Detection and Genomic Characterisation of Clostridioides difficile from Spinach Fields


1
Teagasc Food Research Centre, Ashtown, D15 DY05 Dublin, Ireland
2
School of Veterinary Medicine, University College Dublin, Belfield, D04 V1W8 Dublin, Ireland
*
Author to whom correspondence should be addressed.
Pathogens 2022, 11(11), 1310; https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11111310
Submission received: 30 September 2022
/
Revised: 27 October 2022
/
Accepted: 2 November 2022
/
Published: 8 November 2022
Abstract
:Despite an increased incidence of Clostridioides difficile infections, data on the reservoirs and dissemination routes of this bacterium are limited. This study examined the prevalence and characteristics of C. difficile isolates in spinach fields. C. difficile was detected in 2/60 (3.3%) of spinach and 6/60 (10%) of soil samples using culture-based techniques. Whole genome sequencing (WGS) analysis identified the spinach isolates as belonging to the hypervirulent clade 5, sequence type (ST) 11, ribotypes (RT) 078 and 126 and carried the genes encoding toxins A, B and CDT. The soil isolates belonged to clade 1 with different toxigenic ST/RT (ST19/RT614, ST12/RT003, ST46/RT087, ST16/RT050, ST49/RT014/0) strains and one non-toxigenic ST79/RT511 strain. Antimicrobial resistance to erythromycin (one spinach isolate), rifampicin (two soil isolates), clindamycin (one soil isolate), both moxifloxacin and rifampicin (one soil isolate), and multi-drug resistance to erythromycin, vancomycin and rifampicin (two soil isolates) were observed using the E test, although a broader range of resistance genes were detected using WGS. Although the sample size was limited, our results demonstrate the presence of C. difficile in horticulture and provide further evidence that there are multiple sources and dissemination routes for these bacteria.
1. Introduction
Clostridioides difficile is an enteric spore-forming and toxigenic pathogen that was historically classified as a hospital-acquired infection [1]. Common symptoms range from watery non-bloody diarrhoea with abdominal pain to life-threatening fulminant colitis [2]. However, the steady increase in community acquired C. difficile infection(s) (CDI) in recent years has motivated investigative research on other sources and dissemination routes. As a result, specific C. difficile ribotypes, such as RT078, are now being classified as community-acquired [3,4].
Virulence in C. difficile is mainly determined by the production of toxin A (tcdA) and B (tcdB), which alter the host’s gut by causing damage to the epithelial barrier, leading to the translocation of commensal bacteria and cell death [5,6,7]. In addition, accessory genes tcdR and tcdC are part of the pathogenicity locus (PaLoc) and play a key role in regulating toxin production [8,9]. Apart from toxins A and B, it is estimated that up to 30% of C. difficile strains can produce the transferase C. difficile binary toxin (CDT), which belongs to the binary toxin family and is generally associated with hypervirulent strains [10,11,12].
Epidemiological investigations of C. difficile cases often use ribotyping that differentiates strains based on polymorphisms in the 16S-23S rRNA intergenic spacer region [13]. Certain ribotypes are more often linked with a higher occurrence and severity of disease in humans and some of these have been isolated from the food chain. Multi-drug resistant RT027, for example, is often isolated from clinical samples and is frequently associated with more severe illnesses and has been isolated from different food animals [14,15,16]. Moreover, RT014/020 and RT001/072 are confirmed endemic ribotypes in Europe [17].
In addition to its toxigenic virulence, antimicrobial resistance among the C. difficile strains is common worldwide, driven by the increased use of antibiotics as a treatment therapy for CDI [18]. For example, the emergence of C. difficile RT027 and its high resistance to fluoroquinolones is linked to the extensive use of these antibiotics in human cases [19]. Indeed, clinical isolates are often resistant to clindamycin, cephalosporins and penicillins, in addition to fluoroquinolones [20], with emergent strains also showing resistance to several antibiotics simultaneously [21].
While it is known that the bacterium colonizes the gut after transmission to a new host, the primary habitat of C. difficile is soil [22,23]. Thus, C. difficile spores have been frequently reported in soil on farms, in gardens and in forests [24,25,26]. Its ability to form spores facilitates survival and dissemination of this organism via organic fertilizers, wind and water. Thus, crops including vegetables are inevitably contaminated, resulting in a potential risk of infection for consumers [27,28,29,30]. Several studies have reported the detection of C. difficile in vegetables at retail [31,32,33], including salad leaves [24,34] and ready-to-eat (RTE) salads [31,35,36]. The C. difficile ribotypes identified in these studies include RT056, which has been associated with human illness [37]. However, further information is required to facilitate a better understanding of the role of vegetables in human CDI. Moreover, additional data on the genomic profile of the isolates from such sources would add to our knowledge of this emergent pathogen.
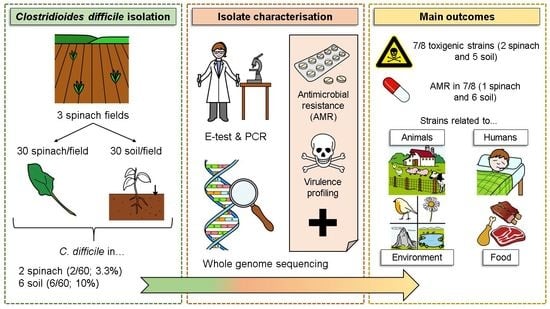
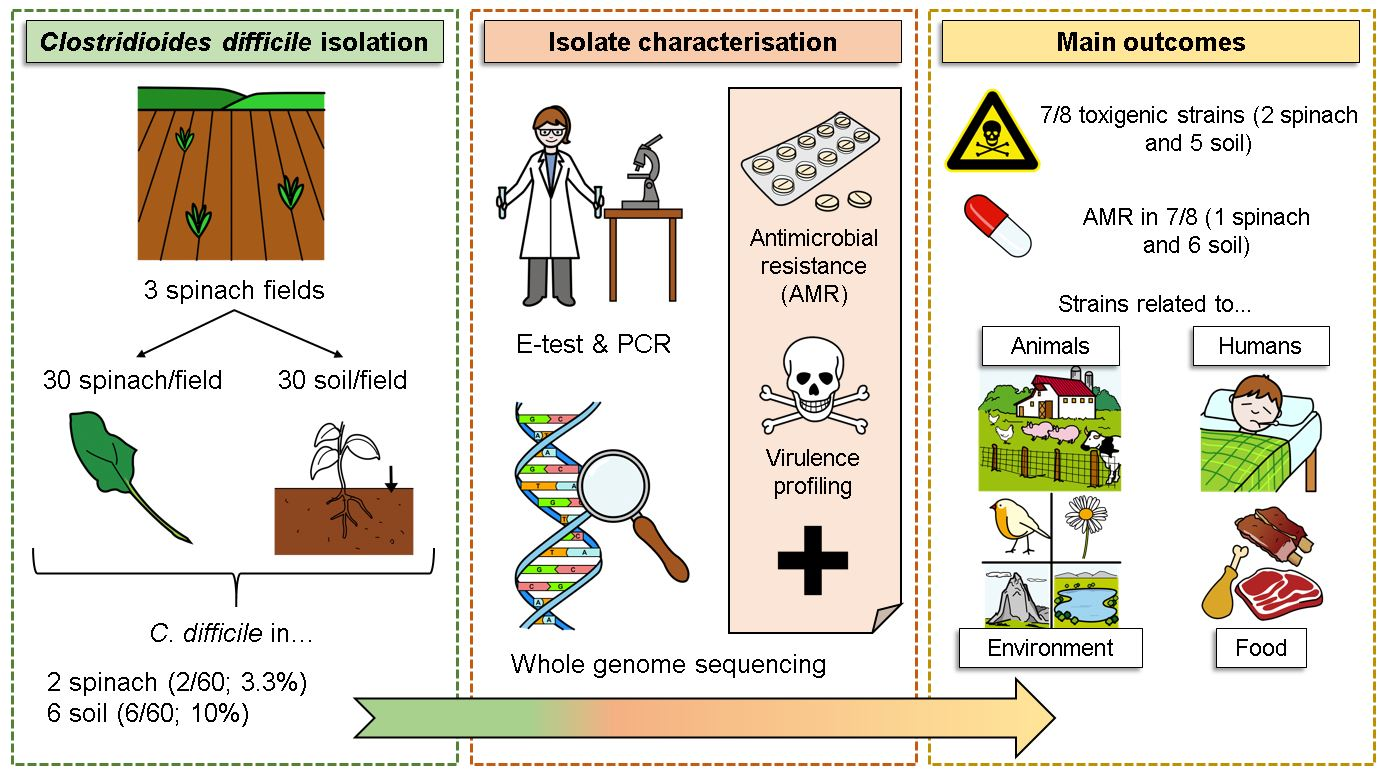
The objectives of this study were to (i) determine the prevalence of C. difficile in spinach and soil samples from fields where spinach was grown, (ii) characterise the C. difficile isolates originating from spinach and associated soil to determine their virulence and antimicrobial resistance using laboratory-based techniques and WGS, and (iii) compare the ribotypes and strains detected to those previously reported to establish if they are limited to the vegetable production environment or are part of the growing collection of C. difficile isolated from environmental, animal, food and/or human sources.
2. Materials and Methods
2.1. Sample Collection
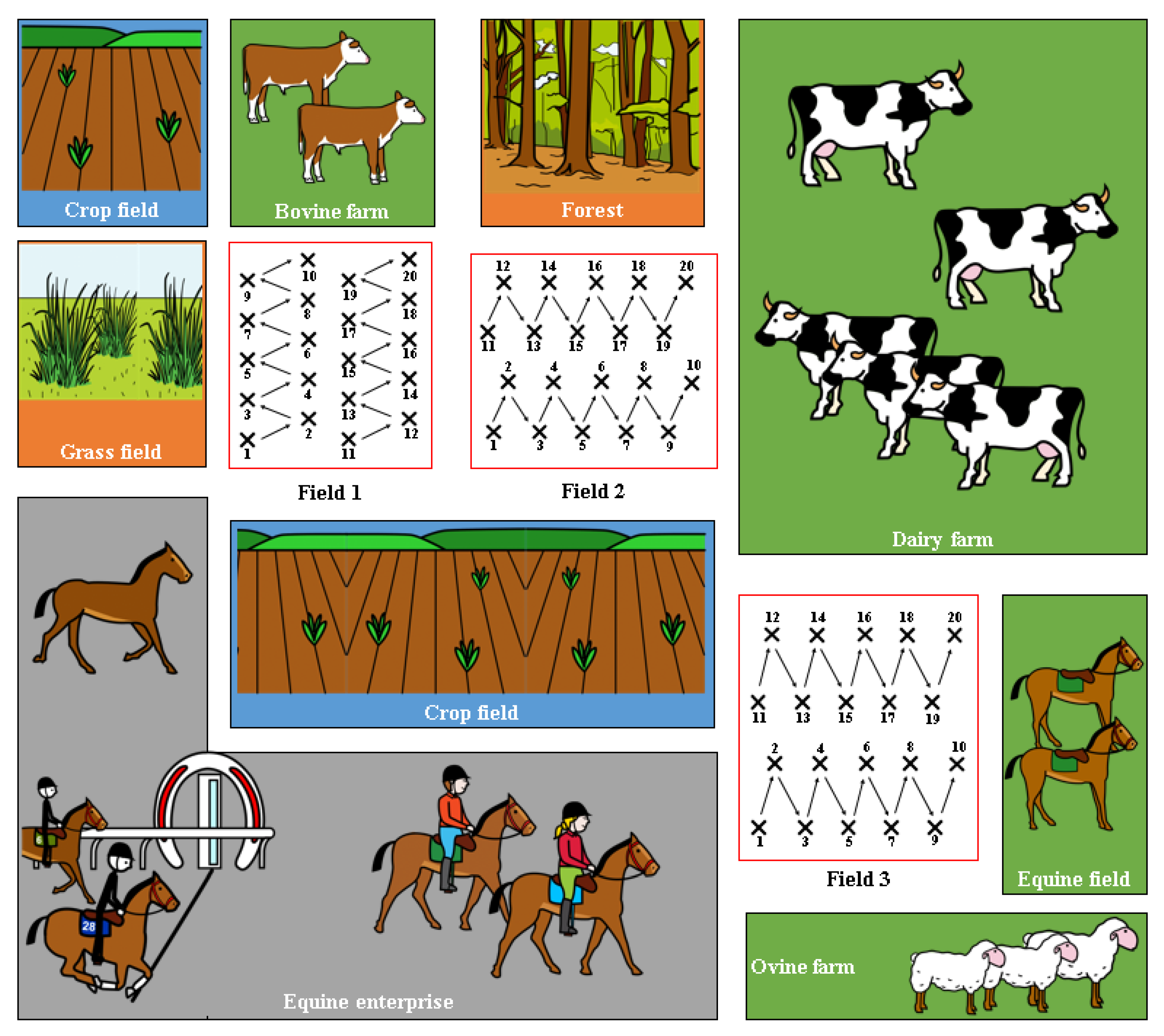
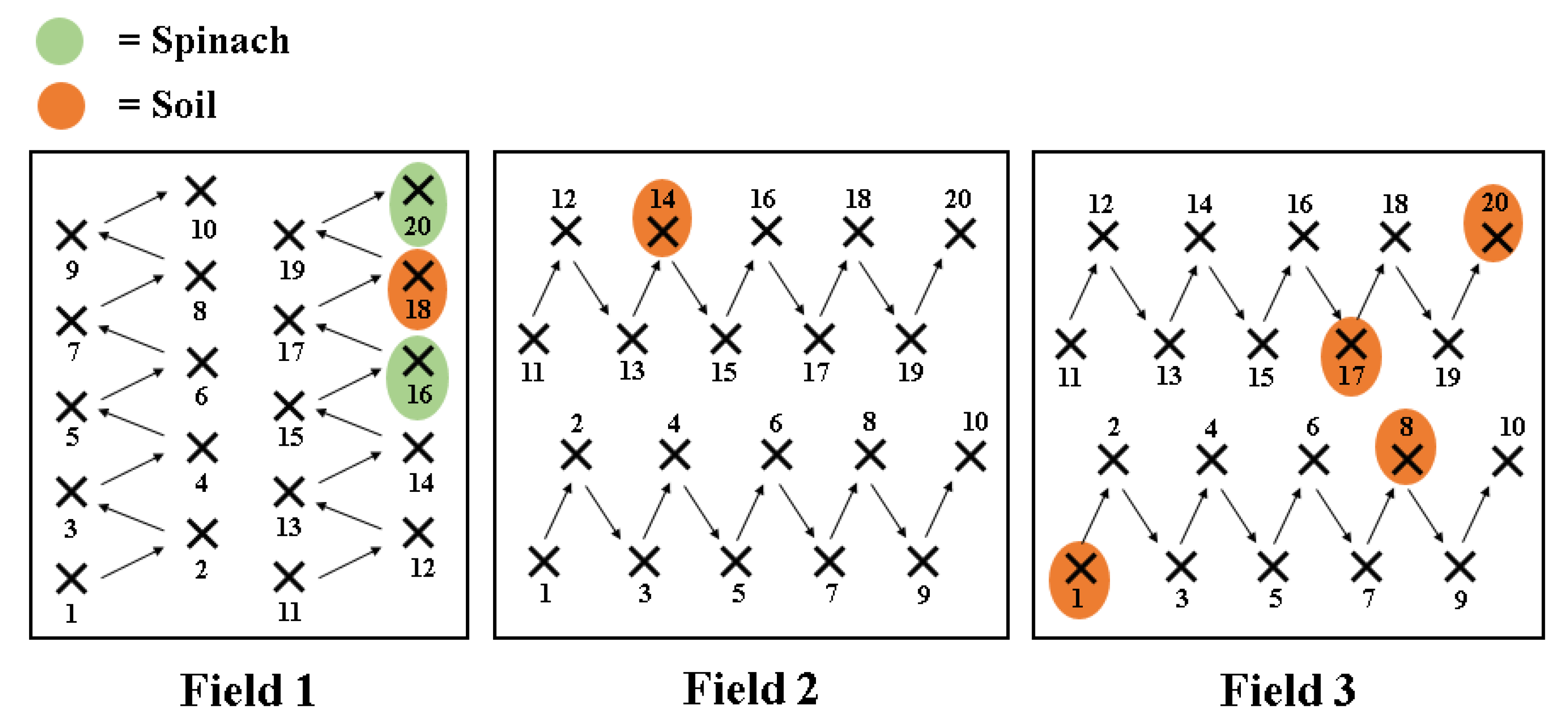
Soil (n = 60) and spinach (n = 60) samples were obtained from three different spinach fields in the same area in Co. Meath (north-east Ireland). Field 1 and Field 2 were adjacent to each other, while Field 3 was bordered by a sheep farm on one side and a dairy farm on the other (Figure 1). In each field, 20 soil and 20 spinach samples were collected in 250 mL Ramboldi sterile containers (Sparks Lab Supplies, Rathcoole, Dublin, Ireland) using sterile sampling scoops (VWR, Ballycoolin, Dublin, Ireland). Soil samples were collected from directly under the spinach plants tested. In addition, sampling was carried out in a zig-zag pattern to cover the entirety of the field (Figure 1). Samples were then transported in a cool box (at approximately 4 °C) to the laboratory for testing within 24 h. None of the fields had been amended with slurry in the 12 months prior to the sampling date.
2.2. C. difficile Isolation and Confirmation
Soil and spinach samples were tested for C. difficile by adding 90 mL of maximum recovery diluent (MRD) to 10 g of each sample in a stomacher bag and blending for 1 min in a Star Blender LB 400 Stomacher (VWR, Lutterworth, Leicestershire, UK). To ensure the selection of C. difficile spores, vegetative cells were eliminated by heating at 60 °C for 25 min. To encourage spore germination, 10 mL of the heat-treated mixture was added to 90 mL of brain heart infusion broth (BHI) (Oxoid, Basingstoke, Hampshire, UK (CM1135B)) with 0.1% sodium taurocholate (Sigma-Aldrich, Gillingham, UK (86339-25G)) and selective C. difficile supplements (containing 8 mg/L of cefoxitin and 250 mg/L of D-cycloserine) (Oxoid, Basingstoke, Hampshire, UK (SR0096E)). Samples were incubated at 37 °C for 3–5 days under anaerobic conditions in an A35 Anaerobic workstation (Don Whitley, Victoria Works, West Yorkshire, UK) [38].
Exactly 10 µL of the broth culture was then streaked onto C. difficile moxalactam norfloxacin agar (Oxoid, Basingstoke, Hampshire, UK (CM0601B)) supplemented with 0.1% sodium taurocholate (Sigma-Aldrich, Gillingham, UK (86339-25G)) (CDMN-T), 32 mg/L moxalactam, 12 mg/L norfloxacin, 500 mg/L cysteine hydrochloride (Oxoid, Basingstoke, Hampshire, UK (SR0173)) and 5% defibrinated horse blood (TCS Biosciences Limited, Botolph Claydon, Buckingham, UK (HB034)). These plates were incubated at 37 °C for 48 h under anaerobic conditions as previously described [15].
DNA was extracted from isolated colonies with the typical C. difficile morphology (irregular ground-glass colonies) using the DNeasy blood & tissue kit (QIAGEN GmbH, Hilden, Germany (69504)) and was confirmed as C. difficile by testing for the tpi gene using PCR [39]. Briefly, 12.5 µL of Mastermix (QIAGEN Ltd., Manchester, UK (206143)), 0.625 µL of each primer (0.5 µM), 9.25 µL of nuclease-free water (Invitrogen, Biosciences Ltd., Dún Laoghaire, Dublin, Ireland (LSKNF0500)) and 2 µL of template were mixed together and amplified using a Veriti 96-Well Thermal Cycler (Applied Biosystems, Warrington, Cheshire, UK). The PCR cycle started with denaturation for 3 min at 95 °C, followed by 40 cycles of 30 s at 95 °C, 30 s at 55 °C and 30 s at 72 °C, and a final extension of 30 s at 72 °C. Electrophoresis in a 2% agarose gel stained with SYBR™ Safe DNA Gel Stain (Biosciences Ltd., Dún Laoghaire, Dublin, Ireland (S33102)) was then used to confirm the tpi PCR fragment generated (230-bp) under UV light. C. difficile RT078, obtained from a clinical isolate at St. James’s Hospital (Dublin, Ireland) and supplied by Prof. Thomas Rogers from Trinity College Dublin, and C. sporogenes, available in the Teagasc Ashtown Food Research Centre collection, were used as the positive and negative controls, respectively.
2.3. Characterization of Toxin and Accessory Genes
The confirmed C. difficile isolates obtained were further characterised for toxin (tcdA, tcdB, cdtA and cdtB) and accessory genes (tcdC and tcdR) by conventional PCR. Primer sequences, concentration and amplification protocols for toxin A (tcdA), toxin B (tcdB), toxin CDT (cdtA and cdtB) and accessory genes (tcdC and tcdR) were as described by Marcos et al. [24]. The total PCR mixture for every reaction had a final volume of 25 µL with the corresponding volume for each primer depending on its concentration, 12.5 µL of Mastermix (QIAGEN), 2 µL of template and nuclease-free water to reach the total volume. As previously described, a 2% agarose gel stained with SYBR™ Safe DNA Gel Stain (Biosciences) was carried out to separate the PCR fragments: 369-bp for tcdA, 160-bp for tcdB, 375-bp for cdtA, 510-bp for cdtB, 718 bp for tcdC and 300 bp for tcdR. C. difficile RT078 and C. sporogenes were the corresponding positive and negative controls, respectively.
2.4. Antimicrobial Susceptibility Testing
The susceptibility of isolates to a range of relevant antibiotics (vancomycin, erythromycin, metronidazole, clindamycin, moxifloxacin and rifampicin) was tested using the E-test strips (bioMérieux, Marcy-l’Étoile, France), following the protocol described by the manufacturer. The aforementioned antibiotics were selected for testing due to their common use in CDI treatment [40,41,42,43,44].
Isolates were cultured in Mueller–Hinton broth (Oxoid, Basingstoke, Hampshire, UK (CM0405)) at 37 °C for 24 h or until an OD600 nm = 0.5 (108 CFU/mL) was achieved, measured using a DeNovix DS-C spectrophotometer (DeNovix Inc., Wilmington, NC, USA). A sterile swab (Sparks Lab Supplies, Dublin, Ireland (SW001)) was dipped into the broth solution and then spread on Brucella agar plates with vitamin K and haemin (Sigma-Aldrich, Gillingham, UK (B2926-500G)) and 5% defibrinated horse blood (TCS Biosciences Limited, Botolph Claydon, Buckingham, UK (HB034)). Plates were allowed to dry for 15 min before the E-test strip was placed on top of the inoculated agar with a sterile forceps and incubated for 48 h at 37 °C in anaerobic conditions as previously described. After incubation, MIC values were read using the scale (µg/mL) provided by the manufacturer. Values obtained for each antibiotic were compared to the breakpoint values described in EUCAST [45] and classified as susceptible or resistant according to the criteria.
2.5. PCR-Ribotyping
Ribotype testing of all isolates obtained was carried out following the PCR ribotyping protocol by ECDC [46], with minor changes. Amplification of the 16S and 23S rRNA genes was undertaken to identify the specific intergenic spacer region (ISR). Primers for the amplification of the C. difficile 16S (forward primer) and 23S (reverse primer) rRNA genes were as described by Bidet et al. [47] (5′ GCTGGATCACCTCCTTT CTAAG (6FAM-16S) and 5′ TGACCAGTTAAAAAGG TTTGATAGATT (23S)). The PCR required 12.5 µL of HotStarTaq Mastermix (QIAGEN Ltd., Manchester, UK), 0.25 µL of each primer (10 μM), 10 µL of RNase-free water (QIAGEN Ltd., Manchester, UK) and 2 µL of the template. The mixture was added to a MicroAmpTM Optical 96-well reaction plate (Applied Biosystems, Warrington, Cheshire, UK) before being sealed and inserted in a Veriti 96-Well Thermal Cycler (Applied Biosystems, Warrington, Cheshire, UK). Amplification consisted of a denaturation of 15 min at 95 °C, 30 cycles of 1 min at 94 °C, 1 min at 60 °C and 1 min at 72 °C, and a final extension of 30 min at 72 °C.
After the PCR, denaturation of the generated fragments was necessary before analysis in an automated sequence and fragment analysis system was undertaken. Exactly 2 µL of the PCR product was added to 9.5 µL of Highly Deionized (Hi-Di) Formamide (Applied Biosystems, Warrington, Cheshire, UK) and 0.5 µL of GeneScan 1200 LIZ Size Standard (Applied Biosystems, Warrington, Cheshire, UK). The mix was denatured for 2 min at 95 °C in a thermal cycler, followed by cooling of the plate for 10 min in a fridge before the PCR products were analysed on an ABI 3500 Genetic Analyzer (Applied Biosystems, Warrington, Cheshire, UK) with default settings for POP7 and 50 cm capillary length. The raw data files (*fsa files) from the ABI 3500 Genetic Analyzer were uploaded to the free-to-use website WEBRIBO (https://webribo.ages.at/; accessed on 7 July 2022) to compare our isolates with existing ribotypes stored on the database [13].
2.6. Whole Genome Sequencing (WGS)
The C. difficile isolates (n = 8) obtained from 2 spinach leaves and 6 soil samples were selected for WGS in order to identify genetic determinants of virulence and resistance. WGS was performed in Teagasc Food Research Centre Moorepark by Dr. Fiona Crispie and Dr. Gaston Allende. DNA libraries were prepared with the Illumina DNA preparation kit as per the manufacturer’s instructions. This was followed by 2 × 150 bp sequencing on the NextSeq 2000 with P2 reagents. Nucleotide sequence data reported are available in the GenBank database under the accession number PRJNA884639.
After quality analysis (FastQC v0.11.8) [48], sequences were trimmed using Trimmomatic (v.0.38) [49]. Paired sequences were subjected to taxonomic annotation using Kraken2 (v.2.1.1) [50] to initially identify the isolates as C. difficile, and fragments were de novo assembled using SPAdes (v.3.15.3) [51]. After the contigs’ quality was analysed (Quast v.5.1.0) [52], functional gene annotation was carried out via Prokka (v.1.14) [53]. Further taxonomic analysis was carried out with the contigs using KmerFinder (v.3.2) [54,55,56]. The presence of putative toxin genes (tcdA, tcdB, cdtA, cdtB) and plasmids was analysed using ABRicate (v.1.0.1) (PlasmidFinder and Virulence Factor Database (Vfdb)) [57,58,59]. The presence of antimicrobial resistance genes was determined using the ResFinder function in ABRicate and Resistance Gene Identifier (RGI) and CARD [60,61]. Allelic profile analysis was carried out via MLST (v.2.18.0) [62] and uploaded to the open-access online platform PubMLST (http://pubmlst.org, accessed on 23 June 2022) to determine the sequence type (ST) and clade. Single nucleotide polymorphism (SNP) analysis was performed using Snippy (v.4.3.6), using the genome of the C. difficile 630 strain as a reference [63]. The pangenome analysis of the C. difficile isolates was performed using anvi’o (v7.1) as per the workflow for microbial pangenomics (https://merenlab.org/2016/11/08/pangenomics-v2/; accessed on 27 September 2022) [64]. For this purpose, assembled scaffold.fasta files were transformed into anvi’o contig databases using the ‘anvi-gen-contigs-database’ program. Then, genes in scaffolds were identified via Prodigal to screen for open reading frames and were annotated using the NCBI’s Clusters of Orthologous Groups database (‘anvi-run-ncbi-cogs’ program) [65,66] against four provided HMM profiles of anvi’o using hidden Markov models (‘anvi-run-hmms’ program). The pangenome was computed by calculating the amino acid sequence similarities and comparing them across all genomes via NCBI blastp. In addition, weak matches between amino acid sequences were eliminated using minbit heuristics of 0.5 [67] and clusters were identified using the MCL algorithm [68] (‘anvi-pan-genome’ program). The pangenome was visualized and edited using the anvi’o interactive interface and the program ‘anvi-display-pan’, and the arrangement and quality of figures was improved using Affinity Designer (v1.8.5.703, Serif). A phylogenetic tree was created using the NDtree 1.2 program available in the Center for Genomic Epidemiology [69,70,71].
3. Results
3.1. Characterisation of the C. difficile Isolates Using Conventional PCR, E-Test and PCR-Ribotyping
A total of 8 (two spinach and six soil) out of 120 samples analysed (6.7%) were confirmed as C. difficile positive (Figure 2). In Field 1, two spinach and one soil samples were positive, while one positive soil sample was obtained in Field 2 (Figure 2). Half of the positive samples (four soils) were obtained in Field 3.
The presence or absence of toxin and accessory genes in the C. difficile isolates is shown in Table 1. Toxin-encoding genes were detected by PCR in seven of the eight isolates. The tcdA and tcdB genes were detected in five out of the six soil isolates, while tcdB, cdtA and cdtB were present in the two spinach isolates. Accessory genes (tcdC and tcdR) were detected in all isolates except in isolate 7, which also lacked the PaLoc.
The resistance or susceptibility of the C. difficile isolates to the six antibiotics tested and their corresponding ribotype is also shown in Table 1. All isolates, except for one spinach isolate (ribotype 078) from Field 1, were resistant to at least one antibiotic. Resistance to rifampicin was observed in 5/8 of the isolates, followed by erythromycin (3/8), vancomycin (2/8), clindamycin and moxifloxacin (1/8 each). All isolates were susceptible to metronidazole. Two soil isolates from Fields 2 (RT003) and 3 (RT050) were resistant to the same three antibiotics (erythromycin, vancomycin and rifampicin). The rest of the isolates presented varied resistance and ribotype patterns.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Ribotype (PCR-ribotyping) and antibiotic susceptibility testing profile using E-test against erythromycin (ERY), metronidazole (MET), clindamycin (CLIN), moxifloxacin (MOX), vancomycin (VAN) and rifampicin (RIF) (S = susceptible; R = resistant) and virulence gene (tcdA, tcdB, cdtA, cdtB, tcdC, tcdR) of the C. difficile strains isolated from spinach fields.
Table 1.
Ribotype (PCR-ribotyping) and antibiotic susceptibility testing profile using E-test against erythromycin (ERY), metronidazole (MET), clindamycin (CLIN), moxifloxacin (MOX), vancomycin (VAN) and rifampicin (RIF) (S = susceptible; R = resistant) and virulence gene (tcdA, tcdB, cdtA, cdtB, tcdC, tcdR) of the C. difficile strains isolated from spinach fields.
| Collection Field | Sample Type | ID 1 | RT 2 | Antibiotic Resistance Profile (MIC) | Toxin Genes 3 | Accessory Genes 3 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ERY | MET | CLIN | MOX | VAN | RIF | tcdA | tcdB | cdtA | cdtB | tcdC | tcdR | ||||
| Field 1 | Soil | 1 | 614 | S (0.25) | S (0.125) | S (8) | S (1) | S (0.75) | R (0.25) | + | + | − | − | + | + |
| Spinach | 2 | 078 | S (0.25) | S (<0.016) | S (<0.016) | S (0.38) | S (1.5) | S (<0.002) | − | + | + | + | + | + | |
| 3 | 126 | R (>256) | S (0.094) | S (4) | S (0.75) | S (1.5) | S (0.003) | − | + | + | + | + | + | ||
| Field 2 | Soil | 4 | 003 | R (8) | S (0.25) | S (1.5) | S (1) | R (>256) | R (0.125) | + | + | − | − | + | + |
| Field 3 | Soil | 5 | 087 | S (0.25) | S (0.125) | S (<0.016) | R (4) | S (1.5) | R (0.047) | + | + | − | − | + | + |
| Soil | 6 | 050 | R (4) | S (0.094) | S (1.5) | S (0.5) | R (3) | R (0.25) | + | + | − | − | + | + | |
| Soil | 7 | 511 | S (0.75) | S (0.094) | S (3) | S (0.75) | S (1.5) | R (0.19) | − | − | − | − | − | − | |
| Soil | 8 | 014/0 | S (<0.016) | S (<0.016) | R (16) | S (0.5) | S (1) | S (<0.002) | + | + | − | − | + | + | |
1 ID: number assigned to each isolate. 2 RT: Ribotype.3 (+/−): Presence (+) or absence (−) of the gene indicated.
3.2. Characterisation of the C. difficile Isolates by Whole Genome Sequencing (WGS)
The initial taxonomic annotation using Kraken2 confirmed all isolates were Clostridioides difficile (81.68–93.90%) (data not shown). Further species identification using KmerFinder suggested our isolates (1–8) had a high degree of similarity (>95%) with previously sequenced strains including C. difficile DSM 27639, M120, DSM 29020, W0023a, CFSAN096664, CFSAN096664, W0003a and DSM 105001, respectively.
The presence of toxin-encoding genes, sequence type (ST) and clade of the spinach (isolates 2 and 3) and soil (isolates 1, 4, 5, 6, 7, 8) isolates are shown in Table 2. The detection of toxin-encoding genes by WGS analysis was in broad agreement with the results obtained by PCR, with the exception of tcdA in the spinach isolates. Isolates 2 and 3 (both spinach isolates from Field 1) belonged to clade 5 and ST11. The rest of the isolates (obtained from soil from Fields 1, 2 and 3) belonged to clade 1 and had different STs (19, 12, 46, 16, 79 and 49).
Antibiotic resistance genes detected in the C. difficile isolates via ResFinder, Prokka and CARD software tools are shown in Table 3. ResFinder and Prokka analysis detected that isolates 2 and 3 (spinach from Field 1) carried resistance genes for aminoglycosides (D19aph(3’)-III_1, ant(6)-Ia_1, ant(6)-Ia_2, ant(6)-Ia_3) and tetracyclines (tet(M)_10, tet(40)_1, tet(M)_4) and all soil isolates, except for 7 and 8, had the vancomycin resistance gene (vanB). Genes encoding antibiotic resistance were not detected in isolates 7 and 8 (soil from Field 3) using ABRicate and Prokka. However, according to the analysis of the isolates using Resistance Gene Identifier (RGI) and CARD, all isolates encoded CDD-1 or CCD-2 (carbapenem resistance), qacG (disinfecting agents and antiseptics resistance) and a mutation in the 23S rRNA that confers the resistance to erythromycin and clindamycin. A spinach isolate (isolate 3) had resistance to nucleoside antibiotics (SAT-4), and soil isolates (1, 4, 5, 6, 7 and 8) carried resistance to glycopeptide antibiotics (vanXY and vanR in the vanG cluster). Isolate 5 (soil from Field 3) also encoded the cdeA gene, which confers resistance against fluoroquinolones.
A further comparison between the antimicrobial susceptibility profiles found by E-test and CARD is presented in Table 4.
Table 3.
Antimicrobial resistance (AMR) genes detected in C. difficile isolates from spinach (isolates 2 and 3) and soil (isolates 1, 4, 5, 6, 7, 8) samples using ResFinder, Prokka and CARD.
Table 3.
Antimicrobial resistance (AMR) genes detected in C. difficile isolates from spinach (isolates 2 and 3) and soil (isolates 1, 4, 5, 6, 7, 8) samples using ResFinder, Prokka and CARD.
| ID 1 | AMR Using ResFinder and Prokka | AMR Using CARD | ||
|---|---|---|---|---|
| Gene | Confers Resistance to | Gene | Confers Resistance to | |
| 1 | vanB2 | Vancomycin | vanXY gene in vanG cluster | Vancomycin |
| vanR gene in vanG cluster | ||||
| ND 3 | CDD-1 | Beta-lactams | ||
| ND 3 | qacG | Disinfecting agents and antiseptics | ||
| ND 3 | Clostridioides difficile 23S rRNA mutation | Erythromycin and clindamycin | ||
| 2 | tet(M)_10 | Tetracyclines | tet(M) | Tetracyclines |
| ant(6)-Ia_2 | Aminoglycosides | ant(6)-Ia | Aminoglycosides | |
| ND 3 | CDD-1 | Beta-lactams | ||
| ND 3 | qacG | Disinfecting agents and antiseptics | ||
| ND 3 | Clostridioides difficile 23S rRNA mutation | Erythromycin and clindamycin | ||
| 3 | D19aph(3′)-III_1 | Aminoglycosides | APH(3′)-IIIa | Aminoglycosides |
| ant(6)-Ia_1 | aad(6) | |||
| ant(6)-Ia_3 | ||||
| tet(40)_1 | Tetracyclines | tet(40) | Tetracyclines | |
| tet(M)_4 | ||||
| ND 3 | CDD-1 | Beta-lactams | ||
| ND 3 | SAT-4 | Nucleosides | ||
| ND 3 | qacG | Disinfecting agents and antiseptics | ||
| ND 3 | Clostridioides difficile 23S rRNA mutation | Erythromycin and clindamycin | ||
| 4 | vanB 2 | Vancomycin | vanXY gene in vanG cluster | Vancomycin |
| vanR gene in vanG cluster | ||||
| ND 3 | CDD-2 | Beta-lactams | ||
| ND 3 | qacG | Disinfecting agents and antiseptics | ||
| ND 3 | Clostridioides difficile 23S rRNA mutation | Erythromycin and clindamycin | ||
| 5 | vanB 2 | Vancomycin | vanXY gene in vanG cluster | Vancomycin |
| vanR gene in vanG cluster | ||||
| ND 3 | CDD-1 | Beta-lactams | ||
| ND 3 | cdeA | Fluoroquinolones, disinfecting agents and antiseptics | ||
| ND 3 | qacG | Disinfecting agents and antiseptics | ||
| ND 3 | Clostridioides difficile 23S rRNA mutation | Erythromycin and clindamycin | ||
| 6 | vanB 2 | Vancomycin | vanXY gene in vanG cluster | Vancomycin |
| vanR gene in vanG cluster | ||||
| ND 3 | CDD-1 | Beta-lactams | ||
| ND 3 | qacG | Disinfecting agents and antiseptics | ||
| ND 3 | Clostridioides difficile 23S rRNA mutation | Erythromycin and clindamycin | ||
| 7 | ND 3 | vanXY gene in vanG cluster | Vancomycin | |
| vanR gene in vanG cluster | ||||
| ND 3 | CDD-1 | Beta-lactams | ||
| ND 3 | qacG | Disinfecting agents and antiseptics | ||
| ND 3 | Clostridioides difficile 23S rRNA mutation | Erythromycin and clindamycin | ||
| 8 | ND 3 | vanXY gene in vanG cluster | Vancomycin | |
| vanR gene in vanG cluster | ||||
| ND 3 | CDD-1 | Beta-lactams | ||
| ND 3 | qacG | Disinfecting agents and antiseptics | ||
| ND 3 | Clostridioides difficile 23S rRNA mutation | Erythromycin and clindamycin | ||
1 ID: number assigned to each isolate. 2: The corresponding gene was found only in Prokka analysis. 3 ND: Not detected.
In addition, PlasmidFinder detected the genes of the replicon repUS43_1_CDS12738(DOp1) in the spinach isolates 2 and 3 (99.83% and 100% identity), respectively.
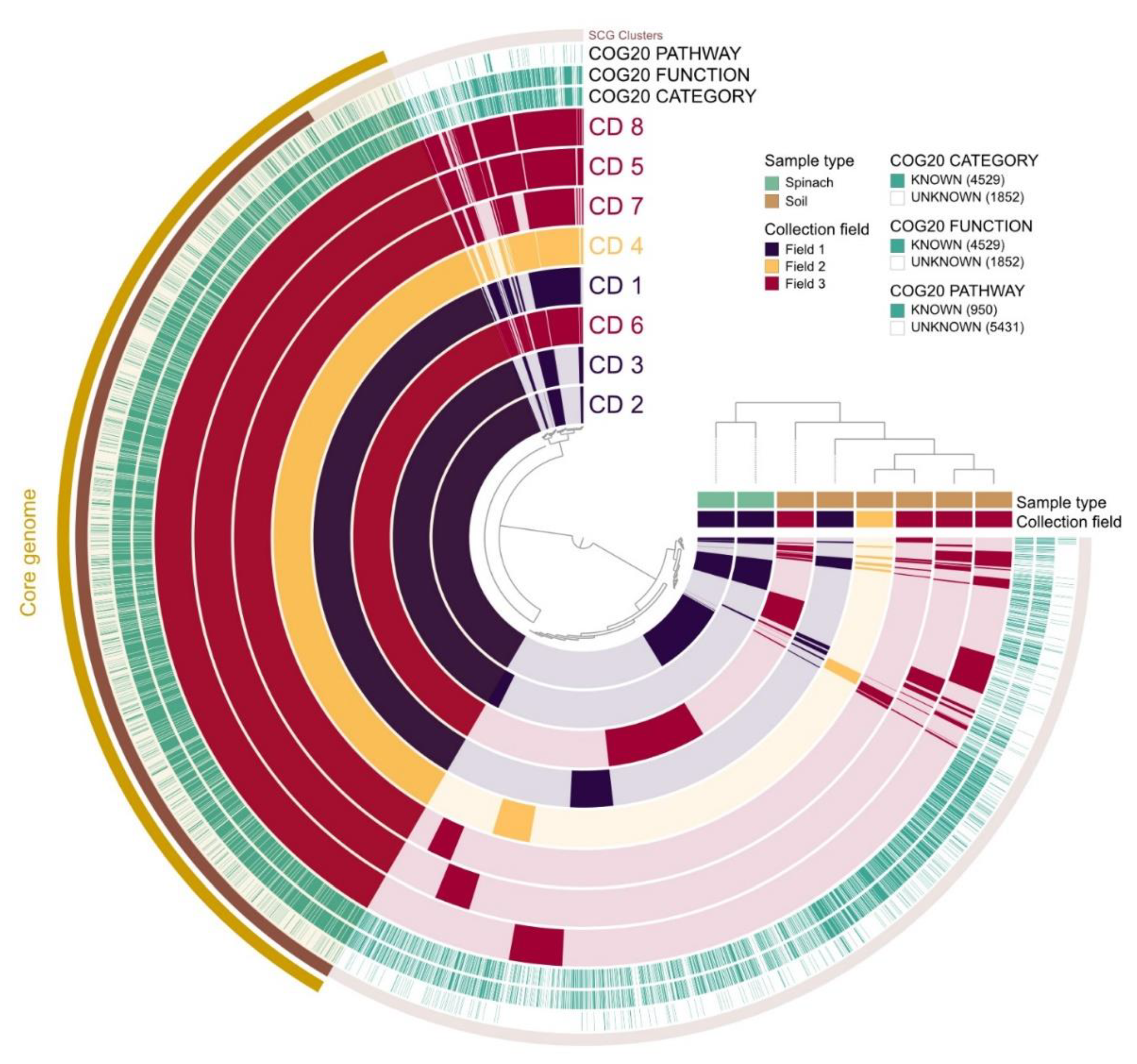
The pangenome analysis carried out for the 8 C. difficile isolates is shown in Figure 3. The core genome consisted of 3010 gene clusters and 25,718 genes. In the total pan-genome comparison, 6381 gene clusters were detected with 33,243 genes identified. Two distinct clusters were observed: the first cluster included isolates 2 and 3 (spinach), while the second cluster included the soil isolates (1, 4–8).
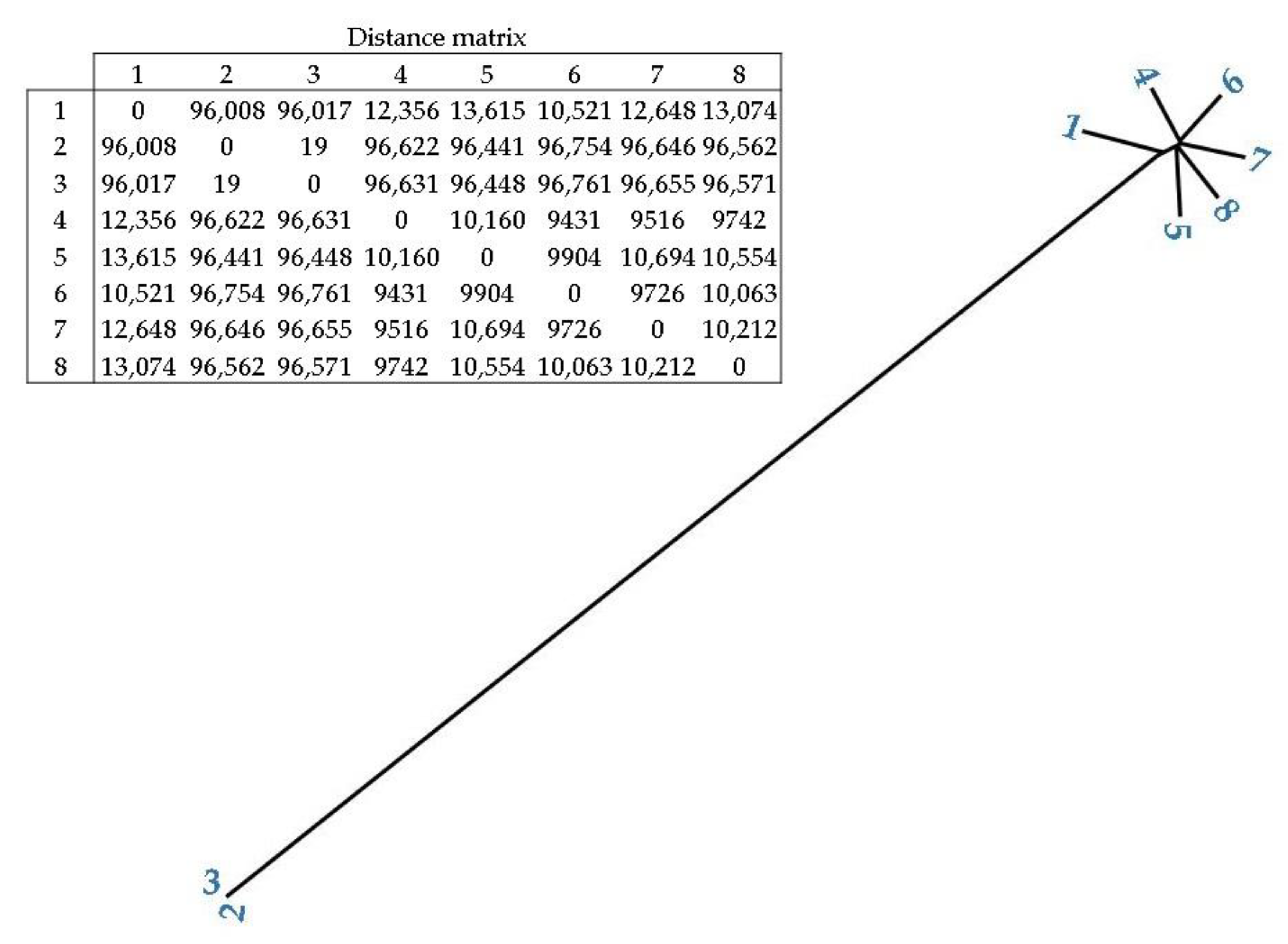
In addition, Figure 4 shows the phylogenetic relationship and distance between the C. difficile isolates. Isolates 2 and 3 (spinach isolates) had the closest distance (19), followed by soil isolates 4 and 6 (9431), and 4 and 7 (9516). Spinach isolates (2 and 3) presented a high distance from all of the soil isolates (>96,008).
4. Discussion
4.1. Prevalence of C. difficile
C. difficile was detected in 3.3% of spinach and 10% of soil samples. This was not unexpected as these bacteria have been previously isolated in compost (22.5%) [37], garden soil (79%), manure (83%) [26] and from soil samples in rural and urban areas (36.7%) [8]. Eckert et al. [31] reported a similar prevalence of C. difficile in salads purchased from retail stores in France (3.3%). A slightly lower prevalence was reported in lettuce (1.9%) in an Italian study by Primavilla et al. [34], but higher levels have also been found in salad leaves and ready-to-eat salads in Iran (5.6%) [36], in Ireland (6%) [24] and in Scotland (7.5%) [35]. Root vegetables are more frequently contaminated with C. difficile (10–75%) than leafy greens [31,33,35,74], most likely due to soil residues [74,75].
4.2. Ribotypes of C. difficile
In our study, ribotypes 078 and 126 were detected on spinach leaves while RT614, 003, 087, 050, 511 and 014/0 were found in the soil. The majority of these ribotypes have been previously linked to animal, food and human sources.
Ribotype 078 is frequently reported in farm animals such as cattle [76,77,78], poultry [79,80] and pigs [81,82,83], with no distinguishable differences between human and porcine RT078 isolates [84,85]. It has also been found in food products derived from animals purchased at retail such as ground pork, turkey [86] and beef [87]. In vegetables, it was detected in ginger and carrots from Canada [33] and in potatoes from Ireland and Italy [28]. In humans, hypervirulent C. difficile RT078 accounted for 17% of ribotyped isolates in 2021 in Ireland [88], with similar data reported for Germany (16.7%) by Rabold et al. [89]. Interestingly, the RT078 strain isolated in our study was identified as similar to C. difficile strain M120 by WGS, a strain that has been associated with clinical infection in the UK and Ireland [19], as well as in calves in Canada [90] and the pig farm environment in Spain [91].
Ribotype 126 has been described in cattle [76,92,93] and pigs [94,95]. It may not be a coincidence that both ribotypes RT078 and RT126, which are frequently reported in cattle, were detected in the spinach isolates from Field 1 that was located adjacent to fields with bovine animals. Apart from farm animals, Agnoletti et al. [96] and Troiano et al. [97] isolated RT126 from shellfish. Tkalec et al. [28] confirmed the presence of ribotype 126 in potatoes from France, Spain, Austria and Romania, and Primavilla et al. [34] found this ribotype in lettuce served in Italian hospitals. In Spain, RT126 was one the most common ribotypes among clinical isolates [98], while in Portugal, it accounted for 3.8% of the CDI cases [99]. Recently, Azimirad et al. [100] identified RT126 as the predominant ribotype (11.2%) in hospital patients in Tehran. Hypervirulent RT126 was observed in clinical isolates in Taiwan by Hung et al. [101], in Kuwait by Jamal and Rotimi [102] and in Australia by Knight et al. [82]. The ribotype 126 detected in spinach leaves in our study was similar to C. difficile strain DSM 29020, which has been reported in hospital patients in Korea and China [103,104].
Of our soil isolates, ribotype 003 was previously reported in poultry faeces (1%) [105] and meat (28.6%) [106], calves [76] and horses (5.6%) [107]. Indeed, the close proximity of the field where it was isolated to a stud farm might highlight the potential for its zoonotic transfer. Regarding its link to humans, in Germany, RT003 accounted for 11.1% of CDI cases in 2012–2013 [89], and strain W0023a, which is similar to our isolate by WGS, has been detected in clinical isolates from Korea and the USA [103,108].
RT087, the ribotype of another of our soil isolates, was identified in poultry meat by de Boer et al. [106] and Tkalec et al. [109] (14.3 and 20%, respectively). The latter authors also reported that ribotype 087 was more common in clinical isolates than RT078 in a Slovenian study.
To the best of our knowledge, this is the first time RT050 has been reported in soil samples, although it has been reported in pigs (12%) [110] and was recently associated with hospitalized adult patients in Mexico [111] and in a CDI outbreak in the intensive care unit of the Amsterdam University Medical Centre [112]. Furthermore, both RT087 and RT050 were similar to C. difficile CFSAN096664, which was previously isolated from hospital patients in the USA [113].
Ribotype 014, previously described in retail ground beef (8.3%) [114], was found in soil in Field 3 that was located next to a dairy farm. A variant of this ribotype, RT014/020, was the predominant ribotype among soil isolates obtained from domestic gardens in Australia by Shivaperumal et al. [26]. In humans, a study from Germany noted RT014/0 as the most prevalent ribotype among CDI patients (22.2%) [89], and our WGS analysis suggested that this isolate was similar to C. difficile DSM 105001, which has been reported before in US hospital patients [115].
Isolates of RT614 have not been previously reported in soil but have been found in clinical samples in Portugal [116]. This isolate was similar to DSM 27639, which has been reported in German hospital patients [117].
Our remaining soil isolate was ribotype 511 and has not been associated with animal, food or human sources. However, due to the ribotype’s lack of toxin genes, it may be carried asymptomatically [118] or be associated with hospital-acquired diarrhoea [119]. WGS analysis suggested that RT511 was most closely related to strain C. difficile W0003a, previously observed in clinical isolates in Korea [103].
4.3. Toxigenicity of the C. difficile Isolates
The C. difficile population consists of six distinct phylogenetic clades labelled 1, 2, 3, 4, 5, and C-I [120,121]. Most genotypes within clades 1–3 produce both toxins A and B and, in addition to these, strains from clades 2, 3, and 5 have genes encoding the binary toxin, CDT. Clade 5 includes ribotype 078, which is typically associated with community-acquired cases and has a higher rate of complications and mortality in humans [3,110,122]. In the present study, spinach isolates (isolates 2 and 3) both belonged to this highly toxigenic clade 5 (ST11) and contained the binary toxin CDT, characteristic of hypervirulent strains [10], apart from toxins A and B. Interestingly, while conventional PCR yielded a negative result for the tcdA gene in the spinach isolates, this gene was detected using WGS. Thus, SNP analysis was carried out to search for possible mutations in the primer region used to amplify tcdA that could have affected the PCR analysis and might explain the negative result. However, no SNPs were detected in the primer or promoter region. The use of a positive tcdA control (C. difficile RT078) and a negative control belonging to C. sporogenes in the conventional PCR analysis ruled out issues with the PCR reaction (data not shown), and thus, further research is required to explain this apparent anomaly.
All soil isolates (isolates 1, 4–8) belonged to clade 1 and presented the toxigenic sequence types and ribotypes ST19/RT614, ST12/RT003, ST46/RT087, ST16/RT050, and ST49/RT014/0 and one non-toxigenic ST79/RT511 (isolate 7). Clade 1 comprises more than 200 toxigenic and non-toxigenic sequence types, with most toxigenic strains encoding genes for toxin A and B production, as we observed in the majority of the soil isolates. Moreover, a few STs (ST2, ST8 and ST17) from clade 1 have been associated with CDI cases worldwide [82].
Pangenome and phylogenetic analysis showed the existence of two clusters among our samples. The first cluster included the spinach samples belonging to clade 5 and the second cluster comprised the soil samples that belonged to clade 1. Thus, the soil samples were closely related to each other but were phylogenetically distinct from the spinach isolates (who were, in turn, closely related). However, due to the limited sample size of this study, further studies would need to investigate this potential relationship considering a larger number of isolates.
4.4. Antibiotic Susceptibility of the C. difficile Isolates
An interesting finding in our study was the resistance differences observed in antibiotics tested phenotypically and genotypically, supporting previous similar observations in C. difficile by Muñoz et al. [123].
Phenotypic resistance to erythromycin (RT126) was observed in a spinach isolate, while soil isolates were resistant to erythromycin, vancomycin and rifampicin (RT003 and RT050), moxifloxacin and rifampicin (RT087), and rifampicin (RT614 and RT511) and clindamycin (RT014/0). While phenotypic analysis was carried out for a limited range of antibiotics (erythromycin, metronidazole, moxifloxacin, clindamycin, vancomycin and rifampicin), genotypic analysis suggested the existence of more resistance determinants. In spinach isolates (RT078 and RT126), resistance genes against tetracycline (tet(M) and tet(40)), aminoglycosides (APH(3′) and ANT(6)), beta-lactam (CDD-1 and CDD-2), disinfecting agents and antiseptics (qacG), nucleoside (SAT-4) and clindamycin antibiotics were detected, while in soil isolates (RT614, 003, 087, 050, 511 and 014/0), resistance genes against vancomycin (vanR and vanXY), fluoroquinolones (cdeA), beta-lactam, disinfecting agents and antiseptics, erythromycin and clindamycin antibiotics were detected. Interestingly, with the exception of erythromycin resistance in isolates 3, 4 and 6, in clindamycin resistance in isolate 8 and vancomycin resistance in isolates 4 and 6, there was little agreement between the phenotypic and genotypic assessment of AMR.
In spinach isolates, similar susceptibilities to those observed in this study in RT078 against all antibiotics tested were described previously in isolates from humans by Álvarez-Pérez et al. [98], rabbits by Drigo et al. [124] and pigs by Zhang et al. [83]. Freeman et al. [14] observed vancomycin resistance in RT078 isolates while we observed susceptibility, although the lack of vancomycin resistance genes we reported in this ribotype would explain our results. In addition, Zhang et al. [83] found resistance to tetracycline in RT078 isolates from pigs in China.
In RT126, phenotypic resistance to erythromycin had been previously reported by Álvarez-Pérez et al. [98] and can be explained by the existence of a mutation in the target site of C. difficile 23S rRNA that confers resistance against the antibiotic. Furthermore, the gene SAT-4, which encodes nucleoside antibiotic resistance and was identified in RT126 in our study, was detected in Campylobacter coli and Enterococcus faecium by other authors [125,126].
Apart from this, both spinach isolates (RT078 and RT126) contained components of the replicon repUS43_1_CDS12738(DOp1), which has been linked with resistance against tetracycline, methicillin and penicillin in methicillin-resistant Staphylococcus aureus (MRSA) LA-MRSA SA0385 [127].
In all soil isolates (RT614, RT003, RT087, RT050, RT511, RT014/0), vancomycin resistance genes (vanXY and vanR) were detected, even though phenotypic resistance was only observed in isolates of RT003 and RT050, contrary to the susceptibility reported by Kecerova et al. [107] in RT003. These results support the findings of Suzuki et al. [128] in C. difficile RT027 strains that carried vanRG and vanG genes but were phenotypically susceptible to vancomycin. Therefore, the presence of vancomycin resistance genes in C. difficile does not always result in their expression in vitro.
Phenotypic resistance to clindamycin in RT014/0, previously reported in human and pig isolates in Australia [129,130] and to erythromycin in RT003 and RT050, is probably due to the mutation detected in the antibiotic target site of C. difficile 23S rRNA, which confers resistance to both antibiotics. The resistance of C. difficile strains to erythromycin and clindamycin has been widely reported in human isolates and treatment with clindamycin is a risk factor for developing CDI [18,131].
Additionally, the in vitro resistance to moxifloxacin in RT087, in contrast to the results reported by Freeman et al. [14] for this ribotype, could be explained by the presence of the cdeA gene. This gene confers resistance against fluoroquinolones in E. coli and has been previously detected in C. difficile [132].
To the best of our knowledge, no previous authors have described antimicrobial resistance patterns for RT614 and RT511, possibly due to a lower frequency of reports in humans, even though they both showed phenotypic resistance to rifampicin in our study and have been associated with an increased severity of CDI [133].
The CDD-1 and CDD-2 genes, detected in all of the isolates analysed in this study, confer a high resistance against a broad range of beta-lactams [134]. While beta-lactam antibiotics were not tested by E-test in this study, these enzymes previously reported in C. difficile allow the bacteria to have intrinsic resistance to antibiotics such as penicillin, monobactams and cephalosporins [135].
Interestingly, the qacG gene present in every isolate was identified previously in staphylococci isolated from the food industry, conferring resistance to benzalkonium chloride that is a frequently used quaternary ammonium disinfectant [136]. Resistance to quaternary ammonium compounds is plasmid-borne in Gram-positive bacteria, linked to the Small Multi-drug Resistance (SMR) family transporters (qacC/D and qacEΔ1, qacG, qacH and qacJ) [137]. While the qacG gene has not been reported in C. difficile before, in 2013, it was detected in a S. aureus MRSA strain isolated from pork [138]. However, Seier-Petersen et al. [139] described no reduced susceptibility in vitro to biocides in S. aureus MRSA strains where the qacG gene was present. More recently, the gene was found in the Staphylococcus species in Turkey [140] and carbapenem-resistant A. baumannii in China [141].
5. Conclusions
It was concluded that the prevalence of C. difficile was 10% in soil and 3.3% in spinach, both of which were lower than the results reported in other similar studies. Both spinach isolates (toxigenic RT078 and RT126) carried virulence genes (toxins A, B and CDT) and RT126 was phenotypically resistant to erythromycin. Soil isolates included toxigenic ribotypes (003, 014/0, 050, 087 and RT614) and a novel non-toxigenic ribotype (RT511). RT003 and RT050 were resistant to erythromycin, vancomycin and rifampicin, RT087 to moxifloxacin and rifampicin, and RT014/0 to clindamycin. WGS suggested there were inconsistencies between AMR phenotypes and genotypes. Moreover, the soil isolates were closely related but genetically distinct from the spinach strains.
Author Contributions
P.M.: Conceptualization, investigation, formal analysis, writing—original draft, writing—review and editing. P.W.: Methodology, supervision, writing—review and editing. C.B.: Methodology, supervision, writing—review and editing. D.E.: Formal analysis, visualization. D.B.: Conceptualization, funding acquisition, experimental design, methodology, project administration, supervision, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the Food Institutional Research Measure (FIRM) administered by the Department for Agriculture, Food and the Marine (DAFM) (Grant number 17F206). Pilar Marcos was supported by the Teagasc Walsh Scholarship Scheme (number 2018210).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Nucleotide sequence data reported are available in the GenBank database under the accession number PRJNA884639.
Acknowledgments
The pictographic symbols used are the property of the Government of Aragón and have been created by Sergio Palao for ARASAAC (http://www.arasaac.org; accessed on 27 September 2022) that distributes them under Creative Commons License BY-NC-SA. The authors acknowledge Aoife Doyle for doing the PCR-ribotyping analysis in St James's Hospital (Dublin, Ireland).
Conflicts of Interest
The authors declare that they have no known competing financial interest or personal relationship that could have appeared to influence the work reported in this paper.
References
- Enoch, D.A.; Aliyu, S.H. Is Clostridium difficile infection still a problem for hospitals? Can. Med. Assoc. J. 2012, 184, 17–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czepiel, J.; Dróżdż, M.; Pituch, H.; Kuijper, E.J.; Perucki, W.; Mielimonka, A.; Goldman, S.; Wultańska, D.; Garlicki, A.; Biesiada, G. Clostridium difficile infection: Review. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1211–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.M.; Kuijper, E.J.; Wilcox, M.H. Clostridium difficile: A European perspective. J. Infect. 2013, 66, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Tunis, M.C.; Frenette, C.; Katz, K.; Amaratunga, K.; Rhodenizer Rose, S.; House, A.; Quach, C. Epidemiology of Clostridioides difficile infection in Canada: A six-year review to support vaccine decision-making. CCDR 2019, 45, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Donta, S.T.; Sullivan, N.; Wilkins, T.D. Differential effects of Clostridium difficile toxins on tissue-cultured cells. J. Clin. Microbiol. 1982, 15, 1157–1158. [Google Scholar] [CrossRef] [Green Version]
- Feltis, B.A.; Wiesner, S.M.; Kim, A.S.; Erlandsen, S.L.; Lyerly, D.L.; Wilkins, T.D.; Wells, C.L. Clostridium difficile toxins A and B can alter epithelial permeability and promote bacterial paracellular migration through HT-29 enterocytes. Shock 2000, 14, 629–634. [Google Scholar] [CrossRef]
- Riggs, M.M.; Sethi, A.K.; Zabarsky, T.F.; Eckstein, E.C.; Jump, R.L.P.; Donskey, C.J. Asymptomatic carriers are a potential source for transmission of epidemic and nonepidemic Clostridium difficile strains among longterm care facility residents. Clin. Infect. Dis. 2007, 45, 992–998. [Google Scholar] [CrossRef] [Green Version]
- Rupnik, M.; Janezic, S. An update on Clostridium difficile toxinotyping. J. Clin. Microbiol. 2016, 54, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Voth, D.; Ballard, J.D. Clostridium difficile Toxins: Mechanism of Action and Role in Disease. Clin. Microbiol. Rev. 2005, 2, 247–263. [Google Scholar] [CrossRef] [Green Version]
- Kuehne, S.A.; Collery, M.M.; Kelly, M.L.; Cartman, S.T.; Cockayne, A.; Minton, N.P. Importance of toxin A, toxin B, and CDT in virulence of an epidemic Clostridium difficile strain. J. Infect. Dis. 2014, 209, 83–86. [Google Scholar] [CrossRef]
- Gerding, D.N.; Johnson, S.; Rupnik, M.; Aktories, K. Clostridium difficile binary toxin CDT: Mechanism, epidemiology, and potential clinical importance. Gut Microbes 2014, 5, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Stiles, B.G.; Wigelsworth, D.J.; Popoff, M.R.; Barth, H. Clostridial binary toxins: Iota and c2 family portraits. Front. Cell. Infect. Microbiol. 2011, 1, 11. [Google Scholar] [CrossRef] [Green Version]
- Dingle, T.C.; MacCannell, D.R. Chapter 9—Molecular Strain Typing and Characterisation of Toxigenic Clostridium difficile. In Methods in Microbiology; Sails, A., Tang, Y.-W., Eds.; Academic Press: San Diego, CA, USA, 2015; Volume 42, pp. 329–357. [Google Scholar]
- Freeman, J.; Vernon, J.; Morris, K.; Nicholson, S.; Todhunter, S.; Longshaw, C.; Wilcox, M.H. Pan-European Longitudinal Surveillance of Antibiotic Resistance among Prevalent Clostridium difficile Ribotypes’ Study Group. Clin. Microbiol. Infect. 2015, 21, 248.e9–248.e16. [Google Scholar] [CrossRef] [Green Version]
- Hampikyan, H.; Bingol, E.B.; Muratoglu, K.; Akkaya, E.; Cetin, O.; Colak, H. The prevalence of Clostridium difficile in cattle and sheep carcasses and the antibiotic susceptibility of isolates. Meat Sci. 2018, 139, 120–124. [Google Scholar] [CrossRef]
- Herbert, R.; Hatcher, J.; Jauneikaite, E.; Gharbi, M.; d’Arc, S.; Obaray, N.; Rickards, T.; Rebec, M.; Blandy, O.; Hope, R.; et al. Two-year analysis of Clostridium difficile ribotypes associated with increased severity. J. Hosp. Infect. 2019, 103, 388–394. [Google Scholar] [CrossRef] [Green Version]
- Couturier, J.; Davies, K.; Gateau, C.; Barbut, F. Ribotypes and New Virulent Strains Across Europe. Adv. Exp. Med. Biol. 2018, 1050, 45–58. [Google Scholar]
- Peng, Z.; Jin, D.; Kim, H.B.; Stratton, C.W.; Wu, B.; Tang, Y.-W.; Sun, X. Update on Antimicrobial Resistance in Clostridium difficile: Resistance Mechanisms and Antimicrobial Susceptibility Testing. J. Clin. Microbiol. 2017, 55, 1998–2008. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Miyajima, F.; Roberts, P.; Ellison, L.; Pickard, D.J.; Martin, M.J.; Connor, T.R.; Harris, S.R.; Fairley, D.; Bamford, B.; et al. Emergence and global spread of epidemic healthcare-associated Clostridium difficile. Nat. Genet. 2013, 45, 109–113. [Google Scholar] [CrossRef]
- Owens, R.C.; Donskey, C.J.; Gaynes, R.P.; Loo, V.G.; Muto, C.A. Antimicrobial-associated risk factors for Clostridium difficile infection. Clin. Infect. Dis. 2008, 46, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spigaglia, P.; Barbanti, F.; Mastrantonio, P. European Study Group on Clostridium difficile (ESGCD) Multidrug resistance in European Clostridium difficile clinical isolates. J. Antimicrob. Chemother. 2011, 66, 2227–2234. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.A.; Stahl, T.J. Persistent and Recurrent Clostridium difficile Colitis. Clin. Colon Rectal Surg. 2015, 28, 65–69. [Google Scholar] [CrossRef]
- Haagsma, J. Pathogenic anaerobic bacteria and the environment. Rev. Sci. Tech. (Int. Off. Epizoot.) 1991, 10, 749–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos, P.; Whyte, P.; Rogers, T.; McElroy, M.; Fanning, S.; Frias, J.; Bolton, D. The prevalence of Clostridioides difficile on farms, in abattoirs and in retail foods in Ireland. Food Microbiol. 2021, 98, 103781. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, C.; Bouchafa, L.; Soumillion, K.; Ngyuvula, E.; Taminiau, B.; Van Broeck, J.; Delmée, M.; Daube, G. Seasonality of Clostridium difficile in the natural environment. Transbound. Emerg. Dis. 2019, 66, 2440–2449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivaperumal, N.; Chang, B.J.; Riley, T.V. High Prevalence of Clostridium difficile in Home Gardens in Western Australia. Appl. Environ. Microbiol. J. 2020, 87, e01572-20. [Google Scholar] [CrossRef]
- Bazaid, F. Distribution and Sources of Clostridium difficile Present in Water Sources. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2013. [Google Scholar]
- Tkalec, V.; Janezica, S.; Skoka, B.; Simonica, T.; Mesarica, S.; Vrabica, T.; Rupnik, M. High Clostridium difficile contamination rates of domestic and imported potatoes compared to some other vegetables in Slovenia. Food Microbiol. 2019, 78, 194–200. [Google Scholar] [CrossRef]
- Warriner, K.; Xu, C.; Habash, M.; Sultan, S.; Weese, S.J. Dissemination of Clostridium difficile in food and the environment: Significant sources of, C. difficile community acquired infection? J. Appl. Microbiol. 2016, 122, 542–555. [Google Scholar] [CrossRef]
- Xu, C.; Weese, J.S.; Flemming, C.; Odumeru, J.; Warriner, K. Fate of Clostridium difficile during wastewater treatment and incidence in Southern Ontario watersheds. J. Appl. Microbiol. 2014, 117, 891–904. [Google Scholar] [CrossRef]
- Eckert, C.; Burghoffer, B.; Barbut, F. Contamination of ready-to-eat raw vegetables with Clostridium difficile, France. J. Med. Microbiol. 2013, 62, 1435–1438. [Google Scholar] [CrossRef]
- Lim, S.C.; Foster, N.F.; Elliott, B.; Riley, T.V. High prevalence of Clostridium difficile on retail root vegetables, Western Australia. J. Appl. Microbiol. 2017, 124, 585–590. [Google Scholar] [CrossRef]
- Metcalf, D.; Costa, M.C.; Dew, W.M.; Weese, J.S. Clostridium difficile in vegetables, Canada. Lett. Appl. Microbiol. 2010, 51, 600–602. [Google Scholar] [CrossRef]
- Primavilla, S.; Farneti, S.; Petruzzelli, A.; Drigo, I.; Scuota, S. Contamination of hospital food with Clostridium difficile in Central Italy. Anaerobe 2019, 55, 8–10. [Google Scholar] [CrossRef]
- Bakri, M.M.; Brown, D.J.; Butcher, J.P.; Sutherland, A.D. Clostridium difficile in ready-to-eat salads, Scotland. Emerg. Infect. Dis. 2009, 15, 817–818. [Google Scholar] [CrossRef]
- Yamoudy, M.; Mirlohi, M.; Isfahani, B.N.; Jalali, M.; Esfandiari, Z.; Hosseini, N.S. Isolation of toxigenic Clostridium difficile from ready-to-eat salads by multiplex polymerase chain reaction in Isfahan, Iran. Adv. Biomed. Res. 2015, 4, 87. [Google Scholar]
- Lim, S.C.; Knight, D.R.; Moono, P.; Foster, N.F.; Riley, T.V. Clostridium difficile in soil conditioners, mulches and garden mixes with evidence of a clonal relationship with historical food and clinical isolates. Environ. Microbiol. Rep. 2020, 12, 672–680. [Google Scholar] [CrossRef]
- Le Maréchal, C.; Druilhe, C.; Repérant, E.; Boscher, E.; Rouxel, S.; Le Roux, S.; Poëzévara, T.; Ziebal, C.; Houdayer, C.; Nagard, B.; et al. Evaluation of the occurrence of sporulating and nonsporulating pathogenic bacteria in manure and in digestate of five agricultural biogas plants. Microbiol. Open 2019, 8, e872. [Google Scholar] [CrossRef]
- Lemee, L.; Dhalluin, A.; Testelin, S.; Mattrat, M.A.; Maillard, K.; Lemeland, J.F.; Pons, J.L. Multiplex PCR targeting tpi (triose phosphate isomerase), tcdA (toxin A), and tcdB (toxin B) genes for toxigenic culture of Clostridium difficile. J. Clin. Microbiol. 2004, 42, 5710–5714. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, G.; Tang-Feldman, Y.J.; Schaumann, R.; Henderson, J.P.; Rodloff, A.C.; Silva, J.; Cohen, S.H. Antecedent use of fluoroquinolones is associated with resistance to moxifloxacin in Clostridium difficile. Clin. Microbiol. Infect. 2003, 9, 526–530. [Google Scholar] [CrossRef] [Green Version]
- Oldfield, E.C., IV; Oldfield, E.C., III; Johnson, D.A. Clinical update for the diagnosis and treatment of Clostridium difficile infection. World J. Gastrointest. Pharmacol. Ther. 2014, 5, 1–26. [Google Scholar]
- Johnson, S.; Samore, M.H.; Farrow, K.A.; Killgore, G.E.; Tenover, F.C.; Lyras, D.; Rood, J.I.; DeGirolami, P.; Baltch, A.L.; Rafferty, M.E.; et al. Epidemics of diarrhea caused by a clindamycin-resistant strain of Clostridium difficile in four hospitals. N. Engl. J. Med. 1999, 341, 1645–1651. [Google Scholar] [CrossRef]
- Surawicz, C.M.; Brandt, L.J.; Binion, D.G.; Ananthakrishnan, A.N.; Curry, S.R.; Gilligan, P.H.; McFarland, L.V.; Mellow, M.; Zuckerbraun, B.S. Guidelines for diagnosis, treatment, and prevention of Clostridium difficile infections. Am. J. Gastroenterol. 2013, 108, 478–498. [Google Scholar] [CrossRef] [PubMed]
- Vardakas, K.Z.; Polyzos, K.A.; Patouni, K.; Rafailidis, P.I.; Samonis, G.; Falagas, M.E. Treatment failure and recurrence of Clostridium difficile infection following treatment with vancomycin or metronidazole: A systematic review of the evidence. Int. J. Antimicrob. Agents 2012, 40, 1–8. [Google Scholar] [CrossRef] [PubMed]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of MICs and Zone Diameters Version 11.0, Valid from 2021-01-01. 2021. Available online: http://www.eucast.org (accessed on 23 November 2021).
- European Centre for Disease Prevention and Control (ECDC). Laboratory Procedures for Diagnosis and Typing of Human Clostridium difficile Infection. Technical Report. 2018. Available online: www.ecdc.europa.eu (accessed on 23 November 2021).
- Bidet, P.; Barbut, F.; Lalande, V.; Burghoffer, B.; Petit, J.C. Development of a new PCR-ribotyping method for Clostridium difficile based on ribosomal RNA gene sequencing. FEMS Microbiol. Lett. 1999, 175, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data [Online]. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 20 May 2022).
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [Green Version]
- Clausen, P.T.L.C.; Aarestrup, F.M.; Lund, O. Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinform. 2018, 19, 307. [Google Scholar] [CrossRef]
- Hasman, H.; Saputra, D.; Sicheritz-Ponten, T.; Lund, O.; Svendsen, C.A.; Frimodt-Møller, N.; Aarestrup, F.M. Rapid whole-genome sequencing for detection and characterization of microorganisms directly from clinical samples. J. Clin. Microbiol. 2014, 52, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Ponten, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of methods for genomic taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2020, 72, 2764–2768. [Google Scholar] [CrossRef] [Green Version]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Snippy: Fast Bacterial Variant Calling from NGS Reads. 2015. Available online: https://github.com/tseemann/snippy (accessed on 1 June 2022).
- Eren, A.M.; Kiefl, E.; Shaiber, A.; Veseli, I.; Miller, S.E.; Schechter, M.S.; Fink, I.; Pan, J.N.; Yousef, M.; Fogarty, E.C.; et al. Community-led, integrated, reproducible multi-omics with anvi’o. Nat. Microbiol. 2021, 6, 3–6. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Benedict, M.N.; Henriksen, J.R.; Metcalf, W.W.; Whitaker, R.J.; Price, N.D. ITEP: An integrated toolkit for exploration of microbial pan-genomes. BMC Genom. 2014, 15, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Dongen, S.; Abreu-Goodger, C. Using MCL to extract clusters from networks. Methods Mol. Biol. 2012, 804, 281–295. [Google Scholar] [PubMed]
- Leekitcharoenphon, P.; Nielsen, E.M.; Kaas, R.S.; Lund, O.; Aarestrup, F.M. Evaluation of whole genome sequencing for outbreak detection of Salmonella enterica. PLoS ONE 2014, 9, e87991. [Google Scholar] [CrossRef] [PubMed]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]
- Olaitan, A.; Dureja, C.; Youngblom, M.; Topf, M.; Shen, W.-J.; Gonzales-Luna, A.; Deshpande, A.; Hevener, K.; Freeman, J.; Wilcox, M.; et al. Decoding a cryptic mechanism of metronidazole resistance among globally disseminated fluoroquinolone-resistant Clostridioides difficile. bioRxiv 2022. [Google Scholar] [CrossRef]
- Dang, U.T.; Zamora, I.; Hevener, K.E.; Adhikari, S.; Wu, X.; Hurdle, J.G. Rifamycin Resistance in Clostridium difficile Is Generally Associated with a Low Fitness Burden. Antimicrob. Agents Chemother. 2016, 60, 5604–5607. [Google Scholar] [CrossRef] [Green Version]
- Al Saif, N.; Brazier, J.S. The distribution of Clostridium difficile in the environment of South Wales. J. Med. Microbiol. 1996, 45, 133–137. [Google Scholar] [CrossRef] [Green Version]
- Simango, C. Prevalence of Clostridium difficile in the environment in a rural community in Zimbabwe. R. Soc. Trop. Med. Hyg. 2006, 100, 1146–1150. [Google Scholar] [CrossRef]
- Blasi, F.; Lovito, C.; Albini, E.; Bano, L.; Dalmonte, G.; Drigo, I.; Maresca, C.; Massacci, F.R.; Orsini, S.; Primavilla, S.; et al. Clostridioides difficile in Calves in Central Italy: Prevalence, Molecular Typing, Antimicrobial Susceptibility and Association with Antibiotic Administration. Animals 2021, 11, 515. [Google Scholar] [CrossRef]
- Hammitt, M.C.; Bueschel, D.M.; Keel, M.K.; Glock, R.D.; Cuneo, P.; DeYoung, D.W.; Reggiardo, C.; Trinh, H.T.; Songer, J.G. A possible role for Clostridium difficile in the etiology of calf enteritis. Vet. Microbiol. 2008, 127, 343–352. [Google Scholar] [CrossRef]
- Magistrali, C.F.; Maresca, C.; Cucco, L.; Bano, L.; Drigo, I.; Filippini, G.; Dettori, A.; Broccatelli, S.; Pezzotti, G. Prevalence and risk factors associated with Clostridium difficile shedding in veal calves in Italy. Anaerobe 2015, 33, 42–47. [Google Scholar] [CrossRef]
- Harvey, R.; Norman, K.; Andrews, K.; Hume, M.; Scanlan, C.; Callaway, T. Clostridium difficile in poultry and poultry meat. Foodborne Pathog. Dis. 2011, 8, 1321–1323. [Google Scholar] [CrossRef]
- Hussain, I.; Borah, P.; Sharma, R.K.; Rajkhowa, S.; Rupnik, M.; Saikia, D. P Molecular characteristics of Clostridium difficile isolates from human and animals in the North Eastern region of India. Mol. Cell. Probes 2016, 30, 306–311. [Google Scholar] [CrossRef]
- Álvarez-Pérez, S.; Blanco, J.L.; Peláez, T.; Astorga, R.J.; Harmanus, C.; Kuijper, E.; García, M.E. High prevalence of the epidemic Clostridium difficile PCR ribotype 078 in Iberian free-range pigs. Res. Vet. Sci. 2013, 95, 358–361. [Google Scholar] [CrossRef]
- Knight, D.R.; Elliott, B.; Chang, B.J.; Perkins, T.T.; Riley, T.V. Diversity and evolution in the genome of Clostridium difficile. Clin. Microbiol. Rev. 2015, 28, 721–741. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.-J.; Yang, L.; Gu, X.-X.; Chen, P.-X.; Fu, J.-L.; Jiang, H.-X. The first isolation of Clostridium difficile RT078/ST11 from pigs in China. PLoS ONE 2019, 14, e0212965. [Google Scholar]
- Debast, S.B.; van Leengoed, L.A.; Goorhuis, A.; Harmanus, C.; Kuijper, E.J.; Bergwerff, A.A. Clostridium difficile PCR ribotype 078 toxinotype, V. found in diarrhoeal pigs identical to isolates from affected humans. Environ. Microbiol. 2009, 11, 505–511. [Google Scholar] [CrossRef]
- Keessen, E.C.; Harmanus, C.; Dohmen, W.; Kuijper, E.J.; Lipman, L.J. Clostridium difficile infection associated with pig farms. Emerg. Infect. Dis. 2013, 19, 1032–1034. [Google Scholar] [CrossRef]
- Songer, J.G.; Trinh, H.T.; Killgore, G.E.; Thompson, A.D.; McDonald, L.C.; Limbago, B.M. Clostridium difficile in Retail Meat Products, USA, 2007. Emerg. Infect. Dis. 2009, 15, 819–821. [Google Scholar] [CrossRef]
- Weese, J.; Avery, B.P.; Rousseau, J.; Reid-Smith, J. Detection and Enumeration of Clostridium difficile Spores in Retail Beef and Pork. Appl. Environ. Microbiol. 2009, 15, 5009–5011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Health Service Executive (HSE)—Health Protection Surveillance Centre (HPSC). Enhanced Surveillance of Clostridioides (Clostridium) difficile Infection in Ireland: Q2 2021 National Report; HSE-HPSC: Dublin, Ireland, 2021. [Google Scholar]
- Rabold, D.; Espelage, W.; Abu Sin, M.; Eckmanns, T.; Schneeberg, A.; Neubauer, H.; Möbius, N.; Hille, K.; Wieler, L.H.; Seyboldt, C.; et al. The zoonotic potential of Clostridium difficile from small companion animals and their owners. PLoS ONE 2018, 13, e0193411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Davenport, K.W.; Vuyisich, G.; Kunde, Y.A.; Johnson, S.L.; Chain, P.S.G.; Dichosa, A.E.K.; Rodriguez-Palacios, A. Complete Genome Sequences of Historic Clostridioides difficile Food-Dwelling Ribotype 078 Strains in Canada Identical to That of the Historic Human Clinical Strain M120 in the United Kingdom. Microbiol. Resour. Announc. 2018, 7, e00853-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín-Burriel, I.; Andrés-Lasheras, S.; Harders, F.; Mainar-Jaime, R.C.; Ranera, B.; Zaragoza, P.; Falceto, V.; Bolea, Y.; Kuijper, E.; Bolea, R.; et al. Molecular analysis of three Clostridium difficile strain genomes isolated from pig farm-related samples. Anaerobe 2017, 48, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Knight, D.R.; Putsathit, P.; Elliott, B.; Riley, T.V. Contamination of Australian newborn calf carcasses at slaughter with Clostridium difficile. Clin. Microbiol. Infect. 2016, 22, 266.e1–266.e7. [Google Scholar] [CrossRef] [Green Version]
- Masarikova, M.; Simkova, I.; Plesko, M.; Eretova, V.; Krutova, M.; Cizek, A. The Colonisation of Calves in Czech Large-Scale Dairy Farms by Clonally-Related Clostridioides difficile of the Sequence Type 11 Represented by Ribotypes 033 and 126. Microorganisms 2020, 8, 901. [Google Scholar] [CrossRef]
- Avberšek, J.; Janezic, S.; Pate, M.; Rupnik, M.; Zidaric, V.; Logar, K.; Vengust, M.; Zemljic, M.; Pirs, T.; Ocepek, M. Diversity of Clostridium difficile in pigs and other animals in Slovenia. Anaerobe 2009, 15, 252–255. [Google Scholar] [CrossRef]
- Andrés-Lasheras, S.; Bolea, R.; Mainar-Jaime, R.C.; Kuijper, E.; Sevilla, E.; Martín-Burriel, I.; Chirino-Trejo, M. Presence of Clostridium difficile in pig faecal samples and wild animal species associated with pig farms. J. Appl. Microbiol. 2017, 122, 462–472. [Google Scholar] [CrossRef]
- Agnoletti, F.; Arcangelia, G.; Barbantib, F.; Barcoa, L.; Brunettaa, R.; Cocchia, M.; Conederaa, G.; D’Estea, L.; Drigoa, I.; Spigagliab, P.; et al. Survey, characterization and antimicrobial susceptibility of Clostridium difficile from marine bivalve shellfish of North Adriatic Sea. Int. J. Food Microbiol. 2019, 298, 74–80. [Google Scholar] [CrossRef]
- Troiano, T.; Harmanus, C.; Sanders, I.; Pasquale, V.; Dumontet, S.; Capuano, F.; Romano, V.; Kuijper, E. J Toxigenic Clostridium difficile PCR ribotypes in ediblemarine bivalve molluscs in Italy. Int. J. Food Microbiol. 2015, 208, 30–34. [Google Scholar] [CrossRef]
- Álvarez-Pérez, S.; Blanco, J.L.; Harmanus, C.; Kuijper, E.; García, M.E. Subtyping and antimicrobial susceptibility of Clostridium difficile PCR ribotype 078/126 isolates of human and animal origin. Vet. Microbiol. 2017, 199, 15–22. [Google Scholar] [CrossRef]
- Santos, A.; Isidro, J.; Silva, C.; Boaventura, L.; Diogo, J.; Faustino, A.; Toscano, C.; Oleastro, M. Molecular and epidemiologic study of Clostridium difficile reveals unusual heterogeneity in clinical strains circulating in different regions in Portugal. Clin. Microbiol. Infect. 2016, 22, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Azimirad, M.; Krutova, M.; Yadegar, A.; Shahrokh, S.; Olfatifar, M.; Aghdaei, H.A.; Fawley, W.N.; Wilcox, M.H.; Zali, M.R. Clostridioides difficile ribotypes 001 and 126 were predominant in Tehran healthcare settings from 2004 to 2018: A 14-year-long cross-sectional study. Emerg. Microbes Infect. 2020, 9, 1432–1443. [Google Scholar] [CrossRef]
- Hung, Y.P.; Lin, H.J.; Tsai, B.Y.; Liu, H.C.; Liu, H.C.; Lee, J.C.; Wu, Y.H.; Wilcox, M.H.; Fawley, W.N.; Hsueh, P.R.; et al. Clostridium difficile ribotype 126 in southern Taiwan: A cluster of three symptomatic cases. Anaerobe 2014, 30, 188–192. [Google Scholar] [CrossRef]
- Jamal, W.Y.; Rotimi, V.O. Surveillance of antibiotic resistance among hospital- and community-acquired toxigenic Clostridium difficile isolates over 5-year period in Kuwait. PLoS ONE 2016, 11, e0161411. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.W.; Lee, S.H.; Kim, U.J.; Jang, H.C.; Choi, H.J.; Choy, H.E.; Kang, S.J.; Roh, S.W. Genomic characterization of nine Clostridioides difficile strains isolated from Korean patients with Clostridioides difficile infection. Gut Pathog. 2021, 13, 55. [Google Scholar] [CrossRef]
- Su, T.; Chen, W.; Wang, D.; Cui, Y.; Ni, Q.; Jiang, C.; Dong, D.; Peng, Y. Complete Genome Sequencing and Comparative Phenotypic Analysis Reveal the Discrepancy Between Clostridioides difficile ST81 and ST37 Isolates. Front. Microbiol. 2021, 12, 776892. [Google Scholar] [CrossRef]
- Koene, M.; Mevius, D.; Wagenaar, J.; Harmanus, C.; Hensgens, M.; Meetsma, A.; Putirulan, F.; van Bergen, M.; Kuijper, E. Clostridium difficile in Dutch animals: Their presence, characteristics and similarities with human isolates. Clin. Microbiol. Infect. 2012, 18, 778–784. [Google Scholar] [CrossRef] [Green Version]
- de Boer, E.; Zwartkruis-Nahuis, A.; Heuvelink, A.E.; Harmanus, C.; Kuijper, E.J. Prevalence of Clostridium difficile in retailed meat in the Netherlands. Int. J. Food Microbiol. 2011, 144, 561–564. [Google Scholar] [CrossRef]
- Kecerova, Z.; Cizek, A.; Nyc, O.; Krutova, M. Clostridium difficile isolates derived from Czech horses are resistant to enrofloxacin; cluster to clades 1 and 5 and ribotype 033 predominates. Anaerobe 2019, 56, 17–21. [Google Scholar] [CrossRef]
- Yin, C.; Chen, D.S.; Zhuge, J.; McKenna, D.; Sagurton, J.; Wang, G.; Huang, W.; Dimitrova, N.; Fallon, J.T. Complete Genome Sequences of Four Toxigenic Clostridium difficile Clinical Isolates from Patients of the Lower Hudson Valley, New York, USA. Genome Announc. 2018, 6, e01537-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tkalec, V.; Jamnikar-Ciglenecki, U.; Rupnik, M.; Vadnjal, S.; Zelenik, K.; Biasizzo, M. Clostridioides difficile in national food surveillance, Slovenia, 2015 to 2017. Euro Surveill. 2020, 25, 1900479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, K.; Egan, S.; Lynch, H.; Harmanus, C.; Kyne, L.; Herra, C.; McDermott, S.; Kuijper, E.; Fitzpatrick, F.; FitzGerald, S.; et al. PCR-ribotype distribution of Clostridium difficile in Irish pigs. Anaerobe 2017, 48, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Zamora, E.; Weimer, B.C.; Torres, R.C.; Gómez-Delgado, A.; Ortiz-Olvera, N.; Aparicio-Ozores, G.; Barbero-Becerra, V.J.; Torres, J.; Camorlinga-Ponce, M. Molecular Epidemiology and Antimicrobial Resistance of Clostridioides difficile in Hospitalized Patients From Mexico. Front. Microbiol. 2022, 12, 787451. [Google Scholar] [CrossRef] [PubMed]
- van Rossen, T.M.; van Prehn, J.; Koek, A.; Jonges, M.; van Houdt, R.; van Mansfeld, R.; Kuijper, E.J.; Vandenbroucke-Grauls, C.M.J.E.; Budding, A.E. Simultaneous detection and ribotyping of Clostridioides difficile, and toxin gene detection directly on fecal samples. Antimicrob. Resist. Infect. Control 2021, 10, 23. [Google Scholar] [CrossRef]
- National Centre for Biotechnology Information (NCBI). Clostridioides difficile Strain CFSAN096664 Chromosome, Complete Genome; NCBI Reference Sequence: NZ_CP049958.1. 2020. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genome/535?genome_assembly_id=1483731 (accessed on 1 August 2022).
- Rodriguez-Palacios, A.; Staempfli, H.R.; Duffield, T.; Weese, J.S. Clostridium difficile in retail ground meat, Canada. Emerg. Infect. Dis. 2007, 13, 485–487. [Google Scholar] [CrossRef]
- Schwanbeck, J.; Riedel, T.; Laukien, F.; Schober, I.; Oehmig, I.; Zimmermann, O.; Overmann, J.; Groß, U.; Zautner, A.E.; Bohne, W. Characterization of a clinical Clostridioides difficile isolate with markedly reduced fidaxomicin susceptibility and a V1143D mutation in rpoB. J. Antimicrob. Chemother. 2019, 74, 6–10. [Google Scholar] [CrossRef]
- Nazareth, C.; Leitão, I.; Reis, E.; Inácio, H.; Martins, F.; Ramalheira, E.; Cunha, F.; Santos, C.; Lino, S.; Moreira, H.; et al. Epidemiology of Clostridioides difficile Infection in Portugal: A Retrospective, Observational Study of Hospitalized Patients. Acta Med. Port. 2022, 35, 270–278. [Google Scholar] [CrossRef]
- Groß, U.; Brzuszkiewicz, E.; Gunka, K.; Starke, J.; Riedel, T.; Bunk, B.; Spröer, C.; Wetzel, D.; Poehlein, A.; Chibani, C.; et al. Comparative genome and phenotypic analysis of three Clostridioides difficile strains isolated from a single patient provide insight into multiple infection of, C. difficile. BMC Genom. 2018, 19, 1. [Google Scholar] [CrossRef] [Green Version]
- Gerding, D.N.; Sambol, S.P.; Johnson, S. Non-toxigenic Clostridioides (Formerly Clostridium) difficile for Prevention of, C. difficile Infection: From Bench to Bedside Back to Bench and Back to Bedside. Front. Microbiol. 2018, 9, 1700. [Google Scholar] [CrossRef]
- Camorlinga, M.; Sanchez-Rojas, M.; Torres, J.; Romo-Castillo, M. Phenotypic Characterization of Non-toxigenic Clostridioides difficile Strains Isolated From Patients in Mexico. Front. Microbiol. 2019, 10, 84. [Google Scholar] [CrossRef]
- Dingle, K.E.; Griffiths, D.; Didelot, X.; Evans, J.; Vaughan, A.; Kachrimanidou, M.; Stoesser, N.; Jolley, K.A.; Golubchik, T.; Harding, R.M.; et al. Clinical Clostridium difficile: Clonality and pathogenicity locus diversity. PLoS ONE 2011, 6, e19993. [Google Scholar] [CrossRef]
- Stabler, R.A.; Gerding, D.N.; Songer, J.G.; Drudy, D.; Brazier, J.S.; Trinh, H.T.; Witney, A.A.; Hinds, J.; Wren, B.W. Comparative phylogenomics of Clostridium difficile reveals clade specificity and microevolution of hypervirulent strains. J. Bacteriol. 2006, 188, 7297–7305. [Google Scholar] [CrossRef] [Green Version]
- Neely, F.; Lambert, M.L.; Van Broeck, J.; Delmée, M. Clinical and laboratory features of the most common Clostridium difficile ribotypes isolated in Belgium. J. Hosp. Infect. 2017, 95, 394–399. [Google Scholar] [CrossRef]
- Muñoz, M.; Restrepo-Montoya, D.; Kumar, N.; Iraola, G.; Camargo, M.; Díaz-Arévalo, D.; Roa-Molina, N.S.; Tellez, M.A.; Herrera, G.; Ríos-Chaparro, D.I.; et al. Integrated genomic epidemiology and phenotypic profiling of Clostridium difficile across intra-hospital and community populations in Colombia. Sci. Rep. 2019, 9, 11293. [Google Scholar] [CrossRef] [Green Version]
- Drigo, I.; Mazzolini, E.; Bacchin, C.; Tonon, E.; Puiatti, C.; Bano, L.; Spigaglia, P.; Barbanti, F.; Agnoletti, F. Molecular characterization and antimicrobial susceptibility of Clostridium difficile isolated from rabbits raised for meat production. Vet. Microbiol. 2015, 181, 303–307. [Google Scholar] [CrossRef]
- Jacob, J.; Evers, S.; Bischoff, K.; Carlier, C.; Courvalin, P. Characterization of the sat4 gene encoding a streptothricin acetyltransferase in Campylobacter coli BE/G4. FEMS Microbiol. Lett. 1994, 120, 13–17. [Google Scholar] [CrossRef]
- Werner, G.; Hildebrandt, B.; Witte, W. Aminoglycoside-streptothricin resistance gene cluster aadE-sat4-aphA-3 disseminated among multiresistant isolates of Enterococcus faecium. Antimicrob. Agents Chemother. 2001, 45, 3267–3269. [Google Scholar] [CrossRef] [Green Version]
- Cafiso, V.; Lo Verde, F.; Zega, A.; Pigola, G.; Rostagno, R.; Borrè, S.; Stefani, S. Genomic Characterization of a New Biofilm-Forming and Adhesive ST398 Human-Adapted MSSA Lineage Causing Septic Knee Arthritis Following Surgical Reconstruction. Microorganisms 2021, 9, 305. [Google Scholar] [CrossRef]
- Suzuki, H.; Tomita, M.; Tsai, P.J.; Ko, W.C.; Hung, Y.P.; Huang, I.H.; Chen, J.W. Comparative genomic analysis of Clostridium difficile ribotype 027 strains including the newly sequenced strain NCKUH-21 isolated from a patient in Taiwan. Gut Pathog. 2017, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Knight, D.R.; Squire, M.M.; Collins, D.A.; Riley, T.V. Genome Analysis of Clostridium difficile PCR Ribotype 014 Lineage in Australian Pigs and Humans Reveals a Diverse Genetic Repertoire and Signatures of Long-Range Interspecies Transmission. Front. Microbiol. 2017, 7, 2138. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.C.; Androga, G.O.; Knight, D.R.; Moono, P.; Foster, N.F.; Riley, T.V. Antimicrobial susceptibility of Clostridium difficile isolated from food and environmental sources in Western Australia. Int. J. Antimicrob. Agents 2018, 52, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Spigaglia, P. Recent advances in the understanding of antibiotic resistance in Clostridium difficile infection. Ther. Adv. Infect. Dis. 2016, 3, 23–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dridi, L.; Tankovic, J.; Petit, J. CdeA of Clostridium difficile, a new multidrug efflux transporter of the MATE family. Microb. Drug Resist. 2004, 10, 191–196. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, J.R.; Galang, M.A.; Sambol, S.P.; Hecht, D.W.; Vedantam, G.; Gerding, D.N.; Johnson, S. Rifampin and rifaximin resistance in clinical isolates of Clostridium difficile. Antimicrob. Agents Chemother. 2008, 52, 2813–2817. [Google Scholar] [CrossRef] [Green Version]
- Toth, M.; Stewart, N.K.; Smith, C.; Vakulenko, S.B. Intrinsic Class D β-Lactamases of Clostridium difficile. mBio 2018, 6, e01803-18. [Google Scholar] [CrossRef] [Green Version]
- Stewart, N.K.; Smith, C.A.; Toth, M.; Stasyuk, A.; Vakulenko, S.B. The crystal structures of CDD-1, the intrinsic class D β-lactamase from the pathogenic Gram-positive bacterium Clostridioides difficile, and its complex with cefotaxime. J. Struct. Biol. 2019, 208, 107391. [Google Scholar] [CrossRef]
- Heir, E.; Sundheim, G.; Holck, A.L. The qacG gene on plasmid pST94 confers resistance to quaternary ammonium compounds in staphylococci isolated from the food industry. J. Appl. Microbiol. 1999, 86, 378–388. [Google Scholar] [CrossRef]
- Poole, K. Efflux pumps as antimicrobial resistance mechanisms. Ann. Med. 2007, 39, 162–176. [Google Scholar] [CrossRef]
- Wong, T.Z.; Zhang, M.; O’Donoghue, M.; Boost, M. Presence of antiseptic resistance genes in porcine methicillin-resistant Staphylococcus aureus. Vet. Microbiol. 2013, 162, 977–979. [Google Scholar] [CrossRef]
- Seier-Petersen, M.A.; Nielsen, L.N.; Ingmer, H.; Aarestrup, F.M.; Agersø, Y. Biocide Susceptibility of Staphylococcus aureus CC398 and CC30 Isolates from Pigs and Identification of the Biocide Resistance Genes, qacG and qacC. Microb. Drug Resist. 2015, 21, 527–536. [Google Scholar] [CrossRef]
- Ignak, S.; Nakipoglu, Y.; Gurler, B. Frequency of antiseptic resistance genes in clinical staphycocci and enterococci isolates in Turkey. Antimicrob. Resist. Infect. Control 2017, 30, 88. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Fu, L.; Huang, M.; Zhang, J.P.; Wu, Y.; Zhou, Y.S.; Zeng, J.; Wang, G.X. Frequency of antiseptic resistance genes and reduced susceptibility to biocides in carbapenem-resistant Acinetobacter baumannii. J. Med. Microbiol. 2017, 66, 13–17. [Google Scholar] [CrossRef]
Figure 1.
Layout of the 3 spinach fields used in this study and their surroundings. The sampling technique used to cover most of the area is detailed in each field.
Figure 1.
Layout of the 3 spinach fields used in this study and their surroundings. The sampling technique used to cover most of the area is detailed in each field.

Figure 2.
Location of the C. difficile positive samples in the 3 fields tested.

Figure 3.
Pangenome analysis for the 8 C. difficile isolates from spinach fields. Gene clusters are indicated as opaque coloured elements and ordered based on their presence and absence, using the Euclidean distance and Ward clustering (inner dendrogram). Samples are hierarchically clustered based on the frequency of gene clusters (outer dendrogram). CD ‘number’ = C. difficile followed by the isolate ID.
Figure 3.
Pangenome analysis for the 8 C. difficile isolates from spinach fields. Gene clusters are indicated as opaque coloured elements and ordered based on their presence and absence, using the Euclidean distance and Ward clustering (inner dendrogram). Samples are hierarchically clustered based on the frequency of gene clusters (outer dendrogram). CD ‘number’ = C. difficile followed by the isolate ID.

Figure 4.
Phylogenetic tree of the C. difficile spinach (2 and 3) and soil isolates (1, 4–8) with the distances between isolates presented as a distance matrix.
Figure 4.
Phylogenetic tree of the C. difficile spinach (2 and 3) and soil isolates (1, 4–8) with the distances between isolates presented as a distance matrix.

Table 2.
Virulence profiles of the C. difficile isolates from spinach (isolates 2 and 3) and soil (isolates 1, 4, 5, 6, 7, 8) samples after MLST and ABRicate analysis (vdfb).
Table 2.
Virulence profiles of the C. difficile isolates from spinach (isolates 2 and 3) and soil (isolates 1, 4, 5, 6, 7, 8) samples after MLST and ABRicate analysis (vdfb).
| ID 1 | ST 2 | Clade | Toxin Genes 3 | |||
|---|---|---|---|---|---|---|
| tcdA | tcdB | cdtA | cdtB | |||
| 1 | 19 | 1 | + | + | − | − |
| 2 | 11 | 5 | + | + | + | + |
| 3 | 11 | 5 | + | + | + | + |
| 4 | 12 | 1 | + | + | − | − |
| 5 | 46 | 1 | + | + | − | − |
| 6 | 16 | 1 | + | + | − | − |
| 7 | 79 | 1 | − | − | − | − |
| 8 | 49 | 1 | + | + | − | − |
1 ID: number assigned to each isolate. 2 ST: Sequence type. 3 (+/−): Presence (+) or absence (−) of the gene indicated.
Table 4.
Heat map comparing the antimicrobial resistance (AMR) profiles observed by E-test (phenotypic) and CARD (genotypic).
Table 4.
Heat map comparing the antimicrobial resistance (AMR) profiles observed by E-test (phenotypic) and CARD (genotypic).
| Genotypic AMR Profile | ||||||
| ID | 23S rRNA Mutation (Erythromycin) | nimB Mutation 1 | 23S rRNA Mutation (Clindamycin) | cdeA | vanB | RpoB Mutation 2 |
| 1 | ||||||
| 2 | ||||||
| 3 | ||||||
| 4 | ||||||
| 5 | ||||||
| 6 | ||||||
| 7 | ||||||
| 8 | ||||||
| Erythromycin | Metronidazole | Clindamycin | Moxifloxacin | Vancomycin | Rifampicin | |
| Phenotypic AMR profile | ||||||
| Only in phenotypic AMR analysis | ||||||
| Only in genotypic AMR analysis | ||||||
| Found in both genotypic and phenotypic AMR analysis | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Marcos, P.; Whyte, P.; Burgess, C.; Ekhlas, D.; Bolton, D. Detection and Genomic Characterisation of Clostridioides difficile from Spinach Fields. Pathogens 2022, 11, 1310. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11111310
AMA Style
Marcos P, Whyte P, Burgess C, Ekhlas D, Bolton D. Detection and Genomic Characterisation of Clostridioides difficile from Spinach Fields. Pathogens. 2022; 11(11):1310. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11111310
Chicago/Turabian StyleMarcos, Pilar, Paul Whyte, Catherine Burgess, Daniel Ekhlas, and Declan Bolton. 2022. "Detection and Genomic Characterisation of Clostridioides difficile from Spinach Fields" Pathogens 11, no. 11: 1310. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11111310
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.