
Flavonoid-Rich Fractions of Bauhinia holophylla Leaves Inhibit Candida albicans Biofilm Formation and Hyphae Growth

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Phenolic Compounds’ Content
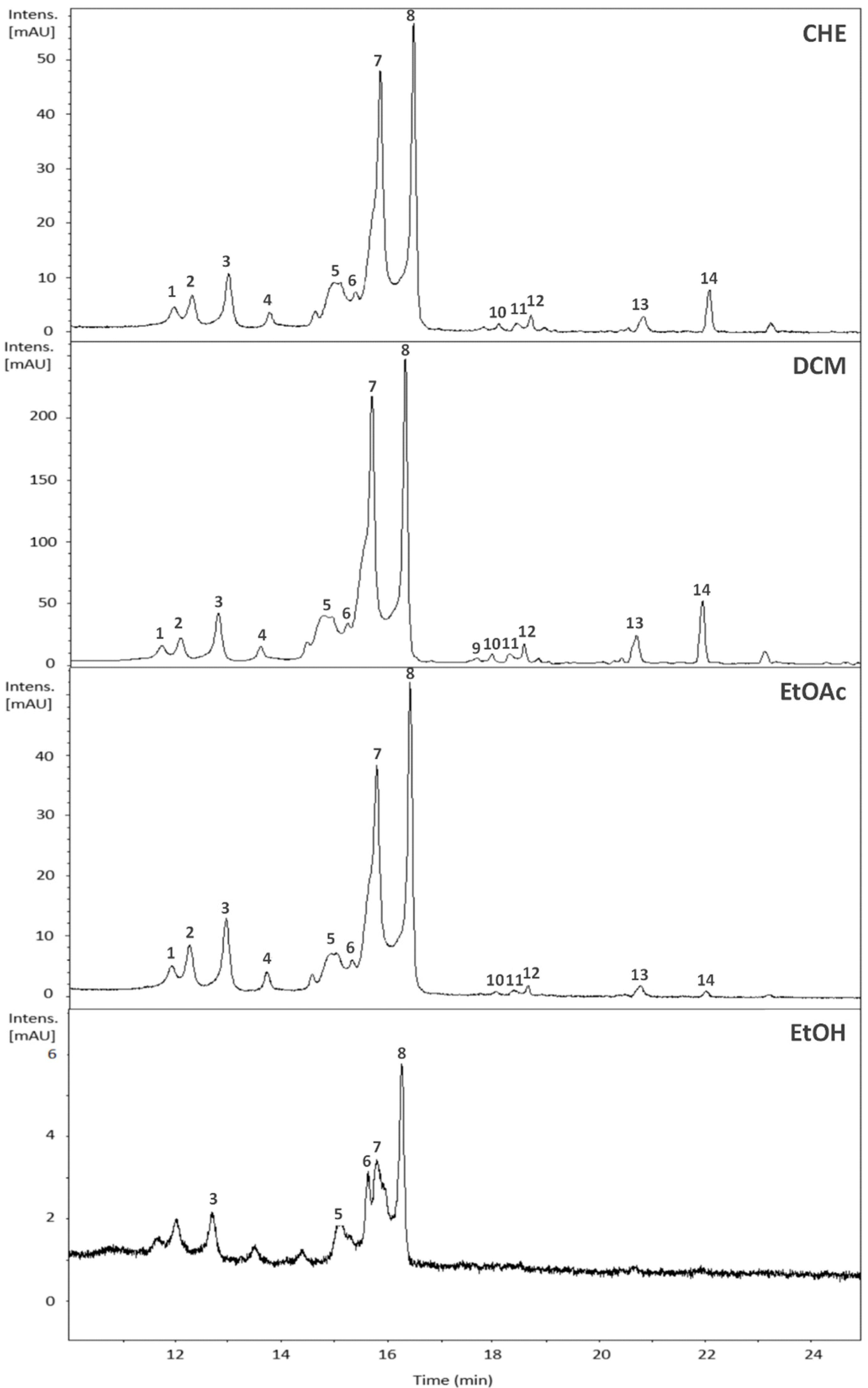
2.2. Chemical Composition of the Crude Extract and Fractions of B. holophylla
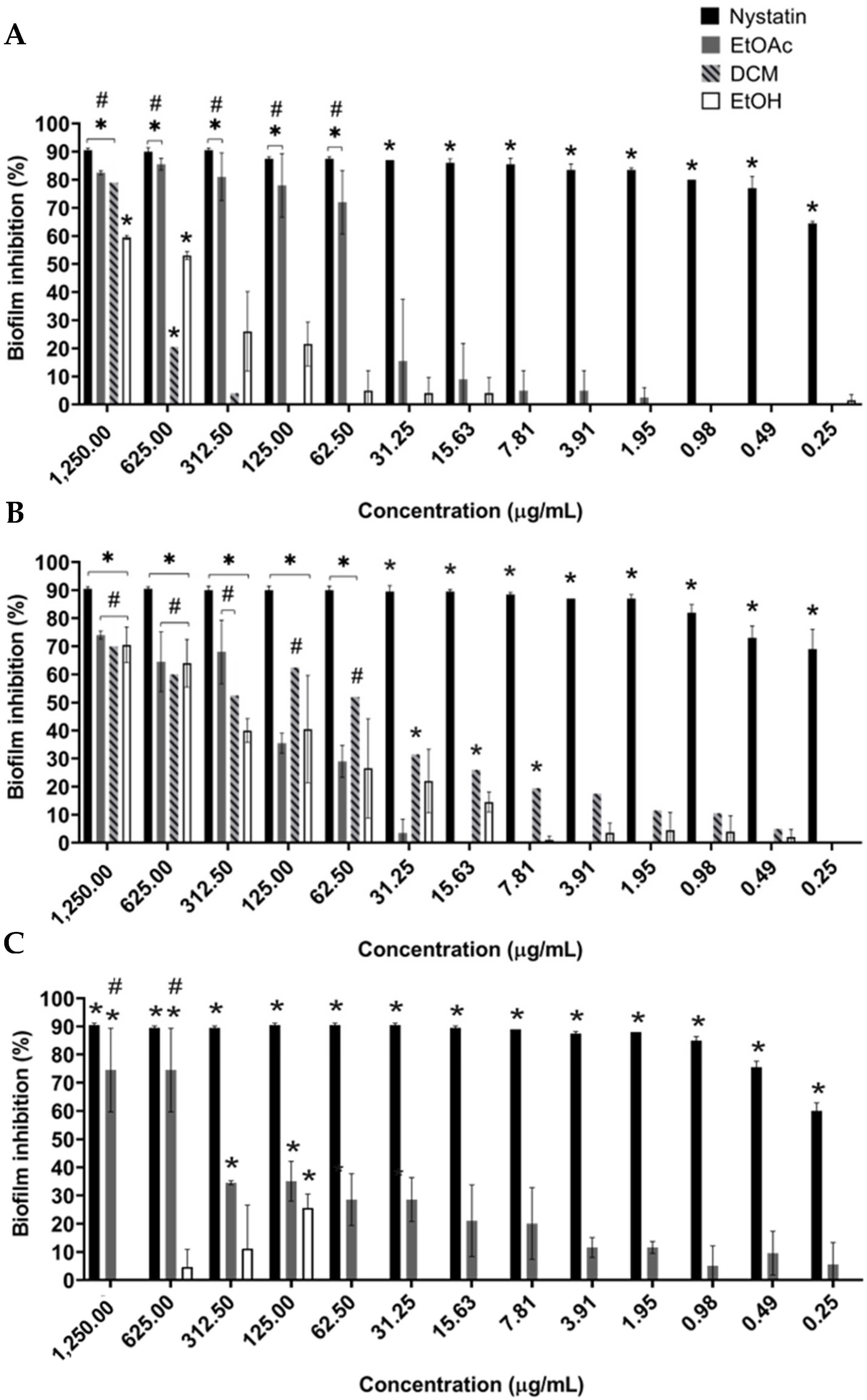
2.3. Evaluation of the Planktonic and Antibiofilm Activity against Candida albicans of Extract and Fractions
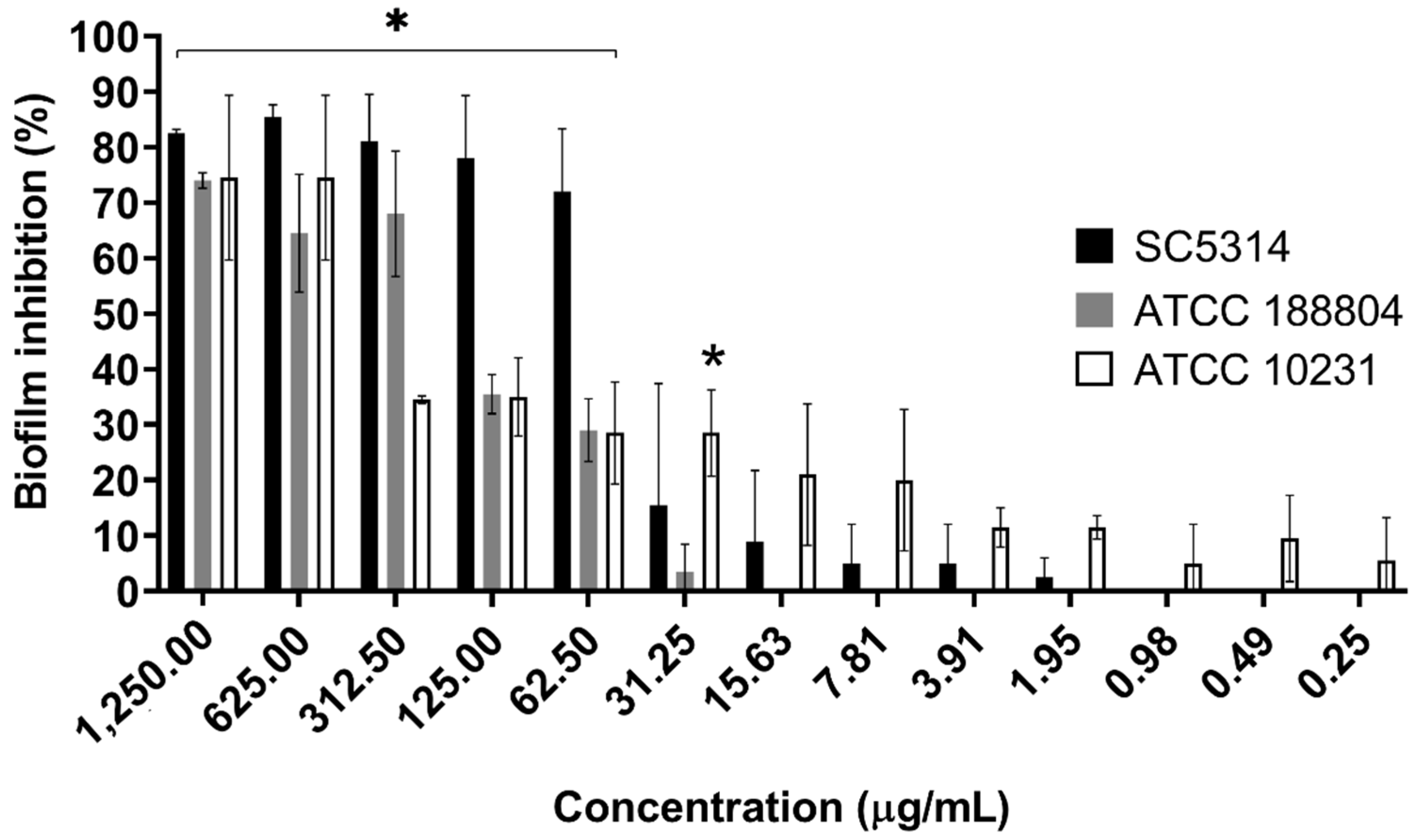
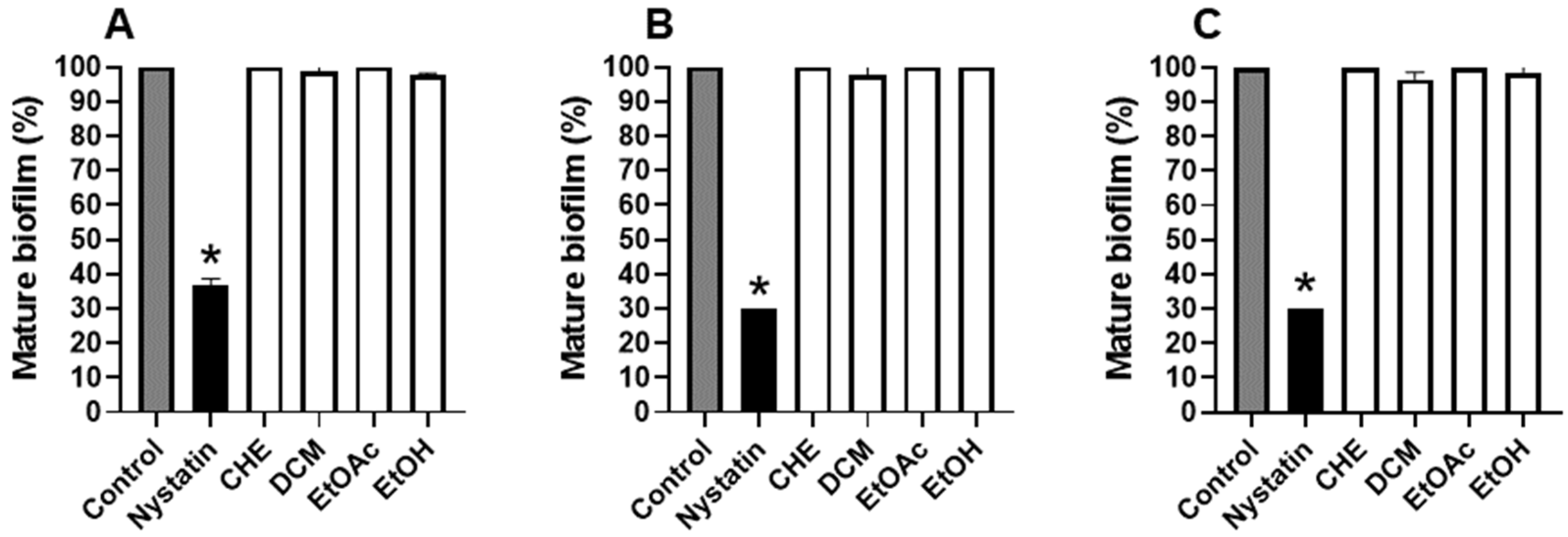
2.4. B. holophylla Fractions Inhibit Biofilm Formation of C. albicans Strains, but Do Not Have Effect on Mature Biofilm
2.5. B. holophylla Fractions Reduce Yeast–Hyphae Transition
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Preparation of the Extract and Fractions
4.3. Phenolic Compounds Content
4.4. HPLC-DAD-MS Analyses
4.5. Antibiofilm Formation Assay
4.5.1. Strains and Culture Conditions
4.5.2. Microdilution Assay: Effect of the B. holophylla Extract and Fractions on C. albicans Planktonic Growth
4.5.3. Antibiofilm Formation Assay
4.5.4. Inhibition of Yeast–Hyphae Transition
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singla, R.K.; Dubey, A.K. Molecules and metabolites from natural products as inhibitors of biofilm in Candida spp. pathogens. Curr. Top. Med. Chem. 2019, 19, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Wall, G.; Montelongo-Jauregui, D.; Bonifacio, B.V.; Lopez-Ribot, J.L.; Uppuluri, P. Candida albicans biofilm growth and dispersal: Contributions to pathogenesis. Curr. Opin. Microbiol. 2019, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Dadar, M.; Tiwari, R.; Karthik, K.; Chakraborty, S.; Shahali, Y.; Dhama, K. Candida albicans-biology, molecular characterization, pathogenicity, and advances in diagnosis and control—An update. Microb. Pathog. 2018, 117, 128–138. [Google Scholar] [CrossRef]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 15, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Sudbery, P.; Gow, N.; Berman, J. The distinct morphogenic states of Candida albicans. Trends Microbiol. 2004, 12, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Correia, I.; Román, E.; Prieto, D.; Hidalgo-Vico, S.; Alonso-Monge, R.; Pla, J. Role of Candida albicans mating in genetic variability and adaptation to the host. Fungal Biol. Rev. 2019, 33, 180–189. [Google Scholar] [CrossRef]
- Stacy, A.; Mcnally, L.; Darch, S.E.; Brown, S.P.; Whiteley, M. The biogeography of polymicrobial infection. Nat. Rev. Microbiol. 2016, 14, 93–105. [Google Scholar] [CrossRef] [Green Version]
- Guzzo, F.; Scognamiglio, M.; Fiorentino, A.; Buommino, E.; D’Abrosca, B. Plant derived natural products against Pseudomonas aeruginosa and Staphylococcus aureus: Antibiofilm activity and molecular mechanisms. Molecules 2020, 25, 5024. [Google Scholar] [CrossRef]
- Aguiar, F.L.L.; Santos, N.C.; de Paula Cavalcante, C.S.; Andreu, D.; Baptista, G.R.; Gonçalves, S. Antibiofilm activity on Candida albicans and mechanism of action on biomembrane models of the antimicrobial peptide Ctn [15–34]. Int. J. Mol. Sci. 2020, 21, 8339. [Google Scholar] [CrossRef]
- D’Acunto, B.; Frunzo, L.; Klapper, I.; Mattei, M.R.; Stoodley, P. Mathematical modeling of dispersal phenomenon in biofilms. Math. Biosci. 2019, 307, 70–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höfs, S.; Mogavero, S.; Hube, B. Interaction of Candida albicans with host cells: Virulence factors, host defense, escape strategies, and the microbiota. J. Microbiol. 2016, 54, 149–169. [Google Scholar] [CrossRef]
- Cavalheiro, M.; Teixeira, M.C. Candida biofilms: Threats, challenges, and promising strategies. Front. Med. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farag, M.A.; Sakna, S.T.; El-Fiky, N.M.; Shabana, M.M.; Wessjohann, L.A. Phytochemical, antioxidant and antidiabetic evaluation of eight Bauhinia L. species from Egypt using UHPLC–PDA–qTOF-MS and chemometrics. Phytochemistry 2015, 119, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure-activity relationship: An uptade review. Phytother. Res. 2018, 33, 13–40. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y. Recent advances in natural antifungal flavonoids and their derivatives. Bioorg. Med. Chem. Lett. 2019, 29, 126589. [Google Scholar] [CrossRef]
- Ninfali, P.; Antonelli, A.; Magnani, M.; Scarpa, E.S. Antiviral properties of flavonoids and delivery strategies. Nutrients 2020, 12, 2534. [Google Scholar] [CrossRef]
- Matilla-Cuenca, L.; Gil, C.; Cuesta, S.; Rapún-Araiz, B.; Ziemyté, M.; Mira, A.; Lasa, I.; Vale, J. Antibiofilm activity of flavonoids on staphylococcal biofilms through targeting BAP amyloids. Sci. Rep. 2020, 10, 18968. [Google Scholar] [CrossRef]
- Pruteanu, M.; Lobato, J.I.H.; Stach, T.; Hengge, R. Common plant flavonoids prevent the assembly of amyloid curli fibres and can interfere with bacterial biofilm formation. Environ. Microbiol. 2020, 22, 5280–5299. [Google Scholar] [CrossRef]
- Kanchanapiboon, J.; Kongsa, U.; Pattamadilok, D.; Kamponchaidet, S.; Wachisunthon, D.; Poonsatha, S.; Tuntoaw, S. Boesenbergia rotunda extract inhibits Candida albicans biofilm formation by pinostrobin and pinocembrin. J. Ethnopharmacol. 2020, 261, 113193. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Yan, S.; Zhou, C.; Liu, Q.; Zhu, H.; Wen, Z. Total flavonoids from Potentilla kleiniana wight et arn inhibits biofilm formation and virulence factors production in methicillin-resistant Staphylococcus aureus (MRSA). J. Ethnopharmacol. 2021, 279, 114383. [Google Scholar] [CrossRef] [PubMed]
- Vaz, A.M.S.F.; Bortulozzi, R.L.C.; da Silva, L.A.E. Checklist of Bauhinia sensu stricto (Caesalpiniaceae) in Brazil. Plant Ecol. Evol. 2010, 143, 212–221. [Google Scholar] [CrossRef]
- Pinheiro, M.S.; Rodrigues, L.S.; Neto, L.S.; Moraes-Souza, R.Q.; Soares, T.S.; Americo, M.F. Effect of Bauhinia holophylla treatment in streptozotocin-induced diabetic rats. An. Acad. Bras. Cienc. 2017, 89, 263–272. [Google Scholar] [CrossRef]
- Camaforte, N.A.D.P.; Saldanha, L.L.; Vareda, P.M.P.; Rezende-Neto, J.M.; Senger, M.R.; Delgado, A.Q.; Morgan, H.J.N.; Violato, N.M.; Pieroni, L.G.; Dokkedal, A.L.; et al. Hypoglycaemic activity of Bauhinia holophylla through GSK3-β inhibition and glycogenesis activation. Pharm. Biol. 2019, 57, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Saldanha, L.L.; Delgado, A.Q.; Marcourt, L.; Camaforte, N.A.P.; Vareda, P.M.P.; Ebrahimi, S.N.; Vilegas, W.; Dokkedal, A.L.; Queiroz, E.F.; Wolfender, J.; et al. Hypoglycemic active principles from the leaves of Bauhinia holophylla: Comprehensive phytochemical characterization and in vivo activity profile. PLoS ONE 2021, 16, e0258016. [Google Scholar] [CrossRef]
- Rozza, A.L.; Cesar, D.A.S.; Pieroni, L.G.; Saldanha, L.L.; Dokkedal, A.L.; De-Faria, F.M.; Souza-Brito, A.R.M.; Vilegas, W.; Takahira, R.K.; Pellizzon, C.H. Antiulcerogenic activity and toxicity of Bauhinia holophylla hydroalcoholic extract. Evid. Based Complement. Altern. Med. 2015, 2015, 439506. [Google Scholar] [CrossRef]
- dos Santos, M.; Teixeira, T.R.; Santos, F.R.S.; Lima, W.G.; Ferraz, A.C.; Silva, N.L.; Leite, F.J.; Siqueira, J.M.; Luyten, W.; Castro, A.H.F.; et al. Bauhinia holophylla (Bong.) Steud. leaves-derived extracts as potent anti-dengue serotype 2. Nat. Prod. Res. 2021, 35, 2804–2809. [Google Scholar] [CrossRef]
- Ribeiro, D.L.; Cilião, H.L.; Specian, A.F.L.; Serpeloni, J.M.; de Oliveira, M.T.; Varanda, E.A.; Vilegas, W.; Saldanha, L.L.; Martínez-López, W.; Dokkedal, A.L.; et al. Phytochemical study and evaluation of cytotoxicity, mutagenicity, cell cycle kinetics and gene expression of Bauhinia holophylla (Bong.) Steud. in HepG2 cells in vitro. Cytotechnology 2018, 70, 713–728. [Google Scholar] [CrossRef] [Green Version]
- Marena, G.D.; Saldanha, L.L.; Ramos, M.A.S.; de Grandis, R.A.; Dokkedal, A.L.; Bauab, T.M.; Pavan, F.R.; Resende, F.A. Antimicrobial, cytotoxic and mutagenic activities of Bauhinia holophylla hydroalcoholic extract. Am. J. Essent. Oil. Nat. Prod. 2021, 9, 1–6. [Google Scholar]
- Filho, V.C. Chemical Composition and biological potential of plants from the genus Bauhinia. Phytother. Res. 2009, 23, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Andersen, O.M.; Markham, K.R. Flavonoids: Chemistry, Biochemistry and Applications, 1st ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 617–856. ISBN 0-8493-2021-6. [Google Scholar]
- Ivanov, M.; Kannan, A.; Stojkovic, D.S.; Glamoclija, J.; Calhelha, R.C.; Ferreira, I.C.F.R.; Sanglard, D.; Sokovic, M. Flavones, flavonols, and glycosylated derivatives-impact on Candida albicans growth and virulence, expression of CDR1 and ERG11, cytotoxicity. Pharmaceuticals 2020, 14, 27. [Google Scholar] [CrossRef]
- Ansari, M.A.; Fatima, Z.; Hameed, S. Sesamol: A natural phenolic compound with promising anticandidal potential. J. Pathog. 2014, 2014, 895193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miceli, N.; Buongiorno, L.P.; Celi, M.G.; Cacciola, F.; Dugo, P.; Donato, P.; Mondello, L.; Bonaccorsi, I.; Taviano, M.F. Role of the flavonoid-rich fraction in the antioxidant and cytotoxic activities of Bauhinia forficata Link. (Fabaceae) leaves extract. Nat. Prod. Res. 2015, 30, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, S.T.D.; Teixeira, T.R.; Ferreira, J.M.S.; Lima, L.A.R.S.; Siqueira, J.M.; Luyten, W.; Castro, A.H.F. Phenolic compounds and antioxidant and antibacterial activities of Bauhinia holophylla (Fabaceae:Cercideae). In Phenolic Compounds in Health and Disease, 1st ed.; Linn, R.C., Ed.; Nova Science Publishers: New York, NY, USA, 2021; pp. 153–173. ISBN 978-1-68507-137-0. [Google Scholar]
- Rochefort, S.J.; Imsic, M.; Jones, R.; Trenerry, V.C.; Thomkins, B. Characterization of flavonol conjugates in immature leaves of pak choi [Brassica rapa L. sp. chinensis L. (Hanelt.)] by HPLC- DAD and LC-MS/MS. J. Agric. Food. Chem. 2006, 54, 4655–4860. [Google Scholar] [CrossRef]
- Mishra, B.; Leishangthem, G.D.; Gill, K.; Singh, A.K.; Das, S.; Singh, K.; Xess, I.; Dinda, A.; Kapil, A.; Patro, I.K.; et al. A novel antimicrobial peptide derived from modified N-terminal domain of bovine lactoferrin: Design, synthesis, activity against multidrug-resistant bacteria and Candida. Biochim. Biophys. Acta. Biomembr. 2013, 1828, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Smiljković, M.; Kostić, M.; Stojković, D.; Glamočlija, J.; Soković, M. Could flavonoids compete with synthetic azoles in diminishing Candida albicans infections? A comparative review based on in vitro studies. Curr. Med. Chem. 2019, 26, 2536–2554. [Google Scholar] [CrossRef]
- Wall, G.; Chaturvedi, A.K.; Wormley, F.L.; Wiederhold, N.P.; Patterson, H.P.; Patterson, T.F.; Lopez-Ribot, J.L. Screening a repurposing library for inhibitors of multidrug-resistant Candida auris identifies ebselen as a repositionable candidate for antifungal drug development. Antimicrob. Agents Chemother. 2018, 62, e01084-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobile, C.J.; Johnson, A.D. Candida albicans biofilms and human disease. Annu. Ver. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [Green Version]
- Patel, M.; Srivastava, V.; Ahmad, A. Dodonaea viscosa var angustifolia derived 5,6,8-trihydroxy-7,4′dimethoxy flavone inhibits ergosterol synthesis and the production of hyphae and biofilm in Candida albicans. J. Ethnopharmacol. 2020, 259, 112965. [Google Scholar] [CrossRef]
- Lewis, K. Persister cells, dormancy and infectious disease. Nat. Ver. Microbiol. 2007, 5, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.N.; Upreti, D.K.; Singh, B.R.; Pandey, G.; Verma, S.; Roy, S.; Naqvi, A.H.; Rawat, A.K.S. Quercetin sensitizes fluconazole-resistant Candida albicans to induce apoptotic cell death by modulating quorum sensing. Antimicrob. Agents Chemother. 2015, 59, 2153–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, E.P.; Lima, R.F.; de Almeida, C.M.; Freires, I.A.; Rosalen, P.L.; Ruiz, A.L.; Granville-Garcia, A.F.; Godoy, G.P.; Pereira, J.V.; Costa, E.M.M.B. Antimicrobial and antiproliferative activity of Bauhinia forficata Link and Cnidoscolus quercifolius extracts commonly used in folk medicine. J. Contemp. Dent. Pract. 2017, 18, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Zhang, M.; Wang, T.; Li, Y.; Wang, C. The roles of CDR1, CDR2, and MDR1 in kaempferol-induced suppression with fluconazole-resistant Candida albicans. Pharm. Biol. 2016, 54, 984–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semwal, D.; Semwal, R.; Combrinck, S.; Viljoen, A. Myricetin: A dietary molecule with diverse biological activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [Green Version]
- Imran, M.; Rauf, A.; Shah, Z.A.; Saeed, F.; Imran, A.; Arshad, M.U.; Ahmad, B.; Bawazeer, S.; Atif, M.; Peters, D.G.; et al. Chemo-preventive and therapeutic effect of the dietary flavonoid kaempferol: A comprehensive review. Phytother. Res. 2018, 33, 263–275. [Google Scholar] [CrossRef]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; Mahdy, N.M.E.; Kiliç, C.S.; Sytar, O.; et al. Therapeutic potential of quercetin: New insights and perspectives for human health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef]
- Gao, M.; Wang, H.; Zhu, L. Quercetin assists fluconazole to inhibit biofilm formations of fluconazole-resistant Candida albicans in in vitro and in vivo antifungal managements of vulvovaginal candidiasis. Cell. Physiol. Biochem. 2016, 40, 727–742. [Google Scholar] [CrossRef]
- Song, X.; Tan, L.; Wang, M.; Ren, C.; Guo, C.; Yang, B.; Ren, Y.; Cao, Z.; Li, Y.; Pei, J. Myricetin: A review of the most recent research. Biomed. Pharmacother. 2021, 134, 111017. [Google Scholar] [CrossRef]
- Mo, F.; Ma, J.; Yang, X.; Zhang, P.; Li, Q.; Zhang, J. In vitro and in vivo effects of the combination of myricetin and miconazole nitrate incorporated to thermosensitive hydrogels, on C. albicans biofilms. Phytomedicine 2020, 71, 153223. [Google Scholar] [CrossRef]
- Mishra, S.; Rastogi, S.K.; Singh, S.; Panwar, S.L.; Shrivash, M.K.; Misra, K. Controlling pathogenesis in Candida albicans by targeting EFG1 and glyoxylate pathway through naturally occurring polyphenols. Mol. Biol. Rep. 2019, 46, 5805–5820. [Google Scholar] [CrossRef] [PubMed]
- Pastrana-Bonilla, E.; Akoh, C.C.; Sellappan, S.; Krewer, G. Phenolic content and antioxidant capacity of muscadine grapes. J. Agric. Food Chem. 2003, 51, 5497–5503. [Google Scholar] [CrossRef] [PubMed]
- Woisky, R.G.; Salatino, A. Analysis of propolis: Some parameters and procedures for chemical quality control. J. Apic. Res. 1998, 37, 99–105. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standarts Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 90th ed.; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2012; CLSI, Document M07-A9. [Google Scholar]
- Clinical and Laboratory Standarts Institute (CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, NJ, USA, 2008; CLSI, Document M27-A3. [Google Scholar]
- Xu, K.; Wang, J.L.; Chu, M.P.; Jia, C. Activity of coumarin against Candida albicans biofilms. J. Mycol. Med. 2018, 29, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhang, N.; Zhang, S.; Zhang, L.; Liu, Q. Phloretin inhibited the pathogenicity and virulence factors against Candida albicans. Bioengineered 2021, 12, 2420–2431. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.T.; Santos, F.R.S.; Lima, W.G.; Sousa, C.D.F.; Oliveira, L.S.F.M.; Ribeiro, R.I.M.A.; Gomes, A.J.P.S.; Araújo, M.G.F.; Villar, J.A.F.P.; Ferreira, J.M.S. Design, synthesis, biological activity and structure-activity relationship studies of chalcone derivatives as potential anti-Candida agentes. J. Antibiot. 2018, 71, 702–712. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Phenols (µg GAEq mg−1 DS) | Total Flavonoids (µg QEq mg−1 DS) |
|---|---|---|
| CHE | 39.08 ± 1.77 b | 3.91 ± 0.13 c |
| DCM | 37.61 ± 2.54 b | 9.93 ± 0.53 a |
| EtOAc | 47.45 ± 1.51 a | 6.03 ± 0.35 b |
| EtOH | 20.46 ± 2.32 c | 2.37 ± 0.13 c |
| Peak | Rt | UVmax | [M + H]+ (m/z) | [M + H]+ (m/z) | Error | MS/MSn | Compound | Samples | Molecular Formula | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (min.) | (nm) | Experimental | Theoretical | (ppm) | CHE | DCM | EtOAc | EtOH | ||||
| 1 | 12.0 | 254/350 | 451.0855 | 451.0871 | 3.5 | 319.0412; 303.0472 | Myricetin-O-pentoside | + | + | + | - | C20H18O12 |
| 2 | 12.3 | 255/348 | 465.1013 | 465.1028 | 3.2 | 319.0441; 303.0511 | Myricetin-O-deoxyhexoside | + | + | + | - | C21H20O12 |
| 3 | 13.0 | 254/353 | 465.1004 | 465.1028 | 5.2 | 303.0496 | Quercetin-O-hexoside | + | + | + | + | C21H20O12 |
| 4 | 13.8 | 254/350 | 465.1024 | 465.1028 | 0.9 | 303.0503 | Quercetin-O-hexoside | + | + | + | - | C21H20O12 |
| 5 | 15.0 | 255/350 | 435.0900 | 435.0922 | 5.0 | 303.0493 | Quercetin-O-xilopyranose | + | + | + | + | C20H18O11 |
| 6 | 15.6 | 255/349 | 449.1062 | 449.1078 | 3.6 | 287.0559 | Kaempferol-3-O-glucoside | + | + | + | + | C21H20O11 |
| 7 | 15.9 | 255/348 | 435.0901 | 435.0922 | 4.8 | 303.0499 | Quercetin-O-pentoside | + | + | + | + | C20H18O11 |
| 8 | 16.5 | 254/348 | 449.1056 | 449.1078 | 4.9 | 303.0470 | Quercetin-O-deoxyhexoside | + | + | + | + | C21H20O11 |
| 9 | 17.7 | 264/346 | 419.0959 | 419.0973 | 3.3 | 287.0505 | Kaempferol-O-pentoside | - | + | - | - | C20H18O10 |
| 10 | 18.1 | 264/346 | 479.1168 | 479.1184 | 3.3 | 317.0624; 302.0381 | Isorhamnetin-3-O-hexoside | + | + | + | - | C22H22O12 |
| 11 | 18.4 | 264/348 | 479.0938 | - | - | 317.0619 | Undentified | + | + | + | - | NI |
| 12 | 18.7 | 264/345 | 433.1108 | 433.1129 | 4.8 | 287.0517 | Luteolin-deoxyhexose | + | + | + | - | C21H20O10 |
| 13 | 20.8 | 254/362 | 303.0493 | 303.0499 | 2.0 | 181.9561 | Quercetin | + | + | + | - | C15H10O7 |
| 14 | 22.0 | 254/356 | 317.0645 | 317.0656 | 3.5 | 302.0408 | Isorhamnetin | + | + | + | - | C16H12O7 |
| Candida albicans | MIC (µg/mL) | Drugs | ||||
|---|---|---|---|---|---|---|
| CHE | DCM | EtOAc | EtOH | FLU | NYS | |
| SC5314 | >1250 c | >1250 c | >1250 c | >1250 c | 1.95 a | - |
| ATCC 18804 | >1250 c | >1250 c | >1250 c | >1250 c | 1.95 a | - |
| ATCC 10231 | >1250 c | >1250 c | >1250 c | >1250 c | - | 3.91 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Fonseca, S.T.D.; Teixeira, T.R.; Ferreira, J.M.S.; Lima, L.A.R.d.S.; Luyten, W.; Castro, A.H.F. Flavonoid-Rich Fractions of Bauhinia holophylla Leaves Inhibit Candida albicans Biofilm Formation and Hyphae Growth. Plants 2022, 11, 1796. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11141796
da Fonseca STD, Teixeira TR, Ferreira JMS, Lima LARdS, Luyten W, Castro AHF. Flavonoid-Rich Fractions of Bauhinia holophylla Leaves Inhibit Candida albicans Biofilm Formation and Hyphae Growth. Plants. 2022; 11(14):1796. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11141796
Chicago/Turabian Styleda Fonseca, Sara Thamires Dias, Thaiz Rodrigues Teixeira, Jaqueline Maria Siqueira Ferreira, Luciana Alves Rodrigues dos Santos Lima, Walter Luyten, and Ana Hortência Fonsêca Castro. 2022. "Flavonoid-Rich Fractions of Bauhinia holophylla Leaves Inhibit Candida albicans Biofilm Formation and Hyphae Growth" Plants 11, no. 14: 1796. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11141796