
Vaccinium virgatum Aiton Leaves Extract Suppressed Lipid Accumulation and Uric Acid Production in 3T3-L1 Adipocytes

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
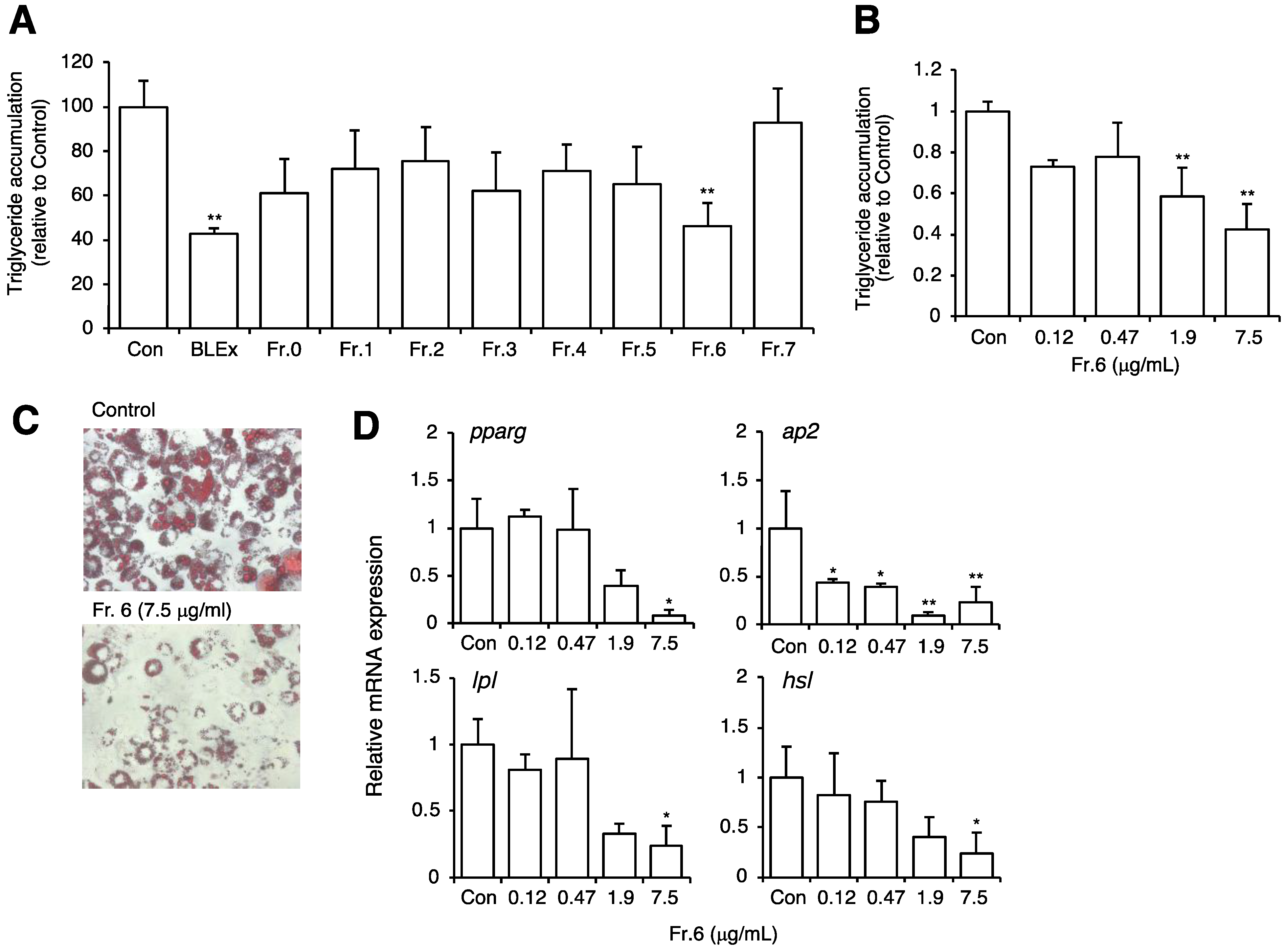
2.1. Effect of BLEx on Adipocyte Maturation
2.2. Effect of Fractionated BLEx on Adipocyte Maturation
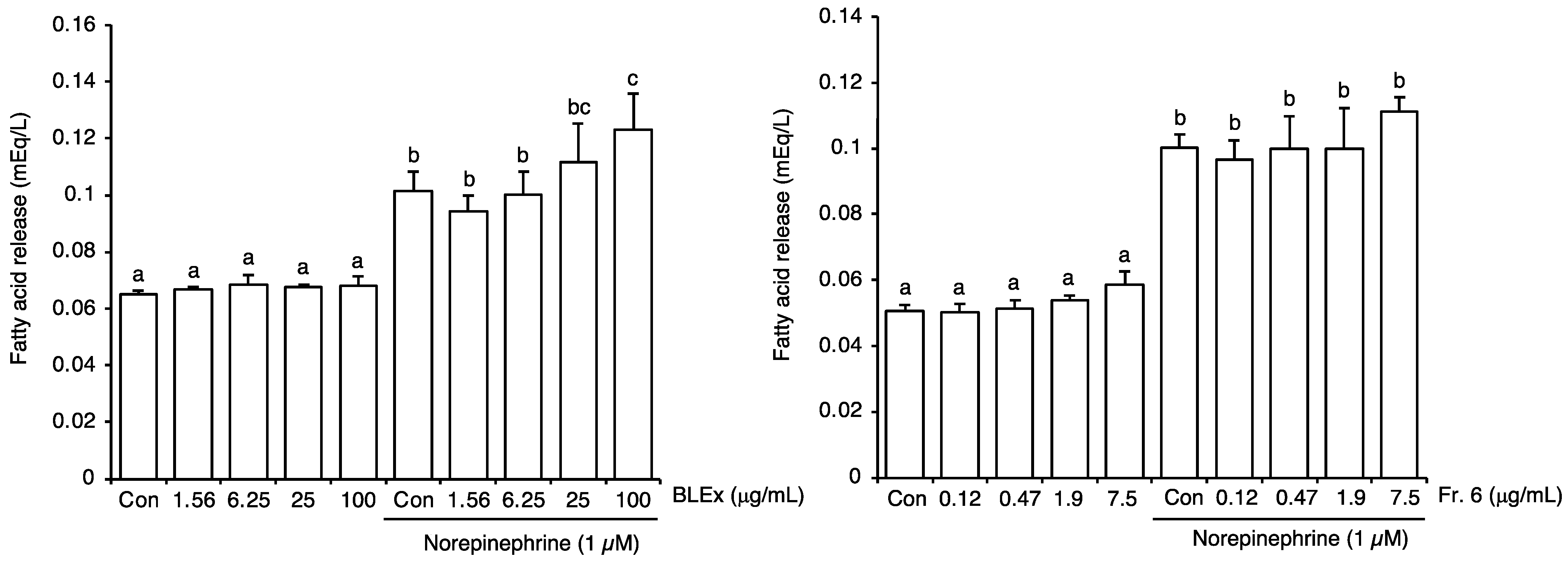
2.3. Effect on Lipolytic Activity of Adipocytes
2.4. Analysis of Proanthocyanidins
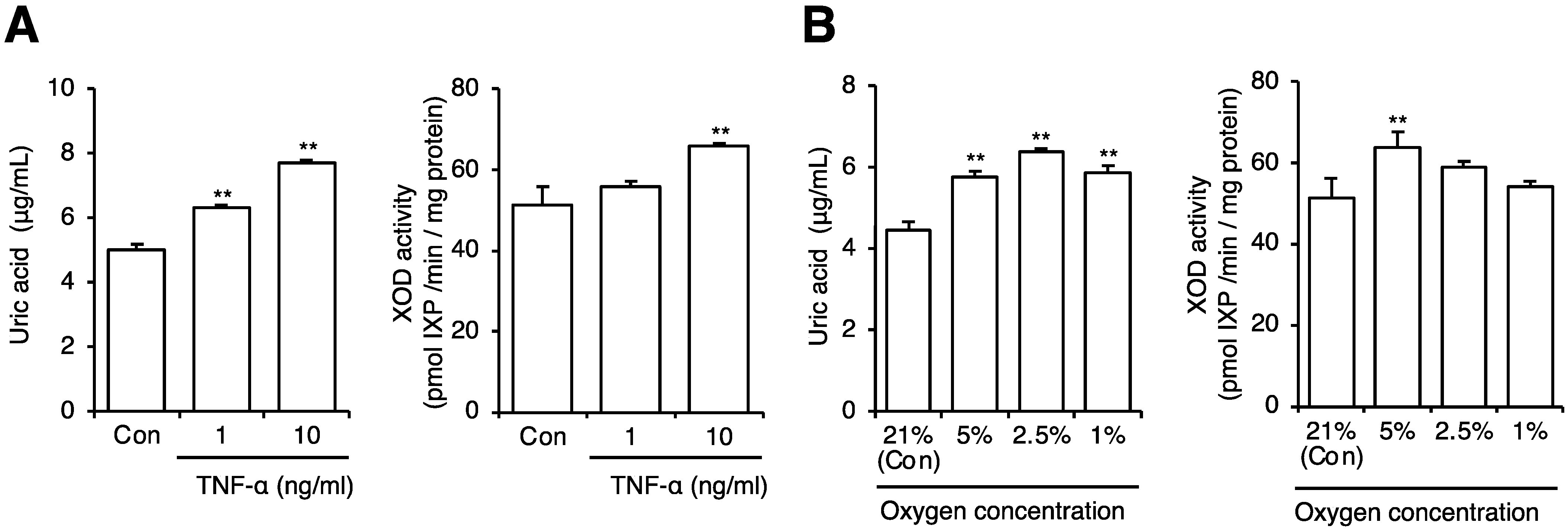
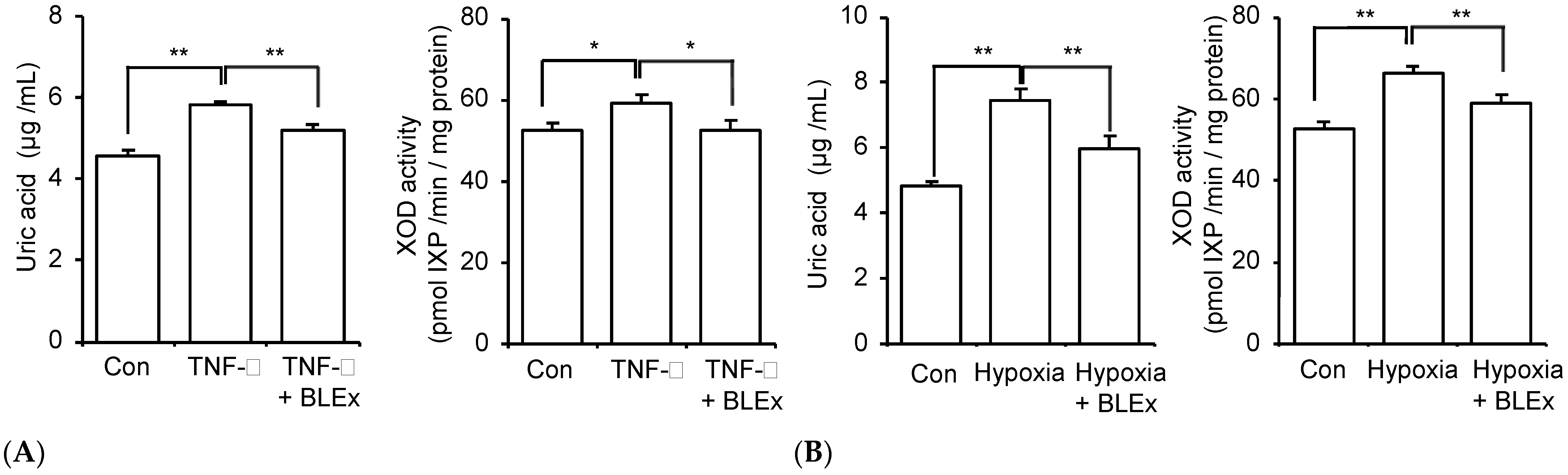
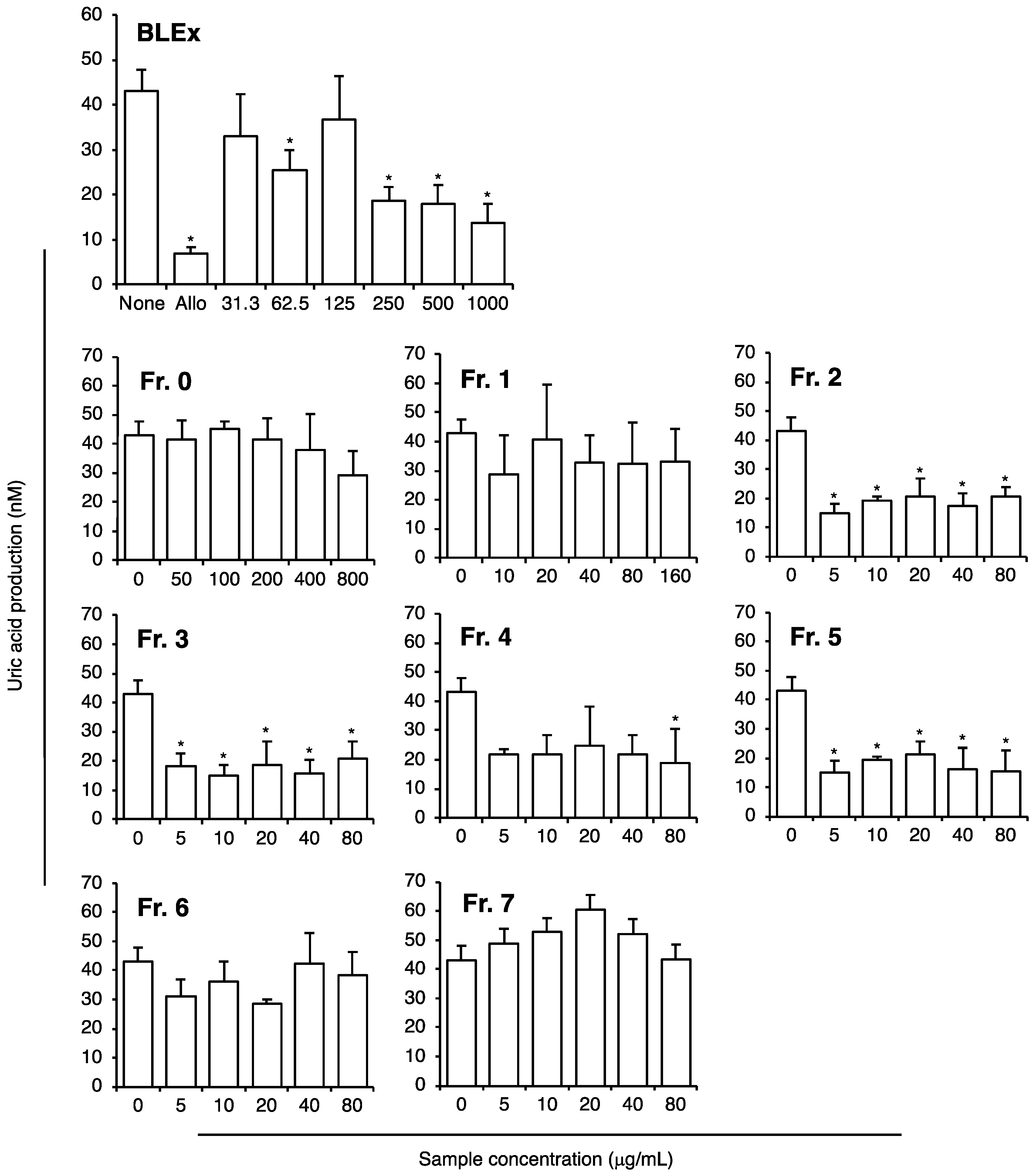
2.5. Effect on Uric Acid Production from Adipocytes
2.6. Direct Inhibitory Effect on XOD Activity
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Oil Red O Staining
4.4. Real-Time PCR
4.5. Fractionation of BLEx
4.6. Quantification of Proanthocyanidins
4.7. Analysis of Polyphenol
4.8. Lipolysis Assay
4.9. Uric Acid Measurement by HPLC
4.10. XOR Activity
4.11. XOD Inhibition Assay
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Alkhalidy, H.; Liu, D. The emerging role of polyphenols in the management of type 2 diabetes. Molecules 2021, 26, 703. [Google Scholar] [CrossRef]
- Abenavoli, L.; Larussa, T.; Corea, A.; Procopio, A.C.; Boccuto, L.; Dallio, M.; Federico, A.; Luzza, F. Dietary polyphenols and non-alcoholic fatty liver disease. Nutrients 2021, 13, 494. [Google Scholar] [CrossRef] [PubMed]
- Diotallevi, C.; Fava, F.; Gobbetti, M.; Tuohy, K. Healthy dietary patterns to reduce obesity-related metabolic disease: Polyphenol-microbiome interactions unifying health effects across geography. Curr. Opin. Clin. Nutr. Metab. Care 2020, 23, 437–444. [Google Scholar] [CrossRef]
- Shi, D.; Xu, M.; Ren, M.; Pan, E.; Luo, C.; Zhang, W.; Tang, Q. Immunomodulatory effect of flavonoids of blueberry (Vaccinium corymbosum L.) leaves via the NF-κB signal pathway in LPS-Stimulated RAW 264.7 Cells. J. Immunol. Res. 2017, 2017, 5476903. [Google Scholar] [CrossRef] [Green Version]
- Tetsumura, T.; Kajiwara, Y.; Honsho, C.; Sato-Yamauchi, M.; Sugimoto, Y.; Kunitake, H. Effective micropropagation of rabbiteye blueberries for leaf tea production environment. Environ. Control Biol. 2012, 50, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, M.; Matsuyama, Y.; Hayasegawa, R.; Hamada, K.; Nishiyama, K.; Kai, T.; Kamenaga, K.; Arakawa, T.; Tari, H.; Shimizu, Y.; et al. Blueberry (Vaccinium virgatum Aiton) leaf extract ameliorates insulin resistance in mice fed a high-fat, high-sucrose diet. Food Sci. Technol. Res. 2015, 21, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Fujii, K.; Ota, Y.; Nishiyama, K.; Kunitake, H.; Yamasaki, Y.; Tari, H.; Araki, K.; Arakawa, T.; Yamasaki, M. Blueberry leaf polyphenols prevent body fat accumulation in mice fed High-fat, High-sucrose Diet. J. Oleo Sci. 2019, 68, 471–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, H.; Li, Y.; Chen, H.; Wang, C.; Wong, V.K.W.; Jiang, Z.; Zhang, W. Phytotherapy using blueberry leaf polyphenols to alleviate non-alcoholic fatty liver disease through improving mitochondrial function and oxidative defense. Phytomedicine 2020, 69, 153209. [Google Scholar] [CrossRef]
- Shoji, K.; Yamasaki, M.; Kunitake, H. Effects of dietary blueberry (Vaccinium ashei Reade) leaves on mildly postprandial hypertriglyceridemia. J. Oleo Sci. 2020, 69, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, Y.; Fujita, Y.; Ohnishi, S.; Tanaka, T.; Hirabaru, H.; Kai, T.; Sakaida, H.; Nishizono, S.; Kouno, I. Chemical constituents of the leaves of rabbiteye blueberry (Vaccinium ashei) and characterization of polymeric proanthocyanidins containing phenylpropanoid units and A-type linkages. Food Chem. 2010, 121, 1073–1079. [Google Scholar] [CrossRef] [Green Version]
- Takeshita, M.; Ishida, Y.-I.; Akamatsu, E.; Ohmori, Y.; Sudoh, M.; Uto, H.; Tsubouchi, H.; Kataoka, H. Proanthocyanidin from blueberry leaves suppresses expression of Subgenomic hepatitis C Virus RNA. J. Biol. Chem. 2009, 284, 21165–21176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.-Y.; Fu, L.; Li, C.; Xu, L.-P.; Zhang, L.-X.; Zhang, W.-M. Quercetin, Hyperin, and chlorogenic acid improve endothelial function by antioxidant, Antiinflammatory, and ACE Inhibitory Effects. J. Food Sci. 2017, 82, 1239–1246. [Google Scholar] [CrossRef]
- Lee, J.; Lee, J.-Y.; Lee, J.-H.; Jung, S.-M.; Suh, Y.S.; Koh, J.-H.; Kwok, S.-K.; Ju, J.H.; Park, K.-S.; Park, S.-H. Visceral fat obesity is highly associated with primary gout in a metabolically obese but normal weighted population: A case control study. Arthritis Res. Ther. 2015, 17, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, N.; Xu, H.; Sun, Q.; Yu, X.; Chen, W.; Wei, H.; Jiang, J.; Xu, Y.; Lu, W. The role of oxidative stress in hyperuricemia and xanthine oxidoreductase (XOR) inhibitors. Oxidative Med. Cell. Longev. 2021, 2021, 1470380. [Google Scholar] [CrossRef] [PubMed]
- Furuhashi, M. New insights into purine metabolism in metabolic diseases: Role of xanthine oxidoreductase activity. Am. J. Physiol. Metab. 2020, 319, E827–E834. [Google Scholar] [CrossRef]
- Nishikawa, T.; Nagata, N.; Shimakami, T.; Shirakura, T.; Matsui, C.; Ni, Y.; Zhuge, F.; Xu, L.; Chen, G.; Nagashimada, M.; et al. Xanthine oxidase inhibition attenuates insulin resistance and diet-induced steatohepatitis in mice. Sci. Rep. 2020, 10, 815. [Google Scholar] [CrossRef] [PubMed]
- Tsushima, Y.; Nishizawa, H.; Tochino, Y.; Nakatsuji, H.; Sekimoto, R.; Nagao, H.; Shirakura, T.; Kato, K.; Imaizumi, K.; Takahashi, H.; et al. Uric acid secretion from adipose tissue and its increase in obesity. J. Biol. Chem. 2013, 288, 27138–27149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehmood, A.; Ishaq, M.; Zhao, L.; Safdar, B.; Rehman, A.; Munir, M.; Raza, A.; Nadeem, M.; Iqbal, W.; Wang, C. Natural compounds with xanthine oxidase inhibitory activity: A review. Chem. Biol. Drug Des. 2019, 93, 387–418. [Google Scholar] [CrossRef]
- Lin, C.-M.; Chen, C.-S.; Liang, Y.-C.; Lin, J.-K. Molecular modeling of flavonoids that inhibits xanthine oxidase. Biochem. Biophys. Res. Commun. 2002, 294, 167–172. [Google Scholar] [CrossRef]
- Hatano, T.; Yasuhara, T.; Yoshihara, R.; Agata, I.; Noro, T.; Okuda, T. Effects of interaction of tannins with co-existing substances. VII. Inhibitory effects of tannins and related polyphenols on xanthine oxidase. Chem. Pharm. Bull. 1990, 38, 1224–1229. [Google Scholar] [CrossRef] [PubMed]
- Arimboor, R.; Arumughan, C. Effect of Polymerization on antioxidant and xanthine oxidase inhibitory potential of sea buckthorn (H. rhamnoides) proanthocyanidins. J. Food Sci. 2012, 77, C1036–C1041. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, P.; Navarro-Herrera, D.; Zabala, M.; Miguéliz, I.; Romo-Hualde, A.; López-Yoldi, M.; Martínez, J.A.; Vizmanos, J.L.; Milagro, F.I.; González-Navarro, C.J. 3T3-L1 adipogenesis depending on the stage of differentiation and their binding affinity to PPARγ. Molecules 2019, 24, 1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Gao, M.; Liu, D. Chlorogenic acid improves high fat diet-induced hepatic steatosis and insulin resistance in mice. Pharm. Res. 2015, 32, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.-S.; Jeon, S.-M.; Kim, M.-J.; Yeo, J.; Seo, K.-I.; Choi, M.-S.; Lee, M.-K. Chlorogenic acid exhibits anti-obesity property and improves lipid metabolism in high-fat diet-induced-obese mice. Food Chem. Toxicol. 2010, 48, 937–943. [Google Scholar] [CrossRef]
- Peng, S.-G.; Pang, Y.-L.; Zhu, Q.; Kang, J.-H.; Liu, M.-X.; Wang, Z. Chlorogenic acid functions as a novel agonist of PPARγ2 during the differentiation of mouse 3T3-L1 preadipocytes. BioMed Res. Int. 2018, 2018, 1–14. [Google Scholar] [CrossRef]
- Naowaboot, J.; Chung, C.H.; Choi, R. Rutin stimulates adipocyte differentiation and adiponectin secretion in 3T3-L1 Adipocytes. J. Med. Assoc. Thail. 2015, 98, S1–S6. [Google Scholar]
- Choi, I.; Park, Y.; Choi, H.; Lee, E.H. Anti-adipogenic activity of rutin in 3T3-L1 cells and mice fed with high-fat diet. BioFactors 2006, 26, 273–281. [Google Scholar] [CrossRef]
- Duangjai, A.; Nuengchamnong, N.; Suphrom, N.; Trisat, K.; Limpeanchob, N.; Saokaew, S. Potential of Coffee Fruit Extract and Quinic Acid on Adipogenesis and Lipolysis in 3T3-L1 Adipocytes. Kobe J. Med. Sci. 2018, 64, E84–E92. [Google Scholar] [PubMed]
- Park, J.Y.; Kim, Y.; Im, J.A.; You, S.; Lee, H. Inhibition of adipogenesis by oligonol through Akt-mTOR inhibition in 3T3-L1 adipocytes. Evid.-Based Complementary Altern. Med. 2014, 2014, 895272. [Google Scholar] [CrossRef]
- Tie, F.; Wang, J.; Liang, Y.; Zhu, S.; Wang, Z.; Li, G.; Wang, H. Proanthocyanidins ameliorated deficits of lipid metabolism in Type 2 diabetes mellitus via inhibiting adipogenesis and improving mitochondrial function. Int. J. Mol. Sci. 2020, 21, 2029. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, K.; Olejnik, A.; Rychlik, J.; Grajek, W. Cranberries (Oxycoccus quadripetalus) inhibit lipid metabolism and modulate leptin and adiponectin secretion in 3T3-L1 adipocytes. Food Chem. 2015, 148, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Osaki, N.; Shimotoyodome, A. Green tea catechins enhance norepinephrine-induced lipolysis via a protein kinase A-dependent pathway in adipocytes. Biochem. Biophys. Res. Commun. 2015, 461, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ardévol, A.; Bladé, C.; Salvadó, M.; Arola, L. Changes in lipolysis and hormone-sensitive lipase expression caused by procyanidins in 3T3-L1 adipocytes. Int. J. Obes. 2000, 24, 319–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, S.; Yamamoto, T.; Tsutsumi, Z.; Moriwaki, Y.; Yamakita, J.; Higashino, K. Close correlation between visceral fat accumulation and uric acid metabolism in healthy men. Metabolism 1997, 46, 1162–1165. [Google Scholar] [CrossRef]
- Sodhi, K.; Hilgefort, J.; Banks, G.; Gilliam, C.; Stevens, S.; Ansinelli, H.A.; Getty, M.; Abraham, N.G.; Shapiro, J.I.; Khitan, Z. Uric acid-induced adipocyte dysfunction is attenuated by HO-1 upregulation: Potential role of antioxidant therapy to target obesity. Stem Cells Int. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Moriya, C.; Satoh, H. Teneligliptin decreases uric acid levels by reducing xanthine dehydrogenase expression in white adipose tissue of male wistar rats. J. Diabetes Res. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bulló-Bonet, M.; Garcia-Lorda, P.; López-Soriano, F.; Argilés, J.; Salas-Salvadó, J. Tumour necrosis factor, a key role in obesity? FEBS Lett. 1999, 451, 215–219. [Google Scholar] [CrossRef] [Green Version]
- Kern, P.A.; Ranganathan, S.; Li, C.; Wood, L.; Ranganathan, G. Adipose tissue tumor necrosis factor and interleukin-6 expression in human obesity and insulin resistance. Am. J. Physiol. Metab. 2001, 280, E745–E751. [Google Scholar] [CrossRef]
- Virdis, A.; Colucci, R.L.; Bernardini, N.; Blandizzi, C.; Taddei, S.; Masi, S. Microvascular endothelial dysfunction in human obesity: Role of TNF-α. J. Clin. Endocrinol. Metab. 2018, 104, 341–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, C.E.; Hare, J.M. Xanthine oxidoreductase and cardiovascular disease: Molecular mechanisms and pathophysiological implications. J. Physiol. 2004, 555, 589–606. [Google Scholar] [CrossRef]
- Abooali, M.; Lall, G.S.; Coughlan, K.; Lall, H.S.; Gibbs, B.F.; Sumbayev, V.V. Crucial involvement of xanthine oxidase in the intracellular signalling networks associated with human myeloid cell function. Sci. Rep. 2014, 4, 6307. [Google Scholar] [CrossRef] [PubMed]
- Nagao, H.; Nishizawa, H.; Tanaka, Y.; Fukata, T.; Mizushima, T.; Furuno, M.; Bamba, T.; Tsushima, Y.; Fujishima, Y.; Kita, S.; et al. Hypoxanthine secretion from human adipose tissue and its increase in hypoxia obesity. Obesity 2018, 26, 1168–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yisireyili, M.; Hayashi, M.; Wu, H.; Uchida, Y.; Yamamoto, K.; Kikuchi, R.; Hamrah, M.S.; Nakayama, T.; Cheng, X.W.; Matsushita, T.; et al. Xanthine oxidase inhibition by febuxostat attenuates stress-induced hyperuricemia, glucose dysmetabolism, and prothrombotic state in mice. Sci. Rep. 2017, 7, 1266. [Google Scholar] [CrossRef]
- Yamasaki, M.; Hamada, K.; Fujii, K.; Nishiyama, K.; Yamasaki, Y.; Tari, H.; Araki, K.; Arakawa, T. Vaccinium ashei leaves extract alleviates insulin resistance via AMPK independent pathway in C2C12 myotube model. Biochem. Biophys. Rep. 2018, 14, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Isa, S.S.P.M.; Ablat, A.; Mohamad, J. The antioxidant and xanthine oxidase inhibitory activity of plumeria rubra flowers. Molecules 2018, 23, 400. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.X.; Wang, Y.; Kong, L.D.; Yang, C.; Zhang, X. Effects of Biota orientalis extract and its flavonoid constituents, quercetin and rutin on serum uric acid levels in oxonate-induced mice and xanthine dehydrogenase and xanthine oxidase activities in mouse liver. J. Ethnopharmacol. 2004, 93, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Aucamp, J.; Gaspar, A.; Hara, Y.; Apostolides, Z. Inhibition of xanthine oxidase by catechins from tea (Camellia sinensis). Anticancer Res. 1998, 17, 4381–4385. [Google Scholar]
- Ștefănescu, B.-E.; Călinoiu, L.F.; Ranga, F.; Fetea, F.; Mocan, A.; Vodnar, D.C.; Crișan, G. The chemical and biological profiles of leaves from commercial blueberry varieties. Plants 2020, 9, 1193. [Google Scholar] [CrossRef]
- Wu, H.; Chai, Z.; Hutabarat, R.P.; Zeng, Q.; Niu, L.; Li, D.; Yu, H.; Huang, W. Blueberry leaves from 73 different cultivars in southeastern China as nutraceutical supplements rich in antioxidants. Food Res. Int. 2019, 122, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Páscoa, R.N.; Gomes, M.J.; Sousa, C. Antioxidant activity of blueberry (Vaccinium spp.) cultivar leaves: Differences across the vegetative stage and the application of near infrared spectroscopy. Molecules 2019, 24, 3900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cezarotto, V.S.; Giacomelli, S.R.; Vendruscolo, M.H.; Vestena, A.S.; Cezarotto, C.S.; da Cruz, R.C.; Maurer, L.H.; Ferreira, L.M.; Emanuelli, T.; Cruz, L. Influence of harvest season and cultivar on the variation of phenolic compounds composition and antioxidant properties in Vaccinium ashei leaves. Molecules 2017, 22, 1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kai, H.; Fuse, T.; Kunitake, H.; Morishita, K.; Matsuno, K. Comparison of cultivars and seasonal variation in blueberry (Vaccinium Species) leaf extract on adult T-Cell Leukemia Cell line growth suppression. Medicines 2014, 1, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, K.; Eto, N.; Sakakibara, Y.; Matsushita, Y.; Sugamoto, K.; Morishita, K.; Suiko, M. Phenolic compounds inhibit oligomeric proanthocyanidins from rabbiteye blueberry leaves inhibits the proliferation of human T-cell lymphotropic virus type 1-associated cell lines via apoptosis and cell cycle arrest. J. Funct. Foods 2014, 6, 356–366. [Google Scholar] [CrossRef] [Green Version]
- Toyama, Y.; Toshima, S.; Hirano, T.; Yamasaki, M.; Kunitake, H. Polyphenol contents, antioxidant activities, and anti-cancer cell proliferation properties at each stage of fruit development in intersectional hybrids between highbush blueberry and shashanbo (Vaccinium bracteatum Thunb.). J. Berry Res. 2021, 1–16, preprint. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction Number | mDP * | A-Type Bond (%) | Cinchonain I Unit (%) | B-Type Bond (%) | Proanthocyanidins (mg/g) |
|---|---|---|---|---|---|
| BLEx | 5.85 | 4.5 | 24.3 | 71.2 | 73.0 |
| 4 | 5.74 | 5.2 | 61.5 | 33.3 | 75.2 |
| 5 | 6.05 | 14.7 | 6.6 | 78.7 | 222 |
| 6 | 7.55 | 4.7 | 5.2 | 90.1 | 301 |
| 7 | 12.28 | 4.4 | 3.8 | 91.8 | 241 |
| Fraction Number | Catechin | Epicatechin | Chlorogenic Acid | Caffeic Acid | Rutin |
|---|---|---|---|---|---|
| BLEx | 13.11 | 5.50 | 119.17 | 1.30 | 22.20 |
| 2 | 1.95 | 30.33 | 278.59 | 5.56 | 17.07 |
| 3 | 39.68 | 106.92 | 198.01 | 10.24 | 36.80 |
| 4 | 59.33 | 16.72 | 0.97 | 1.92 | 217.13 |
| Fraction Number | Eluent | Yield (g) | Yield (%) |
|---|---|---|---|
| 0 | Pass through | 26.42 | 52.83 |
| 1 | Water | 6.08 | 12.16 |
| 2 | Water: MeOH (4:1) | 1.35 | 2.70 |
| 3 | Water: MeOH (3:2) | 0.74 | 1.48 |
| 4 | Water: MeOH (2:3) | 1.99 | 3.98 |
| 5 | Water: MeOH (1:4) | 4.17 | 8.34 |
| 6 | MeOH | 3.77 | 7.54 |
| 7 | Water: acetone (2:3) | 1.76 | 3.52 |
| 8 | Water: acetone (2:3) | 0.05 | 0.10 |
| 9 | Acetone | 0.03 | 0.06 |
| Total | 46.36 | 92.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamasaki, M.; Kiue, Y.; Fujii, K.; Sushida, M.; Yamasaki, Y.; Sugamoto, K.; Suzuki, Y.; Koga, Y.; Kunitake, H.; Kai, H.; et al. Vaccinium virgatum Aiton Leaves Extract Suppressed Lipid Accumulation and Uric Acid Production in 3T3-L1 Adipocytes. Plants 2021, 10, 2638. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122638
Yamasaki M, Kiue Y, Fujii K, Sushida M, Yamasaki Y, Sugamoto K, Suzuki Y, Koga Y, Kunitake H, Kai H, et al. Vaccinium virgatum Aiton Leaves Extract Suppressed Lipid Accumulation and Uric Acid Production in 3T3-L1 Adipocytes. Plants. 2021; 10(12):2638. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122638
Chicago/Turabian StyleYamasaki, Masao, Yusei Kiue, Kento Fujii, Moe Sushida, Yumi Yamasaki, Kazuhiro Sugamoto, Yosuke Suzuki, Yasuko Koga, Hisato Kunitake, Hisahiro Kai, and et al. 2021. "Vaccinium virgatum Aiton Leaves Extract Suppressed Lipid Accumulation and Uric Acid Production in 3T3-L1 Adipocytes" Plants 10, no. 12: 2638. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10122638