A Critical Cross-Species Comparison of Pollen from Nelumbo nucifera Gaertn. vs. Nymphaea lotus L. for Authentication of Thai Medicinal Herbal Tea

 ,
, 
Abstract
:
1. Introduction
2. Results and Discussion
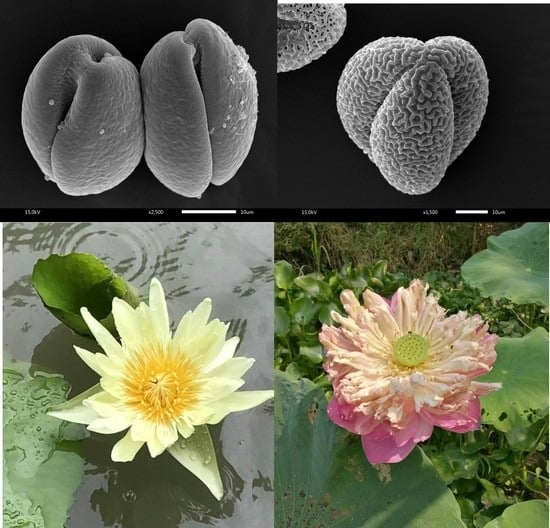
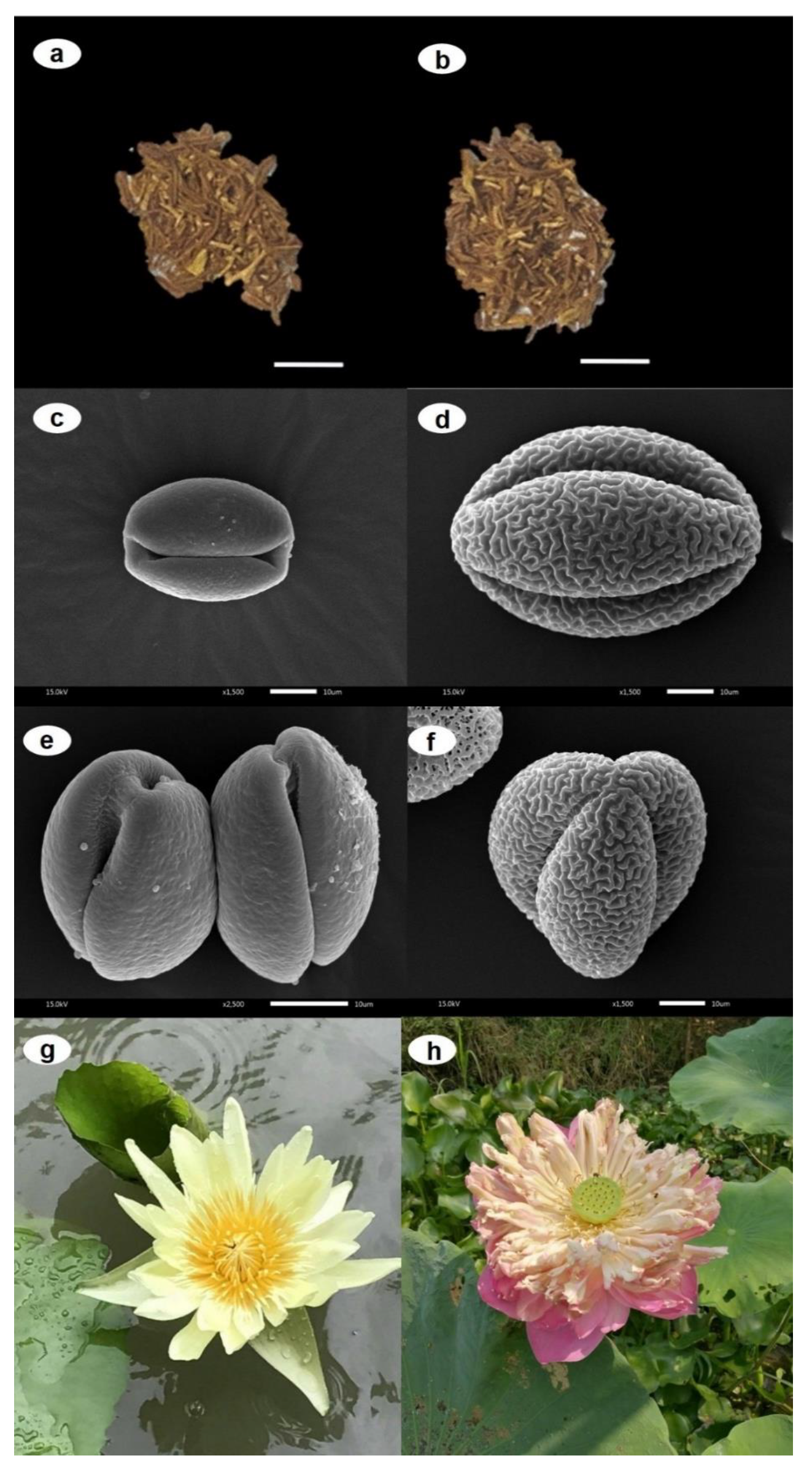
2.1. Macro- and Micro-Morphological Evaluations as Tools for N. nucifera vs. N. lotus Authentication
2.2. Bioinformatic Analysis of Two Nuclear and Plastid DNA Markers as a Tool for the Authentication of N. nucifera vs. N. lotus
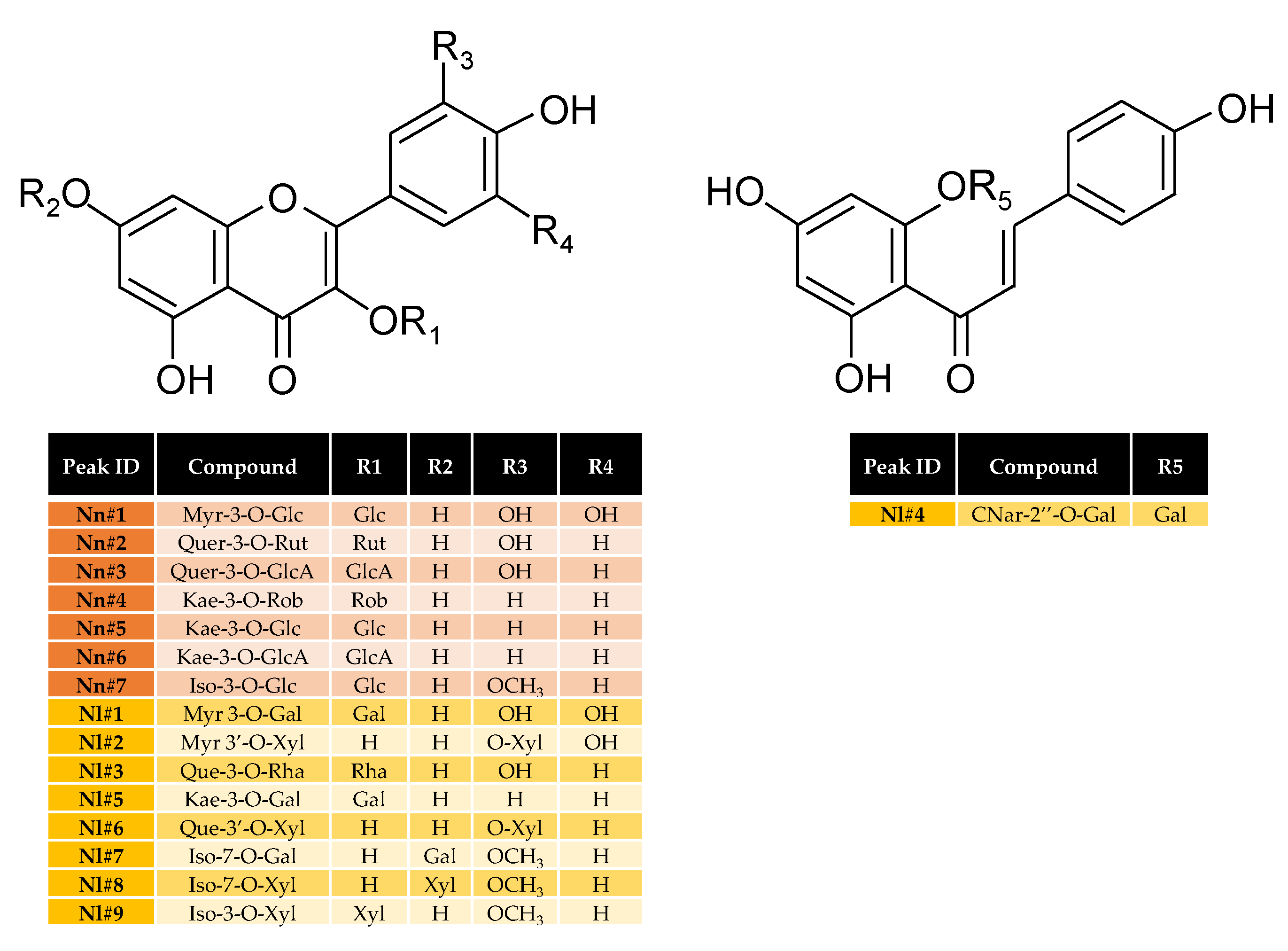
2.3. Flavonoids Profiling in Stamen Extracts as a Tool for the Authentication of N. nucifera vs. N. lotus
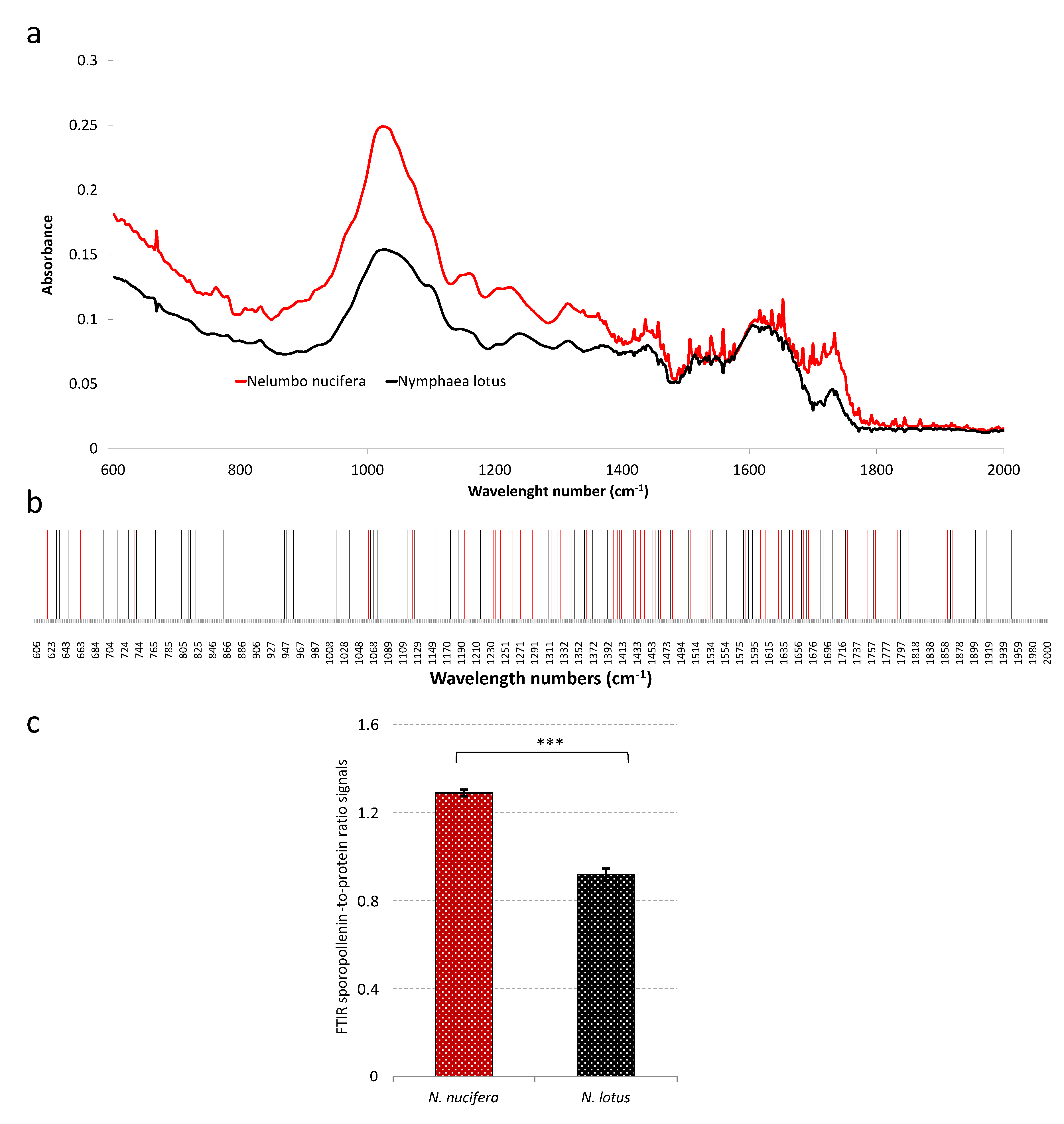
2.4. Pollen FTIR Analysis as a Tool for the Authentication of N. nucifera vs. N. lotus
3. Materials and Methods
3.1. Plant Materials
3.2. Macro- and Micro-Morphological Evaluations as Tools for N. nucifera vs. N. lotus
3.3. Bioinformatic Analysis
3.4. Chemicals
3.5. Flavonoids Extraction
3.6. LC-MS Analysis
3.7. FTIR Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, S.; Zheng, Y.; Fang, J.-B.; Liu, Y.-L.; Li, S.-H. Flavonoids in lotus (Nelumbo) leaves evaluated by HPLC–MSn at the germplasm level. Food Res. Int. 2013, 54, 796–803. [Google Scholar] [CrossRef]
- Sheikh, S.A. Ethno-medicinal uses and pharmacological activities of lotus (Nelumbo nucifera). J. Med. Plants Stud. 2014, 2, 42–46. [Google Scholar]
- Deng, J.; Chen, S.; Yin, X.; Wang, K.; Liu, Y.; Li, S.; Yang, P. Systematic qualitative and quantitative assessment of anthocyanins, flavones and flavonols in the petals of 108 lotus (Nelumbo nucifera) cultivars. Food Chem. 2013, 139, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Shukla, S.; Kim, J.-A.; Kim, M.H. Anti-Angiogenic Effect of Nelumbo nucifera Leaf Extracts in Human Umbilical Vein Endothelial Cells with Antioxidant Potential. PLoS ONE 2015, 10, e0118552. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.-Z.; Wu, W.; Jiao, L.-L.; Yang, P.-F.; Guo, M.-Q. Analysis of Flavonoids in Lotus (Nelumbo nucifera) Leaves and Their Antioxidant Activity Using Macroporous Resin Chromatography Coupled with LC-MS/MS and Antioxidant Biochemical Assays. Molecules 2015, 20, 10553–10565. [Google Scholar] [CrossRef] [Green Version]
- Rai, S.; Wahile, A.; Mukherjee, K.; Saha, B.P.; Mukherjee, P.K. Antioxidant activity of Nelumbo nucifera (sacred lotus) seeds. J. Ethnopharmacol. 2006, 104, 322–327. [Google Scholar] [CrossRef]
- Jung, H.A.; Kim, J.E.; Chung, H.Y.; Choi, J.S. Antioxidant principles of Nelumbo nucifera stamens. Arch. Pharmacal Res. 2003, 26, 279–285. [Google Scholar] [CrossRef]
- Lin, H.-Y.; Kuo, Y.-H.; Lin, Y.-L.; Chiang, W. Antioxidative Effect and Active Components from Leaves of Lotus (Nelumbo nucifera). J. Agric. Food Chem. 2009, 57, 6623–6629. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicine 2018, 5, 93. [Google Scholar] [CrossRef]
- Drouet, S.; Garros, L.; Hano, C.; Tungmunnithum, D.; Renouard, S.; Hagège, D.; Maunit, B.; Lainé, É. A Critical View of Different Botanical, Molecular, and Chemical Techniques Used in Authentication of Plant Materials for Cosmetic Applications. Cosmetics 2018, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Kenđel, A.; Zimmermann, B. Chemical Analysis of Pollen by FT-Raman and FTIR Spectroscopies. Front. Plant Sci. 2020, 11, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tungmunnithum, D.; Kidyoo, M.; Khunwasi, C. Morphological variations in Hoya siamica Craib (Asclepiadaceae) in Thailand. Trop. Nat. Hist. 2011, 11, 29–37. [Google Scholar]
- Depciuch, J.; Kasprzyk, I.; Drzymała, E.; Parlinska-Wojtan, M. Identification of birch pollen species using FTIR spectroscopy. Aerobiology 2018, 34, 525–538. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.U.; Zafar, M.; Ahmad, M.; Anjum, F.; Sultana, S.; Kilic, O.; Ozdemir, F.A.; Nazir, A.; Yaseen, G.; Aabidin, S.Z.U.; et al. Pollen micromorphological analysis of tribe Acacieae (Mimosaceae) with LM and SEM techniques. Microsc. Res. Tech. 2019, 82, 1610–1620. [Google Scholar] [CrossRef]
- Bano, A.; Ahmad, M.; Rashid, S.; Zafar, M.; Ashfaq, S.; Rehman, S.U.; Ali, M.I.; Sultana, S.; Shaheen, S.; Ahmad, S. Microscopic investigations of some selected species of Papilionaceae through SEM and LM from Skardu valley, northern Pakistan. Microsc. Res. Tech. 2018, 82, 452–458. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; Dewaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Boil. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Barcode of Life “Dentifiying Species with DNA Barcoding. Available online: http://www.barcodeoflife.or (accessed on 1 June 2020).
- Quandt, D.; Müller, K.; Štech, M.; Frahm, J.; Frey, W.; Hilu, K.W.; Borsch, T. Molecular evolution of the chloroplast trnL-F region in land plants. Monogr. Syst. Bot. Mo. Bot. Gard. 2004, 98, 13–37. [Google Scholar]
- Borsch, T.; Hilu, K.W.; Wiersema, J.H.; Löhne, C.; Barthlott, W.; Wilde, V. Phylogeny of Nymphaea (Nymphaeaceae): Evidence from Substitutions and Microstructural Changes in the ChloroplasttrnT-trnFRegion. Int. J. Plant Sci. 2007, 168, 639–671. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Liu, X.; Yu, C.; Chen, Y.; Tang, H.; Zhang, L. Water lilies as emerging models for Darwin’s abominable mystery. Hortic. Res. 2017, 4, 17051. [Google Scholar] [CrossRef] [Green Version]
- Mäder, G.; Zamberlan, P.M.; Fagundes, N.J.R.; Magnus, T.; Salzano, F.M.; Bonatto, S.L.; Freitas, L.B. The use and limits of ITS data in the analysis of intraspecific variation in Passiflora L. (Passifloraceae). Genet. Mol. Boil. 2010, 33, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Stuessy, T.F.; Crawford, D.J. Flavonoids and phylogenetic reconstruction. Plant Syst. Evol. 1983, 143, 83–107. [Google Scholar] [CrossRef]
- Escriche, I.; Kadar, M.; Juan-Borrás, M.; Doménech, E. Using flavonoids, phenolic compounds and headspace volatile profile for botanical authentication of lemon and orange honeys. Food Res. Int. 2011, 44, 1504–1513. [Google Scholar] [CrossRef]
- Zhu, M.-Z.; Liu, T.; Zhang, C.; Guo, M.-Q. Flavonoids of Lotus (Nelumbo nucifera) Seed Embryos and Their Antioxidant Potential. J. Food Sci. 2017, 82, 1834–1841. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-S.; Wu, J.; Chen, L.-G.; Du, H.; Xu, Y.-J.; Wang, L.-J.; Zhang, H.-J.; Zheng, X.-C.; Wang, L.-S. Biogenesis of C-Glycosyl Flavones and Profiling of Flavonoid Glycosides in Lotus (Nelumbo nucifera). PLoS ONE 2014, 9, e108860. [Google Scholar] [CrossRef]
- Chen, S.; Wu, B.; Fang, J.-B.; Liu, Y.-L.; Zhang, H.-H.; Fang, L.; Guan, L.; Li, S. Analysis of flavonoids from lotus (Nelumbo nucifera) leaves using high performance liquid chromatography/photodiode array detector tandem electrospray ionization mass spectrometry and an extraction method optimized by orthogonal design. J. Chromatogr. A 2012, 1227, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zheng, X.; Shu, Q.; Li, H.; Zhong, P.; Zhang, H.; Xu, Y.; Wang, L.; Wang, L. Relationship between the Composition of Flavonoids and Flower Colors Variation in Tropical Water Lily (Nymphaea) Cultivars. PLoS ONE 2012, 7, e34335. [Google Scholar] [CrossRef] [Green Version]
- Yin, D.D.; Yuan, R.Y.; Wu, Q.; Li, S.-S.; Shao, S.; Xu, Y.J.; Hao, X.-H.; Wang, L.S. Assessment of flavonoids and volatile compounds in tea infusions of water lily flowers and their antioxidant activities. Food Chem. 2015, 187, 20–28. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Drouet, S.; Kabra, A.; Hano, C. Enrichment in Antioxidant Flavonoids of Stamen Extracts from Nymphaea lotus L. Using Ultrasonic-Assisted Extraction and Macroporous Resin Adsorption. Antioxydant 2020, 9, 576. [Google Scholar] [CrossRef]
- Ferreres, F.; Gil-Izquierdo, A.; Andrade, P.B.; Valentão, P.; Tomas-Barberan, F.A. Characterization of C-glycosyl flavones O-glycosylated by liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2007, 1161, 214–223. [Google Scholar] [CrossRef]
- Rodriguez-Saona, L.; Allendorf, M. Use of FTIR for Rapid Authentication and Detection of Adulteration of Food. Annu. Rev. Food Sci. Technol. 2011, 2, 467–483. [Google Scholar] [CrossRef]
- Baker, M.J.; Trevisan, J.; Bassan, P.; Bhargava, R.; Butler, H.J.; Dorling, K.M.; Fielden, P.R.; Fogarty, S.W.; Fullwood, N.J.; Heys, K.A.; et al. Using Fourier transform IR spectroscopy to analyze biological materials. Nat. Protoc. 2014, 9, 1771–1791. [Google Scholar] [CrossRef] [Green Version]
- Manasi; Mohapatra, S.; Rajesh, N.; Rajesh, V. Impact of heavy metal lead stress on polyamine levels in Halomonas BVR 1 isolated from an industry effluent. Sci. Rep. 2017, 7, 13447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Rosales, M.P.; Ferrol, N.; Roldán, M.; Belver, A.; Donaire, J.P. Cell wall acyl-lipids, proteins and polysaccharides in mature and germinated olive pollen. Rev. espanola de Fisiol. 1990, 46, 371–377. [Google Scholar]
- Thiers, B.; Thiers, B.H.; Cokic, B.B.B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff; New York Botanical Garden’s Virtual Herbarium: New York, NY, USA, 2009. [Google Scholar]
- World Health Organization. Quality Control Methods for Medicinal Plant Materials; World Health Organization: Geneva, Switzerland, 1998; ISBN 9241545100. [Google Scholar]
- Halbritter, H.; Ulrich, S.; Grímsson, F.; Weber, M.; Zetter, R.; Hesse, M.; Buchner, R.; Svojtka, M.; Frosch-Radivo, A. Illustrated Pollen Terminology; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Drouet, S.; Abassi, B.H.; Falguières, A.; Ahmad, W.; Sumaira; Ferroud, C.; Doussot, J.; Vanier, J.R.; Lainé, É.; Hano, C. Single Laboratory Validation of a Quantitative Core Shell-Based LC Separation for the Evaluation of Silymarin Variability and Associated Antioxidant Activity of Pakistani Ecotypes of Milk Thistle (Silybum Marianum L.). Molecules 2018, 23, 904. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Key Morphological Characters | N. nucifera * | N. lotus * |
|---|---|---|
| Leaf surface | pubescent | glabrous |
| Receptacle | torus present | torus absent |
| Ovary | superior | half inferior |
| Number of carpels | 1 | many |
| Placentation | marginal | pareital |
| Peak ID | Retention Time (min) | λmax (nm) | [M-H]− | Propable Identification | Commercial Standard | Reference |
|---|---|---|---|---|---|---|
| Nn#1 | 27.89 | 275, 354 | 479 | Myr-3-O-Glc | +(ES) | [25] |
| Nn#2 | 29.79 | 257, 354 | 609 | Quer-3-O-Rut | +(SA) | [25] |
| Nn#3 | 30.11 | 257, 353 | 477 | Quer-3-O-GlcA | +(SA) | [25] |
| Nn#4 | 30.23 | 266, 346 | 593 | Kae-3-O-Rob | +(SA) | [25] |
| Nn#5 | 30.73 | 266, 347 | 447 | Kae-3-O-Glc | +(SA) | [25] |
| Nn#6 | 31.93 | 269, 345 | 461 | Kae-3-O-GlcA | +(SA) | [25] |
| Nn#7 | 32.34 | 255, 356 | 477 | Iso-3-O-Glc | +(ES) | [25] |
| Nl#1 | 29.11 | 263, 349 | 479 | Myr 3-O-Gal | +(ES) | [27] |
| Nl#2 | 31.67 | 254, 305, 366 | 449 | Myr 3′-O-Xyl | +(ES) | [27] |
| Nl#3 | 32.26 | 257, 348 | 447 | Que-3-O-Rha | +(SA) | [27,28] |
| Nl#4 | 32.87 | 250, 366 | 433 | CNar-2′’-O-Gal | - | [27,28] |
| Nl#5 | 33.52 | 265, 343 | 447 | Kae-3-O-Gal | +(SA) | [27] |
| Nl#6 | 33.91 | 254, 366 | 433 | Que-3′-O-Xyl | +(SA) | [27] |
| Nl#7 | 34.71 | 268, 350 | 477 | Iso-7-O-Gal | - | [27,28] |
| Nl#8 | 35.51 | 252, 268, 352 | 447 | Iso-7-O-Xyl | - | [27,28] |
| Nl#9 | 36.57 | 265, 343 | 447 | Iso-3-O-Xyl | - | [28] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tungmunnithum, D.; Renouard, S.; Drouet, S.; Blondeau, J.-P.; Hano, C. A Critical Cross-Species Comparison of Pollen from Nelumbo nucifera Gaertn. vs. Nymphaea lotus L. for Authentication of Thai Medicinal Herbal Tea. Plants 2020, 9, 921. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9070921
Tungmunnithum D, Renouard S, Drouet S, Blondeau J-P, Hano C. A Critical Cross-Species Comparison of Pollen from Nelumbo nucifera Gaertn. vs. Nymphaea lotus L. for Authentication of Thai Medicinal Herbal Tea. Plants. 2020; 9(7):921. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9070921
Chicago/Turabian StyleTungmunnithum, Duangjai, Sullivan Renouard, Samantha Drouet, Jean-Philippe Blondeau, and Christophe Hano. 2020. "A Critical Cross-Species Comparison of Pollen from Nelumbo nucifera Gaertn. vs. Nymphaea lotus L. for Authentication of Thai Medicinal Herbal Tea" Plants 9, no. 7: 921. https://0-doi-org.brum.beds.ac.uk/10.3390/plants9070921