
Nutritive and Phytochemical Composition of Aromatic Microgreen Herbs and Spices Belonging to the Apiaceae Family
 , ,
, ,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results and Discussion
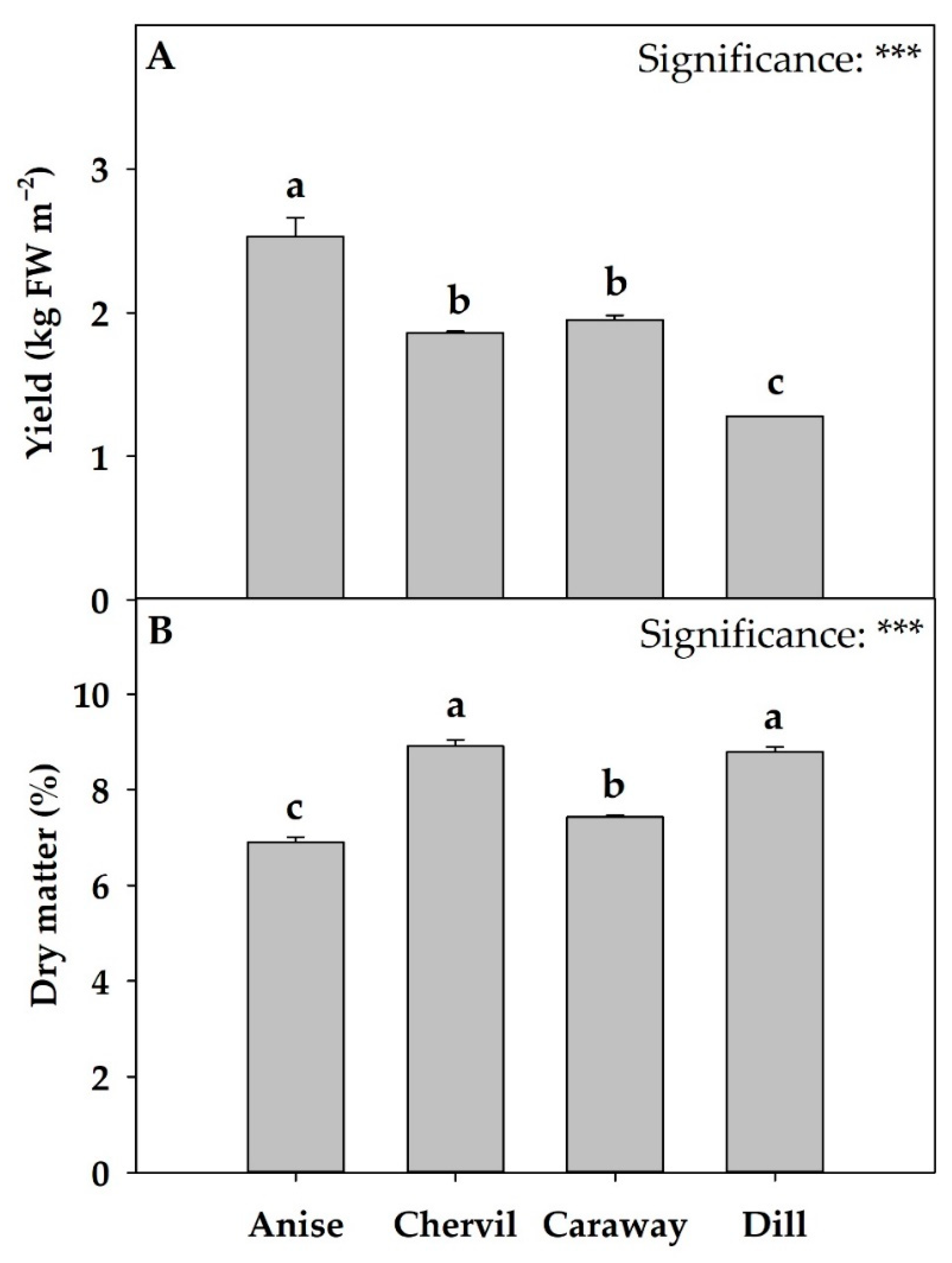
2.1. Growth and Yield
2.2. Macro- and Microminerals
2.3. Chlorophylls, Carotenoids, and Total Ascorbic Acid
2.4. Phenolic Profile
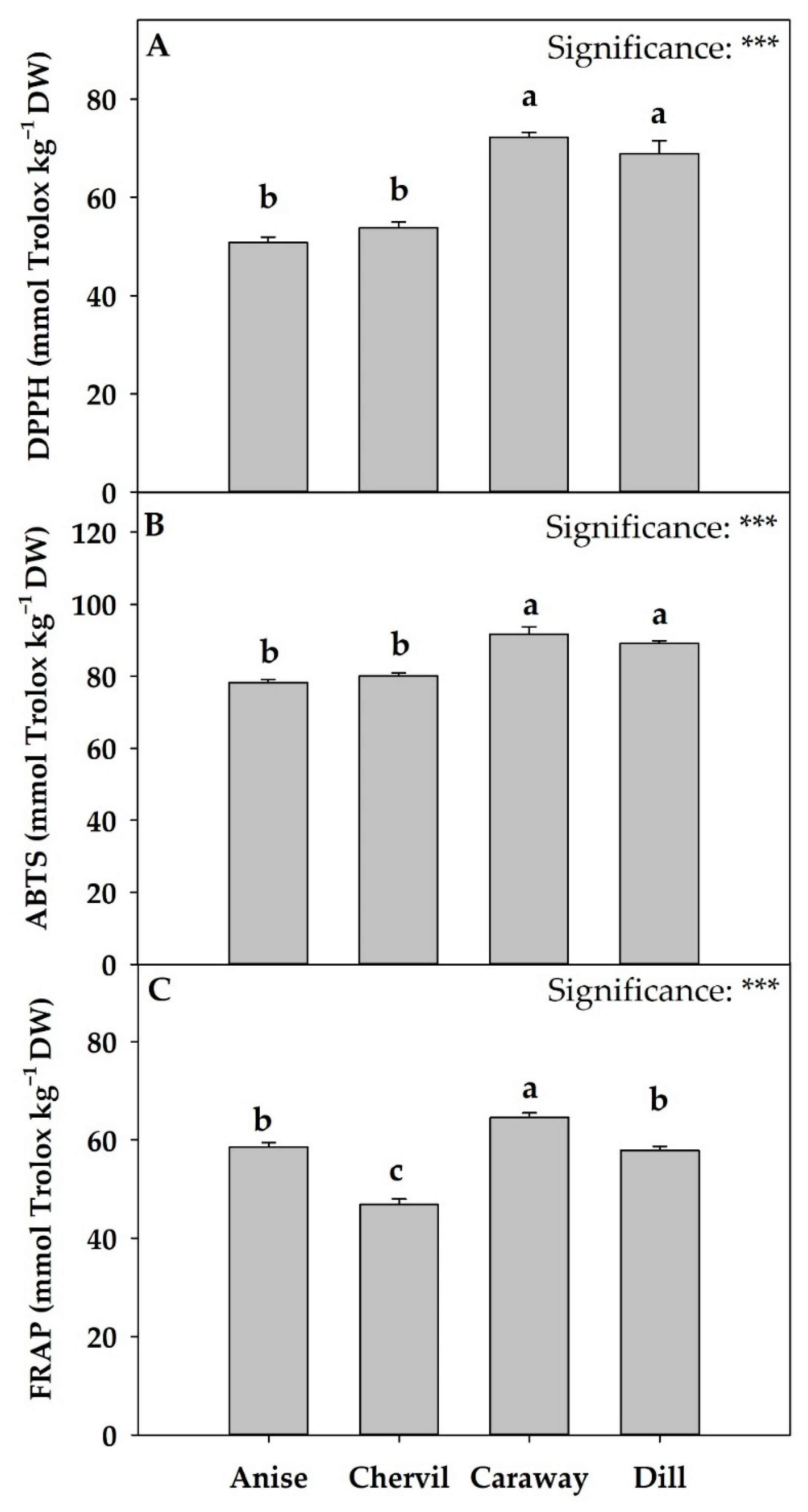
2.5. Antioxidant Activity
3. Materials and Methods
3.1. Plant Material and Growth Condition
3.2. Sampling of Microgreens, Fresh Weight and Dry Matter Determination
3.3. Macro- and Micromineral Analysis by ICP-OES
3.4. Analysis of Chlorophylls, Carotenoids, and Total Ascorbic Acid
3.5. Phenolic Profile by UHPLC-Q-Orbitrap HRMS
3.6. Antioxidant Activity
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Genotype-specific modulatory effects of select spectral bandwidths on the nutritive and phytochemical composition of microgreens. Front. Plant Sci. 2019, 10, 1501. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, F.; Renna, M.; Santamaria, P. Sprouts, microgreens and “baby leaf” vegetables. In Minimally Processed Refrigerated Fruits and Vegetables; Yildiz, F., Wiley, R.C., Eds.; Springer: Boston, MA, USA, 2017; pp. 403–432. [Google Scholar]
- Caracciolo, F.; El-Nakhel, C.; Raimondo, M.; Kyriacou, M.C.; Cembalo, L.; De Pascale, S.; Rouphael, Y. Sensory attributes and consumer acceptability of 12 microgreens species. Agronomy 2020, 10, 1043. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Variation in macronutrient content, phytochemical constitution and in vitro antioxidant capacity of green and red Butterhead lettuce dictated by different developmental stages of harvest maturity. Antioxidants 2020, 9, 300. [Google Scholar] [CrossRef] [Green Version]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-scale vegetable production and the rise of microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Spinozzi, E.; Maggi, F.; Bonacucina, G.; Pavela, R.; Boukouvala, M.C.; Kavallieratos, N.G.; Canale, A.; Romano, D.; Desneux, N.; Wilke, A.; et al. Apiaceae essential oils and their constituents as insecticides against mosquitoes—A review. Crops Prod. 2021, 171, 113892. [Google Scholar] [CrossRef]
- Shahamat, Z.; Abbasi-maleki, S.; Motamed, S.M. Evaluation of antidepressant-like effects of aqueous and ethanolic extracts of Pimpinella anisum fruit in mice. Avicenna J. Phytomed. 2016, 6, 322–328. [Google Scholar]
- Es-safi, I.; Mechchate, H.; Amaghnouje, A.; Elbouzidi, A.; Bouhrim, M. Assessment of antidepressant-like, anxiolytic effects and impact on memory of Pimpinella anisum L. Total extract on Swiss albino mice. Plants 2021, 10, 1573. [Google Scholar] [CrossRef]
- Vyas, A.; Shukla, S.; Pandey, R.; Jain, V.; Joshi, V.; Gidwani, B. Chervil: A multifunctional miraculous nutritional herb. Asian J. Plant Sci. 2012, 11, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Hajlaoui, H.; Arraouadi, S.; Noumi, E.; Aouadi, K.; Adnan, M.; Khan, M.A.; Kadri, A.; Snoussi, M. Antimicrobial, antioxidant, anti-acetylcholinesterase, antidiabetic, and pharmacokinetic properties of Carum carvi L. and Coriandrum sativum L. Essential oils alone and in combination. Molecules 2021, 26, 3625. [Google Scholar] [CrossRef]
- Kabiri, M.; Kamalinejad, M.; Sohrabvand, F.; Bioos, S.; Babaeian, M. Management of breast milk oversupply in traditional persian medicine. J. Evid.-Based Complement. Altern. Med. 2017, 22, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Mahboubi, M. Caraway as important medicinal plants in management of diseases. Nat. Prod. Bioprospecting 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozliman, S.; Yaldiz, G.; Camlica, M.; Ozsoy, N. Chemical components of essential oils and biological activities of the aqueous extract of Anethum graveolens L. grown under inorganic and organic conditions. Chem. Biol. Technol. Agric. 2021, 8, 20. [Google Scholar] [CrossRef]
- Xiao, Z.; Codling, E.E.; Luo, Y.; Nou, X.; Lester, G.E.; Wang, Q. Microgreens of Brassicaceae: Mineral composition and content of 30 varieties. J. Food Comp. Anal. 2016, 49, 87–93. [Google Scholar] [CrossRef] [Green Version]
- AbdElgawad, H.; Okla, M.K.; Al-Amri, S.S.; Al-Hashimi, A.; Al-Qahtani, W.H.; Al-Qahtani, S.M.; Abbas, Z.K.; Al-Harbi, N.A.; Algafar, A.A.; Almuhayawi, M.S.; et al. Effect of elevated CO2 on biomolecules’ accumulation in caraway (Carum carvi L.) plants at different developmental stages. Plants 2021, 10, 2434. [Google Scholar] [CrossRef]
- Okla, M.K.; Abdel-Mawgoud, M.; Alamri, S.A.; Abbas, Z.K.; Al-Qahtani, W.H.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Hassan, A.H.A.; Selim, S.; Alruhaili, M.H.; et al. Developmental stages-specific response of Anise plants to laser-induced growth, nutrients accumulation, and essential oil metabolism. Plants 2021, 10, 2591. [Google Scholar] [CrossRef]
- Balanescu, F.; Busuioc, A.C.; Botezatu, A.V.D.; Gosav, S.; Avramescu, S.M.; Furdui, B.; Dinica, R.M. Comparative study of natural antioxidants from Glycine max, Anethum graveolensand, Pimpinella anisum seed and sprout extracts obtained by ultrasound-assisted extraction. Separations 2022, 9, 152. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Giordano, M.; Kyriacou, M.C.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Mineral and antioxidant attributes of Petroselinum crispum at different stages of ontogeny: Microgreens vs. baby greens. Agronomy 2021, 11, 857. [Google Scholar] [CrossRef]
- Ghoora, M.D.; Babu, D.R.; Srividya, N. Nutrient composition, oxalate content and nutritional ranking of ten culinary microgreens. J. Food Comp. Anal. 2020, 91, 103495. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; El-Nakhel, C.; Pannico, A.; Graziani, G.; Soteriou, G.A.; Giordano, M.; Palladino, M.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Phenolic constitution, phytochemical and macronutrient content in three species of microgreens as modulated by natural fiber and synthetic substrates. Antioxidants 2020, 9, 252. [Google Scholar] [CrossRef] [Green Version]
- Di Gioia, F.; De Bellis, P.; Mininni, C.; Santamaria, P.; Serio, F. Physicochemical, agronomical and microbiological evaluation of alternative growing media for the production of rapini (Brassica rapa L.) microgreens. J. Sci. Food Agric. 2017, 97, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- El-Nakhel, C.; Ciriello, M.; Formisano, L.; Pannico, A.; Giordano, M.; Gentile, B.R.; Fusco, G.M.; Kyriacou, M.C.; Carillo, P.; Rouphael, Y. Protein hydrolysate combined with hydroponics divergently modifies growth and shuffles pigments and free amino acids of carrot and dill microgreens. Horticulturae 2021, 7, 279. [Google Scholar] [CrossRef]
- Pannico, A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Soteriou, G.A.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Selenium biofortification impacts the nutritive value, polyphenolic content, and bioactive constitution of variable microgreens genotypes. Antioxidants 2020, 9, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.J.; Jeong, D.W.; Lee, J.G.; Lee, S.; Jin, Y.; Yi, Y.H.; Cho, Y.H.; Im, S.J.; Bae, M.J. The impact of bladder distension on blood pressure in middle aged women. Korean J. Fam. Med. 2011, 32, 306–310. [Google Scholar] [CrossRef]
- Pinto, E.; Almeida, A.A.; Aguiar, A.A.; Ferreira, I.M.P.L.V.O. Comparison between the mineral profile and nitrate content of microgreens and mature lettuces. J. Food Comp. Anal. 2015, 37, 38–43. [Google Scholar] [CrossRef]
- Kusumitha, V.N.; Rajasree, V.; Swarnapriya, R.; Uma, D.; Meenakshi, P. Nutrient availability of selected leafy vegetables at micro green stage grown in vertical gardening. J. Pharmacogn. Phytochem. 2021, 10, 2226–2228. [Google Scholar]
- Rayman, M.P. The argument for increasing selenium intake. Proc. Nutr. Soc. 2002, 61, 203–215. [Google Scholar] [CrossRef]
- Szerement, J.; Szatanik, A.; Jakub, K.; Monika, M.; Hersztek, M. Agronomic biofortification with Se, Zn, and Fe: An effective strategy to enhance crop nutritional quality and stress defense—A review. J. Soil Sci. Plant Nutr. 2022, 22, 1129–1159. [Google Scholar] [CrossRef]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; Mcgrath, S.P.; Seneviratne, S.; Smith, P.; Winkel, L.H.E. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2848–2853. [Google Scholar] [CrossRef] [Green Version]
- Pannico, A.; Graziani, G.; El-Nakhel, C.; Giordano, M.; Ritieni, A.; Kyriacou, M.C.; Rouphael, Y. Nutritional stress suppresses nitrate content and positively impacts ascorbic acid concentration and phenolic acids profile of lettuce microgreens. Italus Hortus 2020, 27, 41–52. [Google Scholar] [CrossRef]
- Xiao, Z.; Lester, G.E.; Park, E.; Saftner, R.A.; Luo, Y.; Wang, Q. Evaluation and correlation of sensory attributes and chemical compositions of emerging fresh produce: Microgreens. Postharvest Biol. Technol. 2015, 110, 140–148. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Rouphael, Y. The effects of nutrient solution feeding regime on yield, mineral profile, and phytochemical composition of spinach microgreens. Horticulturae 2021, 7, 162. [Google Scholar] [CrossRef]
- Barros, L.; Dueňas, M.; Dias, M.I.; Sousa, M.J.; Santos-Buelga, C.; Ferreira, I.C. Phenolic profiles of in vivo and in vitro grown Coriandrum sativum L. Food Chem. 2012, 132, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Harakotr, B.; Srijunteuk, S.; Rithichai, P.; Tabunhan, S. Effects of light-emitting diode light irradiance levels on yield, antioxidants and antioxidant capacities of indigenous vegetable microgreens. Sci. Technol. Asia 2019, 24, 59–66. [Google Scholar]
- Jana, S.; Shekhawat, G. Phytochemical analysis and antibacterial screening of in vivo and in vitro extracts of indian medicinal herb: Anethum graveolens. Res. J. Med. Plant 2010, 4, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Petropoulos, S.; Fernandes, Â.; Calhelha, R.C.; Di Gioia, F.; Kolovou, P.; Barros, L.; Ferreira, I.C.F.R. Chemical composition and bioactive properties of Cichorium spinosum L. in relation to nitrate/ammonium nitrogen ratio. J. Food Sci. Agric. 2019, 99, 6741–6750. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; El-Nakhel, C.; De Micco, V.; Giordano, M.; Pannico, A.; De Pascale, S.; Graziani, G.; Ritieni, A.; Soteriou, G.; Kyriacou, M.; et al. Morpho-metric and specialized metabolites modulation of parsley microgreens through selective LED wavebands. Agronomy 2022, 12, 1502. [Google Scholar] [CrossRef]
- Volpe, M.G.; Nazzaro, M.; Di Stasio, M.; Siano, F.; Coppola, R.; De Marco, A. Content of micronutrients, mineral and trace elements in some Mediterranean spontaneous edible herbs. Chem. Cent. J. 2015, 9, 17–19. [Google Scholar] [CrossRef] [Green Version]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Kim, H.-J.; Fonseca, J.; Choi, J.-H.; Kubota, C.; Kwon, D. Salt in irrigation water affects the nutritional and visual properties of romaine lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Montagu, M.; Inzé, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.F.; Straint, J.J. The ferric reducing ability of plasma (FRAP) as a measure of ‘Antioxidant Power’: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 9–10. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | P | K | Ca | Mg | Na |
|---|---|---|---|---|---|
| mg g−1 DW | |||||
| Anise | 10.9 ± 0.31 a | 27.7 ± 0.54 ab | 12.6 ± 0.50 b | 2.28 ± 0.11 a | 0.18 ± 0.02 d |
| Chervil | 9.27 ± 0.38 ab | 23.8 ± 0.63 bc | 13.0 ± 0.25 b | 2.24 ± 0.04 a | 0.76 ± 0.02 b |
| Caraway | 10.1 ± 0.64 a | 29.9 ± 1.00 a | 9.6 ± 0.10 c | 1.88 ± 0.05 b | 0.31 ± 0.01 c |
| Dill | 7.17 ± 0.23 b | 21.7 ± 1.00 c | 18.0 ± 0.71 a | 1.81 ± 0.08 b | 2.91 ± 0.04 a |
| Significance | *** | ** | *** | ** | *** |
| Species | Fe | Mn | B | Zn | Se |
|---|---|---|---|---|---|
| µg g−1 DW | |||||
| Anise | 57.0 ± 1.8 a | 37.6 ± 0.7 c | 25.3 ± 0.8 ab | 47.5 ± 0.5 a | 1.75 ± 0.16 |
| Chervil | 45.6 ± 1.8 b | 78.5 ± 0.5 a | 24.8 ± 0.2 bc | 47.0 ± 0.9 a | 1.40 ± 0.07 |
| Caraway | 59.9 ± 0.7 a | 43.8 ± 0.8 b | 21.7 ± 0.4 c | 32.2 ± 1.4 b | 1.44 ± 0.15 |
| Dill | 34.5 ± 1.7 c | 40.1 ± 0.8 bc | 28.3 ± 1.2 a | 27.0 ± 0.5 c | 1.47 ± 0.14 |
| Significance | *** | *** | ** | *** | ns |
| Species | Total Chlorophylls | Lutein | β-Carotene | TAA |
|---|---|---|---|---|
| mg g−1 FW | µg g−1 DW | mg AA 100 g−1 FW | ||
| Anise | 1.10 ± 0.08 | 18.4 ± 0.8 a | 172 ± 1.8 a | 94.5 ± 2.1 b |
| Chervil | 1.01 ± 0.04 | 13.1 ± 0.5 b | 170 ± 0.7 a | 152 ± 1.4 a |
| Caraway | 1.05 ± 0.03 | 13.9 ± 1.1 b | 179 ± 1.5 a | 71.6 ± 0.9 c |
| Dill | 0.99 ± 0.04 | 8.81 ± 0.2 c | 126 ± 2.7 b | 150 ± 2.4 a |
| Significance | ns | *** | *** | *** |
| Phenolic Compounds (μg g−1 DW) | Species | Significance | |||
|---|---|---|---|---|---|
| Anise | Chervil | Caraway | Dill | ||
| PHENOLIC ACID DERIVATIVES | |||||
| salicylic acid hexoside | 150.4 ± 1.0 a | 36.8 ± 0.3 c | 44.3 ± 0.8 b | 16.9 ± 1.5 d | *** |
| caffeic acid hexoside | 4.59 ± 0.21 a | 4.81 ± 0.05 a | 4.21 ± 0.37 a | 1.42 ± 0.10 b | *** |
| dihydroferulic acid | 71.4 ± 1.2 a | 5.75 ± 0.08 b | 3.25 ± 0.21 b | 4.87 ± 0.28 b | *** |
| sinapinic acid hexose | 4.18 ± 0.04 d | 12.9 ± 0.59 b | 7.54 ± 0.64 c | 21.6 ± 0.32 a | *** |
| feruloyl quinic acid | 2937 ± 0.5 b | 491 ± 0.3 d | 1761 ± 0.88 c | 2977 ± 2.3 a | *** |
| caffeoyl quinic acid | 7040 ± 1.2 b | 948 ± 1.2 d | 5447 ± 1.1 c | 7995 ± 3.5 a | *** |
| dicaffeoyl quinic acid | 950.9 ± 0.9 b | 396.7 ± 3.6 c | 3168.1 ± 2.62 a | 61.2 ± 0.92 d | *** |
| caffeic acid | 1.119 ± 0.140 | 1.228 ± 0.050 | 0.961 ± 0.027 | 1.234 ± 0.080 | ns |
| caffeoyl shikimate acid | 6.78 ± 0.26 a | 0.19 ± 0.01 c | 0.64 ± 0.06 bc | 0.85 ± 0.06 b | *** |
| coumaroyl diglucoside | 3.01 ± 0.11 a | 0.64 ± 0.09 b | 0.22 ± 0.03 c | 0.28 ± 0.08 bc | *** |
| ferulic acid | 42.6 ± 1.4 b | 68.7 ± 0.7 a | 17.8 ± 0.7 c | 44.4 ± 0.7 b | *** |
| coumaric acid | 7.29 ± 0.53 a | 1.30 ± 0.17 c | 3.77 ± 0.64 b | 1.08 ± 0.08 c | *** |
| Total phenolic acid derivatives | 11,218.50 ± 1.96 a | 1967.82 ± 3.61 d | 10,459.28 ± 1.95 c | 11,126.23 ± 6.11 b | *** |
| FLAVONOID DERIVATIVES | |||||
| rutin | 32.7 ± 1.09 b | 0.28 ± 0.01 c | 1.13 ± 0.05 c | 49.8 ± 0.81 a | *** |
| hyperoside | 27.5 ± 0.94 b | 0.19 ± 0.02 d | 7.87 ± 0.44 c | 56.4 ± 0.90 a | *** |
| luteolin-7-O-glucoside | 35.9 ± 0.53 a | 0.25 ± 0.00 b | 0.52 ± 0.00 b | 0.29 ± 0.00 b | *** |
| quercetin rhamnoside | 125 ± 1.8 a | 1.61 ± 0.02 b | 1.92 ± 0.02 b | 1.82 ± 0.08 b | *** |
| kaempferol-3-dihexoside | 10.3 ± 0.83 b | 0.10 ± 0.04 c | 0.31 ± 0.05 c | 14.5 ± 0.31 a | *** |
| Total flavonoid derivatives | 231.57 ± 4.53 a | 2.44 ± 0.07 c | 11.77 ± 0.47 c | 122.85 ± 1.46 b | *** |
| TOTAL POLYPHENOLS | 11,450.07 ± 3.01 a | 1970.26 ± 3.65 d | 10,471.05 ± 2.13 c | 11,249.10 ± 6.07 b | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giordano, M.; Petropoulos, S.A.; Kyriacou, M.C.; Graziani, G.; Zarrelli, A.; Rouphael, Y.; El-Nakhel, C. Nutritive and Phytochemical Composition of Aromatic Microgreen Herbs and Spices Belonging to the Apiaceae Family. Plants 2022, 11, 3057. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11223057
Giordano M, Petropoulos SA, Kyriacou MC, Graziani G, Zarrelli A, Rouphael Y, El-Nakhel C. Nutritive and Phytochemical Composition of Aromatic Microgreen Herbs and Spices Belonging to the Apiaceae Family. Plants. 2022; 11(22):3057. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11223057
Chicago/Turabian StyleGiordano, Maria, Spyridon A. Petropoulos, Marios C. Kyriacou, Giulia Graziani, Armando Zarrelli, Youssef Rouphael, and Christophe El-Nakhel. 2022. "Nutritive and Phytochemical Composition of Aromatic Microgreen Herbs and Spices Belonging to the Apiaceae Family" Plants 11, no. 22: 3057. https://0-doi-org.brum.beds.ac.uk/10.3390/plants11223057