NTRC and Thioredoxin f Overexpression Differentially Induces Starch Accumulation in Tobacco Leaves

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
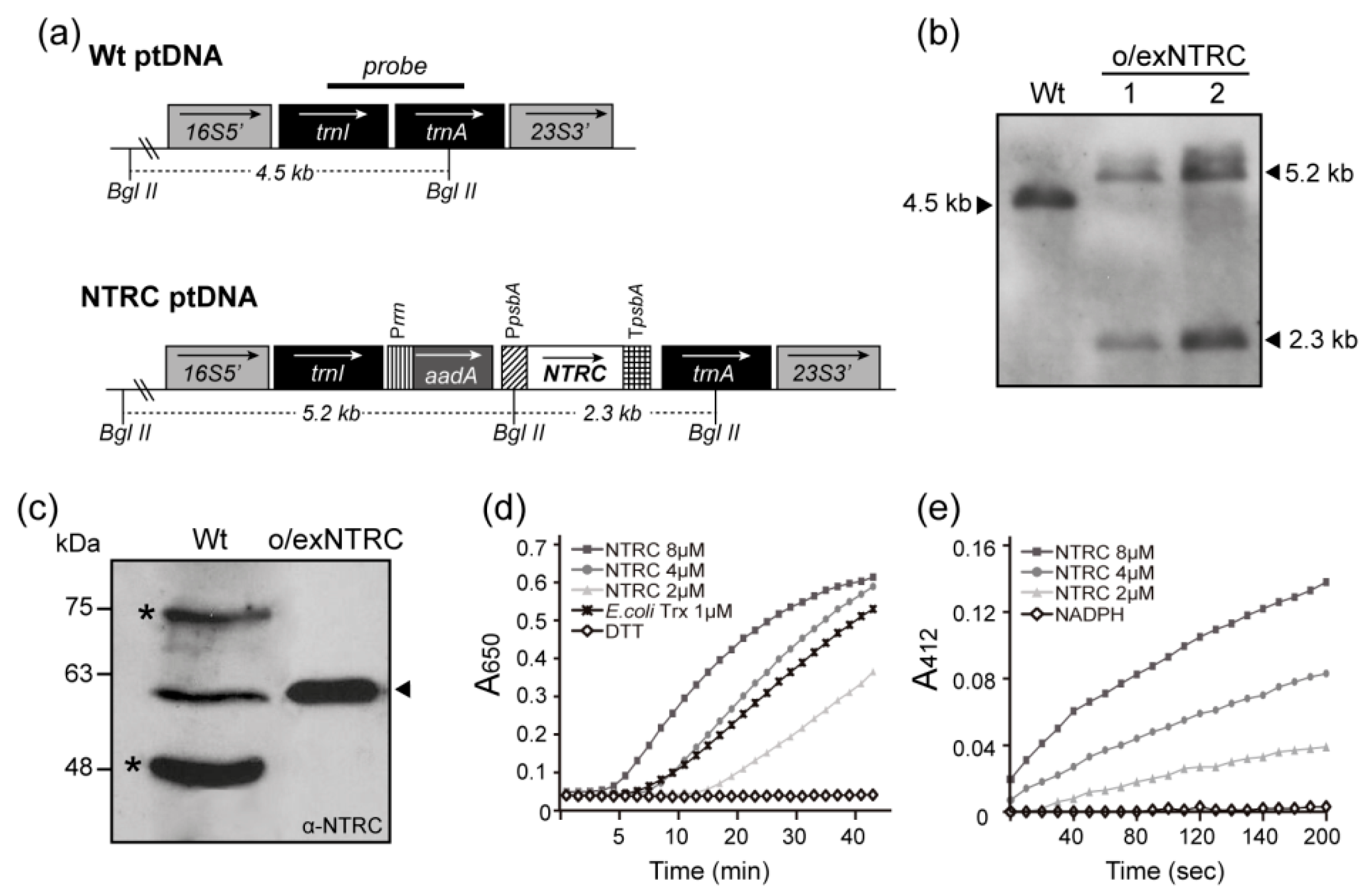
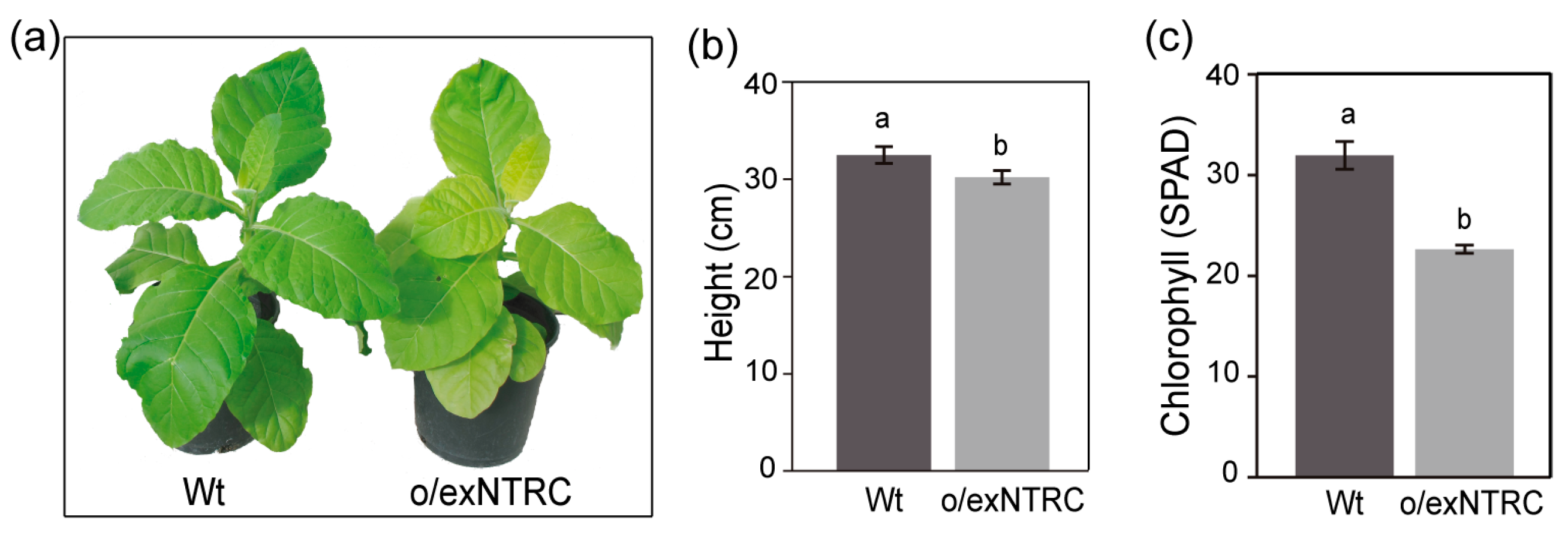
2.1. Generation of Transplastomic Tobacco Plants Overexpressing a Fully Functional NTRC
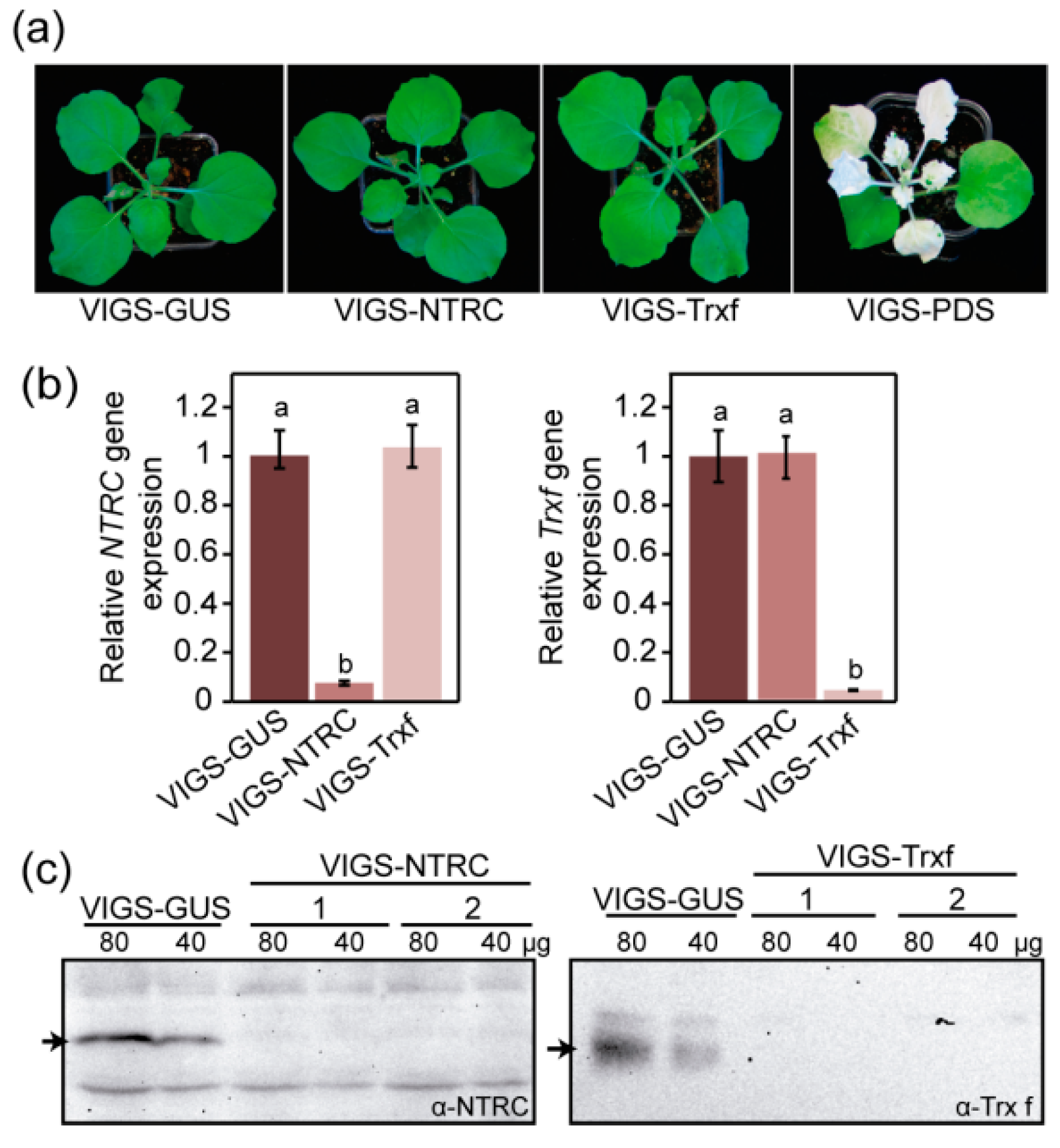
2.2. Production of Nicotiana Benthamiana Plants with Reduced Expression of NTRC or Trx f
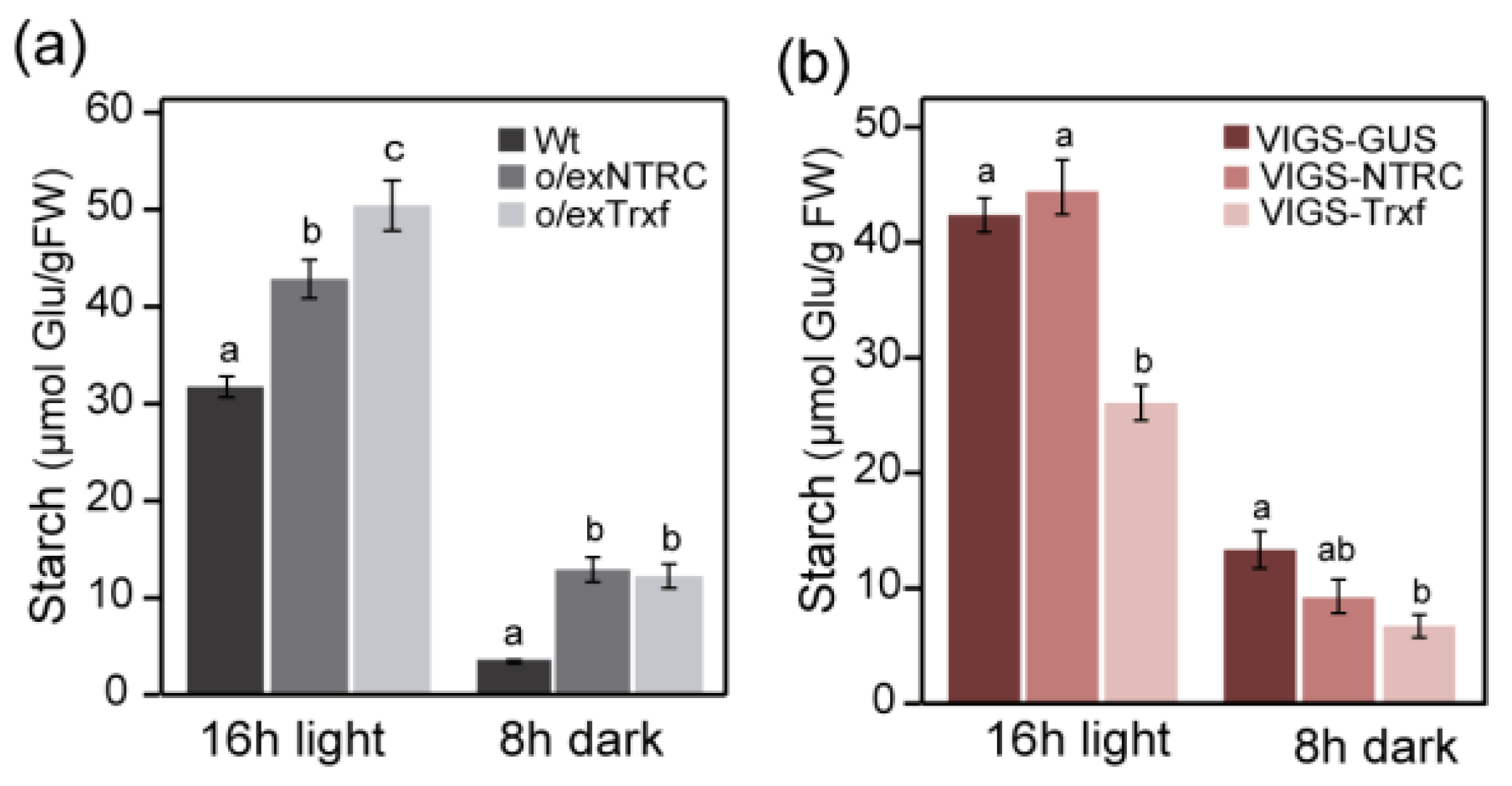
2.3. Transient Starch Accumulation Pattern in Leaves of Plants with Altered NTRC or Trx f Levels
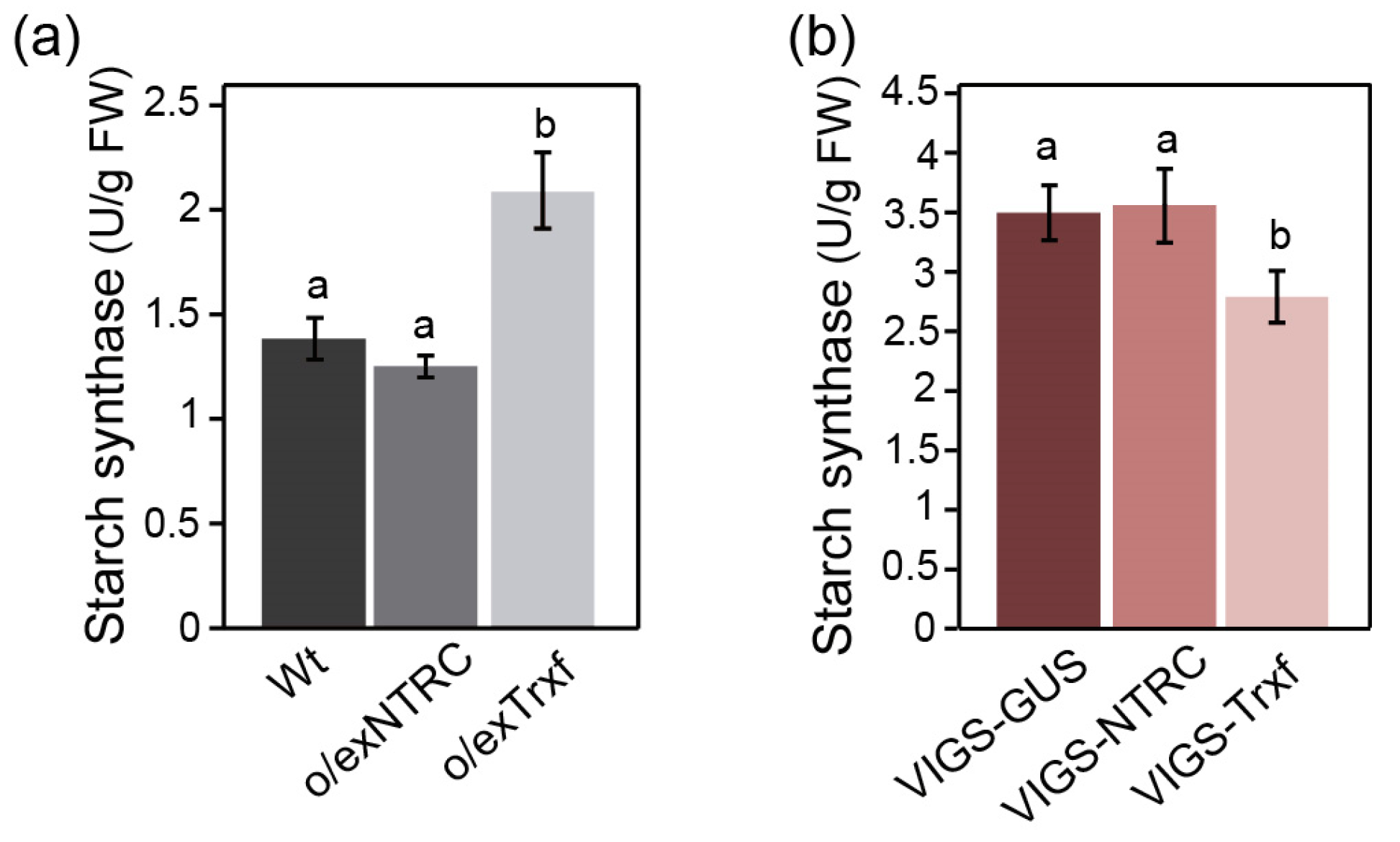
2.4. Comparative Analysis of Enzymes Related to Starch Synthesis
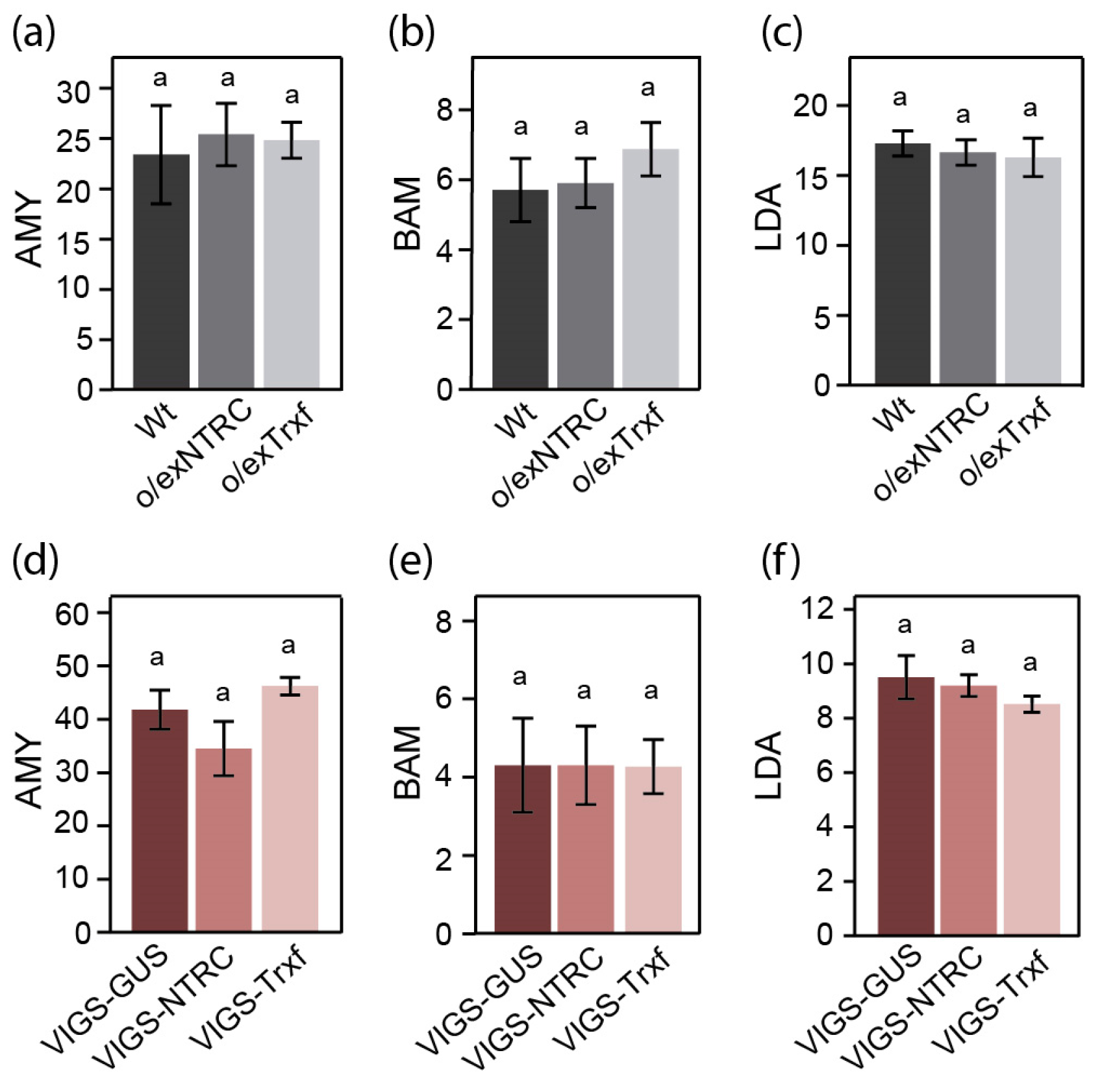
2.5. Comparative Analysis of Enzymes Related to Starch Degradation
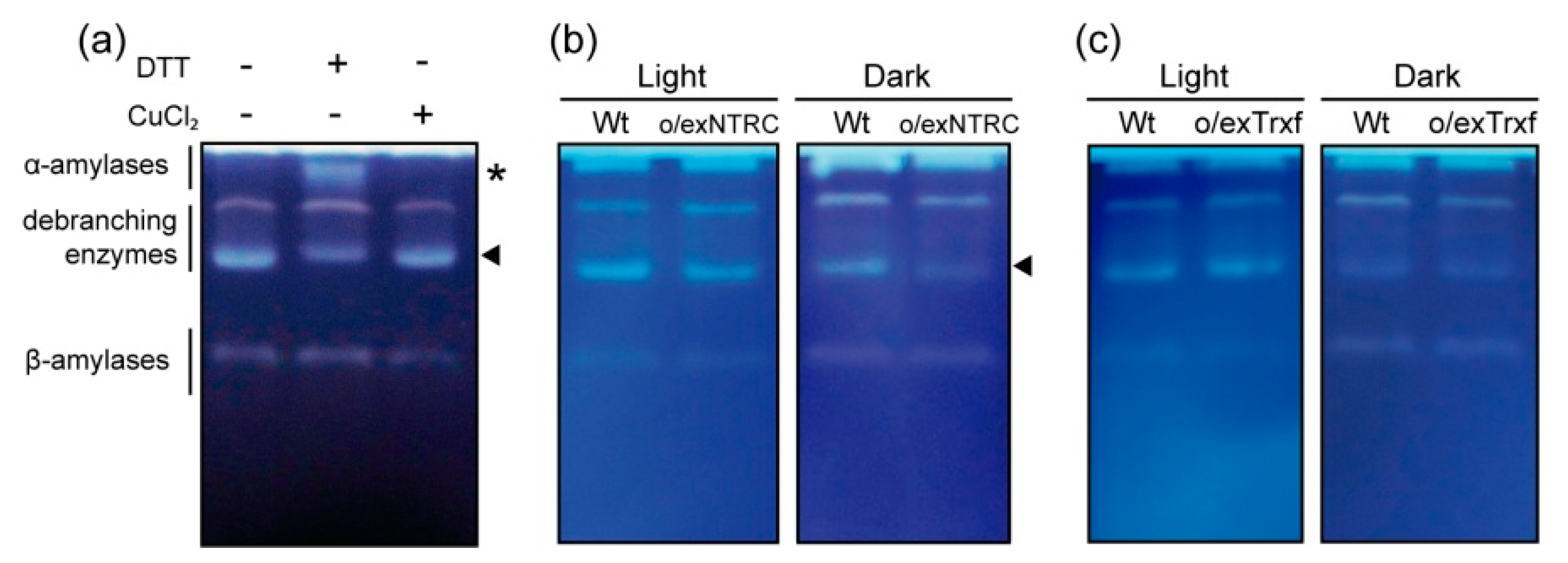
2.6. Qualitative Study of Redox Sensitive Amylolytic Enzymes in Tobacco Plants
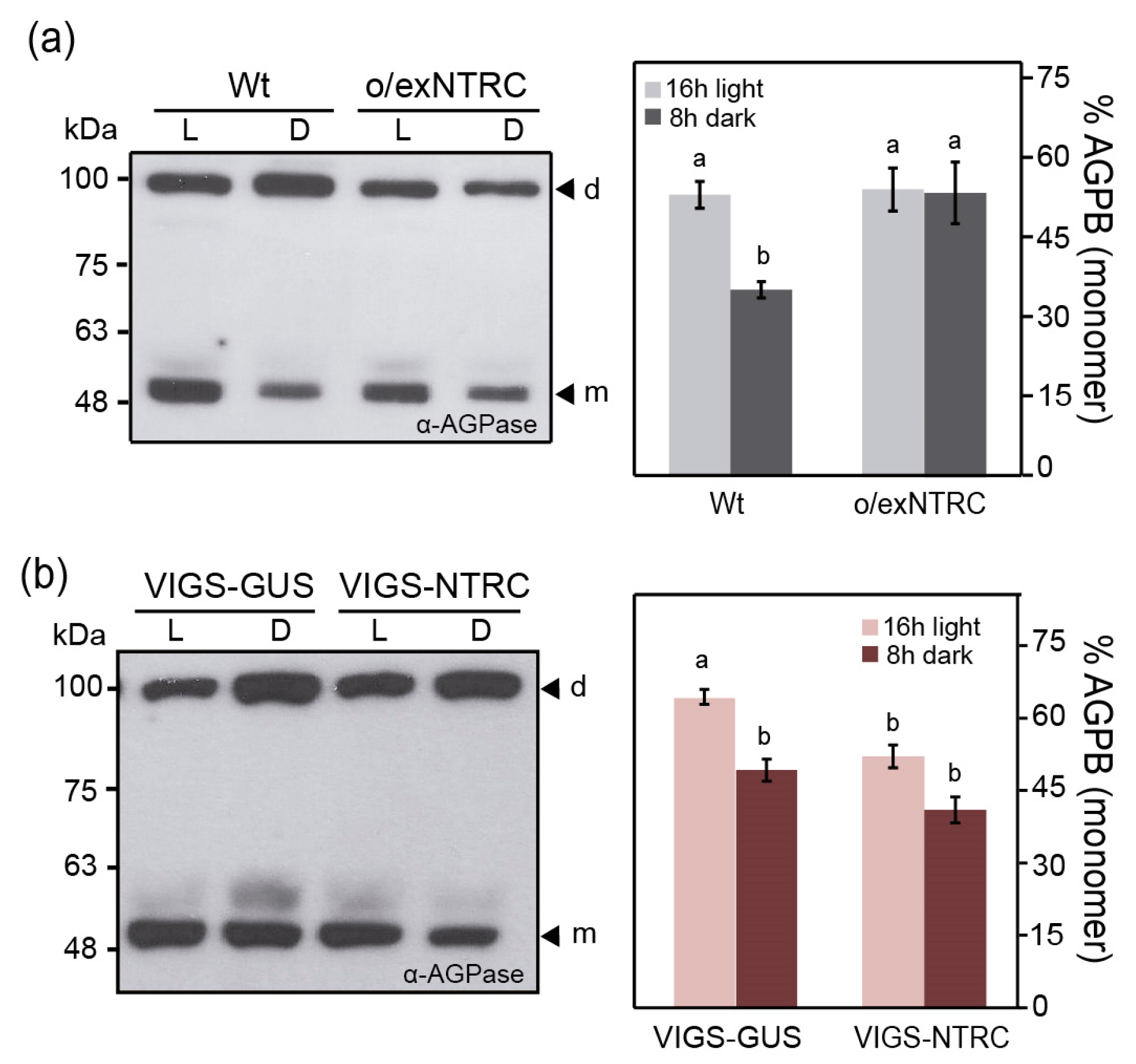
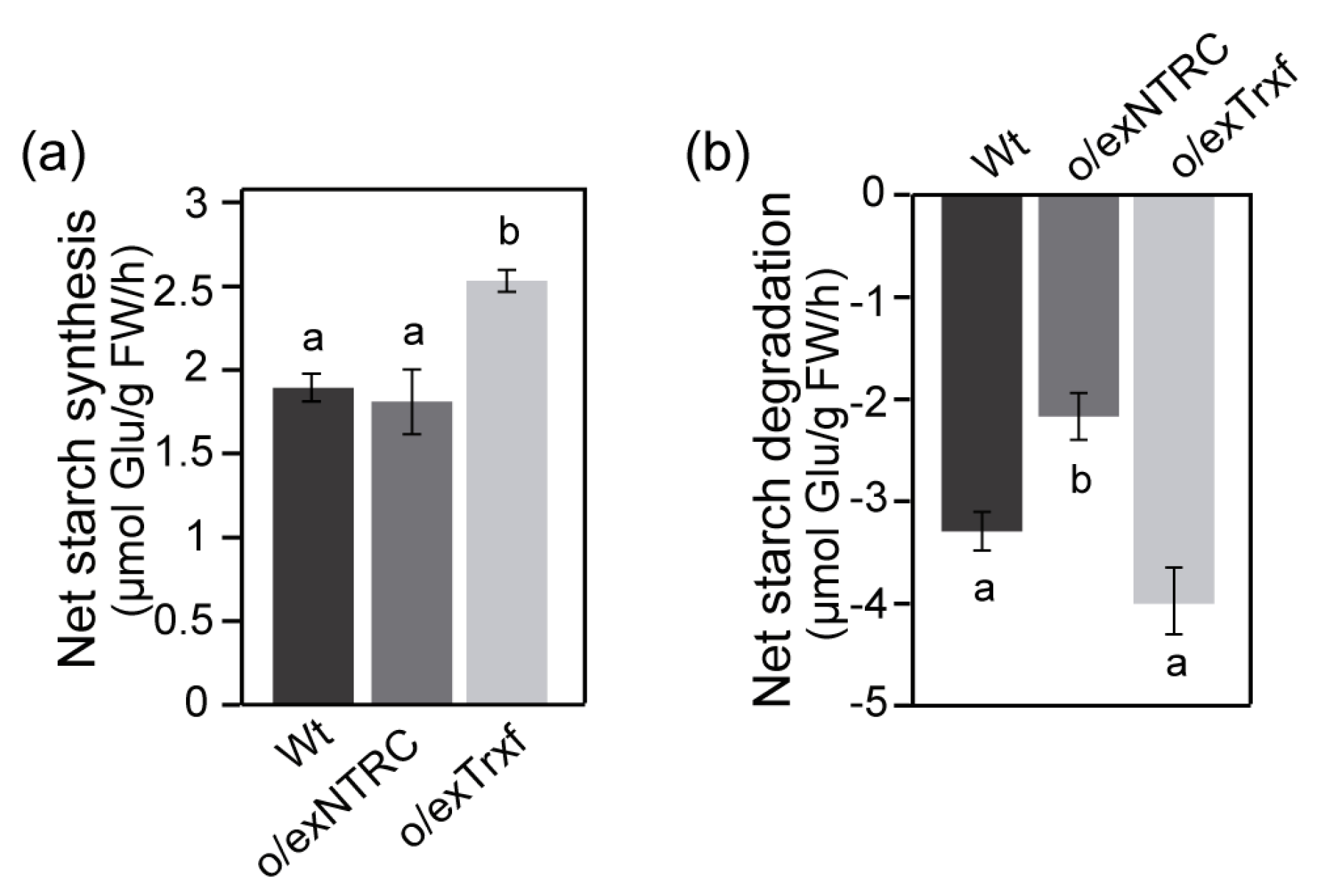
2.7. NTRC and Trx f Overexpression Differentially Alters the Net Starch Synthesis and Degradation
3. Discussion
3.1. Phenotype of N. Tabacum and N. Benthamiana Plants with Altered NTRC Protein Levels
3.2. Enhanced Starch Content in o/exNTRC Leaves as a Consequence of Impaired Starch Metabolism at Night
3.3. Starch Synthase as the Main Determinant of Starch Accumulation in Trx f Transgenic Plants
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. DNA, RNA and Protein Analysis
4.3. Protein Purification and Activity Assays
4.4. Starch Determination
4.5. Enzyme Activities Associated with Starch Synthesis and Degradation
4.6. AGPB Redox Status
4.7. Zymograms of Starch Hydrolytic Activities
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buchanan, B.B.; Holmgren, A.; Jacquot, J.P.; Scheibe, R. Fifty years in the thioredoxin field and a bountiful harvest. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, A. Thioredoxin catalyzes the reduction of insulin disulfides by dithiothreitol and dihydrolipoamide. J. Biol. Chem. 1979, 254, 9627–9632. [Google Scholar] [PubMed]
- Chibani, K.; Wingsle, G.; Jacquot, J.P.; Gelhaye, E.; Rouhier, N. Comparative genomic study of the thioredoxin family in photosynthetic organisms with emphasis on Populus trichocarpa. Mol. Plant 2009, 2, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Montrichard, F.; Alkhalfioui, F.; Yano, H.; Vensel, W.H.; Hurkman, W.J.; Buchanan, B.B. Thioredoxin targets in plants: The first 30 years. J. Proteomics 2009, 72, 452–474. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, B.B. Role of Light in the Regulation of Chloroplast Enzymes. Annu. Rev. Plant Physiol. 1980, 31, 341–374. [Google Scholar] [CrossRef]
- Balmer, Y.; Vensel, W.H.; DuPont, F.M.; Buchanan, B.B.; Hurkman, W.J. Proteome of amyloplasts isolated from developing wheat endosperm presents evidence of broad metabolic capability. J. Exp. Bot. 2006, 57, 1591–1602. [Google Scholar] [CrossRef]
- Arsova, B.; Hoja, U.; Wimmelbacher, M.; Greiner, E.; Üstün, S.; Melzer, M.; Petersen, K.; Lein, W.; Börnke, F. Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: Evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell Online 2010, 22, 1498–1515. [Google Scholar] [CrossRef]
- Collin, V.; Lamkemeyer, P.; Miginiac-Maslow, M.; Hirasawa, M.; Knaff, D.B.; Dietz, K.-J.; Issakidis-Bourguet, E. Characterization of plastidial thioredoxins from Arabidopsis belonging to the new y-type. Plant Physiol. 2004, 136, 4088–4095. [Google Scholar] [CrossRef]
- Meyer, Y.; Belin, C.; Delorme-Hinoux, V.; Reichheld, J.-P.; Riondet, C. Thioredoxin and glutaredoxin systems in plants: Molecular mechanisms, crosstalks, and functional significance. Antioxid. Redox Signal. 2012, 17, 1124–1160. [Google Scholar] [CrossRef]
- Serrato, A.J.; Pérez-Ruiz, J.M.; Spínola, M.C.; Cejudo, F.J. A novel NADPH thioredoxin reductase, localised in the chloroplast, which deficiency causes hypersensitivity to abiotic stress in Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 43821–43827. [Google Scholar] [CrossRef]
- Zaffagnini, M.; Fermani, S.; Marchand, C.H.; Costa, A.; Sparla, F.; Rouhier, N.; Geigenberger, P.; Lemaire, S.D.; Trost, P. Redox Homeostasis in Photosynthetic Organisms: Novel and Established Thiol-Based Molecular Mechanisms. Antioxid. Redox Signal. 2018, 17, ars.2018.7617. [Google Scholar] [CrossRef] [PubMed]
- Spínola, M.C.; Pérez-Ruiz, J.M.; Pulido, P.; Kirchsteiger, K.; Guinea, M.; González, M.; Cejudo, F.J. NTRC new ways of using NADPH in the chloroplast. Physiol. Plant. 2008, 133, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Kirchsteiger, K.; Ferrández, J.; Pascual, M.B.; González, M.; Cejudo, F.J. NADPH Thioredoxin Reductase C Is Localized in Plastids of Photosynthetic and Nonphotosynthetic Tissues and Is Involved in Lateral Root Formation in Arabidopsis. Plant Cell 2012, 24, 1534–1548. [Google Scholar] [CrossRef] [PubMed]
- Alkhalfioui, F.; Renard, M.; Montrichard, F. Unique properties of NADP-thioredoxin reductase C in legumes. J. Exp. Bot. 2007, 58, 969–978. [Google Scholar] [CrossRef]
- Moon, J.C.; Jang, H.H.; Chae, H.B.; Lee, J.R.; Lee, S.Y.; Jung, Y.J.; Shin, M.R.; Lim, H.S.; Chung, W.S.; Yun, D.J.; et al. The C-type Arabidopsis thioredoxin reductase ANTR-C acts as an electron donor to 2-Cys peroxiredoxins in chloroplasts. Biochem. Biophys. Res. Commun. 2006, 348, 478–484. [Google Scholar] [CrossRef]
- Perez-Ruiz, J.M.; Spínola, M.C.; Kirchsteiger, K.; Moreno, J.; Sahrawy, M.; Cejudo, F.J. Rice NTRC is a high-efficiency redox system for chloroplast protection against oxidative damage. Plant Cell 2006, 18, 2356–2368. [Google Scholar] [CrossRef]
- Pérez-Ruiz, J.M.; Cejudo, F.J. A proposed reaction mechanism for rice NADPH thioredoxin reductase C, an enzyme with protein disulfide reductase activity. FEBS Lett. 2009, 583, 1399–1402. [Google Scholar] [CrossRef]
- Stitt, M.; Lunn, J.; Usadel, B. Arabidopsis and primary photosynthetic metabolism - More than the icing on the cake. Plant J. 2010, 61, 1067–1091. [Google Scholar] [CrossRef]
- Baslam, M.; Baroja-Fernández, E.; Ricarte-Bermejo, A.; Sánchez-López, Á.M.; Aranjuelo, I.; Bahaji, A.; Muñoz, F.J.; Almagro, G.; Pujol, P.; Galarza, R.; et al. Genetic and isotope ratio mass spectrometric evidence for the occurrence of starch degradation and cycling in illuminated Arabidopsis leaves. PLoS ONE 2017, 12, e0171245. [Google Scholar] [CrossRef]
- Baroja-Fernandez, E.; Munoz, F.J.; Zandueta-Criado, A.; Moran-Zorzano, M.T.; Viale, A.M.; Alonso-Casajus, N.; Pozueta-Romero, J. Most of ADP-glucose linked to starch biosynthesis occurs outside the chloroplast in source leaves. Proc. Natl. Acad. Sci. USA 2004, 101, 13080–13085. [Google Scholar] [CrossRef]
- Pozueta-Romero, J.; Ardila, F.; Akazawa, T. ADP-Glucose Transport by the Chloroplast Adenylate Translocator Is Linked to Starch Biosynthesis. Plant Physiol. 1991, 97, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Pfister, B.; Zeeman, S.C. Formation of starch in plant cells. Cell. Mol. Life Sci. 2016, 73, 2781–2807. [Google Scholar] [CrossRef] [PubMed]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [PubMed]
- Skryhan, K.; Gurrieri, L.; Sparla, F.; Trost, P.; Blennow, A. Redox Regulation of Starch Metabolism. Front. Plant Sci. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kötting, O.; Kossmann, J.; Zeeman, S.C.; Lloyd, J.R. Regulation of starch metabolism: The age of enlightenment? Curr. Opin. Plant Biol. 2010, 13, 321–329. [Google Scholar] [CrossRef]
- Ballicora, M.A.; Frueauf, J.B.; Fu, Y.; Schürmann, P.; Preiss, J.; Ballicora, M.A.; Frueauf, J.B.; Fu, Y.; Schu, P.; Preiss, J. Activation of the potato tuber ADP-glucose pyrophosphorylase by thioredoxin. J. Biol. Chem. 2000, 275, 1315–1320. [Google Scholar] [CrossRef]
- Hendriks, J.H.M.; Kolbe, A.; Gibon, Y.; Stitt, M.; Geigenberger, P. ADP-glucose pyrophosphorylase is activated by posttranslational redox-modification in response to light and to sugars in leaves of Arabidopsis and other plant species. Plant Physiol. 2003, 133, 838–849. [Google Scholar] [CrossRef]
- Michalska, J.; Zauber, H.; Buchanan, B.B.; Cejudo, F.J.; Geigenberger, P. NTRC links built-in thioredoxin to light and sucrose in regulating starch synthesis in chloroplasts and amyloplasts. Proc. Natl. Acad. Sci. USA 2009, 106, 9908–9913. [Google Scholar] [CrossRef]
- Ojeda, V.; Pérez-Ruiz, J.M.; González, M.; Nájera, V.A.; Sahrawy, M.; Serrato, A.J.; Geigenberger, P.; Cejudo, F.J. NADPH thioredoxin reductase C and thioredoxins act concertedly in seedling development. Plant Physiol. 2017, 174, 1436–1448. [Google Scholar] [CrossRef]
- Thormählen, I.; Meitzel, T.; Groysman, J.; Öchsner, A.B.; von Roepenack-Lahaye, E.; Naranjo, B.; Cejudo, F.J.; Geigenberger, P. Thioredoxin f1 and NADPH-dependent thioredoxin reductase C have overlapping functions in regulating photosynthetic metabolism and plant growth in response to varying light conditions. Plant Physiol. 2015, 169, 1766–1786. [Google Scholar] [CrossRef]
- Naranjo, B.; Diaz-Espejo, A.; Lindahl, M.; Cejudo, F.J. Type-f thioredoxins have a role in the short-term activation of carbon metabolism and their loss affects growth under short-day conditions in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 1951–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thormählen, I.; Ruber, J.; Von Roepenack-Lahaye, E.; Ehrlich, S.M.; Massot, V.; Hümmer, C.; Tezycka, J.; Issakidis-Bourguet, E.; Geigenberger, P. Inactivation of thioredoxin f1 leads to decreased light activation of ADP-glucose pyrophosphorylase and altered diurnal starch turnover in leaves of Arabidopsis plants. Plant Cell Environ. 2013, 36, 16–29. [Google Scholar]
- Hädrich, N.; Hendriks, J.H.M.; Kötting, O.; Arrivault, S.; Feil, R.; Zeeman, S.C.; Gibon, Y.; Schulze, W.X.; Stitt, M.; Lunn, J.E. Mutagenesis of cysteine 81 prevents dimerization of the APS1 subunit of ADP-glucose pyrophosphorylase and alters diurnal starch turnover in Arabidopsis thaliana leaves. Plant J. 2012, 70, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Almagro, G.; Muñoz, F.J.; Baroja-Fernández, E.; Bahaji, A.; Montero, M.; Hidalgo, M.; Sánchez-López, Á.M.; Ezquer, I.; Sesma, M.T.; et al. Post-translational redox modification of ADP-glucose pyrophosphorylase in response to light is not a major determinant of fine regulation of transitory starch accumulation in Arabidopsis leaves. Plant Cell Physiol. 2012, 53, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Lunn, J.E.; Delorge, I.; Figueroa, C.M.; Van Dijck, P.; Stitt, M. Trehalose metabolism in plants. Plant J. 2014, 79, 544–567. [Google Scholar] [CrossRef]
- Glaring, M.A.; Skryhan, K.; Kötting, O.; Zeeman, S.C.; Blennow, A. Comprehensive survey of redox sensitive starch metabolising enzymes in Arabidopsis thaliana. Plant Physiol. Biochem. 2012, 58, 89–97. [Google Scholar] [CrossRef]
- Skryhan, K.; Cuesta-Seijo, J.A.; Nielsen, M.M.; Marri, L.; Mellor, S.B.; Glaring, M.A.; Jensen, P.E.; Palcic, M.M.; Blennow, A. The role of cysteine residues in redox regulation and protein stability of Arabidopsis thaliana starch synthase 1. PLoS ONE 2015, 10, e0136997. [Google Scholar] [CrossRef]
- eung, D.; Thalmann, M.; Sparla, F.; Hachem, M.A.; Lee, S.K.; Issakidis-Bourguet, E.; Svensson, B.; Zeeman, S.C.; Santelia, D. Arabidopsis thaliana AMY3 is a unique redox-regulated chloroplastic α-amylase. J. Biol. Chem. 2013, 288, 33620–33633. [Google Scholar]
- Valerio, C.; Costa, A.; Marri, L.; Issakidis-Bourguet, E.; Pupillo, P.; Trost, P.; Sparla, F. Thioredoxin-regulated β-amylase (BAM1) triggers diurnal starch degradation in guard cells, and in mesophyll cells under osmotic stress. J. Exp. Bot. 2011, 62, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Sparla, F.; Costa, A.; Schiavo, F.L.; Pupillo, P.; Trost, P. Redox regulation of a novel plastid-targeted β-Amylase. Plant Physiol. 2006, 141, 840–850. [Google Scholar] [CrossRef] [Green Version]
- Mikkelsen, R.; Mutenda, K.E.; Mant, A.; Schürmann, P.; Blennow, A. α-glucan, water dikinase (GWD): A plastidic enzyme with redox-regulated and coordinated catalytic activity and binding affinity. Proc. Natl. Acad. Sci. USA 2005, 102, 1785–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sokolov, L.N.; Dominguez-Solis, J.R.; Allary, A.-L.; Buchanan, B.B.; Luan, S. A redox-regulated chloroplast protein phosphatase binds to starch diurnally and functions in its accumulation. Proc. Natl. Acad. Sci. USA 2006, 103, 9732–9737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, D.M.; Silva, L.P.; Issakidis-Bourguet, E.; Glaring, M.A.; Schriemer, D.C.; Moorhead, G. Insight into the redox regulation of the phosphoglucan phosphatase SEX4 involved in starch degradation. FEBS J. 2013, 280, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Skeffington, A.W.; Graf, A.; Duxbury, Z.; Gruissem, W.; Smith, A.M. Glucan, water dikinase exerts little control over starch degradation in Arabidopsis leaves at night. Plant Physiol. 2014, 165, 866–879. [Google Scholar] [CrossRef] [Green Version]
- Stitt, M.; Zeeman, S.C. Starch turnover: Pathways, regulation and role in growth. Curr. Opin. Plant Biol. 2012, 15, 282–292. [Google Scholar] [CrossRef]
- Santelia, D.; Trost, P.; Sparla, F. New insights into redox control of starch degradation. Curr. Opin. Plant Biol. 2015, 25, 1–9. [Google Scholar] [CrossRef]
- Bahaji, A.; Li, J.; Sánchez-López, Á.M.; Baroja-Fernández, E.; Muñoz, F.J.; Ovecka, M.; Almagro, G.; Montero, M.; Ezquer, I.; Etxeberria, E.; et al. Starch biosynthesis, its regulation and biotechnological approaches to improve crop yields. Biotechnol. Adv. 2014, 32, 87–106. [Google Scholar] [CrossRef]
- Sanz-Barrio, R.; Corral-Martínez, P.; Ancin, M.; Segui-Simarro, J.M.; Farran, I. Overexpression of plastidial thioredoxin f leads to enhanced starch accumulation in tobacco leaves. Plant Biotechnol. J. 2013, 11, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Harries, P.A.; Palanichelvam, K.; Bhat, S.; Nelson, R.S. Tobacco mosaic virus 126-kDa Protein Increases the Susceptibility of Nicotiana tabacum to Other Viruses and Its Dosage Affects Virus-Induced Gene Silencing. Mol. Plant-Microbe Interact. 2008, 21, 1539–1548. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Schiff, M.; Marathe, R.; Dinesh-Kumar, S.P. Tobacco Rar1, EDS1 and NPR1/NIM1 like genes are required for N-mediated resistance to tobacco mosaic virus. Plant J. 2002, 30, 415–429. [Google Scholar] [CrossRef]
- Sanz-Barrio, R.; Fernández-San Millán, A.; Corral-Martínez, P.; Seguí-Simarro, J.M.; Farran, I. Tobacco plastidial thioredoxins as modulators of recombinant protein production in transgenic chloroplasts. Plant Biotechnol. J. 2011, 9, 639–650. [Google Scholar] [CrossRef] [Green Version]
- Pulido, P.; Spínola, M.C.; Kirchsteiger, K.; Guinea, M.; Pascual, M.B.; Sahrawy, M.; Sandalio, L.M.; Dietz, K.-J.; González, M.; Cejudo, F.J. Functional analysis of the pathways for 2-Cys peroxiredoxin reduction in Arabidopsis thaliana chloroplasts. J. Exp. Bot. 2010, 61, 4043–4054. [Google Scholar] [CrossRef] [PubMed]
- Häusler, R.E.; Schlieben, N.H.; Nicolay, P.; Fischer, K.; Fischer, K.L.; Flügge, U.I. Control of carbon partitioning and photosynthesis by the triose phosphate/phosphate translocator in transgenic tobacco plants (Nicotiana tabacum L.). I. Comparative physiological analysis of tobacco plants with antisense repression and overexpressi. Planta 2000, 210, 371–382. [Google Scholar]
- Geigenberger, P.; Thormählen, I.; Daloso, D.M.; Fernie, A.R. The unprecedented versatility of the plant thioredoxin system. Trends Plant Sci. 2017, 22, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Hisabori, T. Two distinct redox cascades cooperatively regulate chloroplast functions and sustain plant viability. Proc. Natl. Acad. Sci. USA 2016, 113, E3967–E3976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toivola, J.; Nikkanen, L.; Dahlström, K.M.; Salminen, T.A.; Lepistö, A.; Vignols, F.; Rintamäki, E. Overexpression of chloroplast NADPH-dependent thioredoxin reductase in Arabidopsis enhances leaf growth and elucidates in vivo function of reductase and thioredoxin domains. Front. Plant Sci. 2013, 4, 389. [Google Scholar] [CrossRef] [Green Version]
- Richter, A.S.; Grimm, B. Thiol-based redox control of enzymes involved in the tetrapyrrole biosynthesis pathway in plants. Front. Plant Sci. 2013, 4, 371. [Google Scholar] [CrossRef] [Green Version]
- Lepistö, A.; Pakula, E.; Toivola, J.; Krieger-Liszkay, A.; Vignols, F.; Rintamäki, E. Deletion of chloroplast NADPH-dependent thioredoxin reductase results in inability to regulate starch synthesis and causes stunted growth under short-day photoperiods. J. Exp. Bot. 2013, 64, 3843–3854. [Google Scholar] [CrossRef]
- Naranjo, B.; Mignée, C.; Krieger-Liszkay, A.; Hornero-Méndez, D.; Gallardo-Guerrero, L.; Cejudo, F.J.; Lindahl, M. The chloroplast NADPH thioredoxin reductase C, NTRC, controls non-photochemical quenching of light energy and photosynthetic electron transport in Arabidopsis. Plant Cell Environ. 2016, 39, 804–822. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Ruiz, J.M.; Naranjo, B.; Ojeda, V.; Guinea, M.; Cejudo, F.J. NTRC-dependent redox balance of 2-Cys peroxiredoxins is needed for optimal function of the photosynthetic apparatus. Proc. Natl. Acad. Sci. USA 2017, 114, 12069–12074. [Google Scholar] [CrossRef] [Green Version]
- Nikkanen, L.; Toivola, J.; Rintamäki, E. Crosstalk between chloroplast thioredoxin systems in regulation of photosynthesis. Plant Cell Environ. 2016, 39, 1691–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, M.C.M.; Hejazi, M.; Fettke, J.; Steup, M.; Feil, R.; Krause, U.; Arrivault, S.; Vosloh, D.; Figueroa, C.M.; Ivakov, A.; et al. Feedback inhibition of starch degradation in Arabidopsis leaves mediated by trehalose 6-phosphate. Plant Physiol. 2013, 163, 1142–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Née, G.; Zaffagnini, M.; Trost, P.; Issakidis-Bourguet, E. Redox regulation of chloroplastic glucose-6-phosphate dehydrogenase: A new role for f-type thioredoxin. FEBS Lett. 2009, 583, 2827–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Née, G.; Aumont-Nicaise, M.; Zaffagnini, M.; Nessler, S.; Valerio-Lepiniec, M.; Issakidis-Bourguet, E. Redox regulation of chloroplastic G6PDH activity by thioredoxin occurs through structural changes modifying substrate accessibility and cofactor binding. Biochem. J. 2014, 457, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Schindler, I.; Renz, A.; Schmid, F.X.; Beck, E. Activation of spinach pullulanase by reduction results in a decrease in the number of isomeric forms. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2001, 1548, 175–186. [Google Scholar] [CrossRef]
- Repellin, A.; Båga, M.; Chibbar, R.N. In vitro pullulanase activity of wheat (Triticum aestivum L.) limit-dextrinase type starch debranching enzyme is modulated by redox conditions. J. Cereal Sci. 2008, 47, 302–309. [Google Scholar] [CrossRef]
- Delatte, T.; Umhang, M.; Trevisan, M.; Eicke, S.; Thorneycroft, D.; Smith, S.M.; Zeeman, S.C. Evidence for distinct mechanisms of starch granule breakdown in plants. J. Biol. Chem. 2006, 281, 12050–12059. [Google Scholar] [CrossRef] [Green Version]
- Wattebled, F.; Dong, Y.; Dumez, S.; Delvallé, D.; Planchot, V.; Berbezy, P.; Vyas, D.; Colonna, P.; Chatterjee, M.; Ball, S.; et al. Mutants of Arabidopsis lacking a chloroplastic isoamylase accumulate phytoglycogen and an abnormal form of amylopectin. Plant Physiol. 2005, 138, 184–195. [Google Scholar] [CrossRef] [Green Version]
- Farran, I.; Fernández-San Millán, A.; Ancin, M.; Larraya, L.; Veramendi, J. Increased bioethanol production from commercial tobacco cultivars overexpressing thioredoxin f grown under field conditions. Mol. Breed. 2014, 34, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Aranjuelo, I.; Tcherkez, G.; Jauregui, I.; Gilard, F.; Ancin, M.; Fernández-San Millán, A.; Larraya, L.; Veramendi, J.; Farran, I. Alteration by thioredoxin f over-expression of primary carbon metabolism and its response to elevated CO2 in tobacco (Nicotiana tabacum L.). Environ. Exp. Bot. 2015, 118, 40–48. [Google Scholar] [CrossRef]
- Delvallé, D.; Dumez, S.; Wattebled, F.; Roldán, I.; Planchot, V.; Berbezy, P.; Colonna, P.; Vyas, D.; Chatterjee, M.; Ball, S.; et al. Soluble starch synthase I: A major determinant for the synthesis of amylopectin in Arabidopsis thaliana leaves. Plant J. 2005, 43, 398–412. [Google Scholar] [CrossRef] [PubMed]
- Cuesta-Seijo, J.A.; Nielsen, M.M.; Marri, L.; Tanaka, H.; Beeren, S.R.; Palcic, M.M. Structure of starch synthase I from barley: Insight into regulatory mechanisms of starch synthase activity. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 69, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, X.; Ye, Y.; Ren, G.; Li, F.; Qi, S.; Wang, B.; Fan, S.; Zhou, Q. Enhancement of starch content by constitutive expression of GmTrxF in transgenic Arabidopsis. Plant Growth Regul. 2018, 84, 351–358. [Google Scholar] [CrossRef]
- Wang, F.B.; Kong, W.L.; Fu, Y.R.; Sun, X.C.; Chen, X.H.; Zhou, Q. Constitutive expression of SlTrxF increases starch content in transgenic Arabidopsis. Biol. Plant. 2017, 61, 494–500. [Google Scholar] [CrossRef]
- Fernández-San Millán, A.; Ortigosa, S.M.; Hervás-Stubbs, S.; Corral-Martínez, P.; Seguí-Simarro, J.M.; Gaétan, J.; Coursaget, P.; Veramendi, J. Human papillomavirus L1 protein expressed in tobacco chloroplasts self-assembles into virus-like particles that are highly immunogenic. Plant Biotechnol. J. 2008, 6, 427–441. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Liu, Y.L.; Schiff, M.; Dinesh-Kumar, S.P. Virus-induced gene silencing in tomato. Plant J. 2002, 31, 777–786. [Google Scholar] [CrossRef]
- Liu, Y.; Nakayama, N.; Schiff, M.; Litt, A.; Irish, V.F.; Dinesh-Kumar, S.P. Virus induced gene silencing of a DEFICIENS ortholog in Nicotiana benthamiana. Plant Mol. Biol. 2004, 54, 701–711. [Google Scholar] [CrossRef]
- Wu, C.; Jia, L.; Goggin, F. The reliability of virus-induced gene silencing experiments using tobacco rattle virus in tomato is influenced by the size of the vector control. Mol. Plant Pathol. 2011, 12, 299–305. [Google Scholar] [CrossRef]
- Sul, I.; Korban, S. A highly efficient method for isolating genomic DNA from plant tissues. Plant Tissue Cult. Biotechnol. 1996, 2, 113–116. [Google Scholar]
- Ganusova, E.E.; Rice, J.H.; Carlew, T.S.; Patel, A.; Perrodin-Njoku, E.; Hewezi, T.; Burch-Smith, T.M. Altered Expression of a Chloroplast Protein Affects the Outcome of Virus and Nematode Infection. Mol. Plant-Microbe Interact. 2017, 30, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 45e. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Barrio, R.; Fernández-San Millán, A.; Carballeda, J.; Corral-Martínez, P.; Seguí-Simarro, J.M.; Farran, I. Chaperone-like properties of tobacco plastid thioredoxins f and m. J. Exp. Bot. 2012, 63, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Yuki, K.; Park, S.; Ohya, T. Carbohydrate metabolism in the developing endosperm of rice grains. Plant Cell Physiol. 1989, 30, 833–839. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ancín, M.; Larraya, L.; Fernández-San Millán, A.; Veramendi, J.; Burch-Smith, T.; Farran, I. NTRC and Thioredoxin f Overexpression Differentially Induces Starch Accumulation in Tobacco Leaves. Plants 2019, 8, 543. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120543
Ancín M, Larraya L, Fernández-San Millán A, Veramendi J, Burch-Smith T, Farran I. NTRC and Thioredoxin f Overexpression Differentially Induces Starch Accumulation in Tobacco Leaves. Plants. 2019; 8(12):543. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120543
Chicago/Turabian StyleAncín, María, Luis Larraya, Alicia Fernández-San Millán, Jon Veramendi, Tessa Burch-Smith, and Inmaculada Farran. 2019. "NTRC and Thioredoxin f Overexpression Differentially Induces Starch Accumulation in Tobacco Leaves" Plants 8, no. 12: 543. https://0-doi-org.brum.beds.ac.uk/10.3390/plants8120543