Antibacterial Activity of Chitosan Nanoparticles: A Review




1
Department of Food Science and Biotechnology, Sejong University, Seoul 05006, Korea
2
Department of Plant Biotechnology, Korea University, Seoul 02841, Korea
3
Department of Environmental Health Science, Konkuk University, Seoul 05029, Korea
*
Author to whom correspondence should be addressed.
Processes 2020, 8(9), 1173; https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091173
Submission received: 23 July 2020
/
Revised: 8 September 2020
/
Accepted: 9 September 2020
/
Published: 17 September 2020
(This article belongs to the Section Biological Processes and Systems)
Abstract
:In recent years, nanotechnology has attracted attention in many fields because it has several up-and-coming novel uses. Many researchers have suggested that chitosan nanoparticles (CS-NPs) and their derivatives are one of the best nanomaterials for delivering antibacterial activity. CS-NPs have a broad spectrum of antibacterial activity, but they manifest different inhibitory efficacy against gram-negative (G−) and gram-positive (G+) bacterial species. The mechanism of antibacterial action is an intricate process that varies between G− and G+ bacteria as a result of the differences in cell wall and cell membrane chemistry. In previous studies, greater antibacterial activity was more evident against G− bacteria than G+ bacteria, whereas in some studies G+ bacteria were more sensitive. Researchers predicted that the varied responses of bacteria are caused by the mixed hydrophilicity and negative charge distribution on the bacterial surface. Moreover, its activity depends on a number of variables including bacterial target (i.e., G− or G+ bacteria) and bacterial growth, as well as its concentration, pH, zeta-potential, molecular weight, and degree of acetylation. Therefore, this review examines current research on the mechanisms and factors affecting antibacterial activity, and application of CS-NPs specifically against animal and plant pathogenic bacteria.
1. Introduction
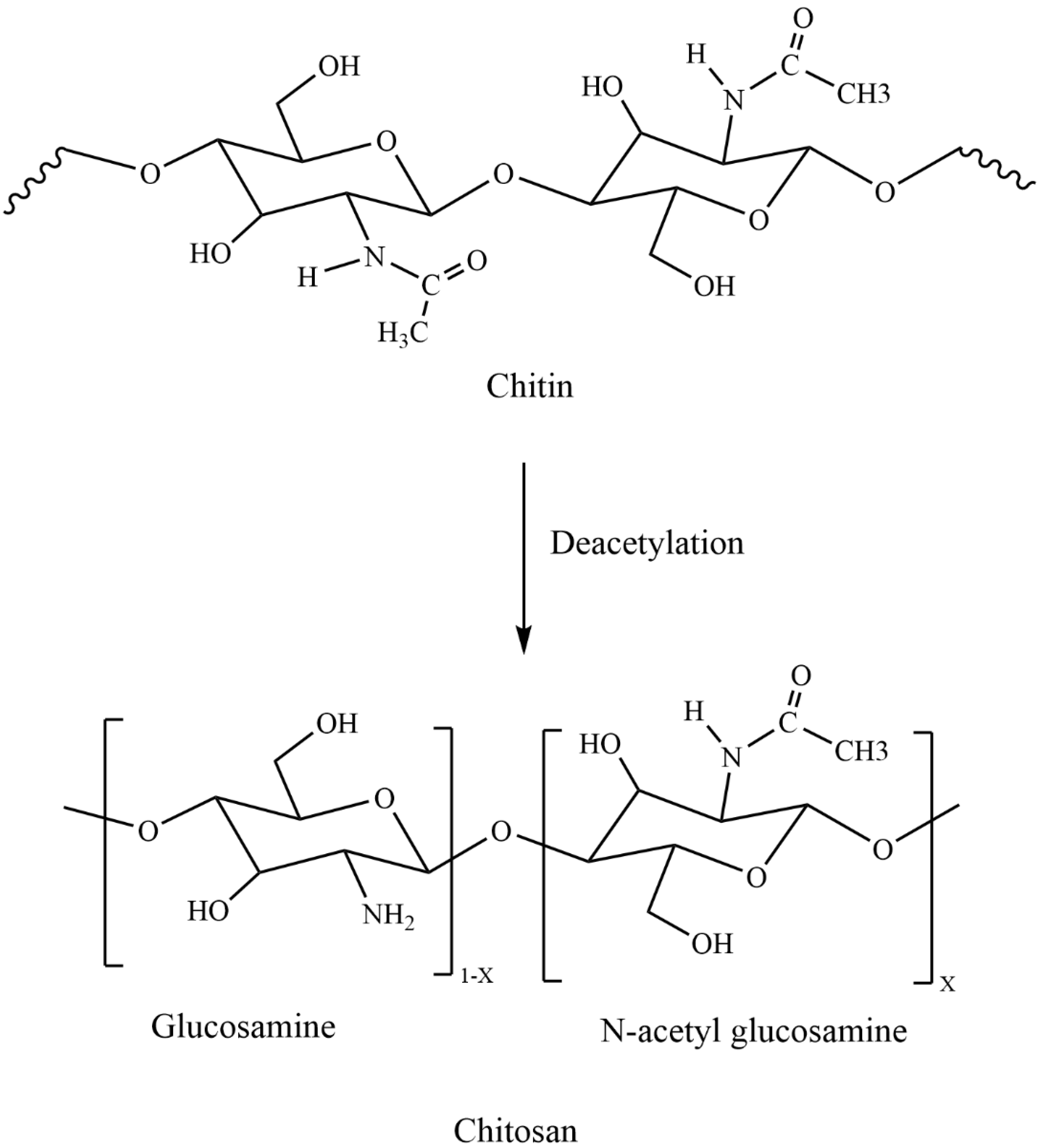
Chitosan (CS) is a natural cationic biopolymer composed of N-acetyl-D-glucosamine and D-glucosamine units connected by β-1,4-glycosidic linkages [1,2] (Figure 1). Previous studies have explored the antibacterial activity of CS [1,2,3,4], and more recently different types of CS derivates have been synthesized to enhance its natural antibacterial activity [5,6,7,8]. CS also exhibits other exceptional biological characteristics, such as biocompatibility, biodegradability, and nontoxicity. These have made it useful in a number of different industries such as medical, food, agriculture, textile, cosmetics and other industries [2,3,9]. Among incredible developments in biological technologies and chemical identification methods, nanotechnology has started to play an increasingly important role as it expands the ability to extend antibacterial studies to the atomic level [2]. Chitosan nanoparticles (CS-NPs) are a derivative of CS with excellent physicochemical properties [10,11,12,13,14]. CS-NPs have been prepared by several approaches, such as ionotropic gelation, microemulsion, emulsification solvent diffusion, polyelectrolyte complex and reverse micellar method [12]. Nanoparticles prepared using various materials and composites have shown antibacterial efficacy against different types of bacteria [15,16]. CS-NPs have exhibited improved biological activities such as antimicrobial [10,17], anticancer [18,19], anti-inflammatory and antioxidant activities [20,21]. Therefore, this review aims to summarize the mechanisms and factors affecting antibacterial activity and the application of CS-NPs against animal and plant pathogens.
2. Chitosan
CS is a linear polysaccharide which is obtained from chitin through demineralization and deproteinization and has an extensive array of medical and agricultural applications [1,2,3,4,22,23,24]. CS has drawn an undue pact of attention in different disciplines because of its exceptional biological properties, which include its biocompatibility [25,26], biodegradability [27], nontoxicity [28], and antimicrobial activity [1,2,29]. CS is the biopolymer that displays a cationic character due to the presence of its amino groups (NH2) [30,31]. Moreover, NH2 and hydroxyl (OH) groups of CS react with solutes present in the solution. However, from the adsorption point of view, the NH2 groups are more important than the OH groups and it is the NH2 groups only that decide the quality of this biopolymer. CS has been chemically modified either at the amino group or at the hydroxyl groups to synthesize derivatives possessing different moieties. According to Vinsová and Vavříková [6], the most frequent chemical modifications include quaternization, acylation, tosylation, Schiff base formation, O-carboxymethylation, N-carboxyalkylation, N-succinylation, and graft copolymerization. For example, CS connected with salicylic acid through an amide bond and produced a derivative called Salicyloyl-CS. Derivatives showed that the diameter of the inhibitory zone against Escherichia coli increased from 9.8 to 13.6 mm and from 21.5 to 27.5 mm against Staphylococcus aureus as the degree of substitution augmented from 0 to 43% [32]. Another study showed N-methylation of N-arylated CS derivatives had solubility at all pH ranges tested and exhibited inhibitory activity against S. aureus and E. coli [33]. These modifications make CS a promising candidate for medicine [34], food [35], cosmetics [36], and biomedical engineering industries [37], as well as for sustainable agriculture [38,39]. CS is an insoluble biopolymer in water, but soluble in acidic solutions at pKa approximately 6.3 or below. At this pH, glucosamine units (-NH2+) of CS convert into the protonated soluble form (-NH3+). Therefore, CS solubility is contingent on its source, molecular weight (MW), and degree of acetylation (DA). Moreover, usage of CS has been restricted to its native form due to its porosity, surface area, and low solubility at neutral pH. To improve activities and functionality, several CS derivatives have been synthesized and evaluated for pioneering applications [5,8,40].
3. Chitosan Nanoparticles (CS-NPs)
Now a days, nanotechnology has attracted attention in various fields as a result of its outstanding potential for novel usage (Figure 2).
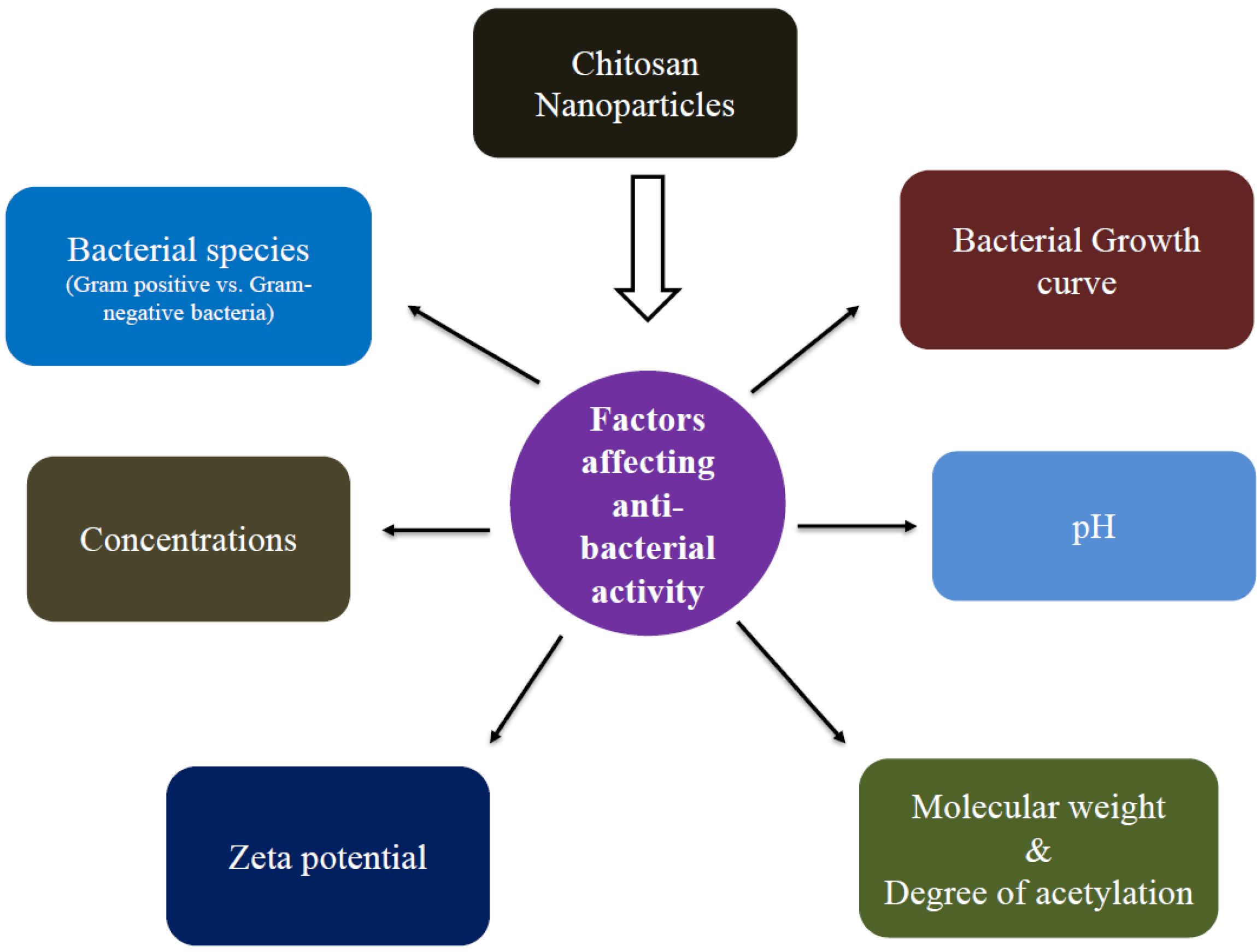
The smaller size of NPs has manifested a significant change in its physical properties compared to its original counterpart. CS-NPs share features of CS and valuable assets of NPs such as small size, increased surface area, and quantum size effects [10,11,12,13,14]. CS-NPs were produced by several procedures such as ionic gelation [41], reverse micellar method [42], microemulsion [43], emulsion droplet coalescence [19], and spray drying [44]. Among several approaches, physical crosslinking by ionic gelation was found to be the best one. The ion gelation method exploits the electrostatic communication between a positively charged group of CS and a negatively charged group of tripolyphosphate (TPP). By changing the ratio of CS to TPP, the size and surface charge of NPs could be modified. Moreover, there is no chemical used for cross-linking, which reduces the toxic side effects. Recently, different types of metal ions and CS complexes (CS-Ag+NPs, CS-Cu2+NPs, CS-Zn2+NPs, CS-Mn2+NPs, and CS-Fe2+NPs) have been made to improve their antimicrobial activity [45,46,47,48]. Moreover, in further pursuit of antimicrobial efficacy, hybrid CS-NPs with protamine [49], lysozyme [50], oleic acid-grafted chitosan oligosaccharide (CSO-OA) NPs [51], essential oil [52], and curcumin [53] NPs, among others, have also been tested. Thus, polycationic CS-NPs with more surface charge density communicate with bacteria to a larger extent than CS alone [54]. Also, CS-NPs interrupt the cell wall and membrane of bacterial cells, which leads to the efflux of intracellular molecules and bacterial cell death. Several parameters control the size of NPs and consequently affect antibacterial responses (Table 1). These include the bacterial species, growth stage, concentration, pH, zeta-potential, MW, and DA.
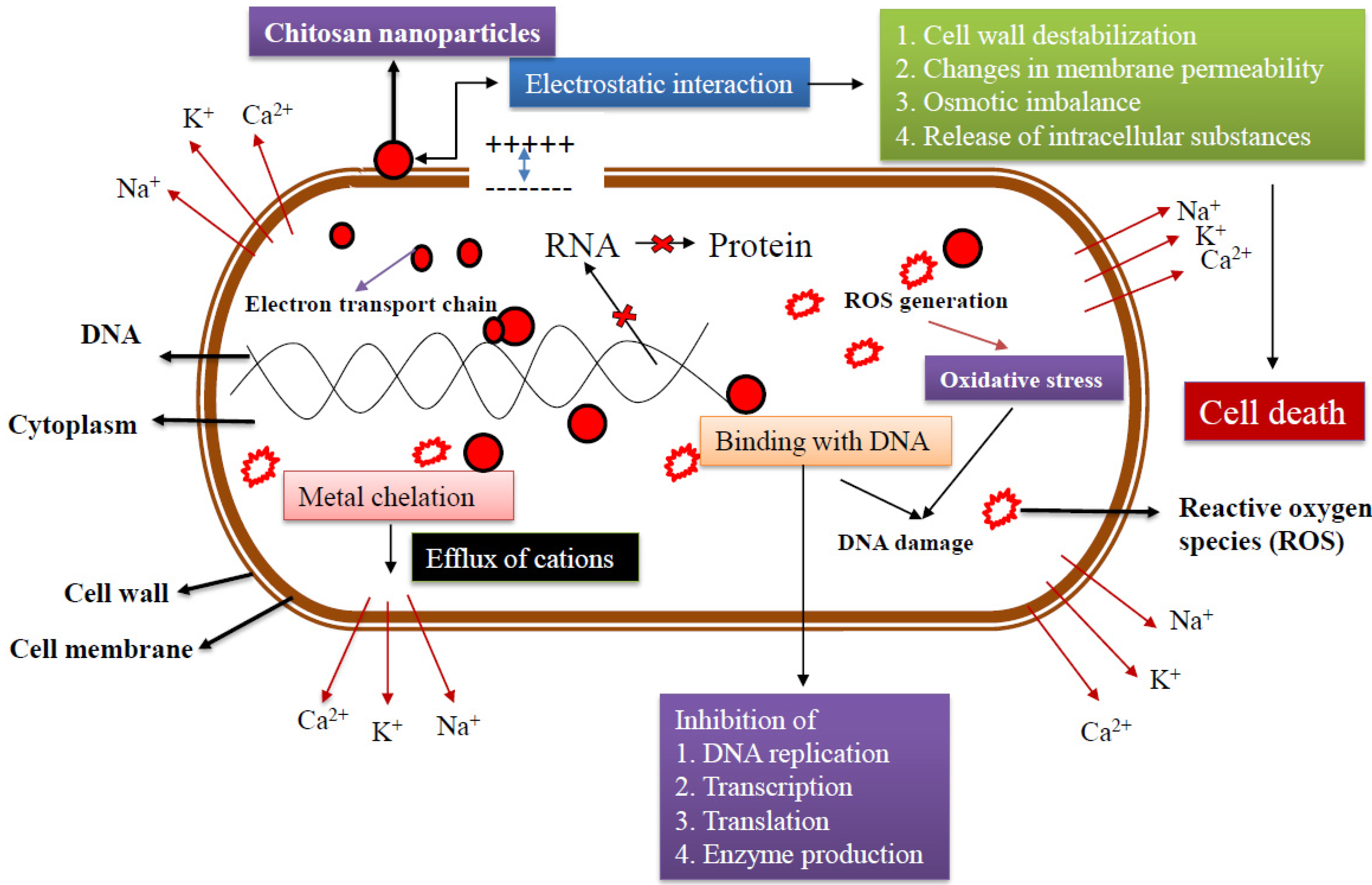
4. Mechanism of Antibacterial Action
The mechanism of antibacterial activity of CS-NPs is probably on account of communications with either the bacterial cell wall or the cell membrane. In order to elucidate this mechanism, different hypothesis have been proposed (Figure 3). The most widely recognized CS-NPs model of antimicrobial action is the electrostatic communication between the amino groups of glucosamine (positively charged) with the cell membranes (negatively charged) of bacteria [59]. This interaction initiates prevalent variations to the surface of the cell, leading to a modification in membrane permeability which sequentially incites osmotic imbalance and efflux of intracellular substances that result in cell death [60,61,62].
The electrostatic force between the CS and bacteria cell wall endorses a closer interaction with charged molecules that leads to the infiltration of CS-NPs through the bacteria cell wall [63]. Consequently, the possibility of CS-NPs accumulating at the place of interaction increases. In addition, CS-NPs is capable of modifying the electron transport chain of bacteria [64,65]. The most predominant proposed antibacterial activity of CS is electrostatic interactions which change the membrane’s permeability. It then binds with DNA and ruins DNA replication, leading to bacterial cell death. In addition, it has been shown that lower MW-CS can enter the cell, bind to DNA, and inhibit replication machinery [65]. Flocculation of electronegative elements by CS in the cell upsets the physiological activities of the bacteria and leads to bacterial cell death [66].
One more likely mechanism is the chelating capacity of CS towards metal ions, which stimulates toxin production and prevents bacterial viability [10]. CS retains greater chelating activity for various ions of metal (including Fe2+, Mg2+, Ni2+, Co2+, Cu2+, and Zn2+) in acid conditions. Metal ions that are associated with molecules of the bacteria cell wall are vital for their stability. Thus, CS-mediated chelation of such metal ions has regularly been concerned as a conceivable mechanism of antibacterial action. Generally, such a mechanism is more effective at high pH where positive ions are seized by CS since the NH2 groups are unprotonated and the electron pair on the amine nitrogen is accessible for contribution to metal ions. Indisputably, CS molecules surround the metal complex and impede vital nutrient flow, leading to cell death. Therefore, the appropriate deployment of CS-NPs is reliant on many factors that can be altered [67].
5. Factors Affecting Antibacterial Activity
The antibacterial activity of CS-NPs is triggered by several factors, including bacterial target (gram-negative (G−) vs. gram-positive (G+) bacteria), growth stage, zeta potential, concentration, pH, MW, and DA (Figure 4). It is known that CS and CS-NPs antibacterial activity is tended by several factors which affect it in a tidy and sovereign way.
5.1. Bacterial Species (Gram-Negative (G−) vs. Gram-Positive Bacteria (G+))
In general, it is believed that if the tested NPs do not kill but inhibit the growth of bacteria (bacteriostatic), bacteria taken out of the NPs solution will form bacterial colonies. If the studied NPs are bactericidal, no bacterial colonies should be spotted. Generally, CS-NPs act as bactericidal or bacteriostatic, often with no distinction between activities. The minimum inhibitory concentration (MIC) is the lowest concentration (mg/mL) at which a tested compound inhibits bacterial growth and the minimum bactericidal concentration (MBC) is the lowest concentration (mg/mL) at which a tested compound kills more than 99% of the added bacteria. A ratio with a value superior to 1 (MBC/MIC > 1) designates that an excessive quantity of compound is required to influence the bactericidal result and this NP is considered a bacteriostatic compound. CS-NPs have a wide spectrum of antibacterial activity, but they manifest different inhibitory efficacy against G− and G+ bacterial species. The bactericidal effectiveness on G− or G+ bacteria is, however, somewhat controversial. The mechanism of antibacterial action is a multifaceted process that varies between G− and G+ bacteria because of the differences in cell wall and cell membrane chemistry. In previous studies, greater antibacterial activity was more evident against G− bacteria than G+ bacteria [55,67,68,69], whereas in some studies G+ bacteria were highly sensitive, probably as a result of the G− bacterial cell membrane [70]. The cell membrane of G− bacteria contains lipopolysaccharide (LPS) which is comprised of anionic groups (phosphate and pyrophosphate groups) which provides more negative charges to the cell surface than that observed for G+ bacteria which is made of peptidoglycan (PG) and teichoic acid (TA). This provides a strong rationale for why the loss of intracellular contents observed with CS-NPs in G− is greater than that stated in G+ bacteria [22,71,72,73].
According to Perelshtein et al. [74], CS-NPs treated on cotton showed minimal activity against E. coli (G− bacteria) but was satisfactory against Enterococcus faecalis (G+ bacteria). Zn–CS-NPs showed great levels of antibacterial activity against both bacteria. However, the G− bacteria was less affected by CS than the G+ bacteria. Mubarak Ali et al. [21] studies also showed that CS-NPs had a robust antibacterial effect on medically important species, Staphylococcus aureus, and E. coli. Devlieghere et al. [75] produced data showing G− bacteria were less affected while the impact on G+ bacteria differed. Whereas Katas et al. [49] reported that CS-Au-NPs antibacterial activity was more effective against G− bacteria than G+ bacteria. Venkatesham et al. [76] reported that green synthesized Ag-NPs from CS had greater antibacterial activity against E. coli and Micrococcus luteus. They also stated that these CS-Ag-NPs had higher antibacterial activity on G− (E. coli) than G+ (M. luteus) bacteria. Rasaee et al. [77] also found that G− bacteria (E. coli) were more sensitive to CS-NPs compared to G+ bacteria (B. vallismortis). Significant (p < 0.05) antibacterial activity against Streptomycin species, Pseudomonas fluorescence, and S. aureus has been identified. CS-NPs also showed a more inhibitory effect on P. fluorescence compared to other species and positive controls [20]. In addition to antimicrobial and antivirulence activity, CS-NPs disturbed the permeability of the outer membrane of the G− ubiquitous opportunistic pathogen P. aeruginosa to various types of reagents [78].
Numerous metal ions and experimental conditions support the synthesis of CS-NP derivatives. Chen et al. [46] showed Ag-CS-NPs had a higher antibacterial activity than CS-NPs against both G+ (S. aureus and Bacillus subtilis) and G− (E. coli and Salmonella choleraesuis) bacteria. For B. subtilis, the MBC of Ag-CS-NPs was only 0.128 mg/mL, while that of CS-NPs was 1.28 mg/mL. Thus, the bactericidal activities of Ag-CS-NPs were ten times higher than CS-NPs. Wei et al. [79] also confirmed that the CS-based Ag-NPs had the highest antibacterial activity against G+ (B. subtilis and S. aureus) and G− (E. coli) bacteria. Compared to CS-NPs, Ag-CS-NPs had both quick and long-lasting antibacterial efficiency against E. coli. Another study also showed that the Ag/CS nanocomposite had a significant antibacterial effect on both G− and G+ bacteria [72]. Badawy et al. [47] also found E. coli was more susceptible to CS-Ag-NPs than S. typhimurium. A study by Venkatesham et al. [78] revealed green synthesized Ag-NPs had significant antibacterial action on E. coli and M. luteus bacteria. These NPs also had significantly more activity against G− (E. coli) than G+ (M. luteus) bacteria.
Du et al. [55] reported that the MIC of CS-NPs against E. coli, S. enterica serotype Choleraesuis, and S. aureus were three to four-fold lower than CS. According to Du et al. [55], except for Fe2+ CS-NPs, CS-NPs loaded with metal ions displayed more antibacterial activity than CS, CS-NPs, and related metal ions alone. It was also noted that CS-NPs-loaded Ag+ exhibited the highest antibacterial activity. The MIC and MBC of Cu2+-loaded CS-NPs against G− and G+ bacteria were 21–42 times lower than that of Cu2+ alone. Manikandan and Sathiyabama [80] also stated that while the CS stabilized Cu-NPs exhibited inhibitory activity towards G− (E. coli, S. paratyphi) and G+ bacteria (Bacillus sp.), G− bacteria was more sensitive to Cu-CS-NPs compared to G+-bacteria. Interestingly, Qi et al. [22] also confirmed the MIC of CS-NPs against E. coli, Salmonella, and S. aureus and showed that only 0.25 to 4 μg/mL of CS-NPs was sufficient to impede the growth of bacterial species (Table 2).
Curcumin-CS-TPP NPs showed the highest zone of inhibition against S. aureus indicating that inhibitory capacity was higher for G+ bacteria than G− bacteria (E. coli) [16]. The MIC value was recorded for growth inhibition of both species and was found to be 12.5 μg/mL and 20 μg/mL for G+ (S. aureus) and G− (E. coli), respectively. Tamara et al. [47] confirmed that adding protamine in the CS-NPs enhanced the antimicrobial specificity and MIC value on E. coli (31.25 µg/mL) and B. cereus (>250 µg/mL). The addition of protamine in the hybrid CS-NPs changed the bacterial cell wall structure and membrane permeability. Moreover, hybrid CS-NPs showed lesser binding affinity to B. cereus than E. coli. Thus, the hybridization of CS with protamine upgraded the antibacterial activity of CS-NPs against pathogenic E. coli, but its effect on B. cereus was significantly reduced.
Tran et al. [73] found a bactericidal action against all strains of G− (E. coli and P. aeruginosa), and G+ bacteria (Lactobacillus fermentum, S. aureus, and B. subtilis) with an MBC/MIC = 1. Moreover, G− bacteria were inhibited more strongly than G+ bacteria. The thinner cell wall of G− bacteria makes them more susceptible to CS-NPs. G+ bacteria have a thicker cell wall because of a peptidoglycan layer that can obstruct the inward bound of CS-NPs [48,76]. Wu et al. [50] showed the addition of lysozyme (Lys) into CS-NPs improved the antibacterial activity against both G− and G+ bacteria. In addition, a transmission electron microscopy morphological study revealed that when E. coli was treated with CS-NPs there was damage in the cell membrane and outflow of some cytoplasmic constituents, whereas CS-Lys-NPs treated cells were changed from round shape to irregular shape and completely ruptured. Also, the antibacterial activity against G+ bacteria (B. subtilis) may be attributed to the impact of metabolic enzyme activities and restrictions on bacterial metabolism. Hence, the diverse metabolic changes of bacteria are formed by means of the negative charge distribution and mixed hydrophilicity on the bacterial surface. Therefore, increases in CS absorption would result in more changes to cell wall morphology and membrane permeability. The greater antimicrobial activity of CS-NPs related with CS was constant across several species of bacteria. This suggests that the mechanism of antibacterial action and effectiveness is specifically dependent on the type of bacterial species and CS derivatives.
5.2. Bacterial Growth Stage
For certain bacterial species, the growth stage can impact antibacterial efficiency of nanoparticles. Recently, Orellano et al. [82] reported that a notable decrease in colony-forming units (CFUs) of S. aureus was observed after 2 h of co-incubation with CS-NPs, whereas 4 h were required to achieve the same results with CS (p < 0.05). Similar results were observed for S. xylosus. The cell viability of S. mutans decreased significantly at 3 h and 18 h after incubation, along with increasing concentrations of CS-NPs [83]. When exploring the antibacterial activity of CS and its derivatives, most bacterial species are in the late log phase. However, some investigators have confirmed that bacterial growth curves affect the sensitivity of bacteria to CS-NPs. Yang et al. [84] revealed the consequence of growth phase on the susceptibility of E. coli O157: H7 to CS. They discovered that the mid-exponential phase was more susceptible, followed by the late-exponential phase. Moreover, cell surface negativity of the bacterial cell changes as growth stage changes and it incites the susceptibility of bacterial cells toward CS-NPs [52]. Therefore, a diverse surface charge on bacterial cells could possibly result in a difference in susceptibility.
5.3. Zeta Potential
Zeta potential is a vital parameter for stability in aqueous nanosuspensions. Studies have shown that CS-NPs with higher zeta potential values had greater effects on inhibition of bacterial growth. According to Nguyen et al. [56], reduced size and higher zeta potential of CS-NPs resulted in higher antibacterial activity. In contrast, Tamara et al. [49], found that increasing the concentration of CS was positively correlated with the size of NPs. Regardless, differences in zeta potential values were not readily obvious among NPs.
Du et al. [55] presented data showing that the potency of antibacterial activity was correlated with the zeta potential of metal associated CS-NPs. They found that the zeta potential of CS-NPs was lower than those of hybrid CS-NPs due to the addition of metal ions. The purpose may be derived from the positive charge carried by metal ions. Zeta potential is used to establish cellular interaction with charged ions or molecules. The presence of negatively changed ions decreases the zeta potential whereas positively charged ions increases the zeta potential of the surface. The zeta potential of the NPs loaded with Ag+ was highest due to its positively charged NP surface and its relation to mass, followed by Cu2+, Zn2+, and Mn2+. The NPs loaded with Fe2+ had the lowest zeta potential due to its surface charge and low molecular mass, but this was still higher than for CS-NPs. The antibacterial activity of CS-NPs loaded with metal ions was directly proportional to the zeta potential. Thus, zeta potential could easily be associated with antibacterial activity.
Qi et al. [22] also revealed that Cu2+-loaded NPs zeta potential is markedly higher (96 mV) than that of CS-NPs (51 mV). Moreover, Cu2+-loaded CS-NPs had higher antibacterial activity than native CS, and a little higher than CS-NPs. Therefore, the negatively charged bacterial cell membrane had a high affinity for Cu-loaded NPs, which is responsible for their higher antibacterial activity. Another study showed that CS-NPs loaded with Fe2+ had higher antimicrobial activity than Fe3+ loaded CS-NPs. They found that the zeta potential of CS (+45.88 mV) is higher than those for CS-NPs loaded with Fe2+ and Fe3+ (+28.82 and +28.26 mV, respectively), but its bactericidal performance was inferior [43]. As the Fe-loaded CS-NPs is a cationic polymer, it can attach to the negatively charged E. coli cell wall by electrostatic interaction and upset the cell membrane, which leads to the release of intracellular molecules, thus killing the bacteria cells.
5.4. pH
CS-NPs have a polycationic nature due to protonation of the −NH2 group at the C-2 position of D-glucosamine under acidic conditions [5,51,61,85]. CS displays its antibacterial activity only in an acidic pH due to its meager solubility above pH 6.5. In contrast to CS, CS-NPs could be well spread in aqueous solutions and a smaller amount pretentious by the pH. The antibacterial activity of CS was inversely proportional to pH, with more activity at lower pH levels [67]. Xing et al. [85] reported that the antibacterial activity of OCS-NPs increased as the pH increased (4.0 to 6.0) and resulted in higher activity at pH 6.0 for both G− (E. coli) and G+ (S. aureus) bacteria. P. aeruginosa showed significant antimicrobial effects at acidic pH 5.0 for CS-NPs, however the activity was not found at a pH > 7.0 [80]. Tsai and Su [59] showed the bactericidal effect of CS against E. coli was increased at an acidic pH. Moreover, the antibacterial activity of CS against foodborne pathogens (S. aureus, E. coli, Y. enterocolitica, L. monocytogenes, S. typhimurium) was stronger at pH 5.5 than at pH 6.5 [86].
Gomes et al. [87] stated that the MIC values estimated at pH 7.0 and pH 5.0 for CS medium MW (CS-M-MW) were 0.47 and 0.40 mg/mL, respectively, whereas for CS-M-MW30 the values ranged from 0.40 to 0.28 mg/mL. When considering CS low MW (CS-L-MW), values of 0.30 and 0.33 mg/mL were obtained, whereas for CS-L-MW30, the MIC values were 0.30 and 0.20 mg/mL, in pH 7.0 or 5.0, respectively. Manikandan and Sathiyabama, [80] reported that CS-Cu-NPs showed only a small zone of inhibition against both G− and G+ bacterial species at pH 7.4 suggesting a lack of antibacterial activity at higher pH values.
Huang et al. [51] discovered the effect of pH (4.0, 7.0, and 10.0) on the antibacterial activity of oleic acid-grafted CS oligosaccharide (CS-O-OA) NPs. The antibacterial activity of NPs improved upon increasing the pH of NPs from 4.0 to 7.0 and reduced with a further increase to 10.0 for both G− and G+ bacteria. Divya et al. [11] showed K. pneumoniae had the highest susceptibility at pH 7.5 whereas for S. aureus it was at pH 5.5. A MIC value of 0.2 mg/mL was observed at pH 5.0 for CS-L-MW30 which confirmed the presence of charged amine groups [85]. Thus, CS protonation caused its dispersion, leading to more effective activity against E. coli and the MIC values were shown to be pH dependent.
5.5. Concentrations
CS-NPs increased the antibacterial activity as the concentration increased [48,51,88,89]. Xing et al. [85] found that increasing the concentration of OCS-NPs lead to an increase in antibacterial activity, like its source, CS [90]. Sanpui et al. [91] studied the effects of CS-Ag NP composite concentrations on green fluorescent protein expressing E. coli and found that growth was completely inhibited at ≥100 μgmL−1 (MIC) while a concentration of 120 μgmL−1 was found to be the MBC. Moreover, 0.012% of CS in CS-Ag-NP composite preparations was sufficient to inhibit the growth of recombinant E. coli [91]. Mohammadi et al. [89] showed that inhibitory effects increased with increasing concentrations (from 0.009% to 0.15% w/v) of CS and CS-NPs and was significantly (p < 0.05) correlated to the type of CS-NPs and bacteria. Another study showed that CS at 0.8% inhibited the growth of Aeromonas hydrophila, while CS at 0.4% was required to prevent the growth of Edwardsiella ictaluri and Flavobacterium columnare [56]. Alarfaj et al. [92] studied five different concentrations of CS-NPs (10, 20, 50, 100, and 150 µg) against G+ (Bacillus sp.) and G− bacteria (Pseudomonas sp.). They found that the higher concentrations of CS-NPs (100 and 150 µg), inhibited both bacteria. Divya et al. [11] evaluated the CS-NPs antibacterial activity against four different strains (K. pneumoniae, P. aeruginosa, S. aureus, E. coli) and found that activity improved as the concentration of CS-NPs increased. All the CS-NPs had high inhibition rates against tested microorganisms. In another study, the CS-NPs prepared from CS (200 kDa) at different concentrations were found to have a similar MIC against E. coli and B. cereus [50].
Katas et al. [48] also found that CS-Au-NPs by variable concentrations of CS (0.2%, 0.4%, 0.6%, 0.8%, 1.0%, and 1.2% w/v) affected antibacterial activity of both S. aureus, and P. aeruginosa. The largest zone of inhibition was found at 1% (w/v) for S. aureus while P. aeruginosa was inhibited at 1.2% (w/v) of CS. In addition, the in vivo antibacterial activity of CS-Ag-NPs against E. coli in minced meat was concentration-dependent and better than controls [47]. A previous study revealed that this is due to the electrostatic static interactions between the electron-rich oxygen atoms of the polar hydroxyl and ether groups of CS and the electropositive transition cations (Ag+) [91]. Moreover, AgNPs also bind with the cell wall via thiol-containing proteins, some of them penetrating the cell wall and changing the membrane permeability. This leads to the efflux of proteins and other intracellular constituents and prevents bacterial growth. Anitha et al. [88] studied the effect of CS, O- carboxymethyl (O-CMC) and N, O-carboxymethyl (N-OCMC) NPs against S. aureus. They revealed that CS-NPs showed less antibacterial activity compared to O-CMC and N-OCMC NPs. Antibacterial potency of NPs augmented with concentration and N-OCMC NPs exhibited maximum antibacterial activity. Huang et al. [51] studied the outcome of CS-O-OA NPs against E. coli and S. aureus based on concentrations and found the same results. These studies all confirmed that antibacterial activity was augmented as the concentration of NPs increased.
5.6. Influence of Molecular Weight and Degree of Acetylation
Previous studies showed that the antibacterial activity of CS and CS-NPs depends a lot on its MW and DA [1,2,22,49,56]. MW and DA amend the antibacterial activity of CS and CS-NPs separately, but MW still has a larger influence than DA [1]. The MW modifications allowed for satisfactory changes in the antibacterial ability of the synthesized NPs [87]. They concluded that CS-NPs at low and medium MWs can strongly inhibit pathogen growth and that the inhibitory ability can be controlled by pH and MW. These data showed that an increase in MW of CS caused a rise in size and decreased the zeta potential, hence reducing antibacterial activity. A decrease in MW of CS produced smaller sized particles and increased zeta potential. Thus, higher zeta potential supports NPs to attach and communicate with the bacterial cell membrane. The variation in MW and particle size/zeta potential allowed easy manipulation of the NPs physicochemical properties.
Other studies have also confirmed that the size and zeta potential of CS-NPs and their activity against bacterial species depends greatly on MW [57,70,93]. De Paz et al. [94] showed that when treating S. mutans with different MW and DA forms, high MW CS-NPs had decreased antimicrobial activities (20% to 25% of cells damaged) compared to CS-NPs synthesized with low MW (>95% of cells damaged). Increasing the MW of CS material led to an increase in the NPs size and a decline in the zeta potential value [56]. Another study also showed that higher MW CS-NPs had reduced antibacterial activity against S. pneumoniae. They also found that lower MW CS-NPs had a higher zeta potential and higher antibacterial activity. According to Gan et al. [93], when increasing the MW of CS, the size of the NPs increased, and zeta potential decreased. Ngan et al. [57] also found that CS-NPs synthesized from different MWs (low, medium, and high) by spray drying method had an increase in size and a reduction in zeta potential. It led to an increase in turbidity of S. aureus. A previous study suggested that increasing the MW of the CS led to an increase in the average size of the CS-NPs and a reduction in the zeta potential. Moreover, the antibacterial activity of the CS-NPs increased with decreasing MW. Therefore, lower MW CS formed particles of reduced size and increased zeta potential, which led to greater inhibition of bacterial growth. Devlieghere et al. [87] investigated the antibacterial activity of CS with MW of 43 kDa and a DA of 94%. They found that G− bacteria were less affected while the effect on G+ bacteria varied. One more study explored the medium MW CS activity and found it to be effective against G− pathogens of warm water fish [95].
Mohammadi et al. [89] demonstrated that NPs produced to achieve MW modifications such as low MW CS (MW of 70 kDa, DA of 75–85%), and medium MW CS (MW of 190 kDa, DA of 75–85%) had a reasonable variation in the antimicrobial ability. They found a positive correlation between the particle size of CS-NPs and MW of CS, in which the size of NPs increased when a higher MW was used. Reductions in the OD of bacterial cell cultures depends on the type of CS, the type of CS-NPs, and the bacterial species. It is challenging to define an optimal MW for maximal antimicrobial activity. The selection of MW for CS may depend on the intended application. Moreover, medium MW-CS and low MW-CS showed lower MIC and MBC values compared with related microparticles only for P. fluorescens and E. coli, respectively. In contrast, low MW-CS-NPs had improved inhibitory effects against E. carotovora compared to medium MW-CS-NPs. This finding agreed with other research which proved that CS-NPs displayed more activity as a result of specific characteristics, such as small size and more surface charge [22]. The same efficiency was also revealed by low-MW CS cross-linked with sodium sulfate against E. coli O157: H7 [96].
According to Xing et al. [85], OC-NPs synthesized from low MW-CS (38 kDa) had the most effective antibacterial activity against E. coli. This proved that increasing the MW of CS for OC-NPs led to reduced antibacterial activity. However, this tendency was not detected with G+ bacteria (S. aureus). Thus, the impact of CS-MW on the antibacterial activity of OC-NPs against S. aureus was not as noticeable as that seen for E. coli. Alqahtani et al. [97] prepared non-antibiotic DIC-loaded CS-NPs (DIC-CS-NPs) from low and high MW CS. The low MW DIC-CS-NPs and high MW DIC-CS-NPs had inhibition zones of 18 and 15 mm, respectively, against S. aureus and 21 and 16 mm, respectively, against B. subtilis. These results confirmed that the degree of antimicrobial activity exhibited by DIC-CS-NPs depended on the MW of the CS. The studies of Honary et al. [98] also proved that the size of NPs was determined by varying the MW of CS (100, 400, and 600 kDa). The antibacterial activity of the NPs against S. aureus increased with a decrease in particle size owing to an increase in surface area. Moreover, Sarwar et al. [81] studied the antibacterial activity of both low and high MW CS-NPs against six different bacterial strains and found that low MW CS-NPs had better inhibitory effects than high MW CS-NPs. They suggested that surface area rises with reduction in particle size. It might be the key factor in the augmented antibacterial activity of small size nanoparticles.
Greater positive charge density of CS takes the lead with respect to better antimicrobial activity. Also, positive charge density is positively correlated with the DA of CS [2]. It was confirmed that CS with a higher DA (97.5%) was more effective than CS with a lower DA (83.7%) at the same MW [99]. According to Regiel et al. [100], pure CS films (low/medium/high MW), showed a clear bacteriostatic effect against S. aureus strains. Whereas Ag-CS-NPs clearly showed a bactericidal effect dependent on MW of CS, the medium MW-CS was found to be a robust bactericidal agent. They also studied the influence of the bacterial concentration and time. Low and high MW-CS showed similar DA, but medium MW-CS showed the highest DA. The finest outcomes were found with high DA, leading to smaller NPs. As mentioned earlier, the DA determines the solubility and charge development, where the –NH2, and –OH groups in CS are considered the regulatory reactive sites.
6. Applications
6.1. Antibacterial Activity against Animal Pathogens
CS-NPs have a wide spectrum of antimicrobial activity against different pathogenic bacteria (Table 3). Ikono et al. [79] studied CS-NP effects on the dental caries-associated microorganism, S. mutans. After CS-NP treatment, the viability of cells decreased significantly with increasing concentrations of nanoparticles. Covarrubias et al. [101] also studied antibacterial properties of Cu-CS-NPs against S. mutans. Cu-CS-NPs showed higher MIC and MBC values than Cu-NPs [102,103]. It is specifically exciting to note that Cu-CS-NPs were proficient at disturbing adhesion and biofilm formation by S. mutans [104,105]. Cu-CS-NPs displayed greater capacity to inhibit S. mutans growth on the human tooth surface as well as to disrupt the dental biofilm and had a bactericidal effect [101]. The bactericidal properties revealed by Cu-CS-NPs could make it suitable for further investigation of treatments for dental plaque.
Hipalaswins et al. [106] demonstrated antibacterial activity of synthesized CS-NPs against clinically pathogenic bacterial strains P. fluorescens MTCC 1748, Proteus mirabilis MTCC 1429, S. aureus MTCC 7443, K. pneumoniae MTTC 109, E. coli MTTC 1687, and Enterobacter aerogenes MTCC 111. E. aerogenes were found to be most susceptible followed by E. coli, K. pneumoniae, P. fluorescens, and P. mirabilis. The CS-NPs were found to be less toxic towards S. aureus. The results of the antibacterial activity studies revealed that the CS-NPs inhibited G− bacteria more efficiently than G+ bacteria. Nguyen et al. [56] conducted antibacterial activity studies against antibiotic resistant G+ S. pneumoniae using CS-NPs and a CS-NP–amoxicillin complex. The CS-NP–amoxicillin inoculation exhibited increased antibacterial activity compared to amoxicillin and CS-NPs alone. Indeed, it was found to be three-fold better than amoxicillin and completely inhibited the growth of S. pneumoniae. According to Sotelo-Boyás et al. [52], CS-NPs incorporated with lime essential oil exhibited antibacterial activity against four food-borne pathogens (S. aureus, L. monocytogenes, Shigella dysenteriae, and E. coli). S. dysenteriae is highly sensitive to CS-NPs incorporated with lime essential oil and showed the highest susceptibility [52]. CS-NPs used as nanocarriers for the treatment of bovine mastitis caused by S. aureus and coagulase-negative staphylococci [107,108].
CS-NPs exhibited promising antimicrobial activities against pathogenic microorganisms, K. pneumoniae, E. coli, S. aureus, and P. aeruginosa. CS-NPs showed greater antimicrobial activity against all pathogens compared to chitin and CS [10]. Tamara et al. [49] data showed that hybridizing CS with protamine enhanced the antibacterial activity of CS-NPs towards pathogenic E. coli, but their effect against B. cereus was significantly reduced. In addition, curcumin-loaded CS-NPs could be used in drug delivery and useful as an approach to precisely activate antibacterial systems. Curcumin loaded CS-NPs significantly repressed the in vivo growth of P. aeruginosa and S. aureus infections in mice [53]. Tran et al. [73] synthesized and characterized Ag-NPs from CS and evaluated their effect against both G− (E. coli and P. aeruginosa) and G+ (Lactobacillus fermentum, S. aureus and B. subtilis) bacteria.
AMP temporin B (TB)-CS-NPs were studied against four clinical isolates of S. epidermidis. Within 24 h of incubation, CS-NPs demonstrated bactericidal activity and markedly reduced the initial inoculum. Subsequently, TB was distributed to the bacterial surface and further reduced the viability of cells [58]. Pilon et al. [113] disclosed that nanoparticulated CS (CS-NPs) used as a coating inhibited the growth of mesophilic microorganisms compared to the conventional coating. The CS-based NPs (110 nm) coating was found to be more effective in inhibiting microbial growth compared to those of conventional coating (300 nm). As a result, there was no pathogenic, Salmonella sp., or fecal coliforms. This study supported the possible use of CS-NPs as edible coatings for controlling bacterial growth in fresh vegetables and fruits. Therefore, the smaller the NPs, the greater its motion and surface interaction, resulting in increased antibacterial activity against animal pathogens.
6.2. Antibacterial Activity against Plant Pathogens
CS-NPs may promote a deep and steady outcome on plant growth and protection. As a multipurpose material, CS has been exploited to reduce disease severity of phytopathogens [114,115] or to boost plant innate immunity [116,117]. The combination of antimicrobial and immune stimulating properties of CS makes it a potentially powerful antimicrobial agent to control plant diseases [118,119]. Nanotechnology is an auspicious area for the synthesis of materials for controlling phytopathogens. Cu-CS-NPs are considered a promising plant protection and growth promoting agent. Their exceptional capacity to withstand plant growth under pathogenic attack makes them a very effective antimicrobial agent [120]. CS and CS-based NPs have been tested for their inhibition of phytopathogenic bacteria [29,109,118,119,120]. Treatment of CS at 0.10 (mg/mL) concentration against the phytopathogenic bacteria Xanthomonas which causes bacterial leaf spots on Euphorbia pulcherrima, showed significant inhibition [4]. Bacterial wilt of tomato plants caused by Ralstonia solanacearum was reduced to 48% and 72% by seed treatment and soil drenching, respectively, at 10.0 (mg/mL) concentration [121]. Moreover, foliar application (0.20 mg/mL) of two different CS solutions such as solution-A (MW 1129 kDa, DD 85%) and solution-B (607 kDa with DD 75%) effectively controlled rice leaf blight and leaf streak caused by X. oryzae pv. oryzae and X. oryzae pv. Oryzicola, respectively [113]. Another study showed that foliar application of solution -A strongly reduced the disease severity of Acidovorax citrulli (fruit blotch of watermelon) [122].
Li et al. [109] synthesized, characterized, and studied the antibacterial activity of CS/TiO2 NPs against rice phytopathogen X. oryzae pv. oryzae. They found that CS/TiO2 NPs at the ratio of 1:5 had higher disease resistance. However, substantial inhibition of Xanthomonas species, which causes bacterial diseases of E. pulcherrima, was renowned regardless of the different MWs and DD of CS and type of bacteria [123]. Also, the bacterial speck disease (caused by P. syringae pv. tomato DC3000) severity in tomato was significantly decreased in CS pretreated seedlings, showing an auspicious action of CS as a nontoxic biopesticide [115]. Mohammadi et al. [89] confirmed that CS-NPs had potentially high antibacterial activity against soft rot bacterial pathogens P. fluorescens and Erwinia carotovora. The studies of Esyanti et al. [110] showed that CS-NPs were found to be effective in reducing disease severity by X. campestris in chili peppers and could possibly be used as a substitute to chemical bactericide. Oh et al. [111] investigated the antibacterial activities against phytopathogenic bacteria, such as three strains of E. carotovora subsp. carotovora and one strain of X. campestris pv. vesicatoria. The Ag-NP-entrapped CS showed significant antibacterial activity against tomato wilt pathogen R. solanacearum [112]. They confirmed that green synthesized CS-derived NPs containing Ag-NPs from leaf extracts were a promising and sustainable alternative in agriculture. These outcomes validate the potential of CS-NPs for biological control and crop yield without upsetting soil characteristics or the environment.
7. Conclusions and Future Perspectives
Natural biopolymers such as CS have vast aptitude for antibacterial applications. By using suitable NP synthesis procedures, it is prospective to develop firm and vastly active CS-NPs for various industrial applications. The biological usage of CS-NPs either alone or in combination with other compounds have inhibitory effects against both G− and G+ bacteria. This bioactivity is predisposed by numerous factors including chemical nature of CS, concentration, pH, and size of the particles. Studies to date have primarily absorbed features of in vitro studies, so it is essential to accomplish in situ study appraisals to define answers and alternates to the difficulties that both the medical and agricultural fields confront. CS-NPs mechanisms of action against bacteria are not yet completely inferred, thus mandating that study efforts endure. In addition, it is vital to continue to monitor and appraise toxicity arising from the application of CS-NPs in the control of bacteria and to provide guidance for rules and procedures related to their use and applications.
The following types of studies must be done to complete the knowledge gap in antibacterial activity of CS-NPs: (1) Investigations on mechanisms of antibacterial actions of CS-NPs; (2) Determining why CS-NPs are highly efficient against G− bacteria compared to G+ bacteria and by what mechanism this occurs; (3) Determining why medium MW-CS is more efficient against bacteria than high MW-CS; (4) Discovering the mechanism and toxicity behind the synthesis of metal ions associated CS-NPs and hybrid CS-NPs. These studies will support the development of a new group of antibacterial agents and is useful for both animal and plant studies.
Author Contributions
M.C. and S.C.C. conceived idea; M.C. wrote the manuscript; M.C.; S.C.C. and K.D.K. edited, revised and finalized the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Goy, R.C.; De Britto, D.; Assis, O.B.G. A review of the antimicrobial activity of chitosan. Polímeros 2009, 19, 241–247. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Rabea, E.I.; Badawy, M.E.T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, X.; Chen, R.; Huangfu, W.; Xie, G.-L. Antibacterial activity of chitosan solution against Xanthomonas pathogenic bacteria isolated from Euphorbia pulcherrima. Carbohydr. Polym. 2008, 72, 287–292. [Google Scholar] [CrossRef]
- Jia, Z.; Shen, D.; Xu, W. Synthesis and antibacterial activities of quaternary ammonium salt of chitosan. Carbohydr. Res. 2001, 333, 1–6. [Google Scholar] [CrossRef]
- Vinsová, J.; Vavříková, E. Chitosan derivatives with antimicrobial, antitumour and antioxidant Activities—A Review. Curr. Pharm. Des. 2011, 17, 3596–3607. [Google Scholar] [CrossRef]
- Wang, X.; Du, Y.; Fan, L.; Liu, H.; Hu, Y. Chitosan-metal complexes as antimicrobial agent: Synthesis, characterization and structure-activity study. Polym. Bull. 2005, 55, 105–113. [Google Scholar] [CrossRef]
- Yang, T.-C.; Chou, C.-C.; Li, C.-F. Antibacterial activity of N-alkylated disaccharide chitosan derivatives. Int. J. Food Microbiol. 2005, 97, 237–245. [Google Scholar] [CrossRef]
- Kammoun, M.; Haddar, M.; Kallel, T.K.; Dammak, M.; Sayari, A. Biological properties and biodegradation studies of chitosan biofilms plasticized with PEG and glycerol. Int. J. Boil. Macromol. 2013, 62, 433–438. [Google Scholar] [CrossRef]
- Ma, Z.; Garrido-Maestu, A.; Jeong, K.C. Application, mode of action, and in vivo activity of chitosan and its micro- and nanoparticles as antimicrobial agents: A review. Carbohydr. Polym. 2017, 176, 257–265. [Google Scholar] [CrossRef]
- Divya, K.; Vijayan, S.; George, T.K.; Jisha, M.S. Antimicrobial properties of chitosan nanoparticles: Mode of action and factors affecting activity. Fibers Polym. 2017, 18, 221–230. [Google Scholar] [CrossRef]
- Divya, K.; Jisha, M.S. Chitosan nanoparticles preparation and applications. Environ. Chem. Lett. 2017, 16, 101–112. [Google Scholar] [CrossRef]
- Kassem, A.; Ayoub, G.M.; Malaeb, L. Antibacterial activity of chitosan nano-composites and carbon nanotubes: A review. Sci. Total. Environ. 2019, 668, 566–576. [Google Scholar] [CrossRef]
- Rozman, N.A.S.; Tong, W.Y.; Leong, C.R.; Tan, W.N.; Hasanolbasori, M.A.; Abdullah, S.Z. Potential antimicrobial applications of chitosan nanoparticles (ChNP). J. Microbiol. Biotechnol. 2019, 29, 1009–1013. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Aljaeid, B.M. Preparation, characterization, and potential application of chitosan, chitosan derivatives, and chitosan metal nanoparticles in pharmaceutical drug delivery. Drug Des. Dev. Ther. 2016, 10, 483–507. [Google Scholar] [CrossRef] [Green Version]
- Vishwakarma, A.; Sriram, P.; Preetha, S.P.; Tirumurugaan, K.G.; Nagarajan, K.; Pandian, K. Synthesis and characterization of chitosan/TPP encapsulated curcumin nanoparticles and its antibacterial efficacy against colon bacteria. Int. J. Chem. Stud. 2019, 7, 602–606. [Google Scholar]
- Perinelli, D.R.; Fagioli, L.; Campana, R.; Lam, J.K.; Baffone, W.; Palmieri, G.F.; Bonacucina, G. Chitosan-based nanosystems and their exploited anti-microbial activity. Eur. J. Pharm. Sci. 2018, 117, 8–20. [Google Scholar] [CrossRef]
- Kravanja, G.; Primožič, M.; Knez, Ž; Leitgeb, M. Chitosan-Based (nano) materials for novel biomedical applications. Molecules 2019, 24, 1960. [Google Scholar] [CrossRef] [Green Version]
- Tokumitsu, H.; Ichikawa, H.; Fukumori, Y. Chitosan-gadopentetic acid complex nanoparticles for gadolinium neutron-capture therapy of cancer: Preparation by novel emulsion-droplet coalescence technique and characterization. Pharm. Res. 1999, 16, 1830–1835. [Google Scholar] [CrossRef]
- Supraja, N.; Thiruchenduran, S.; Prasad, T. Synthesis and characterization of chitosan nanoparticles and evaluation of antimicrobial activity antioxidant activity. Adv. Bioequiv. Bioavailab. 2018, 2, 88–93. [Google Scholar]
- MubarakAli, D.; LewisOscar, F.; Gopinath, V.; Alharbi, N.S.; Alharbi, S.A.; Thajuddin, N. An inhibitory action of chitosan nanoparticles against pathogenic bacteria and fungi and their potential applications as biocompatible antioxidants. Microb. Pathog. 2018, 114, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Xu, Z.; Jiang, X.; Hu, C.; Zou, X. Preparation and antibacterial activity of chitosan nanoparticles. Carbohydr. Res. 2004, 339, 2693–2700. [Google Scholar] [CrossRef] [PubMed]
- Maluin, F.N.; Hussein, M. Chitosan-based agronanochemicals as a sustainable alternative in crop protection. Molecules 2020, 25, 1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elieh-Ali-Komi, D.; Hamblin, M.R. Chitin and chitosan: Production and application of versatile biomedical nanomaterials. Int. J. Adv. Res. 2016, 4, 411–427. [Google Scholar]
- Hsu, S.; Chang, Y.-B.; Tsai, C.-L.; Fu, K.-Y.; Wang, S.-H.; Tseng, H.-J. Characterization and biocompatibility of chitosan nanocomposites. Colloids Surfaces B Biointerfaces 2011, 85, 198–206. [Google Scholar] [CrossRef]
- Mi, F.-L.; Tan, Y.-C.; Liang, H.-F.; Sung, H. In vivo biocompatibility and degradability of a novel injectable-chitosan-based implant. Biomaterials 2002, 23, 181–191. [Google Scholar] [CrossRef]
- Kim, H.; Tator, C.H.; Shoichet, M.S. Chitosan implants in the rat spinal cord: Biocompatibility and biodegradation. J. Biomed. Mater. Res. Part A 2011, 97, 395–404. [Google Scholar] [CrossRef]
- Shi, Z.; Neoh, K.G.; Kang, E.-T.; Wang, W. Antibacterial and mechanical properties of bone cement impregnated with chitosan nanoparticles. Biomaterials 2006, 27, 2440–2449. [Google Scholar] [CrossRef]
- Rabea, E.; Steurbaut, W. Chemically modified chitosans as antimicrobial agents against some plant pathogenic bacteria and fungi. Plant Prot. Sci. 2010, 46, 149–158. [Google Scholar] [CrossRef]
- Bernkop-Schnürch, A.; Dünnhaupt, S. Chitosan-based drug delivery systems. Eur. J. Pharm. Biopharm. 2012, 81, 463–469. [Google Scholar] [CrossRef]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.; Chen, X.; Yin, Y.; Zheng, H.; Xiong, X.; Du, Y. Synthesis, characterization and antibacterial activity of salicyloyl chitosan. Carbohydr. Polym. 2011, 83, 1274–1278. [Google Scholar] [CrossRef]
- Sajomsang, W.; Tantayanon, S.; Tangpasuthadol, V.; Daly, W.H. Synthesis of methylated chitosan containing aromatic moieties: Chemoselectivity and effect on molecular weight. Carbohydr. Polym. 2008, 72, 740–750. [Google Scholar] [CrossRef]
- Tan, H.-L.; Ma, R.; Lin, C.; Liu, Z.; Tang, T. Quaternized chitosan as an antimicrobial agent: Antimicrobial activity, mechanism of action and biomedical applications in orthopedics. Int. J. Mol. Sci. 2013, 14, 1854–1869. [Google Scholar] [CrossRef]
- Dutta, P.; Tripathi, S.; Mehrotra, G.; Dutta, J. Perspectives for chitosan based antimicrobial films in food applications. Food Chem. 2009, 114, 1173–1182. [Google Scholar] [CrossRef]
- Ray, S.D. Potential aspects of chitosan as pharmaceutical excipient. Acta Pol. Pharm. Drug Res. 2011, 68, 619–622. [Google Scholar]
- Upadhyaya, L.; Singh, J.; Agarwal, V.; Tewari, R.P. Biomedical applications of carboxymethyl chitosans. Carbohydr. Polym. 2013, 91, 452–466. [Google Scholar] [CrossRef]
- Cota-Arriola, O.; Cortez-Rocha, M.O.; Burgos-Hernández, A.; Ezquerra-Brauer, J.M.; Plascencia-Jatomea, M. Controlled release matrices and micro/nanoparticles of chitosan with antimicrobial potential: Development of new strategies for microbial control in agriculture. J. Sci. Food Agric. 2013, 93, 1525–1536. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in plant protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Liu, H.; Du, Y.M.; Wang, X.H.; Sun, L.P. Chitosan kills bacteria through cell membrane damage. Int. J. Food Microbiol. 2004, 95, 147–155. [Google Scholar] [CrossRef]
- Calvo, P.; Remuñan-López, C.; Vila-Jato, J.L.; Alonso, M.J. Chitosan and chitosan/ethylene oxide-propylene oxide block copolymer nanoparticles as novel carriers for proteins and vaccines. Pharm. Res. 1997, 14, 1431–1436. [Google Scholar] [CrossRef] [PubMed]
- Brunel, F.; Varon, L.; David, L.; Domard, A.; Delair, T. A novel syn-thesis of chitosan nanoparticles in reverse emulsion. Langmuir 2008, 24, 11370–11377. [Google Scholar] [CrossRef] [PubMed]
- Maitra, A.; Ghosh, P.K.; De, T.K.; Sahoo, S.K. Process for the Preparation of Highly Monodispersed Polymeric hydrophilic Nanoparticles. U.S. Patent 587411, 23 February 1999. [Google Scholar]
- Davis, L.I.S.S. Sustained release chitosan microspheres prepared by novel spray drying methods. J. Microencapsul. 1999, 16, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Jiang, H.; Ye, H.; Li, J.; Huang, J. Preparation, antibacterial, and antioxidant activities of silver/chitosan composites. J. Carbohydr. Chem. 2014, 33, 298–312. [Google Scholar] [CrossRef]
- Qian, J.; Pan, C.; Liang, C. Antimicrobial activity of Fe-loaded chitosan nanoparticles. Eng. Life Sci. 2017, 17, 629–634. [Google Scholar] [CrossRef] [Green Version]
- Badawy, M.E.I.; Lotfy, T.M.R.; Shawir, S.M.S. Preparation and antibacterial activity of chitosan-silver nanoparticles for application in preservation of minced meat. Bull. Natl. Res. Cent. 2019, 43, 83. [Google Scholar] [CrossRef]
- Katas, H.; Lim, C.S.; Azlan, A.Y.H.N.; Buang, F.; Busra, M.F.M. Antibacterial activity of biosynthesized gold nanoparticles using biomolecules from Lignosus rhinocerotis and chitosan. Saudi Pharm. J. 2018, 27, 283–292. [Google Scholar] [CrossRef]
- Tamara, F.R.; Lin, C.; Mi, F.-L.; Ho, Y.-C. Antibacterial effects of chitosan/cationic peptide nanoparticles. Nanomaterials 2018, 8, 88. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Wu, C.; Fu, S.; Wang, L.; Yuan, C.; Chen, S.; Hu, Y. Integration of lysozyme into chitosan nanoparticles for improving antibacterial activity. Carbohydr. Polym. 2017, 155, 192–200. [Google Scholar] [CrossRef]
- Huang, L.; Cheng, X.; Liu, C.; Xing, K.; Zhang, J.; Sun, G.; Li, X.; Chen, X. Preparation, characterization, and antibacterial activity of oleic acid-grafted chitosan oligosaccharide nanoparticles. Front. Biol. China 2009, 4, 321–327. [Google Scholar] [CrossRef]
- Sotelo-Boyás, M.; Correa-Pacheco, Z.; Bautista-Baños, S.; Corona-Rangel, M. Physicochemical characterization of chitosan nanoparticles and nanocapsules incorporated with lime essential oil and their antibacterial activity against food-borne pathogens. LWT 2017, 77, 15–20. [Google Scholar] [CrossRef]
- Ali, M.J.; Sharafaldin, A.M.; Majid, P.; Mahdi, F.R.; Kazem, A.; Hajar, R.; Zuhair, M.H.; Mahdi, K.M.; Reza, M. Curcumin-loaded chitosan tripolyphosphate nanoparticles as a safe, natural and effective antibiotic inhibits the infection of Staphylococcus aureus and Pseudomonas aeruginosa in vivo. Iran. J. Biotechnol. 2014, 12, 1–8. [Google Scholar]
- Avadi, M.; Sadeghi, A.; Tahzibi, A.; Bayati, K.; Pouladzadeh, M.; Zohuriaan-Mehr, M.; Rafiee-Tehrani, M. Diethylmethyl chitosan as an antimicrobial agent: Synthesis, characterization and antibacterial effects. Eur. Polym. J. 2004, 40, 1355–1361. [Google Scholar] [CrossRef]
- Du, W.-L.; Niu, S.-S.; Xu, Y.-L.; Xu, Z.-R.; Fan, C.-L. Antibacterial activity of chitosan tripolyphosphate nanoparticles loaded with various metal ions. Carbohydr. Polym. 2009, 75, 385–389. [Google Scholar] [CrossRef]
- Nguyen, V.B.; Nguyen, T.T.H.; Wang, S.-L.; Vo, T.P.K.; Nguyen, A.D. Preparation of chitosan nanoparticles by TPP ionic gelation combined with spray drying, and the antibacterial activity of chitosan nanoparticles and a chitosan nanoparticle–amoxicillin complex. Res. Chem. Intermed. 2016, 43, 3527–3537. [Google Scholar] [CrossRef]
- Ngan, L.T.K.; Wang, S.-L.; Hiep, Đ.M.; Luong, P.M.; Nguyen, V.B.; Đinh, T.M.; Dzung, N.A. Preparation of chitosan nanoparticles by spray drying, and their antibacterial activity. Res. Chem. Intermed. 2014, 40, 2165–2175. [Google Scholar] [CrossRef]
- Piras, A.M.; Maisetta, G.; Sandreschi, S.; Gazzarri, M.; Bartoli, C.; Grassi, L.; Esin, S.; Chiellini, F.; Batoni, G. Chitosan nanoparticles loaded with the antimicrobial peptide temporin B exert a long-term antibacterial activity in vitro against clinical isolates of Staphylococcus epidermidis. Front. Microbiol. 2015, 6, 372. [Google Scholar] [CrossRef] [Green Version]
- Tsai, G.-J.; Su, W.-H. Antibacterial activity of shrimp chitosan against Escherichia coli. J. Food Prot. 1999, 62, 239–243. [Google Scholar] [CrossRef]
- Sudarshan, N.; Hoover, D.; Knorr, D. Antibacterial action of chitosan. Food Biotechnol. 1992, 6, 257–272. [Google Scholar] [CrossRef]
- Shahidi, F.; Arachchi, J.K.V.; Jeon, Y.-J. Food applications of chitin and chitosans. Trends Food Sci. Technol. 1999, 10, 37–51. [Google Scholar] [CrossRef]
- Raafat, D.; Von Bargen, K.; Haas, A.; Sahl, H.-G. Insights into the mode of action of chitosan as an antibacterial compound. Appl. Environ. Microbiol. 2008, 74, 3764–3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cava, F.; Lam, H.; De Pedro, M.A.; Waldor, M.K. Emerging knowledge of regulatory roles of d-amino acids in bacteria. Cell. Mol. Life Sci. 2010, 68, 817–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivask, A.; Elbadawy, A.; Kaweeteerawat, C.; Boren, D.; Fischer, H.; Ji, Z.; Chang, C.H.; Liu, R.; Tolaymat, T.; Telesca, D.; et al. Toxicity mechanisms in Escherichia coli vary for silver nanoparticles and differ from ionic silver. ACS Nano 2013, 8, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Birsoy, K.; Wang, T.C.; Chen, W.W.; Freinkman, E.; Abu-Remaileh, M.; Sabatini, D.M. An essential role of the mitochondrial electron transport chain in cell proliferation is to enable aspartate synthesis. Cell 2015, 162, 540–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eltahlawy, K.; El-Bendary, M.A.; Elhendawy, A.; Hudson, S. The antimicrobial activity of cotton fabrics treated with different crosslinking agents and chitosan. Carbohydr. Polym. 2005, 60, 421–430. [Google Scholar] [CrossRef]
- Joshi, M.; Ali, S.W.; Purwar, R. Ecofriendly antimicrobial finishing of textile using bioactive agents based on natural products. Indian J. Fibre Text. Res. 2009, 30, 295–304. [Google Scholar]
- Wang, X.; Du, Y.; Liu, H. Preparation, characterization and antimicrobial activity of chitosan–Zn complex. Carbohydr. Polym. 2004, 56, 21–26. [Google Scholar] [CrossRef]
- Chung, Y.-C.; Su, Y.-P.; Chen, C.-C.; Jia, G.; Wang, H.-L.; Wu, J.C.G.; Lin, J.-G. Relationship between antibacterial activity of chitosan and surface characteristics of cell wall. Acta Pharmacol. Sin. 2004, 25, 932–936. [Google Scholar]
- No, H.K.; Park, N.Y.; Lee, S.H.; Meyers, S.P. Antibacterial activity of chitosans and chitosan oligomers with different molecular weights. Int. J. Food Microbiol. 2002, 74, 65–72. [Google Scholar] [CrossRef]
- Zhong, Z.; Xing, R.; Liu, S.; Wang, L.; Cai, S.; Li, P. Synthesis of acyl thiourea derivatives of chitosan and their antimicrobial activities in vitro. Carbohydr. Res. 2008, 343, 566–570. [Google Scholar] [CrossRef]
- Cao, X.L.; Cheng, C.; Ma, Y.L.; Zhao, C.S. Preparation of silver nanoparticles with antimicrobial activities and the researches of their biocompatibilities. J. Mater. Sci. Mater. Electron. 2010, 21, 2861–2868. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.V.; Tran, L.D.; Ba, C.T.; Vu, H.D.; Nguyen, T.N.; Pham, D.G.; Nguyen, P.X. Synthesis, characterization, antibacterial and antiproliferative activities of monodisperse chitosan-based silver nanoparticles. Colloids Surfaces A Physicochem. Eng. Asp. 2010, 360, 32–40. [Google Scholar] [CrossRef]
- Perelshtein, I.; Ruderman, E.; Perkas, N.; Tzanov, T.; Beddow, J.; Joyce, E.; Mason, T.; Blanes, M.; Mollá, K.; Patlolla, A.; et al. Chitosan and chitosan-ZnO-based complex nanoparticles: Formation, characterization, and antibacterial activity. J. Mater. Chem. B 2013, 1, 1968–1976. [Google Scholar] [CrossRef] [PubMed]
- Devlieghere, F.; Vermeulen, A.; Debevere, J. Chitosan: Antimicrobial activity, interactions with food components and applicability as a coating on fruit and vegetables. Food Microbiol. 2004, 21, 703–714. [Google Scholar] [CrossRef]
- Venkatesham, M.; Ayodhya, D.; Madhusudhan, A.; Babu, N.V.; Veerabhadram, G. A novel green one-step synthesis of silver nanoparticles using chitosan: Catalytic activity and antimicrobial studies. Appl. Nanosci. 2012, 4, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Rasaee, I.; Ghannadnia, M.; Honari, H. Antibacterial properties of biologically formed chitosan nanoparticles using aqueous leaf extract of Ocimum basilicum. Nanomed. J. 2016, 3, 147–154. [Google Scholar]
- Aleanizy, F.S.; Alqahtani, F.Y.; Shazly, G.; Alfaraj, R.; Alsarra, I.; Alshamsan, A.; Abdulhady, H.G. Measurement and evaluation of the effects of pH gradients on the antimicrobial and antivirulence activities of chitosan nanoparticles in Pseudomonas aeruginosa. Saudi Pharm. J. 2017, 26, 79–83. [Google Scholar] [CrossRef]
- Wei, D.; Sun, W.; Qian, W.; Ye, Y.; Ma, X. The synthesis of chitosan-based silver nanoparticles and their antibacterial activity. Carbohydr. Res. 2009, 344, 2375–2382. [Google Scholar] [CrossRef]
- Muthukrishnan, A.M.; Manik, A.; Sathiyabama, M. Green synthesis of copper-chitosan nanoparticles and study of its antibacterial activity. J. Nanomed. Nanotechnol. 2015, 6, 251. [Google Scholar] [CrossRef]
- Sarwar, A.; Katas, H.; Zin, N.M. Antibacterial effects of chitosan–tripolyphosphate nanoparticles: Impact of particle size molecular weight. J. Nanoparticle Res. 2014, 16, 2517. [Google Scholar] [CrossRef]
- Orellano, M.S.; Isaac, P.; Breser, M.L.; Bohl, L.P.; Conesa, A.; Falcone, R.D.; Porporatto, C. Chitosan nanoparticles enhance the antibacterial activity of the native polymer against bovine mastitis pathogens. Carbohydr. Polym. 2019, 213, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ikono, R.; Vibriani, A.; Wibowo, I.; Saputro, K.E.; Muliawan, W.; Bachtiar, B.M.; Mardliyati, E.; Bachtiar, E.W.; Rochman, N.T.; Kagami, H.; et al. Nanochitosan antimicrobial activity against Streptococcus mutans and Candida albicans dual-species biofilms. BMC Res. Notes 2019, 12, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.-C.; Li, C.-F.; Chou, C.-C. Cell age, suspending medium and metal ion influence the susceptibility of Escherichia coli O157:H7 to water-soluble maltose chitosan derivative. Int. J. Food Microbiol. 2007, 113, 258–262. [Google Scholar] [CrossRef]
- Xing, K.; Chen, X.; Li, Y.Y.; Liu, C.; Liu, C.G.; Cha, D.S.; Park, H.J. Antibacterial activity of oleoyl-chitosan nanoparticles: A novel antibacterial dispersion system. Carbohydr. Polym. 2008, 74, 114–120. [Google Scholar] [CrossRef]
- Wang, G.-H. Inhibition and inactivation of five species of foodborne pathogens by chitosan. J. Food Prot. 1992, 55, 916–919. [Google Scholar] [CrossRef]
- Gomes, L.P.; Andrade, C.T.; Del Aguila, E.M.; Alexander, C.; Paschoalin, V.M. Assessing the antimicrobial activity of chitosan nanoparticles by fluorescence-labeling. Int. J. Biotechnol. Bioeng. 2018, 12, 111–117. [Google Scholar]
- Anitha, A.; Rani, V.D.; Krishna, R.; Sreeja, V.; Selvamurugan, N.; Nair, S.; Tamura, H.; Jayakumar, R. Synthesis, characterization, cytotoxicity and antibacterial studies of chitosan, O-carboxymethyl and N,O-carboxymethyl chitosan nanoparticles. Carbohydr. Polym. 2009, 78, 672–677. [Google Scholar] [CrossRef]
- Mohammadi, A.; Hashemi, M.; Hosseini, S.M. Effect of chitosan molecular weight as micro and nanoparticles on antibacterial activity against some soft rot pathogenic bacteria. LWT 2016, 71, 347–355. [Google Scholar] [CrossRef]
- Liu, N.; Chen, X.; Park, H.J.; Liu, C.; Liu, C.; Meng, X.-H.; Yu, L.-J. Effect of MW and concentration of chitosan on antibacterial activity of Escherichia coli. Carbohydr. Polym. 2006, 64, 60–65. [Google Scholar] [CrossRef]
- Sanpui, P.; Murugadoss, A.; Prasad, P.; Ghosh, S.; Chattopadhyay, A. The antibacterial properties of a novel chitosan–Ag-nanoparticle composite. Int. J. Food Microbiol. 2008, 124, 142–146. [Google Scholar] [CrossRef]
- Alarfaj, A.A. Preparation, characterization and antibacterial effect of chitosan nanoparticles against food spoilage bacteria. J. Pure Appl. Microbiol. 2019, 13, 1273–1278. [Google Scholar] [CrossRef] [Green Version]
- Gan, Q.; Wang, T.; Cochrane, C.; McCarron, P.A. Modulation of surface charge, particle size and morphological properties of chitosan–TPP nanoparticles intended for gene delivery. Colloids Surfaces B Biointerfaces 2005, 44, 65–73. [Google Scholar] [CrossRef]
- De Paz, L.E.C.; Resin, A.; Howard, K.A.; Sutherland, D.S.; Wejse, P.L. Antimicrobial effect of chitosan nanoparticles on Streptococcus mutans biofilms. Appl. Environ. Microbiol. 2011, 77, 3892–3895. [Google Scholar] [CrossRef] [Green Version]
- Yildirim-Aksoy, M.; Beck, B.H. Antimicrobial activity of chitosan and a chitosan oligomer against bacterial pathogens of warmwater fish. J. Appl. Microbiol. 2017, 122, 1570–1578. [Google Scholar] [CrossRef]
- Garrido-Maestu, A.; Ma, Z.; Paik, S.-Y.-R.; Chen, N.; Ko, S.-H.; Tong, Z.; Jeong, K.C. Engineering of chitosan-derived nanoparticles to enhance antimicrobial activity against foodborne pathogen Escherichia coli O157:H7. Carbohydr. Polym. 2018, 197, 623–630. [Google Scholar] [CrossRef]
- Alqahtani, F.Y.; Aleanizy, F.S.; El Tahir, E.; AlQuadeib, B.; Alsarra, I.; Alanazi, J.S.; Abdelhady, H.G. Preparation, characterization, and antibacterial activity of diclofenac-loaded chitosan nanoparticles. Saudi Pharm. J. 2018, 27, 82–87. [Google Scholar] [CrossRef]
- Honary, S.; Ghajar, K.; Khazaeli, P.; Shalchian, P. Preparation, characterization and antibacterial properties of silver-chitosan nanocomposites using different molecular weight grades of chitosan. Trop. J. Pharm. Res. 2011, 10, 74. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.; Liu, C.; Meng, X.H.; Yu, L.J.; Liu, C.G. Antibacterial mechanism of chitosan microspheres in a solid dispersing system against Escherchia coli. Colloids Surfaces B Biointerfaces 2008, 65, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Regiel, A.; Irusta, S.; Kyzioł, A.; Arruebo, M.; Santamaria, J. Preparation and characterization of chitosan–silver nanocomposite films and their antibacterial activity against Staphylococcus aureus. Nanotechnology 2012, 24, 015101. [Google Scholar] [CrossRef]
- Covarrubias, C.; Trepiana, D.; Corral, C. Synthesis of hybrid copper-chitosan nanoparticles with antibacterial activity against cariogenic Streptococcus mutans. Dent. Mater. J. 2018, 37, 379–384. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Cristóbal, L.F.; Martinez-Castañon, G.A.; Martínez-Martínez, R.; Loyola-Rodríguez, J.P.; Patiño-Marín, N.; Reyes-Macías, J.; Ruiz, F. Antibacterial effect of silver nanoparticles against Streptococcus mutans. Mater. Lett. 2009, 63, 2603–2606. [Google Scholar] [CrossRef]
- Besinis, A.; De Peralta, T.; Handy, R.D. The antibacterial effects of silver, titanium dioxide and silica dioxide nanoparticles compared to the dental disinfectant chlorhexidine on Streptococcus mutans using a suite of bioassays. Nanotoxicology 2013, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aliasghari, A.; Khorasgani, M.R.; Vaezifar, S.; Rahimi, F.; Younesi, H.; Khoroushi, M. Evaluation of antibacterial efficiency of chitosan and chitosan nanoparticles on cariogenic streptococci: An in vitro study. Iran. J. Microbiol. 2016, 8, 93–100. [Google Scholar] [PubMed]
- Costa, E.; Silva, S.; Tavaria, F.; Pintado, M. Study of the effects of chitosan upon Streptococcus mutans adherence and biofilm formation. Anaerobe 2013, 20, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Hipalaswins, W.M.; Balakumaran, M.D.; Jagadeeswari, S. Synthesis, characterization and antibacterial activity of chitosan nanoparticles and its impact on seed germination. J. Acad. Ind.Res. 2016, 5, 65–71. [Google Scholar]
- Taponen, S.; Pyörälä, S. Coagulase-negative staphylococci as cause of bovine mastitis—Not so different from Staphylococcus aureus? Veter. Microbiol. 2009, 134, 29–36. [Google Scholar] [CrossRef]
- Cardozo, V.F.; Lancheros, C.A.; Narciso, A.M.; Valereto, E.C.; Kobayashi, R.K.; Seabra, A.B.; Nakazato, G. Evaluation of antibacterial activity of nitric oxide-releasing polymeric particles against Staphylococcus aureus and Escherichia coli from bovine mastitis. Int. J. Pharm. 2014, 473, 20–29. [Google Scholar] [CrossRef]
- Li, B.; Zhang, Y.; Yang, Y.; Qiu, W.-J.; Wang, X.; Liu, B.; Wang, Y.; Sun, G. Synthesis, characterization, and antibacterial activity of chitosan/TiO2 nanocomposite against Xanthomonas oryzae pv. oryzae. Carbohydr. Polym. 2016, 152, 825–831. [Google Scholar] [CrossRef]
- Esyanti, R.R.; Farah, N.; Bajra, B.D.; Nofitasari, D.; Martien, R.; Sunardi, S.; Safitri, R. Comparative study of nano-chitosan and synthetic bactericide application on chili pepper (Capsicum annuum L.) infected by Xanthomonas campestris. AGRIVITA J. Agric. Sci. 2020, 42, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.-W.; Chun, S.C.; Chandrasekaran, M. Preparation and in vitro characterization of chitosan nanoparticles and their broad-spectrum antifungal action compared to antibacterial activities against phytopathogens of tomato. Agronomy 2019, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Santiago, T.R.; Bonatto, C.C.; Rossato, M.; Lopes, C.A.P.; Lopes, C.A.; Mizubuti, E.S.G.; Silva, L.P. Green synthesis of silver nanoparticles using tomato leaf extract and their entrapment in chitosan nanoparticles to control bacterial wilt. J. Sci. Food Agric. 2019, 99, 4248–4259. [Google Scholar] [CrossRef] [PubMed]
- Pilon, L.; Spricigo, P.C.; Miranda, M.; De Moura, M.R.; Assis, O.B.G.; Mattoso, L.H.C.; Ferreira, M.D. Chitosan nanoparticle coatings reduce microbial growth on fresh-cut apples while not affecting quality attributes. Int. J. Food Sci. Technol. 2014, 50, 440–448. [Google Scholar] [CrossRef]
- Li, B.; Liu, B.; Shan, C.-L.; Ibrahim, M.; Lou, Y.; Wang, Y.; Xie, G.; Li, H.-Y.; Sun, G. Antibacterial activity of two chitosan solutions and their effect on rice bacterial leaf blight and leaf streak. Pest Manag. Sci. 2012, 69, 312–320. [Google Scholar] [CrossRef]
- Mansilla, A.Y.; Albertengo, L.; Rodriguez, M.S.; Debbaudt, A.; Zúñiga, A.; Casalongué, C.A. Evidence on antimicrobial properties and mode of action of a chitosan obtained from crustacean exoskeletons on Pseudomonas syringae pv. tomato DC3000. Appl. Microbiol. Biotechnol. 2013, 97, 6957–6966. [Google Scholar] [CrossRef]
- El Ghaouth, A. Effect of chitosan on cucumber plants: Suppression of Pythium aphanidermatum and induction of defense reactions. Phytopathology 1994, 84, 313. [Google Scholar] [CrossRef]
- Amborabé, B.-E.; Bonmort, J.; Fleurat-Lessard, P.; Roblin, G. Early events induced by chitosan on plant cells. J. Exp. Bot. 2008, 59, 2317–2324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badawy, M.E.I.; Rabea, E.I.; Taktak, N.E. Antimicrobial and inhibitory enzyme activity of N-(benzyl) and quaternary N-(benzyl) chitosan derivatives on plant pathogens. Carbohydr. Polym. 2014, 111, 670–682. [Google Scholar] [CrossRef]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2014, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, R.C.; Kumaraswamy, R.V.; Kumari, S.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Cu-chitosan nanoparticle boost defense responses and plant growth in maize (Zea mays L.). Sci. Rep. 2017, 7, 9754. [Google Scholar] [CrossRef]
- Algam, S.A.E.; Xie, G.; Li, B.; Yu, S.; Su, T.; Larsen, J. Effects of Paenibacillus strains and chitosan on plant growth promotion and control of Ralstonia wilt in tomato. J. Plant Pathol. 2010, 92, 593–600. [Google Scholar]
- Li, B.; Shi, Y.; Shan, C.; Zhou, Q.; Ibrahim, M.; Wang, Y.; Wu, G.; Li, H.; Xie, G.; Sun, G. Effect of chitosan solution on the inhibition of Acidovorax citrulli causing bacterial fruit blotch of watermelon. J. Sci. Food Agric. 2013, 93, 1010–1015. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, L.; Li, B.; Wu, G.; Tang, Q.; Ibrahim, M.; Li, H.; Xie, G.; Sun, G. Action of chitosan against Xanthomonas pathogenic bacteria isolated from Euphorbia pulcherrima. Molecules 2012, 17, 7028–7041. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of chitosan.

Figure 2.
Applications of chitosan nanoparticles.

Figure 3.
Antibacterial mechanisms of chitosan nanoparticles.

Figure 4.
Factors affecting antibacterial activity of chitosan nanoparticles.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Antibacterial activity of chitosan nanoparticle in relation to size of nanoparticles and zeta potential.
Table 1.
Antibacterial activity of chitosan nanoparticle in relation to size of nanoparticles and zeta potential.
| Nanoparticles | Size of Nanoparticles (nm) | Zeta Potential (mV) | Antibacterial Activity | References |
|---|---|---|---|---|
| CS-Fe2+NPs | 206.4 | +28.82 | Escherichia coli, Staphylococcus aureus, Candida albicans | [46] |
| CS-Fe3+-NPs | 195.2 | +28.26 | ||
| CS-AgNP1 | 76.86 | −0.2250 | Escherichia coli and Salmonella typhimurium | [47] |
| CS-AgNP2 | 63.03 | −0.1930 | ||
| CS-AgNP3 | 61.57 | −0.0695 | ||
| CS-NP1 | 78.4 | +33.77 | Escherichia coli and Bacillus cereus | [49] |
| CS-NP2 | 150.67 | +33.63 | ||
| CS-NP3 | 201 | +32 | ||
| Protamine-CS-NP1 | 114.33 | +32.33 | ||
| Protamine-CS-NP2 | 84.8 | +30.27 | ||
| Protamine-CS-NP3 | 79.4 | +27.67 | ||
| CS-NPs | 476.2 | +17.20 | Escherichia coli and Staphylococcus aureus | [50] |
| CS-NPs | 548.1 | +12.50 | ||
| CS-Lys-NPs | 488.8 | +21.10 | ||
| CS-Lys-NPs | 613.5 | +13.6 | ||
| CS-NPs | 217 | +37.6 | Escherichia coli and Bacillus subtilis | [51] |
| Carvacrol Modified CS-NPs | 26 | +18.8 | ||
| Eugenol Modified-CS-NPs | 235 | +28.4 | ||
| CS-NPs | 53.99 | +51.37 | Escherichia coli, Salmonella choleraesuis and Staphylococcus aureus | [55] |
| CS-Ag-NPs | 90.29 | +92.05 | ||
| CS-Cu2+-NPs | 121.9 | +88.69 | ||
| CS-Zn2+-NPs | 210.9 | +86.65 | ||
| CS-Mn2+-NPs | 102.3 | +75.74 | ||
| CS-Fe2+-NPs | 95.81 | +71.42 | ||
| CS-NPs | 166.7 | +54.7 | Streptococcus pneumoniae | [56] |
| CS-NPs | 189.7 | +47.7 | ||
| CS-NPs | 334.6 | +38.6 | ||
| CS-NPs | 1230 | +34.9 | ||
| CS-NPs | 95.4 | +45.7 | Staphylococcus aureus | [57] |
| CS-NPs | 271.5 | +35.1 | ||
| CS-NPs | 335.9 | +29.6 | ||
| CS-NPs | 124 | +21.9 | Staphylococcus epidermidis | [58] |
| Temporin-B Loaded-CS-NPs | 185 | +8.8 |
Table 2.
Minimum inhibitory concentration (MIC) (μg/mL) and minimum bactericidal concentration (MBC) (μg/mL) values of chitosan nanoparticles and their derivatives against gram-negative (G−) and gram-positive (G+) bacteria.
Table 2.
Minimum inhibitory concentration (MIC) (μg/mL) and minimum bactericidal concentration (MBC) (μg/mL) values of chitosan nanoparticles and their derivatives against gram-negative (G−) and gram-positive (G+) bacteria.
| Nanoparticles | Gram-Negative/Positive Bacteria | Bacterial Species | MIC | MBC | References |
|---|---|---|---|---|---|
| Chitosan nanoparticles (CS-NPs) | G− bacteria | Escherichia coli | 1/16 | 1 | [22] |
| Escherichia coli | 1/32 | 2 | |||
| Salmonella choleraesuis | 1/16 | 2 | |||
| Salmonella typhimurium | 1/8 | 4 | |||
| G+ bacteria | Staphylococcus aureus | 1/8 | 4 | ||
| CS-NPs | G− bacteria | Escherichia coli | 1/32 | 1 | [22] |
| Escherichia coli | 1/32 | 1 | |||
| Salmonella choleraesuis | 1/32 | 1 | |||
| Salmonella typhimurium | 1/16 | 2 | |||
| G+ bacteria | Staphylococcus aureus | 1/6 | 2 | ||
| Ag-CS-NPs | G− bacteria | Escherichia coli | 32 | 128 | [45] |
| Salmonella choleraesuis | 64 | 256 | |||
| G+ bacteria | Staphylococcus aureus | 64 | 256 | ||
| Bacillus subtilis | 32 | 128 | |||
| CS-NPs | G− bacteria | Escherichia coli | 625 | 2500 | [50] |
| G+ bacteria | Bacillus subtilis | 312 | 1250 | ||
| CS-NPs | G− bacteria | Escherichia coli | 117 | 187 | [55] |
| Salmonella choleraesuis | 117 | 187 | |||
| G+ bacteria | Staphylococcus aureus | 234 | 281 | ||
| CS-Ag+NPs | G− bacteria | Escherichia coli | 3 | 6 | [55] |
| Salmonella choleraesuis | 3 | 6 | |||
| G+ bacteria | Staphylococcus aureus | 6 | 12 | ||
| CS-Cu2+NPs | G− bacteria | Escherichia coli | 9 | 12 | [55] |
| Salmonella choleraesuis | 9 | 12 | |||
| G+ bacteria | Staphylococcus aureus | 21 | 24 | ||
| CS-Zn2+NPs | G− bacteria | Escherichia coli | 18 | 24 | [55] |
| Salmonella choleraesuis | 18 | 24 | |||
| G+ bacteria | Staphylococcus aureus | 36 | 48 | ||
| CS-Mn2+NPs | G− bacteria | Escherichia coli | 73 | 97 | [55] |
| Salmonella choleraesuis | 73 | 97 | |||
| G+ bacteria | Staphylococcus aureus | 85 | 97 | ||
| CS-Fe2+NPs | G− bacteria | Escherichia coli | 121 | 195 | [55] |
| Salmonella choleraesuis | 121 | 195 | |||
| G+ bacteria | Staphylococcus aureus | 146 | 195 | ||
| Low MW CS-NPs | G− bacteria | Escherichia coli | 12.5 | 25 | [81] |
| Acinetobacter schindleri | 12.5 | 25 | |||
| Pseudomonas aeruginosa | 3.13 | 6.25 | |||
| G+ bacteria | Staphylococcus aureus | 6.25 | 12.5 | ||
| Bacillus cereus | 3.13 | 6.25 | |||
| Bacillus subtilis | 1.56 | 3.13 | |||
| Low MW CS-NPs | G− bacteria | Escherichia coli | 12.5 | 50 | [81] |
| Acinetobacter schindleri | 12.5 | 50 | |||
| Pseudomonas aeruginosa | 3.13 | 12.5 | |||
| G+ bacteria | Staphylococcus aureus | 12.5 | 25 | ||
| Bacillus cereus | 6.25 | 6.25 | |||
| Bacillus subtilis | 3.13 | 6.25 | |||
| High MW CS-NPs | G− bacteria | Escherichia coli | 25 | 50 | [81] |
| Acinetobacter schindleri | 25 | 50 | |||
| Pseudomonas aeruginosa | 12.5 | 25 | |||
| G+ bacteria | Staphylococcus aureus | 12.5 | 25 | ||
| Bacillus cereus | 6.25 | 25 | |||
| Bacillus subtilis | 6.25 | 25 | |||
| High MW CS-NPs | G− bacteria | Escherichia coli | 25 | 50 | [81] |
| Acinetobacter schindleri | 25 | 50 | |||
| Pseudomonas aeruginosa | 12.5 | 25 | |||
| G+ bacteria | Staphylococcus aureus | 12.5 | 25 | ||
| Bacillus cereus | 12.5 | 0.05 | |||
| Bacillus subtilis | 6.25 | 25 |
Table 3.
Antibacterial activity of chitosan nanoparticles and their derivatives against animal and plant pathogens.
Table 3.
Antibacterial activity of chitosan nanoparticles and their derivatives against animal and plant pathogens.
| Nanoparticles | Pathogen | Disease/Infection | References |
|---|---|---|---|
| CS-NPs-essential oil | Staphylococcus aureus, Listeria monocytogenes, Shigella dysenteriae, and Escherichia coli | Food-borne pathogens | [52] |
| CS-NPs-amoxicillin | Streptococcus pneumoniae | Respiratory infection | [56] |
| CS-NPs | Staphylococcus epidermidi | Opportunistic infections | [58] |
| CS-NPs | Staphylococcus aureus, Staphylococcus xylosus, Staphylococcus chromogenes, Staphylococcus haemolyticus | Bovine mastitis | [82] |
| CS-NPs | Streptococcus mutans, Candida albicans | Dental caries | [83] |
| Cu-CS-NPs | Streptococcus mutans | Dental caries | [94] |
| CS-NPs | Streptococcus mutans, Streptococcus sobrinus, Streptococcus sanguis, and Streptococcus salivarius | Cariogenic bacteria | [101] |
| CS-NPs | Pseudomonas fluorescens, Proteus mirabilis, Staphylococcus aureus, Klebsiella pneumoniae, Escherichia coli and Enterobacter aerogenes | Clinical pathogens | [106] |
| CS-NPs | Staphylococcus aureus | Bovine mastitis | [107] |
| CS-NPs | Staphylococcus aureus, Escherichia coli | Bovine mastitis | [108] |
| CS/TiO2 NPs | Xanthomonas oryzae pv. oryzae | Bacterial spot of tomato | [109] |
| CS-NPs | Xanthomonas campestris | Bacterial spot of chili-pepper | [110] |
| CS-NPs | Xanthomonas campestris and Erwinia carotovora | Bacterial spot and soft rot in tomato | [111] |
| CS-NPs | Ralstonia solanacearum | Bacterial wilt of tomato | [112] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chandrasekaran, M.; Kim, K.D.; Chun, S.C. Antibacterial Activity of Chitosan Nanoparticles: A Review. Processes 2020, 8, 1173. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091173
AMA Style
Chandrasekaran M, Kim KD, Chun SC. Antibacterial Activity of Chitosan Nanoparticles: A Review. Processes. 2020; 8(9):1173. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091173
Chicago/Turabian StyleChandrasekaran, Murugesan, Ki Deok Kim, and Se Chul Chun. 2020. "Antibacterial Activity of Chitosan Nanoparticles: A Review" Processes 8, no. 9: 1173. https://0-doi-org.brum.beds.ac.uk/10.3390/pr8091173
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.