, was included in the study to represent a bean variety with no seed coat polyphenols. Snowdon and Red Hawk are from the same Andean gene pool as the three Africa yellow beans. Snowdon and Red Hawk also share many of the same genetic and agronomic characteristics [
30,
31]. Previous in vitro and in vivo studies have demonstrated that the iron absorption properties between white and red beans are very different from one another, which creates an ideal frame of reference for comparing the iron bioavailability properties of yellow beans [
25,
42]. Relative to the each of the yellow beans in this study, both the white and red kidney bean varieties have longer cooking times in boiling water. The overall nutritional value, as well as the iron bioavailability of dry beans is greatly impacted by the conditions in which they are grown, stored, and processed to become edible [
25,
46,
47,
48]. To limit these factors, beans were grown under the same field conditions, stored in a controlled environment, and their cooking time was standardized for each genotype to avoid over-or-under processing before diet formulation.
As previously demonstrated, the Caco-2 bioassay coupled with the
Gallus gallus in vivo feeding model is an effective two-step system for evaluating the iron bioavailability of staple foods, such as beans [
6,
38,
40,
41,
42,
49]. The Caco-2 bioassay is an initial assessment used to compare varieties and to identify factors in food crops that could potentially impact the absorption of iron in vivo [
38,
39]. For this study, the bioassay shows that iron uptake (via ferritin formation) in Caco-2 cells was negatively impacted when diets were formulated with the darker colored yellow and red beans (
Table 6). The results of the Caco-2 bioassay also matched the same patterns of liver iron and liver ferritin concentrations in each group of animals after the feeding trial (
Table 7). Additionally, the bioassay revealed that diets prepared from the Manteca yellow bean produced a higher iron uptake in Caco-2 cells when compared to the other yellow, white and red bean based diets. The bioassay, however, did not predict the precise patterns of iron absorption between Uyole 98, PI527538 and the kidney beans over the course of the in vivo feeding trial. This exemplifies the need for the
Gallus gallus model to be coupled with the Caco-2 bioassay because total body hemoglobin iron (Hb-Fe) and hemoglobin maintenance efficiency (HME) are two physiological markers that take into consideration an animal’s growth rate, food consumption and adaptation to the bean based diet during the course of a feeding trial [
6,
40,
42,
49].
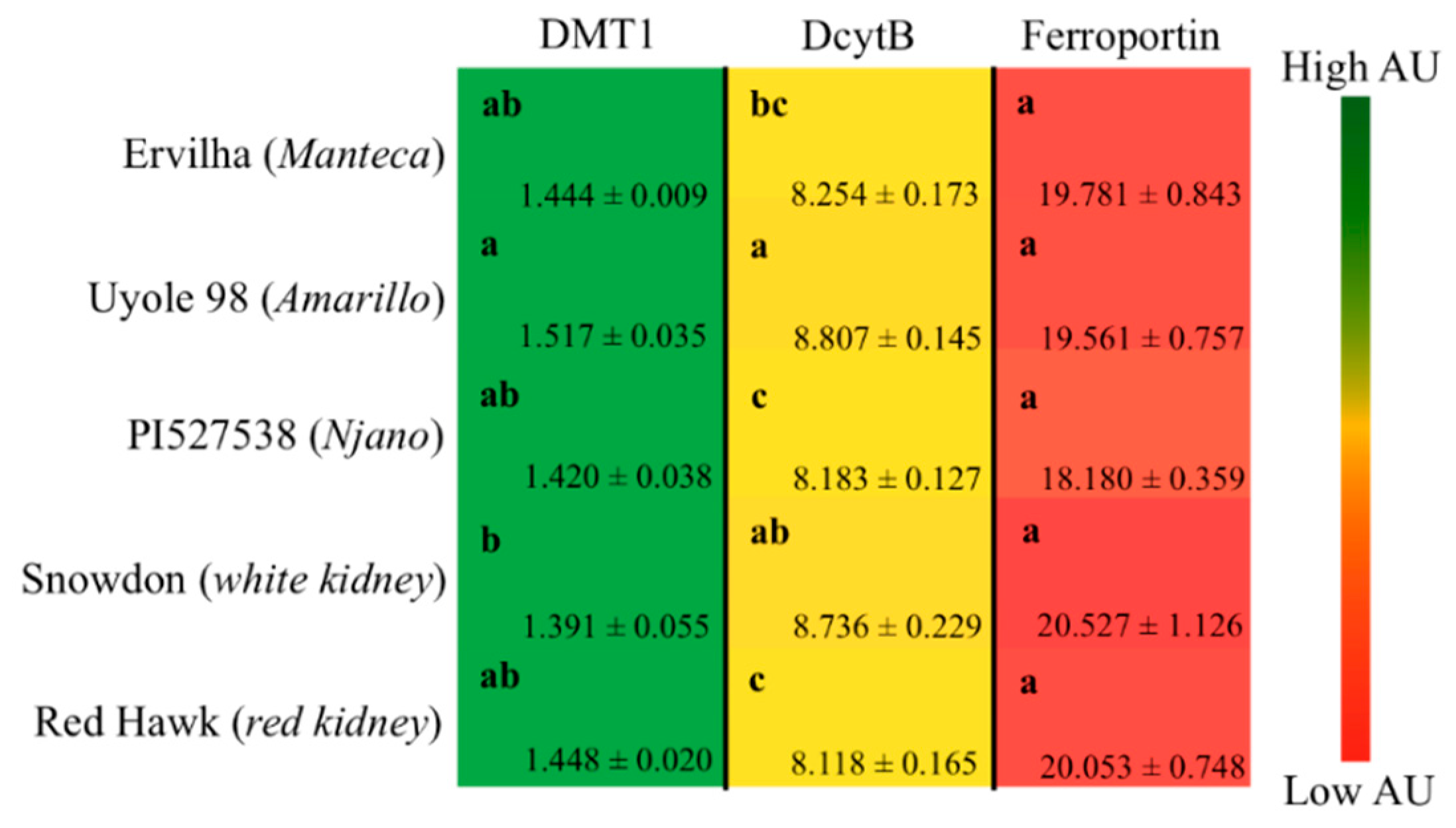
The
Gallus gallus model can also be used to gain insight into the mechanism(s) of iron bioavailability by measuring the hematological, molecular (gene expression) and microbial changes in response to carefully formulated test diets [
14,
38,
40,
41,
42,
50]. Similar to humans, the gene expression of iron absorption proteins and the microbiota profiles of
Gallus gallus are impacted by dietary iron intake [
50,
51]. The
Gallus gallus shares >85% homology with the human gene sequences of the dietary iron import/export proteins DMT-1, DcytB, and ferroportin [
52]. In addition, the phylum levels of gut microbiota between
Gallus gallus and humans are similar, each being dominated by Bacteroidetes, Firmicutes, Proteobacteria and Actinobacteria [
14,
53,
54]. The
Gallus gallus is the first in vivo model to assess the iron nutrition of yellow beans, and this current study represents the first approach to predict the iron bioavailability of yellow beans in humans [
38].
The
Gallus gallus is a useful animal model for testing cooked beans that are combined into diets as either the main ingredient or formulated into a complex meal plan [
40,
41,
42]. For this study, cooked yellow and kidney beans were the main ingredient of bean based diets, contributing to >60% of the total dietary iron in each diet. Bean based diets were formulated with the complementary foods of potato, rice, and cabbage in combinations similar to previous in vivo experiments that have successfully compared the iron bioavailability of different bean varieties in vivo [
14,
41]. Although the combination of diet ingredients used in this study would be quite common in Sub-Saharan Africa, future studies can be designed to evaluate the iron bioavailability of yellows beans within a multiple ingredient meal plan (the food basket approach), which is more precisely tailored to specific regions where yellow beans are already familiar to consumers [
14,
41,
55].
4.2. The Iron Bioavailability of African Yellow Beans
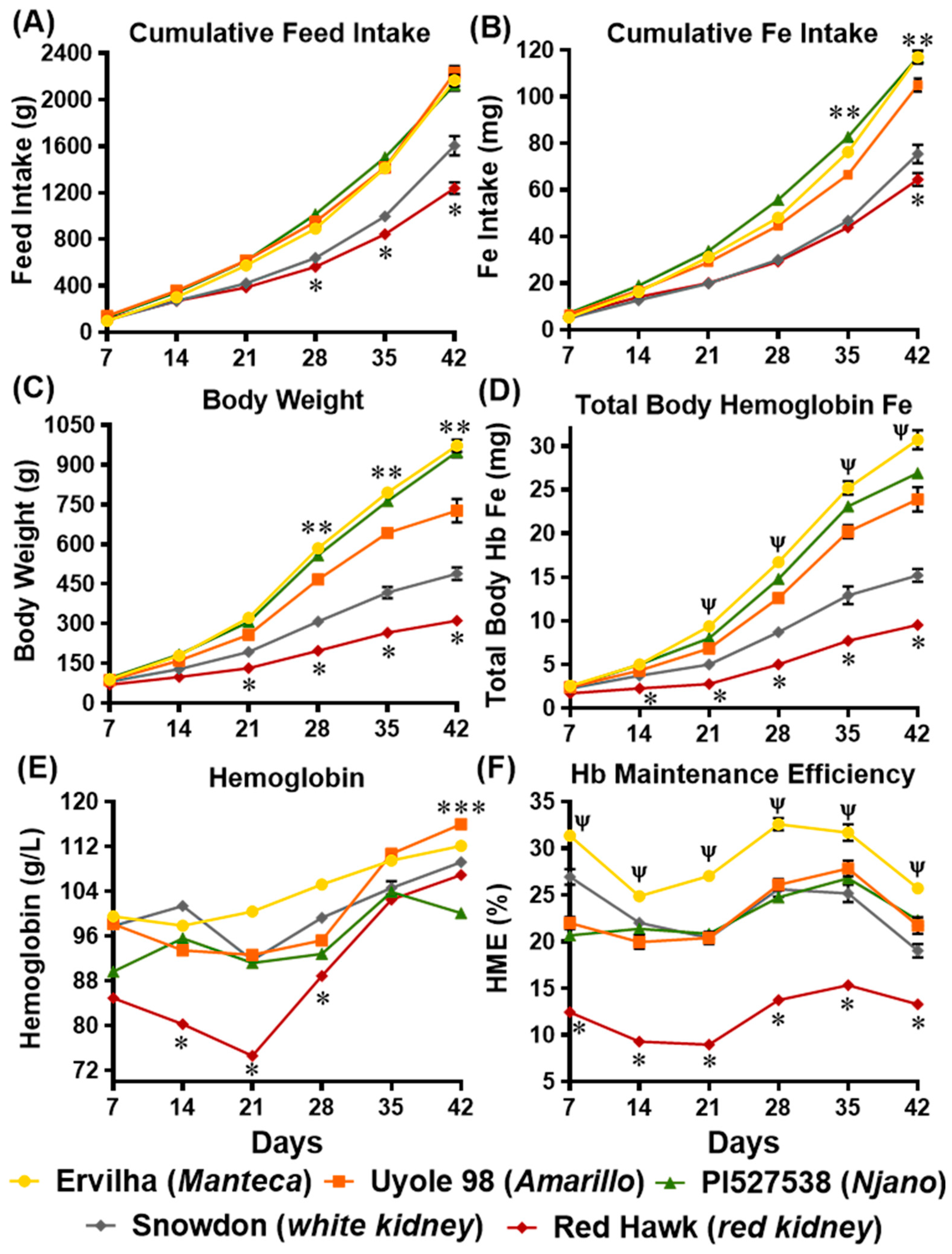
Of the three African yellow beans tested in this study, the fastest cooking Manteca yellow bean (Ervilha) delivered the most absorbable iron for growth and hemoglobin production during the six week feeding trial. The Manteca is a pale-yellow bean native to Chile, characterized by its gray-black hilum ring. Higher prices at the market and anecdotal claims of ‘low-flatulence’ inspired scientists to examine the Manteca seed type’s biochemistry [
20,
56,
57]. They found the Manteca has a different fiber profile with more digestible starch and protein when compared to other black and red beans [
57,
58,
59]. More importantly, they discovered the Manteca carries a recessive allele that closes off the production of procyanidin (condensed tannin) synthesis in the polyphenol pathway [
20,
56,
60].
In addition to producing a darker seed coat color, condensed tannin concentrations influence the cooking time, as well as the digestibility of dry beans [
59,
61,
62]. Although diets formulated with the Ervilha and PI527538 had similar iron concentrations and phytate-iron molar ratios (
Table 2), the results of this experiment indicate that the slowest cooking Njano yellow bean PI527538 did not deliver as much iron as the fastest cooking Manteca yellow bean Ervilha. These results are in agreement with the current and past Caco-2 cell culture experiments, which show how Ervilha and PI528537 have distinct iron bioavailability that is independent of their iron concentrations after cooking [
25]. The same polyphenols in black and red beans that were previously shown to inhibit the absorption of iron in vitro (quercetin 3-glucoside, procyanidin B1, procyanidin B2) were also detected in PI527538 after cooking [
5,
6,
7,
8]. These results provide evidence that the fast cooking Manteca is biochemically unique to other yellow beans, and demonstrates how altering the downstream production of polyphenols to prevent the synthesis of condensed tannins could be a useful mechanism to improve the iron bioavailability of dry beans [
5,
6,
7,
8,
25].
Unlike previous investigations that have only compared the iron bioavailability of non-biofortified and biofortified beans in the same market class [
6,
14,
40,
41,
55], the results of this study demonstrate that the iron bioavailability of yellow beans after cooking is not dependent on seed iron concentrations alone. For example, total dietary iron concentrations (
Table 2) and cumulative iron intakes (
Figure 2B) were lower for animals receiving the Amarillo yellow bean Uyole 98 diet, yet they were able to maintain their hemoglobin, Hb-Fe, and HME values, as well as their liver iron concentrations just as effectively as the animals receiving the slower cooking PI527538 yellow bean diet. The up-regulation of the iron import genes DMT-1 and DcytB in the animals receiving the Uyole 98 diet indicates there was a physiological adaptation to the lower dietary iron concentrations [
6,
40,
41]. Despite the changes in the gene expression of iron import proteins, animals receiving the Uyole 98 diet were still unable to maintain their growth and hemoglobin production as efficiently as the animals receiving the fastest cooking Ervilha bean based diet. Interestingly, Uyole 98 had the highest concentrations of kaempferol 3-glucoside, but small amounts of the precursors to condensed tannins (catechin, epicatechin, procyanidin B1) were also detected in Uyole 98 bean based diets. There is evidence to suggest that these compounds may be more potent inhibitors of dietary iron than kaempferol 3-glucoside is a promotor of iron in vitro [
5,
6,
7].
The contrast between Ervilha, Uyole 98, and PI527538 serves as a good example that not all yellow beans will have the same iron bioavailability. Demonstrating that there is phenotypic diversity in iron bioavailability between the different market classes of yellow beans, however, is important for dry bean breeders to identify targets traits that can lead to genetic improvement, while, at the same time, complementing the biofortification efforts that are currently underway to increase raw bean seed iron concentrations in yellow beans [
4,
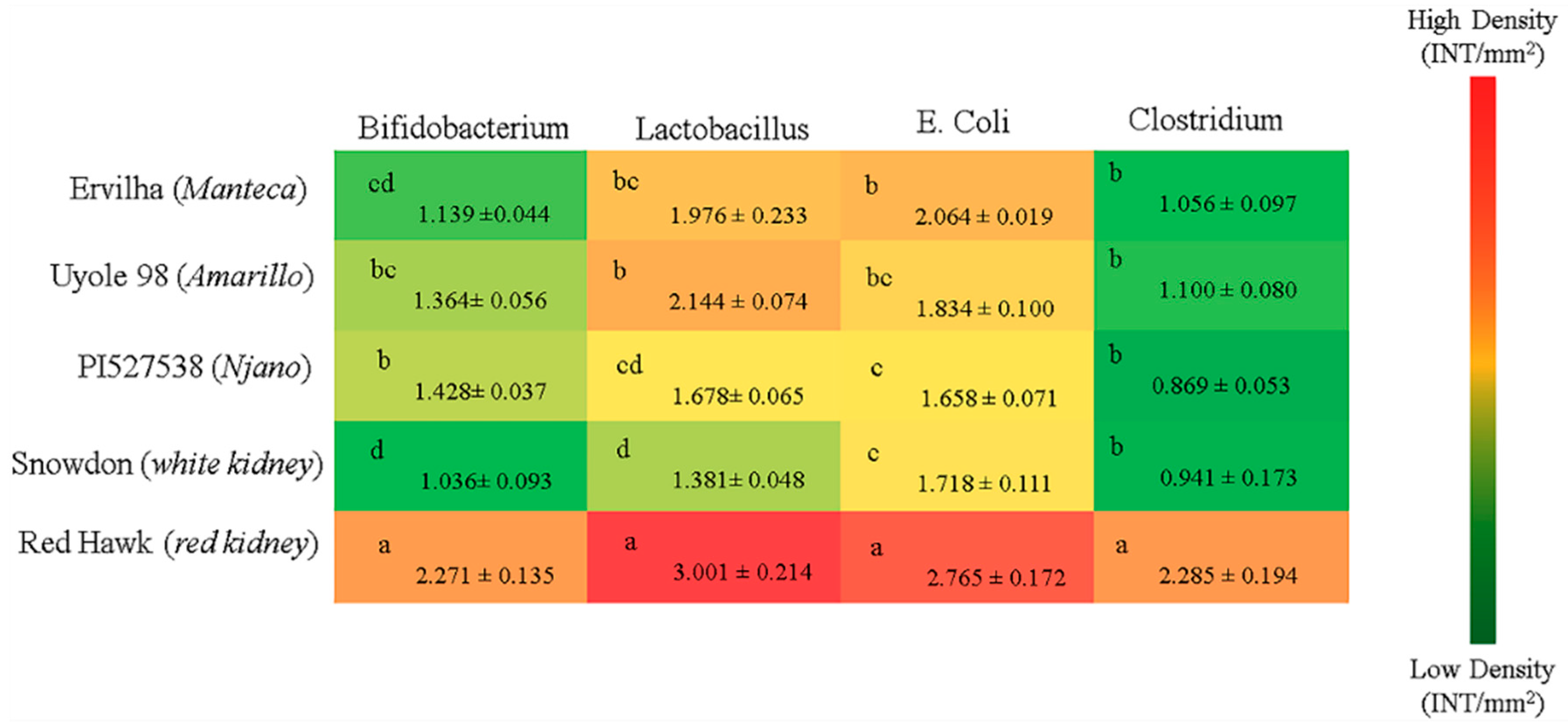
63]. This study is the first to show that differences in polyphenols and dietary fiber concentrations between Ervilha, Uyole 98 and PI527538 can also play a role in determining the microbiota populations of the lower intestine (
Figure 4). For example, the population densities of Bifidobacterium and E. Coli in the cecum were significantly different between animals fed the Ervilha and PI527538 yellow bean diets. This is an important finding because the polyphenols and fiber (prebiotics) found in dry beans are implicated in maintaining the health of the digestive system by influencing bacterial (probiotic) populations in the lower intestine [
9,
10,
11,
12,
13,
14]. While the measurements of microbiota at the species level is too broad of an interpretation to make any specific health recommendations about yellow beans, a more detailed study is currently being conducted to examine the microbial diversity and compositional changes of gut microbiota at the Phylum and Genus levels in animals fed the yellow and kidney bean based diets.
4.3. Iron Bioavailability of the White and Red Kidney Beans
Animals receiving the white and red kidney bean diets had low feed intakes and slower growth rates, which prevented these animals from accumulating the Hb-Fe values that were achieved by the animals receiving yellow bean diets (
Figure 2). With the exception of Uyole 98, the white and red kidney beans tested in this study had significantly more dietary fiber and less protein than the African yellow beans (
Table 3). There is evidence to support that increases in dietary fiber from habitual pulse consumption increase perceived satiety and reduce food intake, while intervention studies show that pulse consumption leads to reductions in body weight with or without energy restriction [
64,
65]. The differences in soluble and insoluble fiber concentrations between the two seed types are also indicative of differences in seed coat thickness and cotyledon cell wall composition, which are additional factors that need to be considered when comparing the digestibility and iron bioavailability of yellow and kidney beans [
66].
The large differences in Hb-Fe between the animals receiving the white kidney Snowdon diet and the yellow Uyole 98 diet, which had the same iron concentrations, phytate-iron molar ratios, and fiber concentrations, suggest that kaempferol 3-glucoside may facilitate the absorption of dietary iron [
5,
6,
7,
8]. The overall digestibility and iron nutrition of African yellow beans with kaempferol 3-glucoside as their most abundant polyphenol appears to be far superior to that of white and red kidney beans-with polyphenol profiles that contain little to no kaempferol compounds. The large differences in growth rates and feed intakes between the two groups receiving the yellow and kidney bean diets, however, makes it difficult to pin-point kaempferol 3-glucoside as the only contributor of improved iron bioavailability among the three African yellow beans. Future investigations are needed to understand kaempferol 3-glucoside’s full potential as a dietary promoter of iron bioavailability in dry beans, as well as in other world food crops that also contain kaempferol compounds, such as the yellow potato or yellow cassava [
15].
Comparing the two kidney beans to each other revealed the iron bioavailability of Red Hawk, with a diverse array of polyphenols detected in its dark red seed coat, was significantly lower than Snowdon, which had little to no polyphenols in its white seed coat (
Table 4). Animals receiving the Red Hawk diet could not absorb enough iron from the diet to maintain hemoglobin production during the first 2–3 weeks of the feeding trial (
Figure 2E). An interesting adaptation in animals receiving the Red Hawk diet, however, was evident between weeks 3 and 4 of the experiment as hemoglobin production began to improve. Of the five beans tested in this study, the highest concentrations of insoluble and soluble fiber were measured in Red Hawk, which is an important observation because a more diverse collection of polyphenols, and more dietary fiber (prebiotics) are important factors that promote the diversity of microbiota populations in the lower intestine [
9,
10,
11,
12,
13,
14]. All four bacterial populations in the cecum at the end of the feeding trial were significantly elevated in the animals receiving the Red Hawk diet, when compared to the animals receiving the white and yellow bean diets (
Figure 4). The probiotic microbial activity and fermentation of fiber in the intestinal lumen produce small chain fatty acids (SCFA) that include acetate, propionate, and butyrate [
67,
68,
69]. These molecules are important to the metabolism and health of the digestive system because they promote intestinal cell proliferation and serve as an energy source for colonic epithelial cells [
67,
68,
69]. The recovery of hemoglobin production in the animals receiving the Red Hawk diet by the end of the feeding trial, in conjunction with increased levels of bacteria populations in the cecum, indicates that the microbial composition of the lower intestine can be a contributing factor in the iron status of the host. These results are consistent with previous animal studies that show how dietary fiber can remodel the microbiota in the lower intestine; which in turn, can improve the morphometric parameters of the upper intestine by stimulating the proliferation of enterocytes and the height of duodenal villi [
12,
13,
14,
67,
69]. These findings continue to support the idea that breeding for specific traits that promote the health and diversity of bacterial populations in the digestive system is an additional strategy that biofortification programs can use to improve the efficacy of biofortified food crops [
14,
70,
71].

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}