The Effect of N-Acetylcysteine on Respiratory Enzymes, ADP/ATP Ratio, Glutathione Metabolism, and Nitrosative Stress in the Salivary Gland Mitochondria of Insulin Resistant Rats

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Mitochondria Isolation
2.3. Mitochondrial Assays
2.4. Activity of Mitochondrial Complexes
2.5. ADP/ATP Ratio and Hydrogen Peroxide Production
2.6. Activity of Citrate Synthase
2.7. Reduced and Oxidized Glutathione and Redox Status
2.8. Pro-Oxidant Enzymes
2.9. Inflammation and Apoptosis
2.10. Nitrosative Stress
2.11. Statistical Analysis
3. Results
3.1. General Characteristics
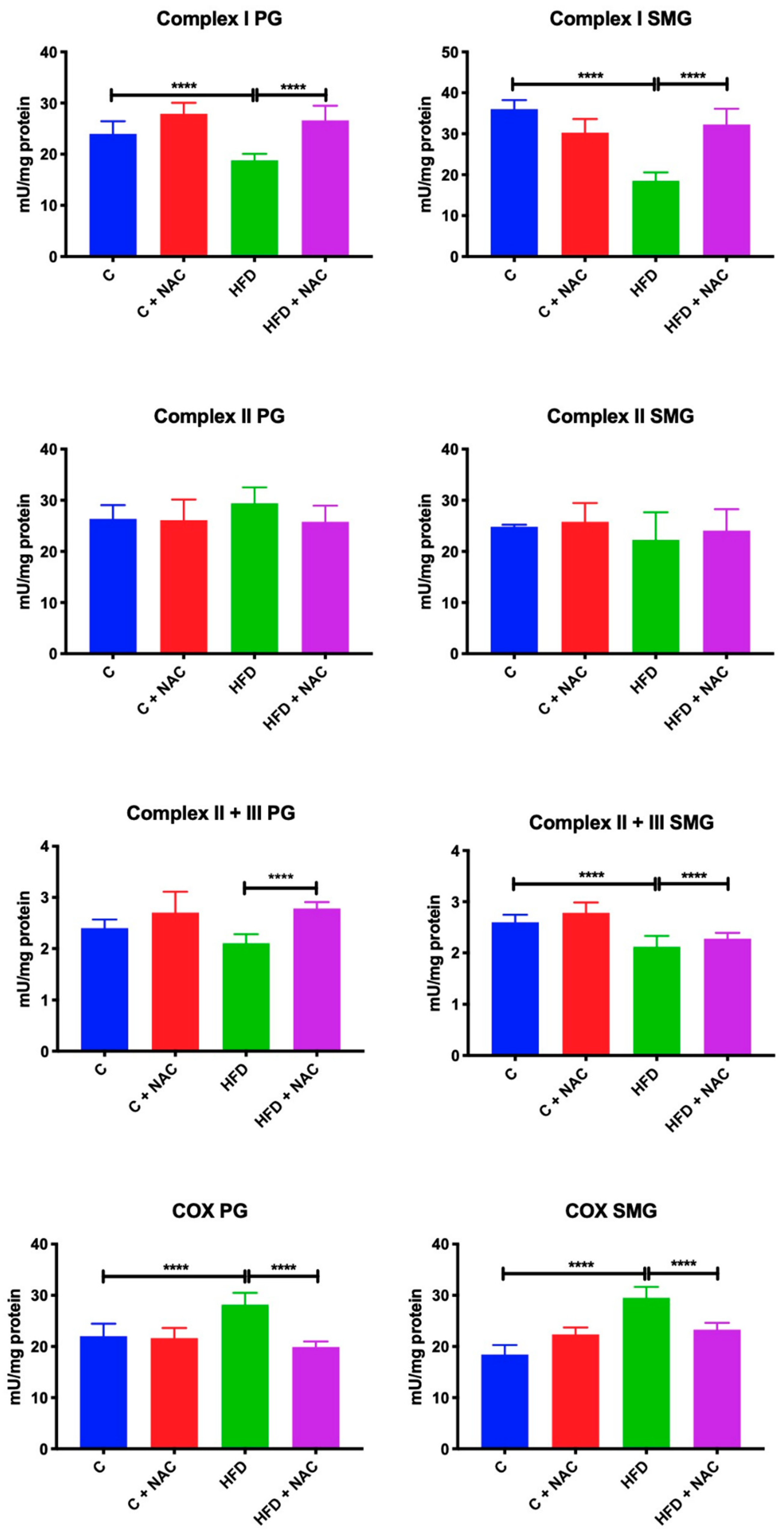
3.2. Activity of Mitochondrial Complexes
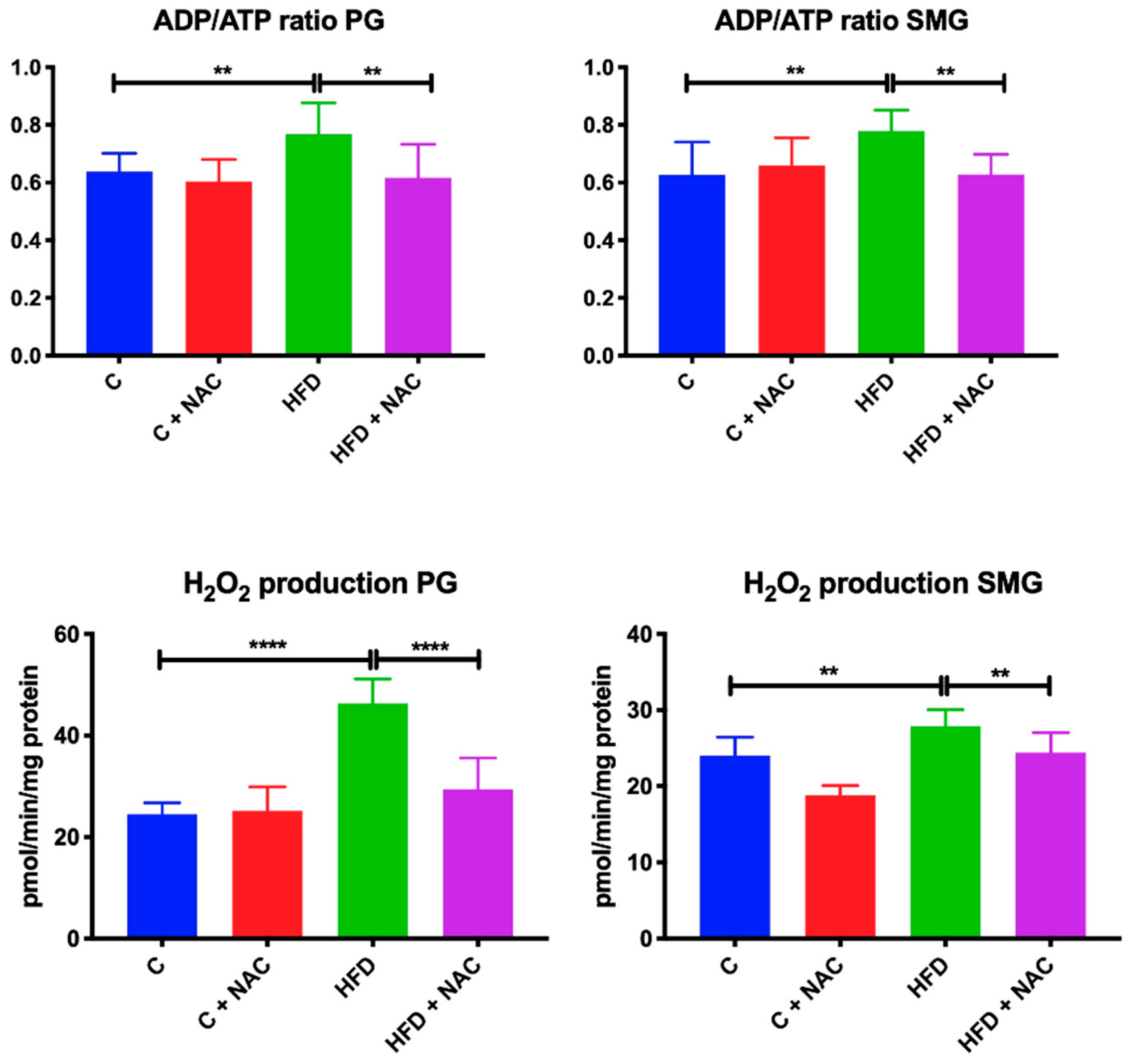
3.3. ADP/ATP Ratio and H2O2 Level
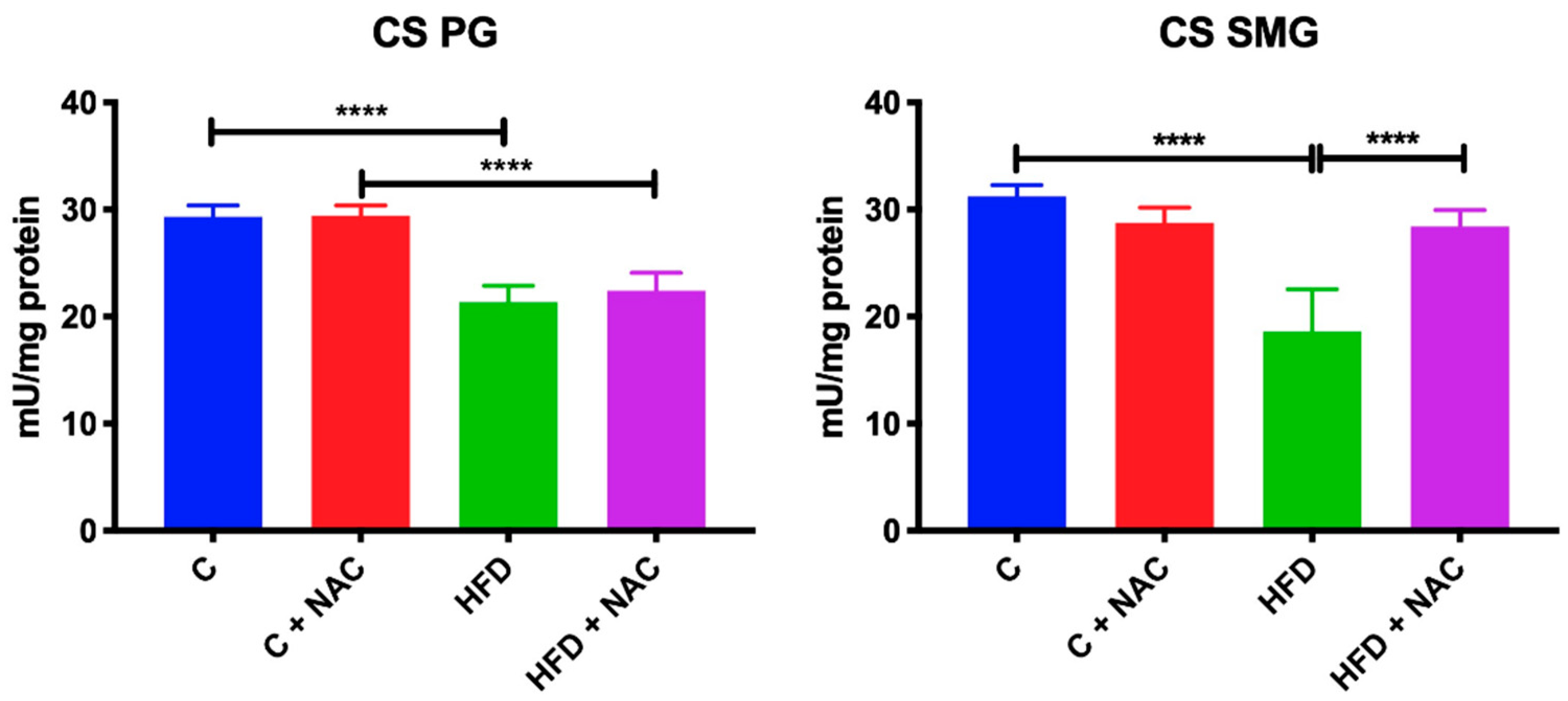
3.4. Activity of Mitochondrial CS
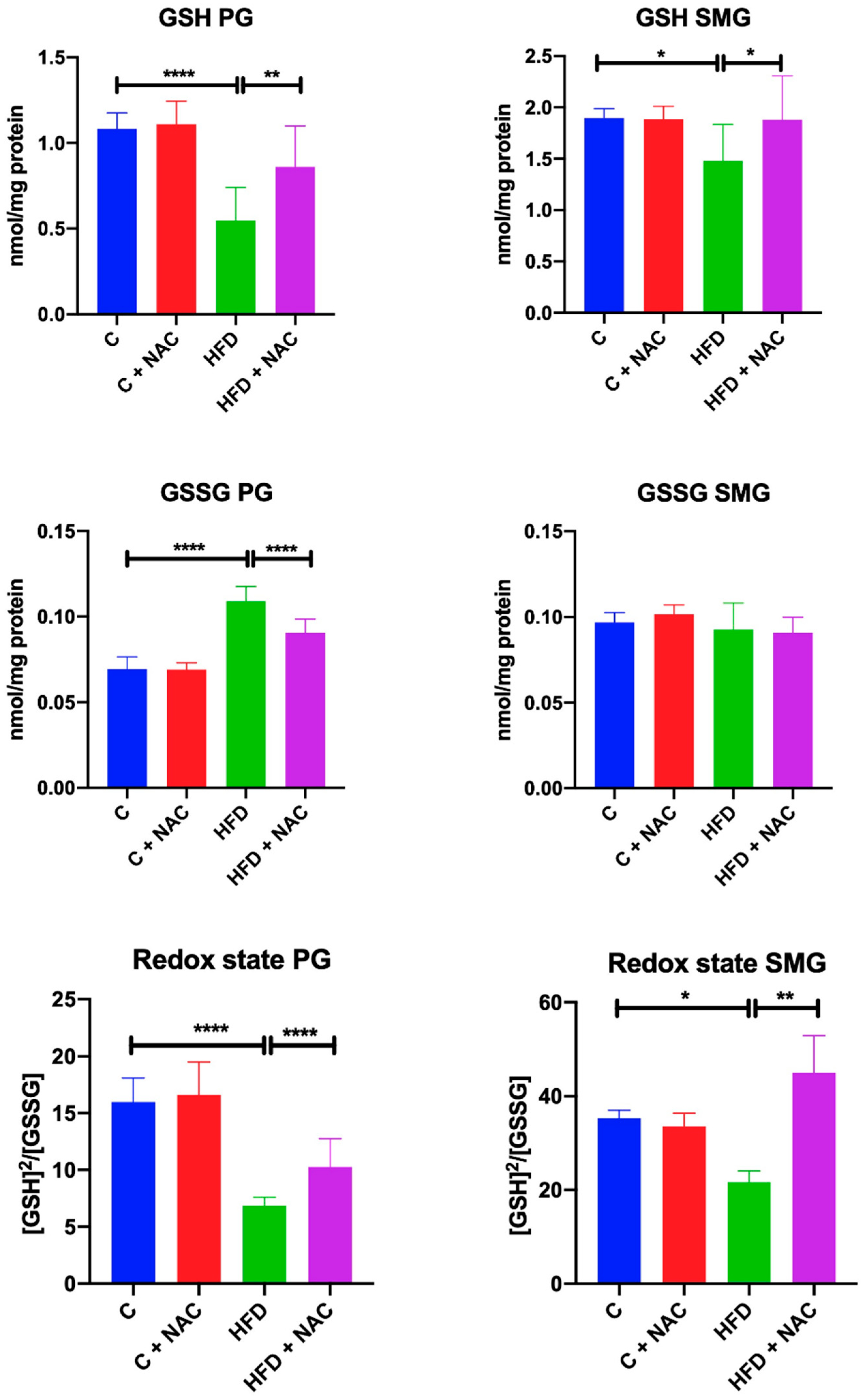
3.5. GSH, GSSG and Redox Status
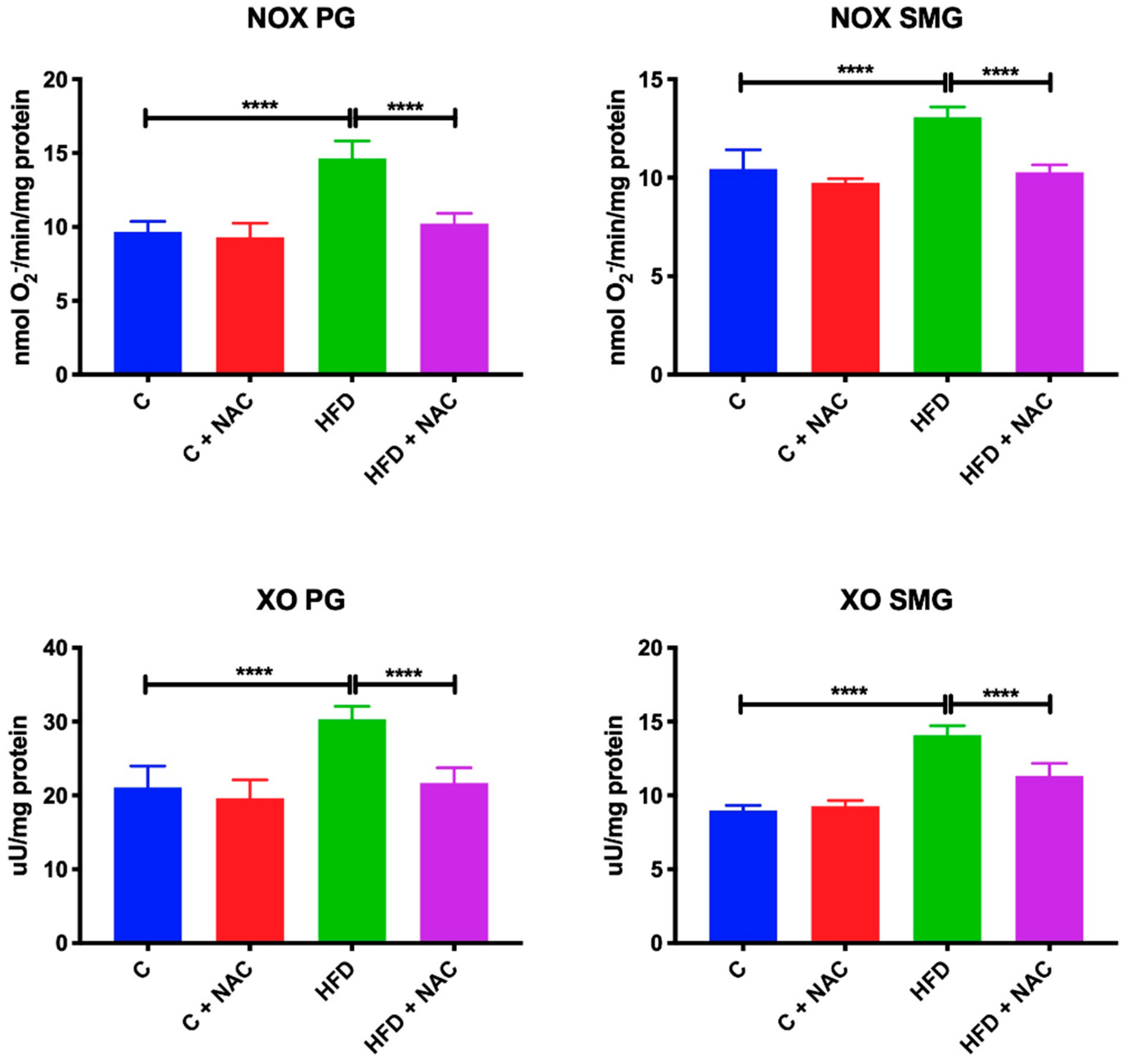
3.6. Activity of NOX and XO
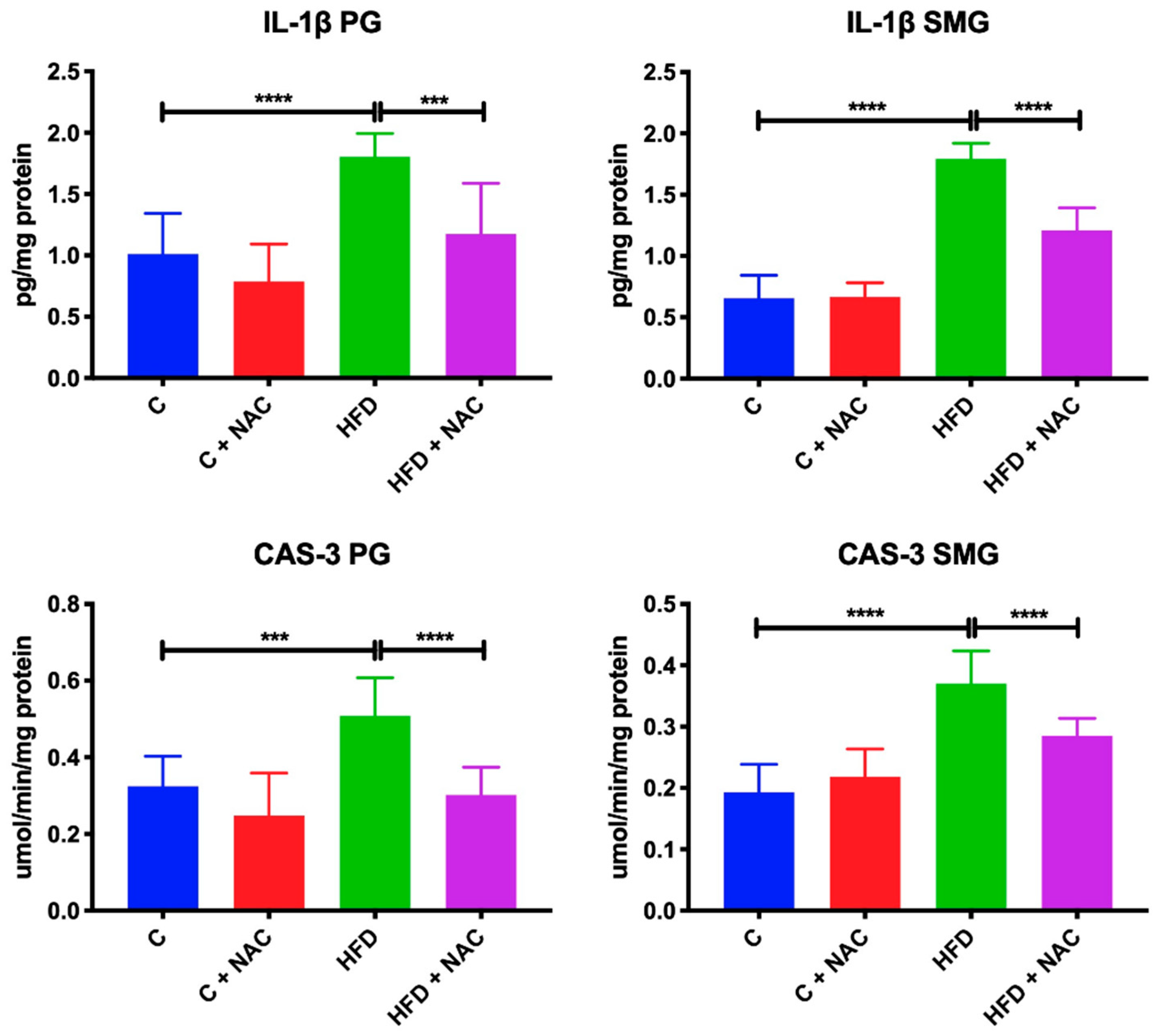
3.7. Inflammation and Apoptosis
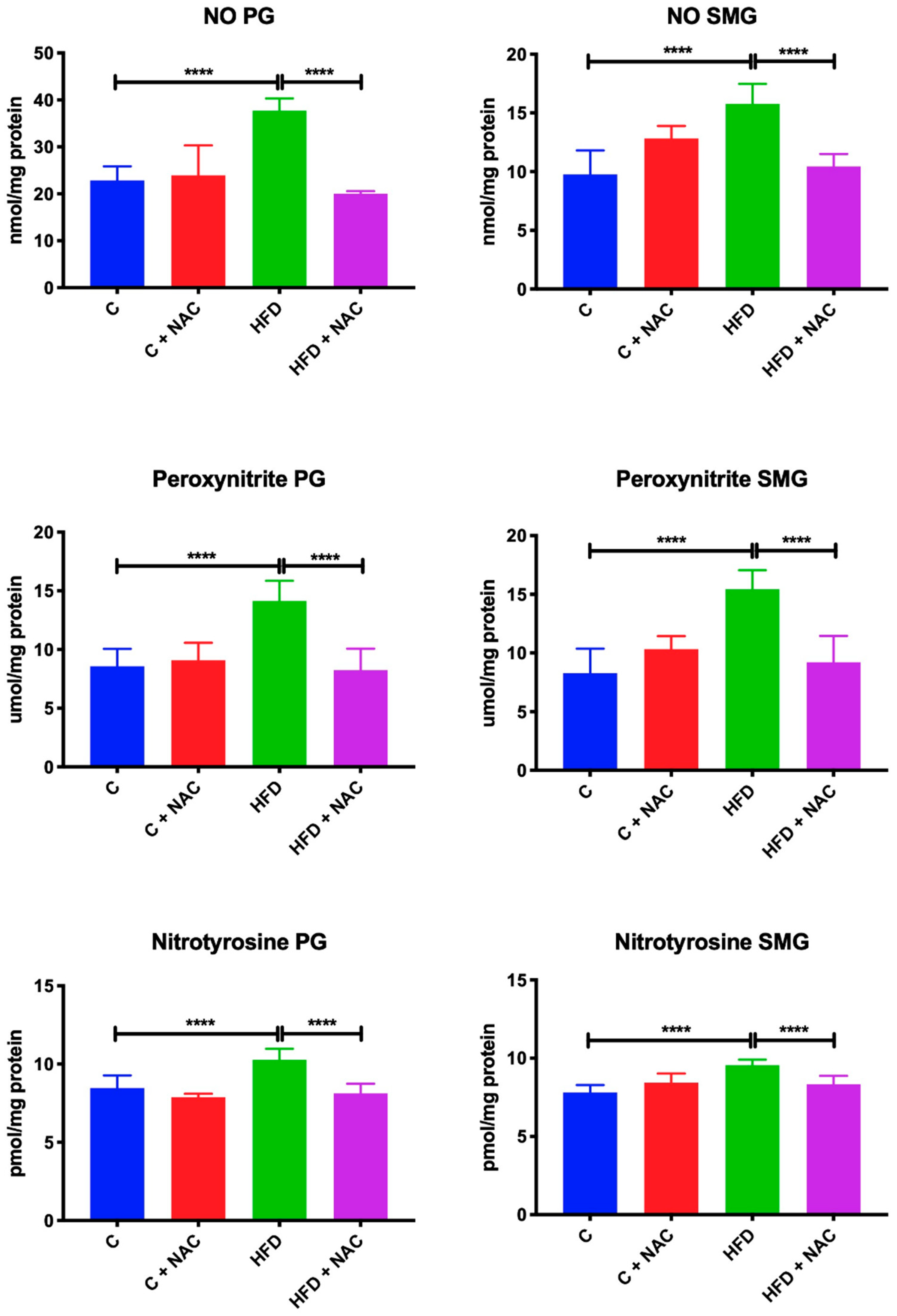
3.8. Nitrosative Stress
3.9. Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lushchak, V.I. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino. Acids. 2012, 2012, 736837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ergin, B.; Guerci, P.; Zafrani, L.; Nocken, F.; Kandil, A.; Gurel-Gurevin, E.; Demirci-Tansel, C.; Ince, C. Effects of N-acetylcysteine (NAC) supplementation in resuscitation fluids on renal microcirculatory oxygenation, inflammation, and function in a rat model of endotoxemia. Int. Care Med. Exp. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Paterson, R.L.; Galley, H.F.; Webster, N.R. The effect of N-acetylcysteine on nuclear factor-kappa B activation, interleukin-6, interleukin-8, and intercellular adhesion molecule-1 expression in patients with sepsis. Crit. Care Med. 2003, 31, 2574–2578. [Google Scholar] [CrossRef] [PubMed]
- Villagrasa, V.; Cortijo, J.; Marti-Cabrera, M.; Ortiz, J.L.; Berto, L.; Esteras, A.; Bruseghini, L.; Morcillo, E.J. Inhibitory effects of N-acetylcysteine on superoxide anion generation in human polymorphonuclear leukocytes. J. Pharm. Pharmacol. 1997, 49, 525–529. [Google Scholar] [CrossRef]
- Shen, H.M.; Yang, C.F.; Ding, W.X.; Liu, J.; Ong, C.N. Superoxide radical-initiated apoptotic signalling pathway in selenite-treated HepG(2) cells: Mitochondria serve as the main target. Free Radic. Biol. Med. 2001, 30, 9–21. [Google Scholar] [CrossRef]
- Cocco, T.; Sgobbo, P.; Clemente, M.; Lopriore, B.; Grattagliano, I.; Di Paola, M.; Villani, G. Tissue-specific changes of mitochondrial functions in aged rats: Effect of a long-term dietary treatment with N-acetylcysteine. Free Radic. Biol. Med. 2005, 38, 796–805. [Google Scholar] [CrossRef]
- Miquel, J.; Ferrandiz, M.L.; De Juan, E.; Sevila, I.; Martinez, M. N-acetylcysteine protects against age-related decline of oxidative phosphorylation in liver mitochondria. Eur. J. Pharmacol. 1995, 292, 333–335. [Google Scholar] [CrossRef]
- Martinez Banaclocha, M.; Martinez, N. N-acetylcysteine elicited increase in cytochrome c oxidase activity in mice synaptic mitochondria. Brain Res. 1999, 842, 249–251. [Google Scholar] [CrossRef]
- Martinez Banaclocha, M. N-acetylcysteine elicited increase in complex I activity in synaptic mitochondria from aged mice: Implications for treatment of Parkinson’s disease. Brain Res. 2000, 859, 173–175. [Google Scholar] [CrossRef]
- Gonzalez, R.; Ferrin, G.; Hidalgo, A.B.; Ranchal, I.; Lopez-Cillero, P.; Santos-Gonzalez, M.; Lopez-Lluch, G.; Briceno, J.; Gomez, M.A.; Poyato, A.; et al. N-acetylcysteine, coenzyme Q10 and superoxide dismutase mimetic prevent mitochondrial cell dysfunction and cell death induced by d-galactosamine in primary culture of human hepatocytes. Chem. Biol. Interact. 2009, 181, 95–106. [Google Scholar] [CrossRef]
- Xiong, Y.; Peterson, P.L.; Lee, C.P. Effect of N-acetylcysteine on mitochondrial function following traumatic brain injury in rats. J. Neurotrauma 1999, 16, 1067–1082. [Google Scholar] [CrossRef]
- Lushchak, V.L. Classification of oxidative stress based on its intensity. Exp. Clin. Sci. 2014, 13, 922–937. [Google Scholar]
- Kołodziej, U.; Maciejczyk, M.; Miąsko, A.; Matczuk, J.; Knaś, M.; Żukowski, P.; Żendzian-Piotrowska, M.; Borys, J.; Zalewska, A. Oxidative modification in the salivary glands of high fat-diet induced insulin resistant rats. Front. Physiol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Zalewska, A.; Knaś, M.; Żendzian-Piotrowska, M.; Waszkiewicz, N.; Szulimowska, J.; Prokopiuk, S.; Waszkiel, D.; Car, H. Antioxidant profile of salivary glands in high fat diet- induced insulin resistance rats. Oral. Dis. 2014, 20, 560–566. [Google Scholar] [CrossRef]
- Żukowski, P.; Maciejczyk, M.; Matczuk, J.; Kurek, K.; Waszkiel, D.; Żendzian-Piotrowska, M.; Zalewska, A. Effect of N-acetylcysteine on antioxidant defense, oxidative modification, and salivary gland function in a rat model of insulin resistance. Oxid. Med. Cell Longev. 2018, 2018. [Google Scholar] [CrossRef]
- Zalewska, A.; Ziembicka, D.; Zendzian-Piotrowska, M.; Maciejczyk, M. The Impact of High-Fat Diet on Mitochondrial Function, Free Radical Production, and Nitrosative Stress in the Salivary Glands of Wistar Rats. Oxid. Med. Cell Longev. 2019, 2019, 2606120. [Google Scholar] [CrossRef] [Green Version]
- Leite, R.S.; Marlow, N.M.; Fernandes, J.K. Oral health and type 2 diabetes. Am. J. Med. Sci. 2013, 345, 271–273. [Google Scholar] [CrossRef] [Green Version]
- Modèer, T.; Blomberg, C.C.; Wondimu, B.; Julihn, A.; Marcus, C. Association between obesity, flow rate of whole saliva, and dental caries in adolescents. Obesity 2010, 18, 2367–2373. [Google Scholar] [CrossRef]
- Su, H.; Velly, A.M.; Salah, M.H.; Benarroch, M.; Trifiro, M.; Schipper, H.M.; Gornitsky, M. Altered redox homeostasis in human diabetes saliva. J. Oral. Pathol. Med. 2012, 41, 235–241. [Google Scholar] [CrossRef]
- Martinez, R.F.; Jaimes-Aveldañez, A.; Hernández-Pèrez, F.; Arenas, R.; Miguel, G.F. Oral Candida spp carriers: Its prevalence in patients with type 2 diabetes mellitus. An Bras Dermatol 2013, 88, 222–225. [Google Scholar] [CrossRef]
- Dursun, E.; Akalin, F.A.; Genc, T.; Cinar, N.; Erel, O.; Yildiz, B.O. Oxidative stress and periodontal disease in obesity. Medicine 2016, 95, e3136. [Google Scholar] [CrossRef]
- Zeigler, C.C.; Persson, G.R.; Wondimu, B.; Marcus, C.; Sobko, T.; Modèer, T. Microbiota in the oral subgingival biofilm is associated with obesity in adolescence. Obesity 2012, 20, 157–164. [Google Scholar] [CrossRef]
- Carda, C.; Mosquera-Lloreda, N.; Salom, L.; Gomez de Ferraris, M.E.; Peydro, A. Structural and functional salivary disorders in type 2 diabetic patients. Med. Oral. Pathol. Oral. Cir. Bucal. 2006, 11, 309–313. [Google Scholar]
- D’Aiuto, F.; Nibali, L.; Parkar, M.; Patel, K.; Suvan, J.; Donos, N. Oxidative stress, systemic inflammation, and severe periodontitis. J. Dent. Res. 2010, 89, 1241–1246. [Google Scholar] [CrossRef] [Green Version]
- Knaś, M.; Maciejczyk, M.; Daniszewska, I.; Klimiuk, A.; Matczuk, J.; Kołodziej, U.; Waszkiel, D.; Ładny, J.R.; Żendzian-Piotrowska, M.; Zalewska, A. Oxidative Damage to the Salivary Glands of Rats with Streptozotocin-Induced Diabetes-Temporal Study: Oxidative Stress and Diabetic Salivary Glands. J. Diabetes Res. 2016. [Google Scholar] [CrossRef] [Green Version]
- Jaccob, A.A. Protective effect of N-acetylcysteine against ethanol-induced gastric ulcer: A pharmacological assessment in mice. J Intercult Ethnopharmacol 2015, 4, 90–95. [Google Scholar] [CrossRef]
- Kononczuk, T.; Lukaszuk, B.; Miklosz, A.; Chabowski, A.; Zendzian-Piotrowska, M.; Kurek, K. Cerulein-Induced Acute Pancreatitis Affects Sphingomyelin Signaling Pathway in Rats. Pancreas 2018, 47, 898–903. [Google Scholar] [CrossRef]
- Maciejczyk, M.; Zebrowska, E.; Zalewska, A.; Chabowski, A. Redox Balance, Antioxidant Defense, and Oxidative Damage in the Hypothalamus and Cerebral Cortex of Rats with High Fat Diet-Induced Insulin Resistance. Oxid. Med. Cell. Longev. 2018, 2018, 6940515. [Google Scholar] [CrossRef] [Green Version]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Zalewska, A.; Maciejczyk, M.; Szulimowska, J.; Imierska, M.; Błachnio-Zabielska, A. High-Fat Diet Affects Ceramide Content, Disturbs Mitochondrial Redox Balance, and Induces Apoptosis in the Submandibular Glands of Mice. Biomolecules 2019, 9, 877. [Google Scholar] [CrossRef] [Green Version]
- Janssen, A.J.M.; Trijbels, F.J.M.; Sengers, R.C.A.; Smeitink, J.A.M.; Van Den Heuvel, L.P.; Wintjes, L.T.M.; Stoltenborg-Hogenkamp, B.J.M.; Rodenburg, R.J.T. Spectrophotometric assay for complex I of the respiratory chain in tissue samples and cultured fibroblasts. Clin. Chem. 2007, 53, 729–734. [Google Scholar] [CrossRef]
- Rustin, P.; Chretien, D.; Bourgeron, T.; Gérard, B.; Rötig, A.; Saudubray, J.M.; Munnich, A. Biochemical and molecular investigations in respiratory chain deficiencies. Clin. Chim. Acta 1994. [Google Scholar] [CrossRef]
- Wharton, D.C.; Tzagoloff, A. Cytochrome oxidase from beef heart mitochondria. Methods Enzymol. 1967. [Google Scholar] [CrossRef]
- Muller, F.L.; Liu, Y.; Van Remmen, H. Complex III releases superoxide to both sides of the inner mitochondrial membrane. J. Biol. Chem. 2004. [Google Scholar] [CrossRef] [Green Version]
- Srere, P.A. Citrate synthase. [EC 4.1.3.7 Citrate oxaloacetate-lyase (CoA-acetylating)]. Methods Enzymol. 1969. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Griendling, K.K.; Minieri, C.A.; Ollerenshaw, J.D.; Alexander, R.W. Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ. Res. 1994, 74, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Prajda, N.; Weber, G. Malignant transformation-linked imbalance: Decreased xanthine oxidase activity in hepatomas. FEBS Lett. 1975, 59, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Meki, A.R.; Esmail, E.-D.; Hussein, A.A.; Hassanein, H.M. Caspase-3 and Heat Shock Protein-70 in Rat Liver Treated with Aflatoxin B1: Effect of Melatonin. Toxicon 2004, 93–100. [Google Scholar] [CrossRef]
- Grisham, M.B.; Johnson, G.G.; Lancaster, J.R. Quantitation of nitrate and nitrite in extracellular fluids. Methods Enzymol. 1996, 268, 237–246. [Google Scholar]
- Beckman, J.S.; Ischiropoulos, H.; Zhu, L.; van der Woerd, M.; Smith, C.; Chen, J.; Harrison, J.; Martin, J.C.; Tsai, M. Kinetics of superoxide dismutase- and iron-catalyzed nitration of phenolics by peroxynitrite. Arch. Biochem. Biophys. 1992, 298, 438–445. [Google Scholar] [CrossRef]
- Kurek, K.; Miklosz, A.; Lukaszuk, B.; Chabowski, A.; Gorski, J.; Zendzian-Piotrowska, M. Inhibition of Ceramide De Novo Synthesis Ameliorates Diet Induced Skeletal Muscles Insulin Resistance. J. Diabetes Res. 2015, 2015, 154762. [Google Scholar] [CrossRef] [Green Version]
- Ebertz, C.E.; Bonfleur, M.L.; Bertasso, I.M.; Mendes, M.C.; Lubaczeuski, C.; Araujo, A.C.; Paes, A.M.; Amorim, E.M.P.; Balbo, S.L. Duodenal jejunal bypass attenuates non-alcoholic fatty liver disease in western diet-obese rats. Acta Cir. Bras. 2014, 29, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Gan, K.X.; Wang, C.; Chen, J.H.; Zhu, C.J.; Song, G.Y. Mitofusin-2 ameliorates high-fat diet- induced insulin resistance in liver of rats. World J. Gastroenterol. 2013, 19, 1572–1581. [Google Scholar] [CrossRef]
- Qu, D.M.; Song, G.Y.; Gao, Y.; Wang, J.; Hu, S.G.; Han, M. Expression pattern of PGC-1 alpha and Mfn2 in insulin resistance state after recovery in rat. Jichu Yixue Yu Linchuang 2008, 28, 133–137. [Google Scholar]
- Ceriello, A.; Testa, R. Antioxidant anti-inflammatory treatment in type 2 diabetes. Diabetes Care 2009, 32, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Haber, C.A.; Lam, T.K.; Yu, Z.; Gupta, N.; Goh, T.; Bogdanovic, E.; Giacca, A.; Fantus, I.G. N-acetylcysteine and taurine prevent hyperglycemia-induced insulin resistance in vivo: Possible role of oxidative stress. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E744–E753. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Hutchings, S.; Pang, C.C.Y. Chronic N-acetylcysteine prevents fructose-induced insulin resistance and hypertension in rats. Eur. J. Pharmacol. 2005, 508, 205–210. [Google Scholar] [CrossRef]
- Diniz, Y.S.; Rocha, K.K.; Souza, G.A.; Galhardi, C.M.; Ebaid, G.M.X.; Rodrigues, H.G.; Novelli Filho, J.V.B.; Cicogna, A.C.; Novelli, E.L.B. Effects of N-acetylcysteine on sucrose-rich diet-induced hyperglycaemia, dyslipidemia and oxidative stress in rats. Eur. J. Pharmacol. 2006, 543, 151–157. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, E.; Davies, K.J. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Xiang, R.L.; Huang, Y.; Zhang, Y.; Cong, X.; Zhang, Z.J.; Wu, L.L.; Yu, G.Y. Type 2 diabetes-induced hyposalivation of the submandibular gland through PINK1/Parkin-mediated mitophagy. J. Cell Physiol. 2020, 235, 232–244. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Marcillat, O.; Giulivi, C.; Ernster, L.; Davies, K.J. The oxidative inactivation of mitochondrial electron transport chain components and ATPase. J. Biol. Chem. 1990, 265, 16330–16336. [Google Scholar] [PubMed]
- Martinez, M.; Martinez, N.; Hernandez, A.I.; Ferrandiz, M.L. Hypothesis: Can N-acetylcysteine be beneficial in Parkinson’s disease? Life Sci. 1999, 64, 1253–1257. [Google Scholar] [CrossRef]
- Oh, S.H.; Lim, S.C. A rapid and transient ROS generation by cadmium tiggers apoptosis via caspase-dependent pathway in HepG2 cells and this is inhibited through N-acetylcysteine-mediated catalase upregulation. Toxicol. Appl. Pharmacol. 2006, 212, 212–223. [Google Scholar] [CrossRef]
- Zaragoza, A.; Diez-Fernandez, C.; Alvarez, A.M.; Andres, D.; Cascales, M. Effect of N-acetylcysteine and deferoxamine on endogenous antioxidant defense system gene expression in a rat hepatocyte model of cocaine cytotoxicity. Biochim. Biophys. Acta Mol. Cell. Res. 2000, 1496, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Martensson, J.; Lai, J.C.; Meister, A. High-affinity transport of glutathione is part of a multicomponent system essential for mitochondrial function. Proc. Natl. Acad. Sci. USA 1990, 87, 7185–7189. [Google Scholar] [CrossRef] [Green Version]
- Selles, B.; Hugo, M.; Trujillo, M.; Srivastava, V.; Wingsle, G.; Jacquot, J.P.; Radi, R.; Rouhier, N. Hydroperoxide and peroxynitrite reductase activity of poplar thioredoxin-dependent glutathione peroxidase 5: Kinetics, catalytic mechanism and oxidative inactivation. Biochem. J. 2012, 442, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, R.Z.; Li, R.Y. Essential of Free Radical Biology and Medicine; Cell Med Press AIMSCI, Inc.: Raleigh, NC, USA, 2017. [Google Scholar]
- Samuni, Y.; Goldstein, S.; Dean, O.M.; Berk, M. The chemistry and biological activities of N-acetylcysteine. Biochim. Biophys. Acta 2013, 1830, 4117–4129. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Metodiewa, D. Reactivity of biologically important thiol compounds with superoxide and hydrogen peroxide. Free Radic. Biol. Med. 1999, 27, 322–328. [Google Scholar] [CrossRef]
- Russell, S.L.; Reisine, S. Investigation of xerostomia in patients with rheumatoid arthritis. JADA 1998, 129, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Burgunder, J.M.; Varriale, A.; Lauterburg, B.H. Effect of N-acetylcysteine on plasma cysteine and glutathione following paracetamol administration. Eur. J. Clin. Pharmacol. 1989, 36, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Haugaard, N.; Lee, N.H.; Kostrzewa, R.; Horn, R.S.; Haugaard, E.S. The role of sulfhydryl groups in oxidative phosphorylation and ion transport by rat liver mitochrondia. Biochim. Biophys. Acta 1969, 172, 198–204. [Google Scholar] [CrossRef]
- Garcia-Ruiz, I.; Solis-Munoz, P.; Fernandez-Moreira, D.; Grau, M.; Colina, F.; Munoz-Yague, T.; Solis-Herruzo, J.A. High-fat diet decreases activity of the oxidative phosphorylation complexes and causes nonalcoholic steatohepatitis in mice. Dis. Models Mech. 2014, 7, 1287–1296. [Google Scholar] [CrossRef] [Green Version]
- Valenti, D.; Manente, G.A.; Moro, L.; Marra, E.; Vacca, R.A. Deficit of complex I activity in human skin fibroblasts with chromosome 21 trisomy and overproduction of reactive oxygen species by mitochondria: Involvement of the cAMP/PKA signalling pathway. Biochem. J. 2011, 435, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Block, K.; Gorin, Y.; Abboud, H.E. Subcellular localization of Nox4 and regulation in diabetes. Proc. Natl. Acad. Sci. USA 2009, 106, 14385–14390. [Google Scholar] [CrossRef] [Green Version]
- Takac, I.; Schroder, K.; Zhang, L.; Lardy, B.; Anilkumar, N.; Lambeth, J.D.; Shah, A.M.; Morel, F.; Brandes, R.P. The E-loop is involved in hydrogen peroxide formation by the NADPH oxidase Nox4. J. Biol. Chem. 2011, 286, 13304–13313. [Google Scholar] [CrossRef] [Green Version]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complication. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Hatanaka, E.; Dermargos, A.; Hirata, A.E.; Vinolo, M.A.; Carpinelli, A.R.; Newsholme, P.; Armelin, H.A.; Curi, R. Oleic, linoleic and linolenic acids increase ros production by fibroblasts via NADPH oxidase activation. PLoS ONE 2013, 8, e58626. [Google Scholar] [CrossRef] [Green Version]
- Forner, L.; Hyldegaard, O.; von Brockdorff, A.S.; Specht, L.; Andersen, E.; Jansen, E.C.; Hillerup, S.; Nauntofte, B.; Jensen, S.B. Does hyperbaric oxygen treatment have the potential to increase salivary flow rate and reduce xerostomia in previously irradiated head and neck cancer patients? A pilot study. Oral Oncol. 2011, 47, 546–551. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | C + NAC | HFD | HFD + NAC | |
|---|---|---|---|---|
| Final body weight (g) | 280.0 ± 12.31 | 278.3 ± 26.62 | 380.9 ± 30.42 * | 314.5 ± 32.78 *,# |
| Plasma glucose (mg/dL) | 90.67 ± 6.19 | 91.77 ± 11.84 | 148.6 ± 7.99 * | 98.50 ± 8.41 *,# |
| Plasma insulin (mIU/mL) | 78.92 ± 8.46 | 76.86 ± 9.75 | 165.4 ± 12.79 * | 84.49 ± 17.07 *,# |
| HOMA-IR | 2.93 ± 1.36 | 2.56 ± 2.45 | 20.11 ± 1.96 * | 2.89 ± 1.83 # |
| Plasma free fatty acids (µmol/L) | 75.59 ± 10.99 | 69.80 ± 8.83 | 173.4 ± 11.99 * | 86.38 ± 12.5 *,# |
| Food intake (mg/day) | 20.98 ± 3.95 | 21.70 ± 6.93 | 11.12 ± 6.1 * | 12.19 ± 5.68 *,# |
| Energy from chow (MJ/day) | 0.19 ± 0.1 | 0.17 ± 0.1 | 0.31 ± 0.12 * | 0.27 ± 0.1 * |
| Non-stimulated salivary flow (µL/min) | 0.4229 ± 0.12 | 0.3813 ± 0.11 | 0.3707 ± 0.14 | 0.4195 ± 0.09 |
| Stimulated salivary flow (µL/min) | 110.2 ± 9.77 | 103.5 ± 10.76 | 70.08 ± 8.14 * | 105.2 ± 12.05 *,# |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zalewska, A.; Szarmach, I.; Żendzian-Piotrowska, M.; Maciejczyk, M. The Effect of N-Acetylcysteine on Respiratory Enzymes, ADP/ATP Ratio, Glutathione Metabolism, and Nitrosative Stress in the Salivary Gland Mitochondria of Insulin Resistant Rats. Nutrients 2020, 12, 458. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020458
Zalewska A, Szarmach I, Żendzian-Piotrowska M, Maciejczyk M. The Effect of N-Acetylcysteine on Respiratory Enzymes, ADP/ATP Ratio, Glutathione Metabolism, and Nitrosative Stress in the Salivary Gland Mitochondria of Insulin Resistant Rats. Nutrients. 2020; 12(2):458. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020458
Chicago/Turabian StyleZalewska, Anna, Izabela Szarmach, Małgorzata Żendzian-Piotrowska, and Mateusz Maciejczyk. 2020. "The Effect of N-Acetylcysteine on Respiratory Enzymes, ADP/ATP Ratio, Glutathione Metabolism, and Nitrosative Stress in the Salivary Gland Mitochondria of Insulin Resistant Rats" Nutrients 12, no. 2: 458. https://0-doi-org.brum.beds.ac.uk/10.3390/nu12020458