
Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments


1
Institut Català de Paleoecologia Humana i Evolució Social (IPHES-CERCA), Zona Educacional 4, Campus Sescelades URV (Edifici W3), 43007 Tarragona, Spain
2
UMR 7269 LAMPEA, CNRS, Aix Marseille Université, Ministry Culture, 13190 Aix-en-Provence, France
Quaternary 2021, 4(3), 23; https://0-doi-org.brum.beds.ac.uk/10.3390/quat4030023
Submission received: 26 May 2021
/
Revised: 12 July 2021
/
Accepted: 14 July 2021
/
Published: 26 July 2021
(This article belongs to the Special Issue Taphonomy and Palaeoecology of Quaternary Vertebrates: Advances in Fossil and Experimental Studies)
Abstract
:Equus is a very sensitive genus which has expanded over a large area and lived in Europe despite the climatic instability of the Pleistocene. Its persistence and abundance are helpful in understanding and describing environmental and climatic regional parameters. In this study, we present the result of dental mesowear and microwear analysis and post-cranial skeleton biometry on Equus populations located in two regions in the South of France from ten sites, corresponding to twelve assemblages dated from MIS 12 to MIS 5. The areas refer to two major climatic zones: the oceanic or subcontinental climate for the South West of France, and the Mediterranean for the South East. The first objective of this study is to integrate and compare biometric data, dental wear, and other already-published environmental proxies. The goal is to discuss the validity of horse body shape adaptations on a small geographical scale. The second objective is to describe the impact of environmental features on the horse population through time in the two regions. We observe that the Equus diet was quite diverse, according to microwear analysis which shows adaptations according to seasonal variations. However, they remained mostly grazers over a long period of time. Estimated body mass of Equus in the localities studied here varies from a mean of 468 up to a mean of 570 kg, but these variations failed to be correlated with the diet, the climatic period, or the geographical position of the horse population, probably because of the sample size or the restricted time-span or geographical scale. However, the conformation of the metapodials and the width of the third phalanges may have been linked with environmental and behavioural parameters. The width of the third phalange may be correlated with the recurrence of the snow cover, while the robustness of the metapodial co-occurs with a humid climate. Also, diet may influence the conformation of the bones, since the tall and slender horses seem to be preferentially grazers all year long and seasonally browser horses are tall and robust. Seasonally mixed-feeder horses, all coming from the Mediterranean area, were found to be smaller, perhaps in relation to a less productive environment. The correspondence of the dietary and morphometrical data could suggest high pressure on the horse population, which caused rapid body adaptation. Thus, the combination of these different proxies allows us to suggest more accurate large mammal paleoenvironmental reconstructions.
1. Introduction
This paper addresses Equus from the caballoid lineage populations from the end of the Middle Pleistocene to the beginning of the Late Pleistocene succession in the South of France. This area can be divided into two major climatic zones: the South East, subject to a Mediterranean climate, and the South West, with an oceanic to subcontinental climate, depending on distance from the sea [1]. In addition, the two zones are characterised by different hydrographic basins and reliefs. These parameters, along with latitude and longitude, affect temperature, precipitation [2], and the composition of the plant communities [3]. The South East is known for its relative climatic stability across time, allowing the persistence of temperate plant species during glacial periods [4]. The South West climate is much more sensitive to global variations [5,6,7]. The interglacial stages covered by this paper are isotopic stages 9, 7 and 5. The first two of these are slightly cooler than today, and the last one is warmer, with a mean of 5 Celsius degrees more than today [8]. Despite the fact that temperatures vary a lot during the phases, the vegetation cover generally remains similar to current cover [9]. The main vegetal cover in the South West is a continental acidophilous oak woodland. In the South East, the vegetation is mainly medio-Mediterranean vegetation (with pine, evergreen oak, rosemary, rock rose and lavender) [3]. The glacial stages relevant to this paper are isotopic stages 12, 10 and 6. Temperatures have been modelled for the Saale Glaciation (MIS 6) in particular and the means show differences between the two regions: the South East is warmer (approximately 15 °C in summer and −1 °C in winter) than the South West (apparently 10 °C in summer and −10 °C in winter) [10]. The vegetation assemblages have also been modelled, and assume many differences between the two regions. The South East vegetation may have been dominated by a temperate savannah and the South West vegetation may have been part of a taiga Forest and part of a mammoth steppe biotope [11]. The South of France is a contrasted region with distinct environments; thus, it may be a key region for understanding the impact of local environmental conditions on ecological, ethological and morphological adaptations of herbivorous megafauna. During these periods and in these regions, Pleistocene horses were able to occupy these various biotopes, which suggests behavioural flexibility.
Many studies have already shown that Equus is a flexible genus that can live in a wide range of environments and occasionally adopt a diversified diet (e.g., [12,13,14,15,16,17,18]). Some populations are characterised by a high amount of browsing in their diet, for example at Schöningen or Taubach [14,16]. Other horse assemblages, located in the Fairbanks area of Alaska, for example, suggest that the Equus diet is more variable than other ungulates from the same area [13]. While, as a general trend, the size of Equus decreased continuously through the Late Pleistocene [19,20,21], during the Middle Pleistocene horses present various morphometrical features, including size, that vary according to environmental conditions [12]. However, for now, the rate of evolutionary adaptation or the phenotypic plasticity of the skeleton is unknown, and so is the geographical and chronological accuracy of these observations.
In order to discuss this observation, in this paper we describe the morphological features of twelve populations of horses from the caballoid lineage from the South of France. We also reconstruct the dietary habits of those populations with the help of meso- and microwear dental analysis. This will allow us: (1) to determine if interpretations of horses’ biometrical adaptations are still valid for a restricted geographical scale; and (2) to document the change of those features through time in these regions.
2. Paleoenvironmental Parameters of the Study Sites
We have selected ten sites located in South West and South East France, corresponding to twelve fossil assemblages (Figure 1, two sites—Rameaux and Pech de l’Azé II—present different layers). The dates of the studied deposits range between MIS 12 (ca. 478-424 ky) and MIS 5 (ca. 130-71 ky) (dating from [22]) (Table 1), covering the period from the end of the Middle Pleistocene to the beginning of the Late Pleistocene. These sites correspond to different climatic phases, from glacial (Camp-de-Peyre, La Micoque, Rigabe, Suard, Pech de l’Azé II-l.9-6 and Coudoulous II) and interglacial contexts (Rameaux, Lunel-Viel, Payre, Pech de l’Azé II.l.4-2 and Peyre). They are rich in horse fossil material with well-preserved bones and teeth. In this paper, we have referred to the Latin terminology E. caballus rather than E. ferus following the demonstration of J.-L. Guadelli [23]. Moreover, we referred to several species and subspecies described for the caballoid lineage in each site through the latest works on this subject (see below).
Local environmental parameters of some of the sites have already been investigated through various proxies (sediment, pollen, microfauna, etc). However, not all of these proxies were used in the study of all the sites. Thus, the palaeoenvironmental data available for some sites are not as complete as for others.
The site of Camp-de-Peyre accumulated abundant elements such as Rangifer sp., Dicrostonyx torquatus and Microtus gregalis, which are clearly indicative of the cold climatic phase [42,43]. However, the presence of some taxa, such as Apodemus sylvaticus and Clethrionomys glareolus, nonetheless suggests the existence of sheltered places. According to these compositions of the mammal species and sedimentological studies [44], the landscape must have been a steppe with some wooded patches. The horse described in this site is Equus mosbachensis campdepeyri [25,42].
At La Micoque, we studied the levels E, J and H. The faunal association is largely dominated by Equus mosbachensis materials, which represent 75% of the remains [40,41]. The study of sediment suggests that the median ensemble (from which all of the studied bones and teeth come) is the result of river transport [45,46]. Occasionally across the year, the flow was very strong, which is typical of arid or semi-arid climate conditions [45]. Furthermore, some sediment lenses correspond to ice block ersatz. Overall, these observations strongly suggest a very cold and arid climate.
As at La Micoque, Equus mosbachensis is the most abundant taxon in the two parts of the Igue des Rameaux (Amont and Aval), with at least 25 individuals [47]. The other herbivores are: Palaeoloxodon antiquus, Coelodonta antiquitatis, Equus hydruntinus, Haploidoceros mediterraneus, Cervus elaphus, Hemitragus sp. and Bos primigenius [37]. Microfauna studies show differences between the paleoenvironment of the two parts of the Igue des Rameaux [48]. For the Aval ensemble, which is a bit more recent than that of the Amont [25,38], the climate must have been temperate-cool, with more precipitation but less snow cover than in the Amont ensemble, and the forest cover was quite significant (25% of the vegetal composition). For the Amont ensemble, the climate was semi-continental and the steppe more significant. In the two parts of the site the horse was described as E. mosbachensis [25].
At Lunel-Viel, the climate must have been temperate, according to fauna associations (Cervus elaphus, Capreolus cf. süssenbornensis, Sus sp., Equus hydruntinus, etc.); furthermore, some birds such as Hieraaetus fasciatus and Falco naumanni are related to the Mediterranean area [35,49,50,51]. The study of the horses’ skeleton suggests a very humid environment, described as a swamp, similar to the current Camargue [36]. The tooth wear study made it possible to rule out this last hypothesis and suggest a rich biotope instead, both in monocot and dicot plants, allowing strong resource partitioning between bovids and equids, which is not consistent with a swamp biotope [52]. The horse was described as Equus mosbachensis palustris [25,36].
At Payre, the herbivore assemblage is rich and diverse (Palaeoloxodon antiquus, Stephanorhinus kirchbergensis, S. hemitoechus, Bos primigenius, Bison priscus, Hemitragus sp., Rupicapra sp., Cervus elaphus, Megaloceros sp., Dama dama, Capreolus capreolus, and Sus scrofa), probably owing to its geographical position. Indeed, it is located in the Rhône Valley at the crossroads of various biotopes, mainly the plateau and the bottom of the valley. For level F (studied here), microfauna studies show that the climate must have been cool and dry, but not enough to indicate a glacial episode [53]. The isotopic analysis indicates that the horses lived in the bottom of the valley, rather than on the top of the plateau [54,55]. The horse is described as Equus mosbachensis in the layer F [25].
The faunal association of the site of Rigabe’s Rissian levels (X to J) is dominated by Equus sp., Cervus elaphus, Capreolus capreolus and Sus scrofa which suggests a forested area [56]. Moreover, the sedimentary analysis suggests a cold climate [33]. The horse was attributed to E. mosbachensis palustris [25].
At Suard, the faunal association is dominated by Equus caballus piveteaui [25,32], Rangifer tarandus and Coelodonta antiquitatis, and the presence of Mammuthus primigenius and Saiga tatarica is signaled. The pollen and faunal analyses of the Suard cave show that the climate must have been cold and dry, with a mosaic landscape dominated by steppes, with some wooded habitats [57,58,59].
With Equus mosbachensis in layers 9 to 6 and E. caballus in layer 4 [19,25], several herbivore species have been described in the different layers of Pech de l’Azé II: for example, Stephanorhinus kirchbergensis (layer 9 to 7) replaced by Coelodonta antiquitatis (in layer 6), Bison priscus (l. 9 to 5), Bos primigenius (l. 9 to 4), Cervus elaphus (l.9 to 4), Megaloceros sp. (l.9), and Capreolus sp. (l.4). The pollen of the sequence of the site of Pech de l’Azé II has been studied [60] but the sedimentary analysis shows a “ruissellement” (overland flow) phenomenon [46,61], which interferes with layer integrities [62]. Nonetheless, the presence of cryoclastic complexes in layers 9 to 6 suggests very cold conditions, whereas layer 4 corresponds to a temperate climate.
Cold-adapted taxa have been described in layer 9 of Coudoulous II, such as Mammuthus intermedius, Coelodonta antiquitatis and Rangifer tarandus. Moreover, the microfauna analysis indicates a cold period, relatively dry, with a mixed-landscape dominated by a steppe [27,28,63]. The horse was attributed to E. mosbachensis [25,28].
Some taxa present at Peyre are typical of cold environments such as Mammuthus primigenius and Coelodonta antiquitatis [64] but the faunal association is dominated by temperate taxa such as Cervus elaphus, Equus hydruntinus, Bos primigenius, Sus scrofa and Hystrix sp., which more likely to indicate a temperate climate [24,65,66,67,68]. Moreover, the study of the Microtidae, the Muridae and the gastropods suggests a dry climate [24]. The horse was described as E. caballus [25].
3. Biometric and Dental Wear Methods
3.1. Osteologic Measurement Protocols and Taphonomic Limitations
The morphometric characteristics observed on horse long bones (humerus, radius, femur and tibia) provide information on locomotor adaptations [69], and some features also depend on environmental parameters (e.g., [12,70,71,72,73]). As already shown, very few differences between adult horse bones are linked to age or sex [74,75]; only the fully fused long bone epiphyses were selected. Limb bone measurements were taken in accordance with the works of V. Eisenmann [76,77,78]. All of the long bones were measured with a calliper: humerus, radius, femur, tibia, metapodials and phalanges (I, II and III).
Although all of the selected sites are rich in bone remains, their conservation differs and constrains this study. Indeed, some of the sites considered here correspond to accumulations in caves of natural origin or linked to the activity of carnivores (Camp-de-Peyre, Les Rameaux, Lunel-Viel, Coudoulous II and Peyre). The bones are mostly very well preserved and numerous morphometric observations have been possible. On the other hand, the accumulation of other sites is due to human activities (La Micoque, Payre l.F, Suard, Pech de l’Azé II and Rigabe). In these sites, bone fragmentation is much more significant and has limited the observations. The most striking example is the La Micoque assemblage which, in addition to being of anthropogenic origin, is also a cliff foot site. A minimum of twenty individuals have been counted there [40] and no whole long bones are known. Thus, although the selected assemblages are rich, their origin and preservation conditions are factors that may have limited our observations.
3.2. Body Mass Estimation and Limb Proportions
Body mass was estimated following the recommendations of Saarinen et al. [18]. They established that the better equations to estimate body mass were the ones published by Scott [79] using the metapodials. In this paper, we use measurements of the minimum mid-shaft width, maximum proximal width, and maximum distal width of third metacarpals and third metatarsals in order to reconstruct body mass.
The relative proportions of the limb bones were analysed using log-ratio diagrams [80]. This graphic allows us to compare the differences in size and conformation between different populations based on measurements standardised on a referenced population. In this paper, the standard used for comparison is E. hemionus, published by V. Eisenmann on her website [76]. Along with the fossil sample, a sample for Equus przewalskii [81] (p. 26) is included for comparative purposes. The maximal length of the long bones (humerus, femur, radius, tibia, third metacarpal and metatarsal, first anterior and posterior phalanges) and the greatest width of the third phalanges of all the studied populations were compared. Comparing the proportions of the different limb segments allows the development of one segment to be compared to another (e.g., length of radius to metacarpals) and therefore provides information on locomotion [12]. Comparing the greatest width of the third phalanges allows us to compare ground hardness adaptation in several populations [72].
3.3. Mesowear and Microwear Methods
The horses’ diet in each site was studied using dental mesowear and microwear analysis. These two techniques provide information about the diet on two different timescales: mesowear indicates the average diet during the individual’s life or its last months or years [82,83], and microwear reveals the diet of the last few days to months of an animal’s life [84,85,86,87,88]. Combined together, these two methods allow us to understand the evolution of the animal’s diet in relation to seasonal variations over a period of several months. All the meso- and microwear data were collected by the author except for those from Payre, studied by F. Rivals [14,89].
3.3.1. Dental Mesowear
Mesowear analysis is a method based on the observation of wear patterns on ungulate molar cusps that indicate the diet of an individual animal [83,90]. The sharpness and the morphology of cusp apices are correlated with relative attritive and abrasive dental wear. Thus, a diet with low abrasion (and high attrition) shows very sharp and high relief molar buccal cusps and, on the contrary, a diet with high abrasion (due especially to the consumption of siliceous grass [91]) will result in rounded, blunted and low relief cusps.
We selected only upper intact adult teeth. As for the bones, taphonomic processes constrained data collection since the slightest alteration (e.g., breaks or polish) in cusp integrity led to its exclusion from selection. Observation of the cusp shape on horse teeth is easier on upper ones, which decreases the chance of potential mistakes. Unworn (and marginally worn) teeth as well as extremely worn ones are excluded from the following analysis [83,89,90].
For this study, the preference was for observation of the paracone of the second upper molar, as recommended by [83], but we also extended the analysis to the upper fourth premolar and first and third molar, as no differences were detected between the teeth [92], in order to increase the number of samples. Furthermore, only medium-age teeth were selected and juvenile or senescent teeth were excluded, as it was observed in [90] that they were less sensitive to diet.
In this study, we employed the standardised method proposed by the authors of [93] and modified by the authors of [94], already used in other works (e. g. [95,96]). This method categorises dental wear into seven groups (numbered from 0 to 6), according to their shape and topography (0 = high and sharp; 6 = blunt with no relief). The average value of mesowear data from a single sample corresponds to the “mesowear score” noted here “MWS” [93].
3.3.2. Dental Microwear
The microwear analysis study followed the protocol established by [97,98]. The occlusal surface of each tooth was cleaned using acetone and 96% alcohol. Then the surface was moulded with a high-resolution silicone and casts were made using clear epoxy resin. The transparent casts were then observed with a stereomicroscope at magnifications of ×35. Observations were restricted to a standard surface of 0.16 mm2 (using an ocular reticule), localised preferably on the upper tooth paracone and the lower tooth protoconid.
We selected adult molars and premolars, as it was shown that both provide consistent data for dietary studies in Artiodactyla and Perissodactyla, and because the intertooth variation in the microwear pattern is not significant [99].
Teeth with taphonomic alterations (e.g., wide and short striae or pits with irregular morphology) were excluded from this analysis [100].
The observed micro-traces, scratches, and pits in particular are left on the occlusal surfaces during mastication [101]. The variability of the density of these traces is indicative of various diets: grazer, mixed-feeder, and browser. We observed various features following the classification of [97,98] (Figure 2): pits (small and large), scratches (fine, coarse and hypercoarse) and gouges. The scratch width score (SWS) is also calculated with a score of ‘0’ for teeth with predominantly fine scratches per tooth surface, ‘1’ for those with mixed fine and coarse scratches on the tooth surface, and ‘2’ for those with predominantly coarse scratches. In addition, we used the method of [102], based on the observations of [97], which separates the three diet categories according to the number of scratches between 0 and 17 in the 0.16 mm2 area (0–17%). For extant ungulates, the diet categories match the percentages as follows: grazers (0.0–22.2% of scratches between 0 and 17), mixed feeders (20.9–70.0% of scratches between 0 and 17), and browsers (72.7–100% of scratches between 0 and 17).
3.4. Statistical Analysis
Several statistical analyses were performed in order to fully investigate ecological and morphometrical horse adaptations. We use principal component analysis (PCA) to observe the morphometrical variability of the horses metapodials. We used the Pearson correlation coefficient to construct the PCA. The variables used were tested using the Kaiser-Meyer-Olkin (KMO) index, which indicates the degree to which each variable in a set is predicted without error by the other variables. Every value below 0.5 was rejected. The PCA does not accept missing data, which led to the rejection of any incomplete sample in this analysis.
The Kruskal-Wallis test was used to test statistical differences between several groups with the Steel-Dwass-Critchlow-Fligner method of multiple comparisons for the post-hoc test. We used the Mann-Whitney test when we were comparing only two samples. In both cases, since the sample size is quite small, we used the exact p-value.
The Spearman correlation test was used to evaluate the liaison intensity between the diet categories defined by enamel wear studies and the size of the different horse populations. All the statistics were carried out using the Xlstat v. 2014.5.03 software with a risk threshold of 5%.
4. Results
4.1. Horse Size and Ecomorphological Variation through Time
In order to understand the impact of the environment on the morphology of horses from the South of France, we compared the ratio of limb bone proportions, and the body size of the different populations. First, we checked the homogeneity of the morphometric characteristics of the bones in the different levels of a same site. For the length of the limb bones and the greatest width of the phalanx III, only the Lunel-Viel site contained a sufficient number of specimens in each level to perform a Kruskal-Wallis test. The statistical results are non-significant for all bones and level of Lunel-Viel. For the La Micoque and Suard sites (which also contain several levels), we assumed that the bone dimensions used for this study are homogenous as this is the case for all the other bones and measurements (see [25]).
Based on the metapodials, and following the recommendations of [18], we estimated the body mass for each cohort (Figure 3). The average body mass within late Middle Pleistocene and early Late Pleistocene horse populations in southern France varied from 468 kg at Pech de l’Azé II l. 6-4 to 571 kg at Coudoulous II l.9. They therefore fall within the ‘large’ (450–550 kg) and ‘very large’ (>550 kg) weight classes defined by the authors of [18]. The variability of the weights of these populations, of the order of 100 kg, is therefore quite low.
The comparison of the proportion of horse limb length in the Pleistocene South of France populations is still preliminary due to the sample size (Table 2). The missing data and the low sample size prevent further statistical analysis. Nonetheless, the log ratio diagram (Figure 4) gives preliminary indications. It indicates that all of the samples have proportionally long limb bones compared to metapodials and phalanges, which is not the case with E. prezwalskii. The main difference between these cohorts of horses is in the width of the third phalanges. Those for which this data is available can be divided into two groups: (1) the populations with narrow phalanges composed of the horses of Lunel-Viel, La Micoque, Rameaux-Aval, and Pech de l’Azé l.8 (the two extremes of this group are the populations of Lunel-Viel with an average of 78.78 mm and Pech de l’Azé l.8 with a phalanx of 72.81 mm wide); and (2) the populations with wide phalanges composed of the horses of Peyre, Coudoulous II l.9, Rameaux-Amont, Suard and Camp-de-Peyre (the two extremes of this group are the populations of Peyre with an average of 90.26 mm and Camp-de-Peyre with phalanges III of 85.03 mm wide on average). The difference between these two groups is statistically significant (Mann-Whitney test: p-value = 0.0001). This measurement was not possible for the horse population of Rigabe and Pech de l’Azé II l. 6-4.
Principal component analyses were performed on the metapodials. Figure 5 shows the comparison of the conformation of the third metacarpal of the populations studied based on the measurement summarised in Table 3. It was constructed from the maximum length of the bone (M. 1), the width and thickness of the distal (M. 11 and 14) and proximal (M. 5 and 6) epiphyses, and the diaphysis of the bone (M. 3 and 4). The sagittal crest thickness (M. 12) and distal supra-articular width (M. 10) were excluded from the analysis because they were poorly represented on PCA axes 1 and 2. In this way the analysis presents 74.39% of the variability of the populations on these two axes. The third metacarpals of the Lunel-Viel, Rameaux-Aval and Peyre cohorts are located at the top of the graph, and are characterised in particular by the robustness of their diaphysis and the short overall length of the bone. The KMO index displays a value of 0.8354, which confirms the adequacy of the chosen variables. At the bottom left, those of Camp-de-Peyre are characterised by a greater length and a much slenderer appearance at the diaphysis and the distal and proximal ends. In Payre l.F, the metacarpal is long, slender at the diaphysis, but more developed at the extremities. Finally, the third metacarpals of Coudoulous II l.9 and the Rameaux-Amont, located at the bottom right of the graph, are both very long and very robust compared to the other populations.
Figure 6 presents a comparison of the same type of data for the third metatarsals (summarised in Table 4). For this PCA, the data used were: maximum bone length (M. 1), width and thickness of the diaphysis (M. 3 and 4) and distal end (M. 11 and 14), and width of the proximal end (M. 5). In order to keep only well-represented data on axes 1 and 2, we had to exclude the distal sagittal crest (M. 12) and proximal end (M. 6) thickness and the distal supra-articular width (M. 10). Thus, the PCA represents 70.86% of the variability of the third metatarsals in the study populations. The KMO index displays a value of 0.6650, indicating that the chosen variables are suitable for the analysis. As for the third metacarpals, the cohorts at the top of the graph corresponding to the Lunel-Viel, Peyre and Rameaux-Aval are short and robust. The third metatarsals at the bottom left of the graph from the Pech de l’Azé l.6-4, Camp-de-Peyre and Rigabe cohorts are longer and slender, particularly at the diaphysis. Finally, the cohorts of Coudoulous II l.9 and Rameaux-Amont are characterised by long and very robust bones.
4.2. Dietary Traits of Horses
The horses from Coudoulous II l.9, Suard and La Micoque display the highest mesowear values, in particular at Coudoulous II l.9, with a score of 5.36 (Table 5; Figure 7). The lowest values come from Rigabe, Payre l.F, Peyre, Pech de l’Azé II l.4-2, Rameaux-Aval and Rameaux-Amont, with a very low score at Payre l.F (MWS = 3.13). The populations from Lunel-Viel (MWS = 4.88) and Camp-de-Peyre (MWS = 4.18) are in the middle between the two extremes, Coudoulous II l.9 and Payre l.F. Nonetheless, the populations’ error bars (Figure 7) overlap and show that horses from the South of France remained preferential grazers according to mesowear analysis.
The preservation of teeth on a microscopic scale is quite similar from one site to another. At Camp-de-Peyre, 75% of the cast teeth were suitable for study, and this value was 81.8% at La Micoque l.E, 86.6% at La Micoque l.H, 57.8% at La Micoque l.J, 100% at Rameaux-Aval, 85.7% at Rameaux-Amont, 81.8% at Lunel-Viel, 100% at Payre l.F, 66.6% at Rigabe, 100% at Suard l.53, 89.6% at Suard l.52, 100% at Suard l. 51 and l.50, 65.6% at Pech de l’Azé II l.9-6, 100% at Coudoulous II l.9, 60% at Pech de l’Azé l.4, and 87.5% at Peyre. Thus, on the whole, the teeth are well preserved on a microscopic scale, with the exception of the Rigabe and Pech de l’Azé II sites, where about 40% had to be excluded from the analysis.
The microwear patterns of some of the horse populations show an abundant number of teeth with a lot of scratches, for example at: Camp-de-Peyre, La Micoque, Suard l.53-52-51, Pech de l’Azé II and Coudoulous II l.9, whereas this number at Lunel-Viel, Payre l.F, Rigabe and Suard l.50 is lower. It is lowest for the populations of les Rameaux and Peyre. (Figure 7 and Figure 8). This suggests that the first group corresponds to grazer feeding habits, the second to mixed-feeder ones, and the third to a browser diet. Some populations show a relatively high mean number of pits on occlusal tooth surfaces (Figure 8), such as Suard l. 51-52 or Camp-de-Peyre. This suggests the consumption of a larger amount of dusty grass than the other populations [102]. The scratch width score of the Rigabe horses is very low (=0.5), which could indicate the occasional consumption of leaves [103], in addition to a more abrasive diet. On the contrary, the scratch width score of Payre l. F is the highest (=1.1). Moreover, the high percentage of teeth with hypercoarse scratches (63.63%) could indicate the consumption of bark and twigs [102]. Thus, the microwear analysis results show high variations between the different populations.
5. Discussion
5.1. Body Mass and Morphometrical Adaptations to Local Environments
The body mass of late Middle Pleistocene and early Late Pleistocene horses in southern France varied from one population to another by a margin of about 100 kg. On average, horses from south-western France were slightly heavier than those from the South East, and their body mass was similar in the isotopic stages of cold and temperate climates. In the South East, horses in the temperate isotopic stages were on average heavier than those in the cold isotopic stages, but this observation is not well based as only one site represents a cold isotopic stage (Rigabe). Statistically, the variation in body mass of these cohorts cannot be explained by the geographical position of the sites (West/East) or by the general climatic ambience of the associated isotopic stages (Cold/Temperate) (Table 6). Moreover, our data, which are still preliminary, do not allow us to propose a chronological scheme for the evolution of horse size over this period of time.
The Pleistocene Equus paleopopulations from France analysed in this study all have elongated upper limb bones and proportionally short metapodials compared to E. przewalskii. This observation is similar to that made by the authors of [12] regarding Pleistocene horses from Britain and Germany. The elongation of distal limb bones has been linked to the occupation of arid and resource-poor habitats by Prezwalski’s horses and their significant mobility related to their living conditions [12,104]. During the Pleistocene, horses in southern France would have occupied environments richer in plant resources, allowing a less mobile lifestyle, similar to that of German or British populations with seasonal moves, which is also supported by microwear analysis [105]. The width of the third phalanx is much more variable in the cohorts studied. We have observed two groups: the first including La Micoque, Rameaux-Aval, Lunel-Viel and Pech de l’Azé II l. 8 with narrow third phalanges; and the second including Camp-de-Peyre, Rameaux-Amont, Suard, Coudoulous II l.9 and Peyre with wider third phalanges similar in proportion to E. prezwalskii. The width of the third phalanges is considered an adaptation to the hardness of the substrate. Narrow phalanges are thought to be an adaptation to hard ground [72]. In general, the phalanges of caballoid equids are wider than those of stenonian, asinian and zebra equids, and thus adapted to softer or more humid ground than the other ones, which are adapted to harder and more arid substrate [12,71,106,107]. The largest third phalanges are identified in a majority of cohorts from the glacial isotopic stages: Camp-de-Peyre (MIS 12), Suard and Coudoulous II l. 9 (MIS 6), which could indicate a regular snow cover. This is not systematic since two temperate isotopic stage cohorts also have very large third phalanges: Rameaux-Amont (MIS 9) and Peyre (MIS 5). At Rameaux-Amont, the study of the microfauna describes a cold interstage and an increase of the snow cover [48], which could be consistent with the increase of the width of the third phalange from Rameaux-Aval to Rameaux-Amont. At Peyre, not enough information about the paleoenvironment is available to discuss the width of the third phalange with regard to the significance of the snow cover or the softness of the substrate. On the other hand, narrower third phalanges can be found in both glacial (La Micoque, MIS 10 and Pech de l’Azé l. 9-6, MIS 6) and temperate isotopic stages (Rameaux-Aval, MIS 9 and Lunel-Viel, MIS 7). At La Micoque, the climate was suspected to be really dry [45], which could indicate a smaller period of snow cover compared to Camp-de-Peyre, for example, and be consistent with the morphology of the third phalange. At Pech de l’Azé II l.9-6, paleoenvironmental information is too limited to establish any correlation. At Lunel-Viel, based on the morphology of the horses, the paleoenvironment was first suspected to be very humid and similar to today’s Camargue wetland [36]. The comparison of the dietary habits of ungulates from the site and ones from Camargue allows us to discard this hypothesis and suggest a mosaic environment rich in monocot and dicot plants [52]. The morphology of the third phalanx of the horse from Lunel-Viel could indicate a harder substrate than horses confronted with significant snow cover or simply soft soils.
The morphology of the metapodials also shows a different pattern in the studied cohorts. The PCAs (Figure 5 and Figure 6) show that the limb bones of the horses from Lunel-Viel and Rameaux-Aval are short and robust, while the ones from Pech de l’Azé II l.6-4, Camp-de-Peyre and Rigabe are longer and slenderer, especially at the diaphysis. Finally, the metapodials of the horses from Coudoulous II l.9 and Rameaux-Amont are longer and more robust than the ones from the other populations. Limb length was correlated with various variables: those from a cold climate and/or a forested biotope could have short limbs [12,15,108], while long limb bones could indicate a rich environment with diversified dietary resources, as in mosaic steppe habitats [12], or strong seasonal variations [109]. The robustness of the bones can be interpreted as a wet climate indicator, according to the authors of [70,71], but [12] shows that, during glacial periods, North European horses tend to always be more robust than those from temperate periods. Moreover, temperate North European horses seem to be slenderer under oceanic conditions related to increasing humidity. Some temperate populations living in a mosaic steppe habitat can grow to a larger size, according to their limb bones length and robusticity, owing to a very suitable environment.
The variations in the slenderness of horse metapodials from the South of France are neither linked to the isotopic stage nor site location. Indeed, cold isotopic stage horses from Camp-de-Peyre (South West, MIS 12) and Rigabe (South East, MIS 6) are slenderer, while the horses from Coudoulous II l.9 (South West, MIS 6) are much more robust. During temperate stages, some populations are slender, as in Payre (South East, MIS 7), Pech de l’Azé II l.4, and Peyre (South West, MIS 5), and others have very robust limb bones, even superior to some glacial period populations, as at Rameaux-Amont and Rameaux-Aval (South West, MIS 9) and Lunel-Viel (South East, MIS 7). The slenderness of the horses from Camp-de-Peyre, Payre and Peyre is associated with a dry climate [24,34,43,110,111]. On the contrary, robust horses from Rameaux-Aval and Rameaux-Amont and Lunel-Viel are associated with a humid climate [36,48,52]. While the correlation between humidity and robusticity of the bones is repeated in our data in accordance with [70,71], it is not always the case. The metapodials of the horse from Coudoulous II l.9 are very robust and this is associated with humid conditions [27]. This feature may be related to very cold temperature adaptations associated with a diverse steppe vegetation [12].
5.2. Horse Dietary Traits at the End of Middle Pleistocene and the Beginning of Late Pleistocene in the South of France
Mesowear analysis shows that the horses from the South of France usually ate abrasive plants like grass during the year, which corresponds to the general trend of this genus [112], even if mixed-feeder populations have been investigated [113,114,115]. Microwear analysis, on the other hand, shows that their diet varies a lot over a shorter time scale. The flexibility of horses’ diet and their ability to incorporate grass and browse in their feeding or rely exclusively on browse has already been shown in several Pleistocene populations, such as Schöningen (MIS 9) [16] and Taubach (MIS 5) [89] in Germany. In a landscape where both browse and grass are available, they seem to prefer to select the latter [52,116], but in the context of the mammoth-steppe landscape, their diet seems to be less exclusive than that of extant horses [115].
In this study, often we found that horses from the South West of France from glacial periods, such as those from Camp-de-Peyre, La Micoque, Suard l.53-52-51, Pech de l’Azé II and Coudoulous II l.9, mainly ate grass all year long (with the notable exception of Suard l.50), whereas horses from the South East, from glacial and interglacial periods, were mixed-feeders shortly before their death. Finally, some horses from the South West from interglacial periods (Rameaux-Amont, Rameaux-Aval and Peyre) were browsers at the time of their death. In addition, in Suard l.51-52 and Camp-de-Peyre, located in the South West and characterised by a particularly cold and rigorous climate [42,43,58,59,117], the horses probably ate dusty or short grass. This behaviour seems to have been quite rare among horses from the South of France and corresponds to a specific local context. Likewise, south-eastern horses display dietary particularities that may be related to local environmental conditions. Indeed, at Rigabe, horses may have occasionally eaten leaves to supplement a more abrasive diet. At Payre l. F, microwear analysis shows the presence of bark and twigs in the horses’ diet, which is consistent with their habitat in the forested bottom of the valley below the site [55].
While Middle to Late Pleistocene horses from the South of France remained preferentially grazers, their diet was strongly impacted by seasonal changes in the environment. They displayed a variety of behaviour from grazers to browsers in accordance with local conditions. In the South West, the influence of the Atlantic Ocean induced colder conditions during the glacial isotopic stages (MIS 12, 10 and 6) [10] which may have favoured open areas and steppes and allowed horse cohorts to remain grazers all year long. In this general scheme, exceptions still exist, like at Suard l.50, where south-western glacial isotopic stage horses could incorporate more browse in their diet. Such behaviour may be the result of competition with other species, as at Canyars with Bos primigenius [96] or the indicator of the development of a forest during the accumulation of this layer. The interglacial periods in this region were more forested, perhaps similar to today’s vegetation with continental acidophilous oak woodland [3], which lead the horses to seasonally eat browse.
In the South East region, in each isotopic stage (MIS 7 and 6), the horses were mixed feeders at the time of their death. This behaviour may be related to the influence of the Mediterranean climate. This region is considered a refuge area where the impact of major global climate changes was buffered by the influence of the Mediterranean Sea [4]. The succession of the different isotopic stages was therefore less marked than in the South West, allowing the horses to maintain the same behaviour from one stage to the next. Their mixed-feeder diet at the time of their death illustrates the composition of the Mediterranean vegetation, rich in both grass and browse, with one rarely taking precedence over the other [3].
5.3. Relations between Horse Dietary Habits, Body Mass, and Morphometrical Parameters
The relationship between body mass, morphometrical skeleton features, and dietary habits can be difficult to establish, as they do not necessarily correspond to the same time span. Indeed, microwear analysis displays information about the final days to months of life [85,86,87,88], and mesowear indicates a general or monthly dietary signal [82,83]. The importance of phenotypic plasticity, and the rate of body adaptation, is largely unknown and may depend on the living condition of each cohort. However, several authors have found fairly large variations in body mass and bone conformation over relatively short time scales within the isotopic stage chronology [12,71,118].
In this study, the variability of the body mass of Middle to Late Pleistocene horses from the South of France was found to be quite low. Probably because of the size of the sample, no trend related to the isotopic stage or the region is apparent. Over larger time-spans and geographical areas, observations have already been made of underlying relationships between horses’ body mass and environmental factors, diet and behaviour. The growth of a large body mass has been suggested to be associated with seasonally highly-productive and high-quality plant communities [119,120,121], or with environments that have a seasonal shortage of dietary resources [109,121,122]. Also, a large body mass can be related to small group size and population densities [18,118,123]. Here, the Spearman correlation test between the mean weight and the MWS of each cohort shows a very light and non-significant positive relationship between those two data (n = 12; p-value = 0.4368; rho = 0.2448) in contrast to the studies described in [18] and [91]. This difference can be explained by the different chronological and geographical scales studied. This work focuses on a shorter period and a smaller area. The variation in body mass of horses according to their diet in the dental mesowear would therefore not be noticeable here. The impact of seasonal variations on horse body mass was also investigated by statistically comparing the body mass of the three dietary categories (G-G: grazer all year long; G-MF: generally, grazer, mixed-feeder at time of death; G-B: generally, grazer, browser at time of death). Each diet-body mass related group was not statistically different one from the other (Table 7).
On the other hand, morphometric analysis concerning metapodials and third phalanges underline a variability between the studied cohorts. Figure 9 displays the centroid of the diet categories according to meso- and microwear data (cf. supra) in relation to metacarpal and metatarsal morphometrics. In the two tests, the G-MF diet was shown to be related to the shortest bones, while the G-G diet was related to the longest. The G-B diet is more represented by the long and robust bones. Open areas rich with grass allow horses to grow tall, since there are no closed-area restrictions. Also, tall and slender metapodials in relation to proximal limb bones indicate cursoriality and an increased ability to escape predators by efficient running in open environments. In more closed-areas, small size may facilitate movement and predator avoidance [108]. However, G-B populations are also tall, but with a more robust conformation. This type of diet indicates strong seasonal variations in the plant composition in a humid climate, allowing the development of wooded areas. The humidity of the climate is one of the strongest factors explaining the growth in robusticity of the horses (cf. supra). The large size of the individuals may be explained by the small group size induced, at least seasonally, by the expansion of the forest cover, as demonstrated by the authors of [18]. The smaller horses in terms of height were shown, in this set of data, to be G-MF and were all located in the South East of France. This effect may be a consequence of the low productivity and quality of Mediterranean plants [124], which may have restrained the horses’ size. On the other hand, the seasonal variability under a Mediterranean climate is less pronounced than under an Atlantic one, which may not have led to a selection of tall horses [109].
In a small timespan and geographical range, the body mass of the horses does not vary enough to describe patterns and adaptations to environmental features. However, the conformation of the bones, here the metapodials, seems to be partly driven by the diet and environmental conditions, which themselves partly conditioned the diet. While body mass may be a good indicator of a larger time span and geographical range, bone conformation seems to be a more sensitive proxy. The relationship between it and the dietary analysis could indicate rapid skeleton adaptations for horses during the end of the Middle Pleistocene and the beginning of the Late Pleistocene in the South of France. While it is not possible to establish the rate of anatomical changes, some must have been very rapid, as illustrated at Les Rameaux. Within the same isotopic stage (MIS 9), the height of the horses and the width of their third phalanges increased in order to face more humid conditions, while their diet remained similar.
6. Conclusions
This study confirms the significant flexibility of horses and their capacity to live in a wide range of habitats. Very common in archaeological and paleontological sites from the end of the Middle and the beginning of the Late Pleistocene in the South of France, they adapted to various environmental and climatic conditions. While at these scales the variation of their body mass did not seem to be a good indicator of their adaptations, metapodials and third phalanges show rapid changes in their conformation in relation to the environment. The width of the third phalanges have been, in a majority of cases, linked with the significance of the snow cover, while the length and robusticity of the metapodials have been linked with the humidity of the climate, the size of the forest cover, the productivity of the environment, and the social and dietary behaviour of the horses.
While the mesowear analysis indicates that the horses remained preferentially grazers, their diet could be impacted by strong seasonal variation. Thus, several populations remained grazers (Camp-de-Peyre, La Micoque, Suard l.53-51, Pech de l’Azé l. 9-4 and Coudoulous II l.9), others could, at their time of death, have been mixed-feeders (Payre l. F, Lunel-Viel, Suard l.50 and Rigabe) or browsers (Rameaux-Aval, Rameaux-Amont and Peyre). Their diet, strongly related to the vegetation, may also have influenced the conformation of their metapodial. Thus, tall horses were grazers all year long and must have inhabited open landscape like the steppe. Tall and robust horses were more likely to be seasonal browsers, with their morphometrical and dietary features explained by the more sizeable forest cover which induces smaller groups. Finally, smaller horses were found in the South East of France, under the Mediterranean climate; their size may be explained by low-quality food resources.
All of the cohorts from this corpus also have similarities. In particular, the ratio of the limb proportions shows elongated upper limb bones compared to the autopodial, which indicates general low-mobility behaviour.
In the case of horses, morphometrical and dental wear studies seem to be good proxies in order to investigate their past behaviour and adaptations in relation to their environment. While the rate of anatomical adaptation cannot be truly established, the concordance between the results of the two proxies could indicate rapid anatomical changes and validate their integration at a paleontological study scale. Moreover, their combination could be a good tool to reconstruct their palaeoenvironment and their eco-ethology.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this paper.
Acknowledgments
I am particularly grateful to Jean-Philip Brugal who directed my PhD, from where this study is extracted. I would also like to thank LAMPEA, ED 355, and Aix-Marseille University for the help and funding they provided during this work. I also would like thank F. Rivals for sharing the teeth wear data from Payre. Finally, I would like to thanks the five reviewers and the academic editors, Juan Rofes, Janine Ochoa and Emmanuelle Stoetzel who help to greatly improve this manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- Ozenda, P. La Végétation Du Continent Européen; Delachaux et Niestlé: Lausanne, Switzerland, 1994; p. 271. [Google Scholar]
- Joannin, S. Changements Climatiques en Méditerranée à la Transition Pléistocène Inférieur-Moyen: Pollens, Isotopes Stables et Cyclostratigraphie. Ph.D. Thesis, Claude Bernard—Lyon I University, Lyon, France, 2007. [Google Scholar]
- Ozenda, P.; Borel, J.-L. An Ecological Map of Europe: Why and How? Compt. Rendus Acad. Sci. Ser. III Sci. Vie 2000, 323, 983–994. [Google Scholar] [CrossRef]
- Petit, R.J.; Aguinagalde, I.; de Beaulieu, J.-L.; Bittkau, C.; Brewer, S.; Cheddadi, R.; Ennos, R.; Fineschi, S.; Grivet, D.; Lascoux, M.; et al. Glacial Refugia: Hotspots But Not Melting Pots of Genetic Diversity. Science 2003, 300, 1563–1565. [Google Scholar] [CrossRef] [Green Version]
- Couchoud, I. Etude Pétrographique et Isotopique Des Spéléothèmes Du Sud-Ouest de La France Formés En Contexte Archéologique: Contribution à La Connaissance Des Paléoclimats Régionaux Du Stade Isotopique 5. Ph.D. Thesis, Bordeaux I University, Bordeaux, France, 2006. [Google Scholar]
- Wainer, K.; Genty, D.; Blamart, D.; Hoffmann, D.; Couchoud, I. A New Stage 3 Millennial Climatic Variability Record from a SW France Speleothem. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 271, 130–139. [Google Scholar] [CrossRef]
- Couchoud, I.; Genty, D.; Hoffmann, D.; Drysdale, R.; Blamart, D. Millennial-Scale Climate Variability during the Last Interglacial Recorded in a Speleothem from South-Western France. Quat. Sci. Rev. 2009, 28, 3263–3274. [Google Scholar] [CrossRef]
- Masson-Delmotte, V. Les grandes oscillations du climat depuis 800,000 ans. In Des Climats et des Hommes; Berger, J.-F., Ed.; La Découverte: Paris, France, 2012; pp. 57–72. [Google Scholar]
- Magri, D.; Palombo, M.R. Early to Middle Pleistocene Dynamics of Plant and Mammal Communities in South West Europe. Quat. Int. 2013, 288, 63–72. [Google Scholar] [CrossRef]
- Colleoni, F.; Krinner, G.; Jakobsson, M.; Peyaud, V.; Ritz, C. Influence of Regional Parameters on the Surface Mass Balance of the Eurasian Ice Sheet during the Peak Saalian (140 Kya). Glob. Planet. Chang. 2009, 68, 132–148. [Google Scholar] [CrossRef]
- Colleoni, F.; Krinner, G.; Jakobsson, M. Sensitivity of the Late Saalian (140 Kyrs BP) and LGM (21 Kyrs BP) Eurasian Ice Sheet Surface Mass Balance to Vegetation Feedbacks. Geophys. Res. Lett. 2009, 36, 1–5. [Google Scholar] [CrossRef]
- van Asperen, E.N. Ecomorphological Adaptations to Climate and Substrate in Late Middle Pleistocene Caballoid Horses. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 297, 584–596. [Google Scholar] [CrossRef]
- Rivals, F.; Mihlbachler, M.C.; Solounias, N.; Mol, D.; Semprebon, G.M.; de Vos, J.; Kalthoff, D.C. Palaeoecology of the Mammoth Steppe Fauna from the Late Pleistocene of the North Sea and Alaska: Separating Species Preferences from Geographic Influence in Paleoecological Dental Wear Analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 286, 42–54. [Google Scholar] [CrossRef]
- Moncel, M.-H.; Rivals, F. On the Question of Short-Term Neanderthal Site Occupations: Payre, France (MIS 8-7), and Taubach/Weimar, Germany (MIS 5). J. Anthropol. Res. 2011, 67, 47–75. [Google Scholar] [CrossRef]
- Van Asperen, E.N. Late Middle Pleistocene Horse Fossils from Northwestern Europe as Biostratigraphic Indicators. J. Archaeol. Sci. 2012, 39, 1974–1983. [Google Scholar] [CrossRef]
- Rivals, F.; Julien, M.-A.; Kuitems, M.; Van Kolfschoten, T.; Serangeli, J.; Drucker, D.G.; Bocherens, H.; Conard, N.J. Investigation of Equid Paleodiet from Schöningen 13 II-4 through Dental Wear and Isotopic Analyses: Archaeological Implications. J. Hum. Evol. 2015, 89, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Rivals, F.; Lister, A.M. Dietary Flexibility and Niche Partitioning of Large Herbivores through the Pleistocene of Britain. Quat. Sci. Rev. 2016, 146, 116–133. [Google Scholar] [CrossRef]
- Saarinen, J.; Cirilli, O.; Strani, F.; Meshida, K.; Bernor, R.L. Testing Equid Body Mass Estimate Equations on Modern Zebras—With Implications to Understanding the Relationship of Body Size, Diet, and Habitats of Equus in the Pleistocene of Europe. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Prat, F. Recherches sur les Equidés Pléistocènes en France. Ph.D. Thesis, Bordeaux Faculty of Sciences, Bordeaux, France, 1968. [Google Scholar]
- Forsten, A. Size Decrease in Late Pleistocene-Holocene Caballoid Horses (Genus Equus), Intra-or Interspecific Evolution? A Discussion of Alternatives. Quat. Int. 1993, 19, 71–75. [Google Scholar] [CrossRef]
- Magniez, P.; Boulbes, N. Environment during the Middle to Late Palaeolithic Transition in Southern France: The Archaeological Sequence of Tournal Cave (Bize-Minervois, France). Quat. Int. 2014, 337, 43–63. [Google Scholar] [CrossRef]
- Lisiecki, L.E.; Raymo, M.E. A Pliocene-Pleistocene Stack of 57 Globally Distributed Benthic Δ18O Records. Paleoceanography 2005, 20, PA1003. [Google Scholar] [CrossRef] [Green Version]
- Brugal, J.-P.; Argant, A.; Boudadi-Maligne, M.; Crégut-Bonnoure, E.; Croitor, R.; Fernandez, P.; Fourvel, J.-B.; Fosse, P.; Guadelli, J.-L.; Labe, B.; et al. Pleistocene Herbivores and Carnivores from France: An Updated Overview of the Literature, Sites and Taxonomy. Ann. Paléontol. 2020, 106, 102384. [Google Scholar] [CrossRef]
- Vernet, J.-L.; Mercier, N.; Bazile, F.; Brugal, J.-P. Travertins et terrasses de la moyenne vallée du Tarn à Millau (Sud du Massif Central, Aveyron, France). Quat. Rev. Assoc. Fr. Pour Létude Quat. 2008, 19, 3–10. [Google Scholar] [CrossRef]
- Uzunidis-Boutillier, A. Grands Herbivores de La Fin Du Pléistocène Moyen Au Début Du Pléistocène Supérieur Dans Le Sud de La France: Implications Anthropologiques Pour La Lignée Néandertalienne. Ph.D. Thesis, Aix-Marseille University, Aix-en-Provence, France, 2017. [Google Scholar]
- Jacobs, Z.; Jankowski, N.R.; Dibble, H.L.; Goldberg, P.; McPherron, S.J.P.; Sandgathe, D.; Soressi, M. The Age of Three Middle Palaeolithic Sites: Single-Grain Optically Stimulated Luminescence Chronologies for Pech de l’Azé I, II and IV in France. J. Hum. Evol. 2016, 95, 80–103. [Google Scholar] [CrossRef] [Green Version]
- Brugal, J.-P.; Aouadi, N.; Mai, B.T.; Cochard, D.; Costamagno, S.; Ellwood, B.; Girard, M.; Jaubert, J.; Jeannet, M.; Kervazo, B.; et al. La Grotte de Coudoulous II (Tour-de-Faure, Lot); Unpublished Excavation Report, Bilan 1997—2000; Pgm P1 Min. de la Culture, SRA Midi-Pyrénées, 2000; 138p. [Google Scholar]
- Uzunidis, A.; Brugal, J.-P. Les grands herbivores (Bovinés, Équidés, Rhinocérotidés, Proboscidiens) de la fin du Pléistocène Moyen: La couche 9 de Coudoulous II (Lot, Quercy, Sud-Ouest France). PALEO 2018, 29, 223–249. [Google Scholar] [CrossRef]
- Schvoerer, M.; Rouanet, J.-F.; Navailles, H.; Debénath, A. Datation Absolue Par Thermoluminescence Des Restes Humains Antéwürmiens de l’abri Suard à La Chaise-de-Vouthon (Charente). Comptes Rendus Acad. Sci. Paris 1977, 284, 1979–1982. [Google Scholar]
- Schwarcz, H.P.; Debénath, A. Datations Absolues Des Restes Humains de La Chaise-de-Vouthon (Charente) Au Moyen Des Déséquilibres Des Séries. Comptes Rendus Acad. Sci. Paris 1979, 288, 1155–1157. [Google Scholar]
- Blackwell, B.; Schwarcz, H.P.; Debénath, A. Absolute Dating of Hominids and Palaeolithic Artifacts of the Cave of La Chaise-de-Vouthon (Charente), France. J. Archaeol. Sci. 1983, 10, 493–513. [Google Scholar] [CrossRef]
- David, P.; Prat, F. Considérations Sur Les Faunes de La Chaise (Commune de Vouthon, Charente), Abris Suard et Bourgeois-Delaunay. Bull. Assoc. Fr. Pour Létude Quat. 1965, 2, 222–232. [Google Scholar] [CrossRef]
- Bonifay, E. Etude Géologique de La Grotte de Rigabe, Artigues (Var). In Proceedings of the Congrès Préhistorique de Monaco, XVIe Session, Monaco, 28 August–5 September 1959; Imprimerie Monnoyer: Le Mans, France, 1959; pp. 265–291. [Google Scholar]
- Moncel, M.-H. Les différentes phases d’occupations humaines à Payre et hypothèses sur les types d’occupation. In Le Site de Payre: Occupations Humaines dans la Vallée du Rhône à la fin du Pléistocène Moyen et au début du Pléistocène Supérieur; Patou-Mathis, M., Moncel, M.-H., Eds.; Société Préhistorique Française: Paris, France, 2008; pp. 309–314. [Google Scholar]
- Brugal, J.-P.; Giuliani, C.; Fosse, P.; Fourvel, J.-B.; Magniez, P.; Pelletier, M.; Uzunidis, A. Preliminary Data on the Middle Pleistocene Site of Lunel Viel I (Hérault, France). Alp. Mediterr. Quat. 2021, 34, 1–13. [Google Scholar] [CrossRef]
- Bonifay, M.-F. Le Cheval du pléistocène moyen des grottes de Lunel-Viel (Hérault). Equus mosbachensis palustris n. ssp. Gall. Préhistoire 1980, 23, 233–281. [Google Scholar] [CrossRef]
- Rouzaud, F.; Soulier, M.; Brugal, J.-P.; Jaubert, J. L’igue Des Rameaux (Saint-Antonin-Noble-Val, Tarn-et-Garonne). Un Nouveau Gisement Du Pléistocène Moyen. Premiers Résultats. PALEO 1990, 2, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Argant, A.; Brugal, J.-P. The Cave Lion Panthera (Leo) Spelaea and Its Evolution: Panthera Spelaea Intermedia Nov. Subspecies. Acta Zool. Crac. 2017, 60, 58–103. [Google Scholar] [CrossRef]
- Falguères, C.; Bahain, J.-J.; Saleki, H. U-Series and ESR Dating of Teeth from Acheulian and Mousterian Levels at La Micoque (Dordogne, France). J. Archaeol. Sci. 1997, 24, 537–545. [Google Scholar] [CrossRef]
- Langlois, A. Au Sujet du Cheval de La Micoque (Dordogne) et des Comportements Humains de Subsistance au Pléistocène Moyen Dans le Nord-est de l’Aquitaine. Ph.D. Thesis, Bordeaux I University, Bordeaux, France, 2004. [Google Scholar]
- Langlois, A. Le Cheval du gisement Pléistocène moyen de La Micoque (Les Eyzies-de-Tayac, Dordogne): Equus mosbachensis micoquii nov. ssp. PALEO 2005, 17, 73–110. [Google Scholar] [CrossRef]
- Guadelli, J.-L.; Prat, F. Le Cheval Du Gisement Pléistocène Moyen de Camp-de-Peyre (Sauveterre-La-Lémance, Lot-et-Garonne): Equus mosbachensis campdepeyri n. ssp. PALEO 1995, 7, 85–121. [Google Scholar] [CrossRef]
- Marquet, J.-C. Paléoenvironnement et Chronologie Des Sites Du Domaine Atlantique Fraçais d’âge Pléistocene Moyen et Supérieur d’après l’étude Des Rongeurs. Ph.D. Thesis, Dijon University, Dijon, France, 1993. [Google Scholar]
- Le Tensorer, J.-M. Le Paléolithique de l’Agenais; Cahiers du Quaternaire; Editions du Centre National de la Recherche Scientifique: Paris, France, 1981; 526p. [Google Scholar]
- Texier, J.-P.; Bertran, P. Nouvelle Interprétation Paléoenvironnementale et Chronostratigraphique Du Site Paléolithique de La Micoque (Dordogne): Implications Archéologiques. Comptes Rendus Acad. Sci. Paris 1993, 316, 1611–1617. [Google Scholar]
- Texier, J.-P. Histoire Géologique de Sites Préhistoriques Classiques Du Périgord: Une Vision Actualisée La Micoque, La Grotte Vaufrey, Le Pech de l’Azé I et II, La Ferrassie, l’abri Castanet, Le Flageolet, Laugerie Haute; Documents Préhistoriques; Comité des Travaux Historiques et Scientifiques: Paris, France, 2009; p. 193. [Google Scholar]
- Coumont, M.-P. Taphonomie Préhistorique: Mammifères Fossiles en Contexte d’avens Pièges. Apport Pour L’étude des Archeofaunes. Ph.D. Thesis, Aix-Marseille I University, Aix-en-Provence, France, 2006. [Google Scholar]
- Jeannet, M.; Mein, P. Les Muridae (Mammalia, Rodentia) Du Pléistocène Moyen de l’Igue Des Rameaux (Tarn-et-Garonne, France). PALEO 2016, 27, 177–205. [Google Scholar] [CrossRef]
- Fosse, P. L’hyène Des Cavernes Comme Agent d’accumulation Des Ossements à Lunel-Viel 1 (Hérault, France): Observations Préliminaires. In Outillage peu Élaboré en os et bois de Cervidés; CERDAC: Treignes, Belgium, 1994; pp. 91–96. [Google Scholar]
- Fosse, P. Taphonomie Paléolithique: Les Grands Mammifères de Soleilhac (Haute-Loire) et de Lunel-Viel 1 (Hérault). Ph.D. Thesis, Provence University Aix-Marseille I, Marseille, France, 1994. [Google Scholar]
- Fosse, P. La Grotte N° 1 de Lunel-Viel (Hérault, France): Repaire d’hyènes Du Pléistocène Moyen. Etude Taphonomique Du Matériel Osseux. PALEO 1996, 8, 47–79. [Google Scholar] [CrossRef]
- Uzunidis, A. Dental Wear Analyses of Middle Pleistocene Site of Lunel-Viel (Hérault, France): Did Equus and Bos Live in a Wetland? Quat. Int. 2020, 557, 39–46. [Google Scholar] [CrossRef]
- Desclaux, E.; El Hazzazi, N.; Vilette, P.; Dubar, M. Le contexte environnemental des occupations humaines. L’apport de la microfaune, des restes aviaires et de la malacofaune. In Le site de Payre. Occupations Humaines dans la Vallée du Rhône à la fin du Pléistocène Moyen et au Début du Pléistocène Supérieur; Moncel, M.-H., Ed.; Mémoires de la Société Préhistorique Française; Société Préhistorique Française: Paris, France, 2008; pp. 91–105. [Google Scholar]
- Ecker, M.; Bocherens, H.; Julien, M.-A.; Rivals, F.; Raynal, J.-P.; Moncel, M.-H. Middle Pleistocene Ecology and Neanderthal Subsistence: Insights from Stable Isotope Analyses in Payre (Ardèche, Southeastern France). J. Hum. Evol. 2013, 65, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Bocherens, H.; Díaz-Zorita Bonilla, M.; Daujeard, C.; Fernandes, P.; Raynal, J.-P.; Moncel, M.-H. Direct Isotopic Evidence for Subsistence Variability in Middle Pleistocene Neanderthals (Payre, Southeastern France). Quat. Sci. Rev. 2016, 154, 226–236. [Google Scholar] [CrossRef]
- Dessart, C. Etude de La Faune de La Grotte de Rigabe (Var) (Fouilles E. Bonifay); Post-Graduate Work, Aix-Marseille I-Université de Provence: Marseille, France, 2001. [Google Scholar]
- Paquereau, M.-M. Flore Des Deux Derniers Interglaciaires Dans Le Sud-Ouest de La France. L’Anthropologie 1976, 80, 201–228. [Google Scholar]
- Griggo, C. Significations Environnementales Des Communautés Animales Pléistocènes Reconnues à l’Abri Suard (Charente) et à La Grotte Du Bois-Ragot (Vienne). Ph.D. Thesis, Bordeaux 1 University, Bordeaux, France, 1995. [Google Scholar]
- Griggo, C. Etablissement de Courbes Climatiques Quantifiées à Partir Des Communautés Animales Pléistocènes Suivi d’une Application Aux Gisements de l’Abri Suard (Charente) et La Grotte de Bois-Ragot (Vienne). PALEO 1996, 8, 81–97. [Google Scholar] [CrossRef]
- Paquereau, M.-M. Etude Palynologique Du Wurm I Du Pech de l’Azé (Dordogne). Quaternaria 1969, XI, 227–233. [Google Scholar]
- Goldberg, P. Micromorphology of Pech-de-l’Azé II Sediments. J. Archaeol. Sci. 1979, 6, 17–47. [Google Scholar] [CrossRef]
- Lenoble, A. Le Rôle Du Ruissellement Dans La Formation Des Sites Préhistoriques: Approche Expérimentale. Ph.D. Thesis, Bordeaux 1 University, Bordeaux, France, 2003. [Google Scholar]
- Uzunidis, A. Les Grands Herbivores de La Couche 9 (Fin Pléistocène Moyen) de Coudoulous II (Tour-de-Faure, Lot): Paléontologie, Paléoécologie, Taphonomie; Post-Graduate Work, Aix-Marseille University: Aix-en-Provence, France, 2013. [Google Scholar]
- Astre, G. Brèche Pléistocène de Peyre (Aveyron), Avec Equus hydruntinus et Megaceros. Bull. Soc. Hist. Nat. Toulouse 1963, 98, 321–331. [Google Scholar]
- Brugal, J.-P. Recherches Préliminaires Sur Les Faunes Des Grands Causses et Vallées Périphériques. Paléobiol. Cont. 1981, 12, 145–158. [Google Scholar]
- Vernet, J.-L.; Ambert, P.; Andre, J.; Brugal, J.-P.; Casanova, J.; Michaux, J.; Semah, F. Les Travertins Pléistocènes de La Vallée Du Tarn, Dans Le Sud Du Massif Central: Intérêts Paléoécologiques. In Proceedings of the Paléoécologie, Lyon, France, 18–23 July 1983. [Google Scholar]
- Brugal, J.-P.; Jaubert, J. Les Gisements Paléontologiques Pléistocènes à Indice de Fréquentation Humaine: Un Nouveau Type de Comportement Prédateur ? PALEO 1991, 3, 15–41. [Google Scholar] [CrossRef]
- Brugal, J.-P. Un repaire de hyène des cavernes et de porc-épic à Peyre (Comprégnac, Aveyron). In Les Hommes et leurs Environnements Quaternaires dans les Causses de l’Aveyron; Ambert, P., Vernet, J.-L., Eds.; Association des amis du Musée de Millau: Millau, France, 1993; pp. 47–48. [Google Scholar]
- Sondaar, P.Y. The Osteology of the Manus of Fossil and Recent Equidae with Special Reference to Phylogeny and Function; Verhandelingen der Koninklijke Nederlandse Akademie van Wetenschappen, Afd. Natuurkunde; N.V. Noord-Hollandsche Utgevers Maatschappij: Amsterdam, The Netherlands, 1968. [Google Scholar]
- Gromova, V. Le Genre “Equus”, Premiere Partie: Revue et Description des Formes. In Annales du Centre d’Etudes et de Documentation Paléontologiques, 1955; n° 13; Pietresson de, S.-A., Translator; Travauxde l’Institut Paléontologique, Académie des Sciences de l’URSS: Paris, France, 1949; Volume 1, t. 17. [Google Scholar]
- Eisenmann, V. Sur Quelques Caractères Adaptatifs Du Squelette d’Equus et Leurs Implications Paléoécologiques. Bull. Muséum Hist. Nat. 1984, 6, 185–195. [Google Scholar]
- Kuzmina, I.E. Horses of North Eurasia from the Pliocene till the present time. In Proceedings of the Zoological Institute; Vereschagin, N.K., Ed.; Russian Academy of Sciences: Saint-Petersbourg, Russia, 1997; Volume 273. [Google Scholar]
- Cramer, B. Morphometrische Untersuchungen an Quartären Pferden in Mitteleuropa. Ph.D. Thesis, Geowissenschaftliche Fakultät, Eberhard-Karls-Universität, Tübingen, Germany, 2002. [Google Scholar]
- Levine, M.A. The use of crown height measurements and eruption-wear sequences to age horse teeth. In Ageing and Sexing of Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; BAR British Series; BAR Publishing: Oxford, UK, 1982; pp. 223–250. [Google Scholar]
- Van Asperen, E.N. Implications of Age Variation and Sexual Dimorphism in Modern Equids for Middle Pleistocene Equid Taxonomy. Int. J. Osteoarchaeol. 2013, 23, 1–12. [Google Scholar] [CrossRef]
- Eisenmann, V. Vera Eisenmann. Available online: https://vera-eisenmann.com/ (accessed on 20 May 2021).
- Eisenmann, V.; Alberdi, M.T.; De Giuli, C.; Staesche, U. Methodology. In Studying Fossil Horses; Woodburne, M.-O., Sondaar, P.Y., Eds.; E. J. Brill: Leiden, The Netherlands, 1988; Volume 1, pp. 1–71. [Google Scholar]
- Eisenmann, V. Les métapodes d’Equus sensu lato (Mammalia, Perissodactyla). Géobios 1979, 12, 863–886. [Google Scholar] [CrossRef]
- Scott, K.M. Postcranial dimensions of ungulates as predictors of body mass. In Body Size in Mammalian Palaeobiology—Estimation and Biological Implications; Damuth, J., MacFadden, B.J., Eds.; Cambridge University Press: New York, NY, USA, 1990; pp. 301–355. [Google Scholar]
- Simpson, G.G. Large Pleistocene Felines of North America; American Museum Novitates; American Museum of Natural History: New York, NY, USA, 1941. [Google Scholar]
- Eisenmann, V. Proportions Squelettiques de Chevaux Quaternaires et Actuels. Géobios 1991, 13, 25–32. [Google Scholar] [CrossRef]
- Ackermans, N.L.; Martin, L.F.; Codron, D.; Hummel, J.; Kircher, P.R.; Richter, H.; Kaiser, T.M.; Clauss, M.; Hatt, J.-M. Mesowear Represents a Lifetime Signal in Sheep (Ovis Aries) within a Long-Term Feeding Experiment. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 553, 109793. [Google Scholar] [CrossRef]
- Fortelius, M.; Solounias, N. Functional Characterization of Ungulate Molars Using the Abrasion-Attrition Wear Gradient: A New Method for Reconstructing Paleodiets. Am. Mus. Novit. 2000, 2000, 1–36. [Google Scholar] [CrossRef]
- Grine, F.E. Dental Evidence for Dietary Differences in Australopithecus and Paranthropus: A Quantitative Analysis of Permanent Molar Microwear. J. Hum. Evol. 1986, 15, 783–822. [Google Scholar] [CrossRef]
- Hoffman, J.M.; Fraser, D.; Clementz, M.T. Controlled Feeding Trials with Ungulates: A New Application of in Vivo Dental Molding to Assess the Abrasive Factors of Microwear. J. Exp. Biol. 2015, 218, 1538–1547. [Google Scholar] [CrossRef] [Green Version]
- Merceron, G.; Ramdarshan, A.; Blondel, C.; Boisserie, J.-R.; Brunetiere, N.; Francisco, A.; Gautier, D.; Milhet, X.; Novello, A.; Pret, D. Untangling the Environmental from the Dietary: Dust Does Not Matter. Proc. Biol. Sci. 2016, 283. [Google Scholar] [CrossRef] [Green Version]
- Teaford, M.F. A Review of Dental Microwear and Diet in Modern Mammals. Scanning Microsc. 1988, 2, 1149–1166. [Google Scholar] [PubMed]
- Winkler, D.E.; Schulz-Kornas, E.; Kaiser, T.M.; Codron, D.; Leichliter, J.; Hummel, J.; Martin, L.F.; Clauss, M.; Tütken, T. The Turnover of Dental Microwear Texture: Testing the “Last Supper” Effect in Small Mammals in a Controlled Feeding Experiment. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 557, 109930. [Google Scholar] [CrossRef]
- Rivals, F.; Moncel, M.-H.; Patou-Mathis, M. Seasonality and Intra-Site Variation of Neanderthal Occupations in the Middle Palaeolithic Locality of Payre (Ardèche, France) Using Dental Wear Analyses. J. Archaeol. Sci. 2009, 36, 1070–1078. [Google Scholar] [CrossRef]
- Rivals, F.; Mihlbachler, M.C.; Solounias, N. Effect of Ontogenetic-Age Distribution in Fossil and Modern Samples on the Interpretation of Ungulate Paleodiets Using the Mesowear Method. J. Vertebr. Paleontol. 2007, 27, 763–767. [Google Scholar] [CrossRef]
- Saarinen, J.; Lister, A.M. Dental Mesowear Reflects Local Vegetation and Niche Separation in Pleistocene Proboscideans from Britain. J. Quat. Sci. 2016, 31, 799–808. [Google Scholar] [CrossRef]
- Kaiser, T.M.; Solounias, N. Extending the Tooth Mesowear Method to Extinct and Extant Equids. Geodiversitas 2003, 25, 321–345. [Google Scholar]
- Mihlbachler, M.C.; Rivals, F.; Solounias, N.; Semprebon, G.M. Dietary Change and Evolution of Horses in North America. Science 2011, 331, 1178–1181. [Google Scholar] [CrossRef] [PubMed]
- Rivals, F.; Rindel, D.; Belardi, J.B. Dietary Ecology of Extant Guanaco (Lama Guanicoe) from Southern Patagonia: Seasonal Leaf Browsing and Its Archaeological Implications. J. Archaeol. Sci. 2013, 40, 2971–2980. [Google Scholar] [CrossRef]
- Uzunidis, A.; Rivals, F.; Brugal, J.-P. Relation between Morphology and Dietary Traits in Horse Jugal Upper Teeth during the Middle Pleistocene in Southern France. Quat. Rev. Assoc. Fr. Pour Létude Quat. 2017, 28, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Rivals, F.; Uzunidis, A.; Sanz, M.; Daura, J. Faunal Dietary Response to the Heinrich Event 4 in Southwestern Europe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 473, 123–130. [Google Scholar] [CrossRef]
- Solounias, N.; Semprebon, G. Advances in the Reconstruction of Ungulate Ecomorphology with Application to Early Fossil Equids. Am. Mus. Novit. 2002, 3366, 49. [Google Scholar] [CrossRef]
- Semprebon, G.; Godfrey, L.R.; Solounias, N.; Sutherland, M.R.; Jungers, W.L. Can Low-Magnification Stereomicroscopy Reveal Diet? J. Hum. Evol. 2004, 115–144. [Google Scholar] [CrossRef]
- Xafis, A.; Nagel, D.; Bastl, K. Which Tooth to Sample? A Methodological Study of the Utility of Premolar/Non-Carnassial Teeth in the Microwear Analysis of Mammals. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 487, 229–240. [Google Scholar] [CrossRef]
- Uzunidis, A.; Pineda, A.; Jiménez-Manchón, S.; Xafis, A.; Ollivier, V.; Rivals, F. The Impact of Sediment Abrasion on Tooth Microwear Analysis: An Experimental Study. Archaeol. Anthropol. Sci. 2021, 13, 134. [Google Scholar] [CrossRef]
- Walker, A.; Hoeck, H.N.; Perez, L. Mecrowear of Mammalian Teeth as an Indicator of Diet. Science 1978, 201, 908–910. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Rivals, F. Was Grass More Prevalent in the Pronghorn Past? An Assessment of the Dietary Adaptations of Miocene to Recent Antilocapridae (Mammalia: Artiodactyla). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 253, 332–347. [Google Scholar] [CrossRef]
- Semprebon, G.M.; Sise, P.J.; Coombs, M.C. Potential Bark and Fruit Browsing as Revealed by Stereomicrowear Analysis of the Peculiar Clawed Herbivores Known as Chalicotheres (Perissodactyla, Chalicotherioidea). J. Mamm. Evol. 2011, 18, 33–55. [Google Scholar] [CrossRef]
- Kaczensky, P.; Ganbaatar, O.; Wehrden, H.V.; Walzer, C. Resource Selection by Sympatric Wild Equids in the Mongolian Gobi. J. Appl. Ecol. 2008, 45, 1762–1769. [Google Scholar] [CrossRef]
- Uzunidis, A. Large Ungulates Mobility and Neanderthal Subsistence Behaviours: A Preliminary Tooth Microwear Analysis. J. Archaeol. Sci. Rep. 2020, 29, 102084. [Google Scholar] [CrossRef]
- Foronova, I. Late Quaternary Equids (Genus Equus) of South-Western and Southcentral Siberia. In Proceedings of the Equids in Time and Space: Papers in Honour of Véra Eisenmann: Proceedings of the 9th Conference of the International Council of Archaeozoology, Durham, UK, 23–28 August 2002; Mashkour, M., Ed.; Août 2002; Oxbow Books: Oxford, UK, 2006; pp. 20–30. [Google Scholar]
- Forsten, A. Middle Pleistocene Replacement of Stenonoid Horses by Caballoid Horses—Ecological Implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. Elsevier Sci. Publ. BV 1988, 65, 23–33. [Google Scholar] [CrossRef]
- Bro-Jørgensen, J. Dense Habitats Selecting for Small Body Size: A Comparative Study on Bovids. Oikos 2008, 117, 729–737. [Google Scholar] [CrossRef]
- Lindstedt, S.L.; Boyce, M.S. Seasonality, Fasting Endurance, and Body Size in Mammals. Am. Nat. 1985, 125, 873–878. [Google Scholar] [CrossRef]
- El Hazzazi, N. Apport Du Genre Arvicola (Rodentia, Mammalia) Dans La Connaissance de La Chronostratigraphie Des Sites Pléistocène Moyen et Supérieur: Orgnac 3, Payre 2 et l’abri Des Pêcheurs (Ardèche, France). In Proceedings of the Theoretical and Methodological Problems—Archaeometry—Paleoecology, Forli, Italy, 8–14 September 1996; Arias, C., Bietti, A., Castelletti, L., Peretto, C., Eds.; A.B.A.C.O. Edizioni: Forli, Italy, 1998; Volume 1, pp. 125–133. [Google Scholar]
- Moncel, M.-H.; Debard, E.; Desclaux, E.; Dubois, J.-M.; Lamarque, F.; Patou-Mathis, M.; Villette, P. Le Cadre de Vie Des Hommes Du Paléolithique Moyen (Stades Isotopiques 6 et 5) Dans Le Site de Payre (Rompon, Ardèche): D’une Grotte à Un Abri Sous Roche Effondré. Bull. Soc. Préhistorique Fr. 2002, 99, 249–273. [Google Scholar] [CrossRef] [Green Version]
- Schulz, E.; Kaiser, T.M. Historical Distribution, Habitat Requirements and Feeding Ecology of the Genus Equus (Perissodactyla). Mammal Rev. 2013, 43, 111–123. [Google Scholar] [CrossRef]
- Kaiser, T.M.; Franz-Odendaal, T.A. A Mixed-Feeding Equus Species from the Middle Pleistocene of South Africa. Quat. Res. 2004, 62, 316–323. [Google Scholar] [CrossRef]
- Valli, A.M.F.; Palombo, M.R.; Alberdi, M.T. How Homogeneous Are Microwear Patterns on a Fossil Horse Tooth? Preliminary Test on a Premolar of Equus Altidens from Barranco Leon 5 (Spain). Alp. Mediterr. Quat. 2012, 25, 25–33. [Google Scholar]
- Kelly, A.; Miller, J.H.; Wooller, M.J.; Seaton, C.T.; Druckenmiller, P.; DeSantis, L. Dietary Paleoecology of Bison and Horses on the Mammoth Steppe of Eastern Beringia Based on Dental Microwear and Mesowear Analyses. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 572, 110394. [Google Scholar] [CrossRef]
- Rivals, F.; Kitagawa, K.; Julien, M.-A.; Patou-Mathis, M.; Bessudnov, A.A.; Bessudnov, A.N. Straight from the Horse’s Mouth: High-Resolution Proxies for the Study of Horse Diet and Its Relation to the Seasonal Occupation Patterns at Divnogor’ye 9 (Middle Don, Central Russia). Quat. Int. 2018, 474, 146–155. [Google Scholar] [CrossRef]
- Delpech, F.; Le Tensorer, J.-M.; Pineda, R.; Prat, F. Un Nouveau Gisement Du Pleistocène Moyen: Camp-de-Peyre à Sauveterre-La-Lémance (Lot-et-Garonne). Comptes Rendus Acad. Sci. Paris D 1978, 286, 1101–1103. [Google Scholar]
- Saarinen, J.; Eronen, J.; Fortelius, M.; Seppä, H.; Lister, A.M. Patterns of Diet and Body Mass of Large Ungulates from the Pleistocene of Western Europe, and Their Relation to Vegetation. Palaeontol. Electron. 2016, 19.3.32A, 1–58. [Google Scholar] [CrossRef]
- Geist, V. Deer of the World: Their Evolution, Behaviour, and Ecology; Stackpole Books: Mechanicsburg, PA, USA, 1998; ISBN 978-0-8117-0496-0. [Google Scholar]
- Guthrie, R.D. Origin and Causes of the Mammoth Steppe: A Story of Cloud Cover, Woolly Mammal Tooth Pits, Buckles, and inside-out Beringia. Quat. Sci. Rev. 2001, 20, 549–574. [Google Scholar] [CrossRef]
- Saarinen, J. Ecometrics of Large Herbivorous Land Mammals in Relation to Climatic and Environmental Changes during the Pleistocene. Ph.D. Thesis, Department of Geosciences and Geography, University of Helsinki, Helsinki, Finland, 2014. [Google Scholar]
- Blackburn, T.M.; Gaston, K.J.; Loder, N. Geographic Gradients in Body Size: A Clarification of Bergmann’s Rule. Divers. Distrib. 1999, 5, 165–174. [Google Scholar] [CrossRef]
- Boulbes, N.; van Asperen, E.N. Biostratigraphy and Palaeoecology of European Equus. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Richerson, P.J.; Boyd, R.; Bettinger, R.L. Was Agriculture Impossible during the Pleistocene but Mandatory during the Holocene? A Climate Change Hypothesis. Am. Antiq. 2001, 66, 387–411. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Location of the sites from the South West and South East of France selected for this study: Camp-de-Peyre, La Micoque, Rameaux, Lunel-Viel, Payre, Rigabe, Suard, Pech de l’Azé II, Coudoulous II and Peyre. Map: IGN 2012_Licence ouverte, modified A. Uzunidis (http://education.ign.fr/primaire/fonds-de-cartes (accessed on 20 May 2018)).
Figure 1.
Location of the sites from the South West and South East of France selected for this study: Camp-de-Peyre, La Micoque, Rameaux, Lunel-Viel, Payre, Rigabe, Suard, Pech de l’Azé II, Coudoulous II and Peyre. Map: IGN 2012_Licence ouverte, modified A. Uzunidis (http://education.ign.fr/primaire/fonds-de-cartes (accessed on 20 May 2018)).

Figure 2.
Photomicrographs of horse teeth from Suard (n° 52-K14-50) and Peyre (n° E9-347) with scratches and pits. Magnification ×35.
Figure 2.
Photomicrographs of horse teeth from Suard (n° 52-K14-50) and Peyre (n° E9-347) with scratches and pits. Magnification ×35.

Figure 3.
Box plot of the estimated weight in kilograms of Southern France Middle and Late Pleistocene caballoid horses based on metacarpal and metatarsal measurements (MC2, MC4, and MC6, MT2, MT4 and MT6 of Scott [79]). Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus. Vertically: in grey, South East sites, in black South West sites. Horizontally: in grey temperate isotopic stages, in black glacial isotopic stages.
Figure 3.
Box plot of the estimated weight in kilograms of Southern France Middle and Late Pleistocene caballoid horses based on metacarpal and metatarsal measurements (MC2, MC4, and MC6, MT2, MT4 and MT6 of Scott [79]). Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus. Vertically: in grey, South East sites, in black South West sites. Horizontally: in grey temperate isotopic stages, in black glacial isotopic stages.

Figure 4.
Log ratio diagram of humerus, femur, radius, tibia, third metacarpal and metatarsal, first anterior and posterior phalanges greatest length and third phalanx greatest width for caballoid horses from the South of France. Reference E. hemionus [78]. The number of specimens is indicated for each population. Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
Figure 4.
Log ratio diagram of humerus, femur, radius, tibia, third metacarpal and metatarsal, first anterior and posterior phalanges greatest length and third phalanx greatest width for caballoid horses from the South of France. Reference E. hemionus [78]. The number of specimens is indicated for each population. Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.

Figure 5.
Principal component analysis (PCA) with measurements of the third metacarpal of Southern Middle and Late Pleistocene horses from the caballoid lineage. The correlation circle illustrates the contribution of each variable. M. 1: Maximal length, M. 3: Breadth at the middle of the diaphysis; M. 4: Depth of the diaphysis at the same level; M. 5: Proximal breadth; M. 6: Proximal depth; M. 11: Distal articular breadth; M.14: Greatest depth of the medial condyle. The large dots correspond to the centroid of each cohort. Camp-de-Peyre: E. m. campdepeyri; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Coudoulous II l.9: E. mosbachensis; Peyre: E. caballus.
Figure 5.
Principal component analysis (PCA) with measurements of the third metacarpal of Southern Middle and Late Pleistocene horses from the caballoid lineage. The correlation circle illustrates the contribution of each variable. M. 1: Maximal length, M. 3: Breadth at the middle of the diaphysis; M. 4: Depth of the diaphysis at the same level; M. 5: Proximal breadth; M. 6: Proximal depth; M. 11: Distal articular breadth; M.14: Greatest depth of the medial condyle. The large dots correspond to the centroid of each cohort. Camp-de-Peyre: E. m. campdepeyri; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Coudoulous II l.9: E. mosbachensis; Peyre: E. caballus.

Figure 6.
Principal component analysis (PCA) with measurements of the third metatarsals of southern Middle and Late Pleistocene horses from the caballoid lineage. The correlation circle illustrates the contribution of each variable. M. 1: Maximal length, M. 3: Breadth at the middle of the diaphysis; M. 4: Depth of the diaphysis at the same level; M. 5: Proximal breadth; M. 11: Distal articular breadth; M.14: Greatest depth of the medial condyle. The large dots correspond to the centroid of each cohort. Camp-de-Peyre: E. m. campdepeyri; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
Figure 6.
Principal component analysis (PCA) with measurements of the third metatarsals of southern Middle and Late Pleistocene horses from the caballoid lineage. The correlation circle illustrates the contribution of each variable. M. 1: Maximal length, M. 3: Breadth at the middle of the diaphysis; M. 4: Depth of the diaphysis at the same level; M. 5: Proximal breadth; M. 11: Distal articular breadth; M.14: Greatest depth of the medial condyle. The large dots correspond to the centroid of each cohort. Camp-de-Peyre: E. m. campdepeyri; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.

Figure 7.
Low scratch percentages (0%–17%) and mesowear scores for each cohort through time. The grey areas correspond to the range of values for extant mixed feeders. Camp-de-Peyre: E. m. campdepeyri; La Micoque l.E, H, J: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard l. 53-50: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
Figure 7.
Low scratch percentages (0%–17%) and mesowear scores for each cohort through time. The grey areas correspond to the range of values for extant mixed feeders. Camp-de-Peyre: E. m. campdepeyri; La Micoque l.E, H, J: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard l. 53-50: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.

Figure 8.
Bivariate plot of the average numbers of pits and scratches in horse cohorts from the South of France. Error bars correspond to standard deviations for the fossil samples. Plain ellipses correspond to Gaussian confidence ellipses (p = 0.95) on the centroid for extant leaf browsers and grazers from Solounias and Semprebon [97]. Camp-de-Peyre: E. m. campdepeyri; La Micoque l.E, H, J: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard l. 53-50: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
Figure 8.
Bivariate plot of the average numbers of pits and scratches in horse cohorts from the South of France. Error bars correspond to standard deviations for the fossil samples. Plain ellipses correspond to Gaussian confidence ellipses (p = 0.95) on the centroid for extant leaf browsers and grazers from Solounias and Semprebon [97]. Camp-de-Peyre: E. m. campdepeyri; La Micoque l.E, H, J: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard l. 53-50: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.

Figure 9.
Principal component analysis (PCA) with measurements of the third metacarpals and metatarsals of Southern Middle and Late Pleistocene caballoid horses with regards to the dietary categories displayed by meso- and microwear analysis. G-G: grazer all year long; G-MF: generally, grazer, mixed-feeder at the time of death; G-B: generally, grazer, browser at the time of death. The large dots correspond to the centroid of each dietary category. For the correlation circles, see Figure 5 for metacarpals and 6 for metatarsals.
Figure 9.
Principal component analysis (PCA) with measurements of the third metacarpals and metatarsals of Southern Middle and Late Pleistocene caballoid horses with regards to the dietary categories displayed by meso- and microwear analysis. G-G: grazer all year long; G-MF: generally, grazer, mixed-feeder at the time of death; G-B: generally, grazer, browser at the time of death. The large dots correspond to the centroid of each dietary category. For the correlation circles, see Figure 5 for metacarpals and 6 for metatarsals.


{kind=link}
{kind=link}
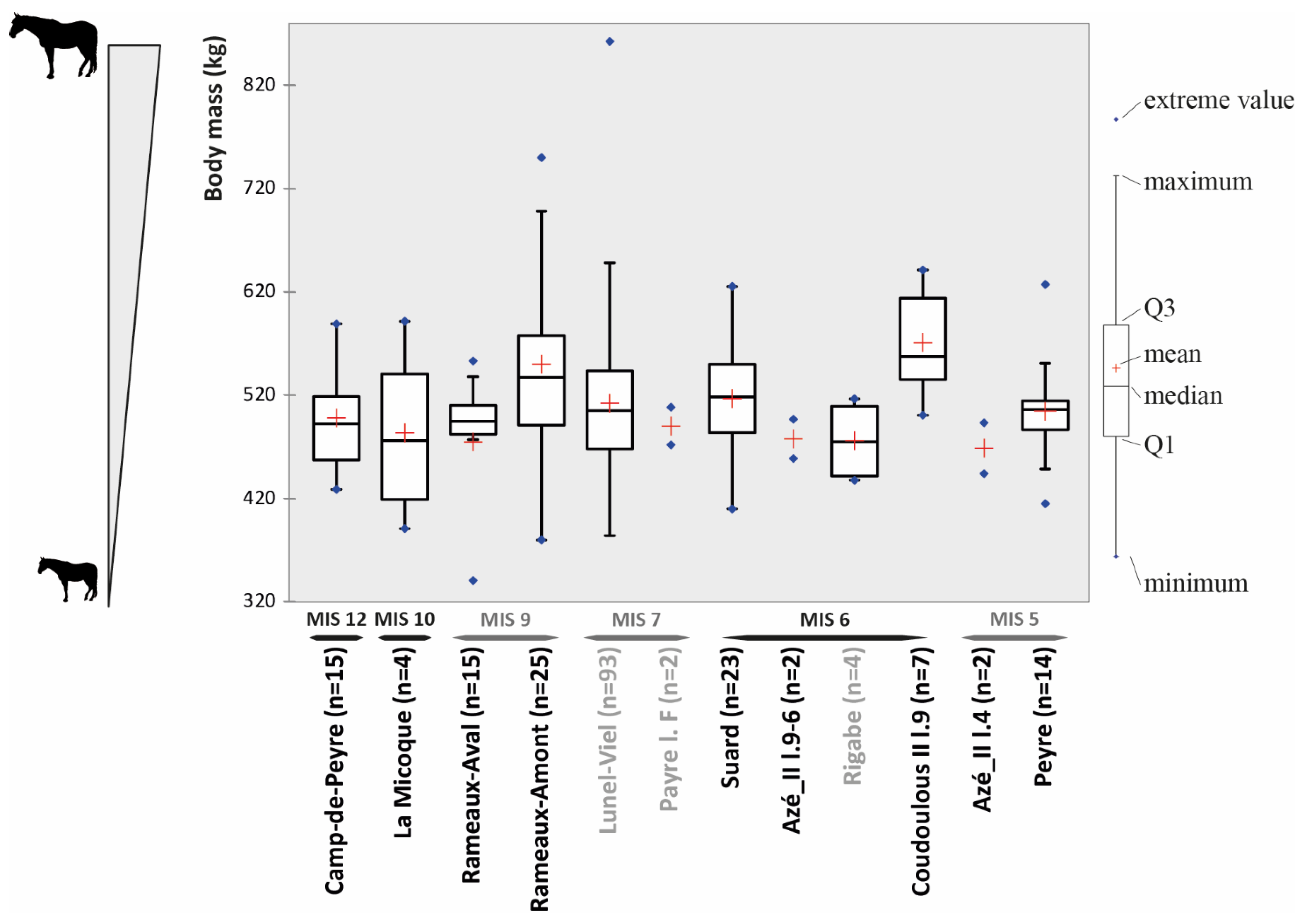{kind=link}
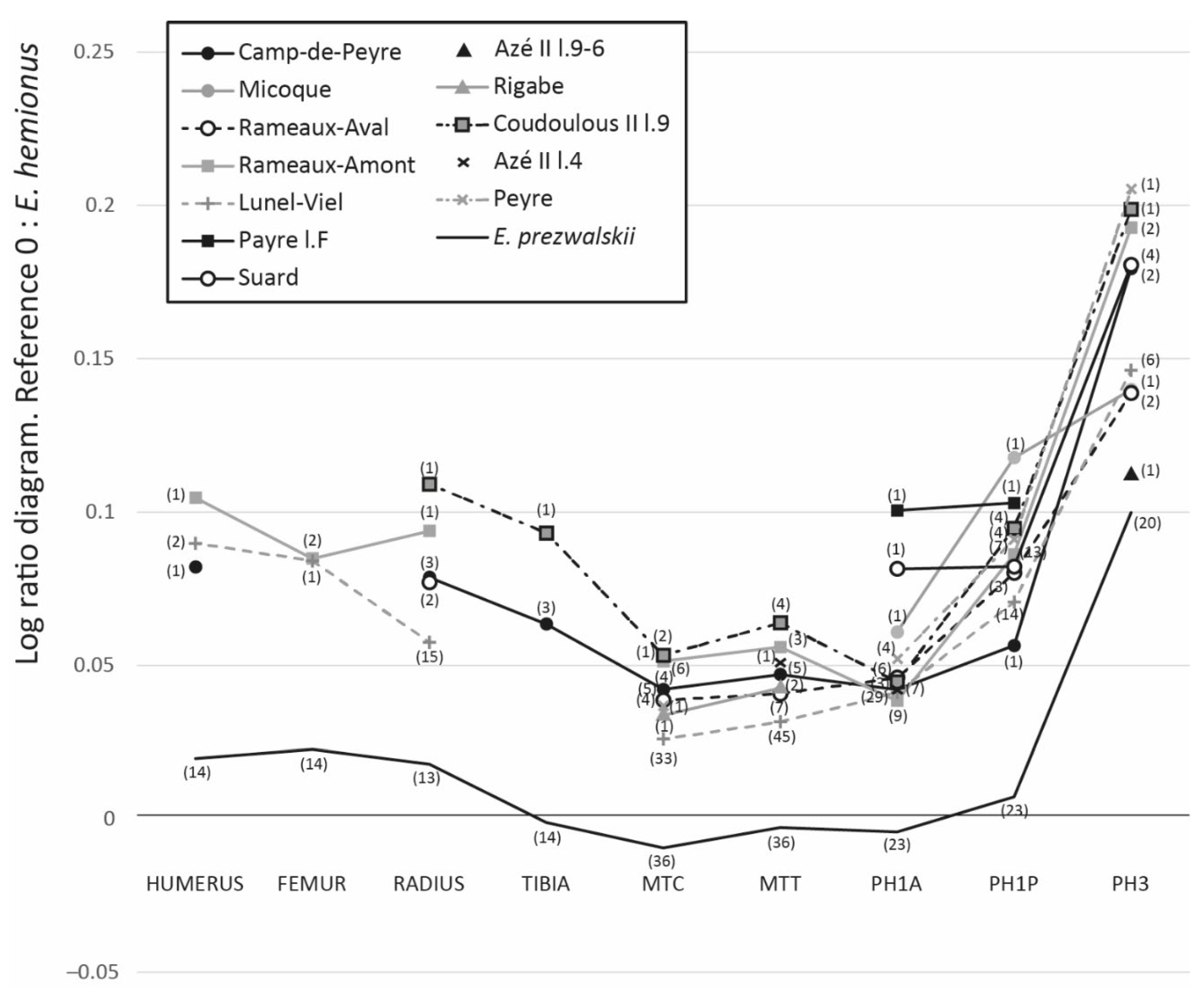{kind=link}
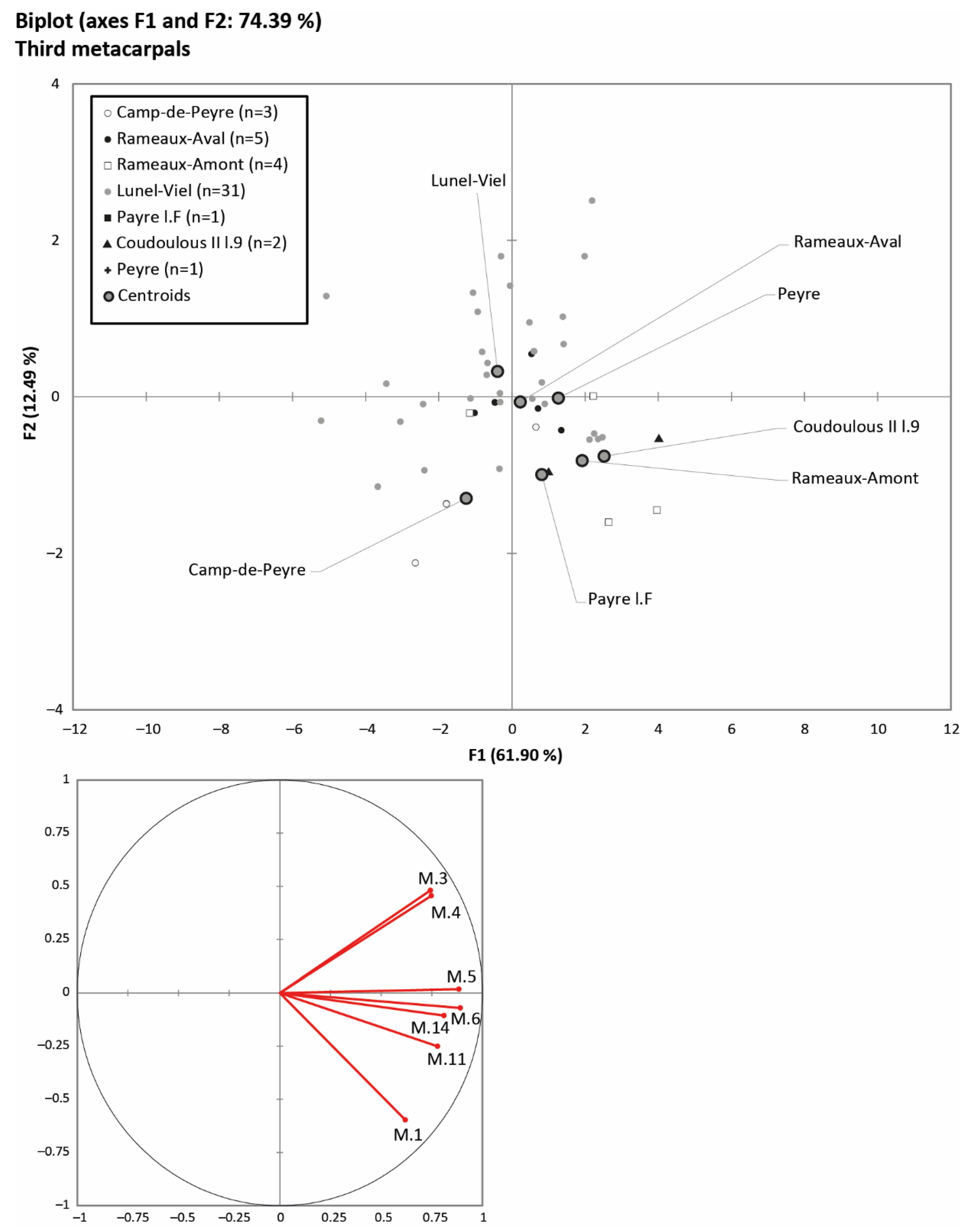{kind=link}
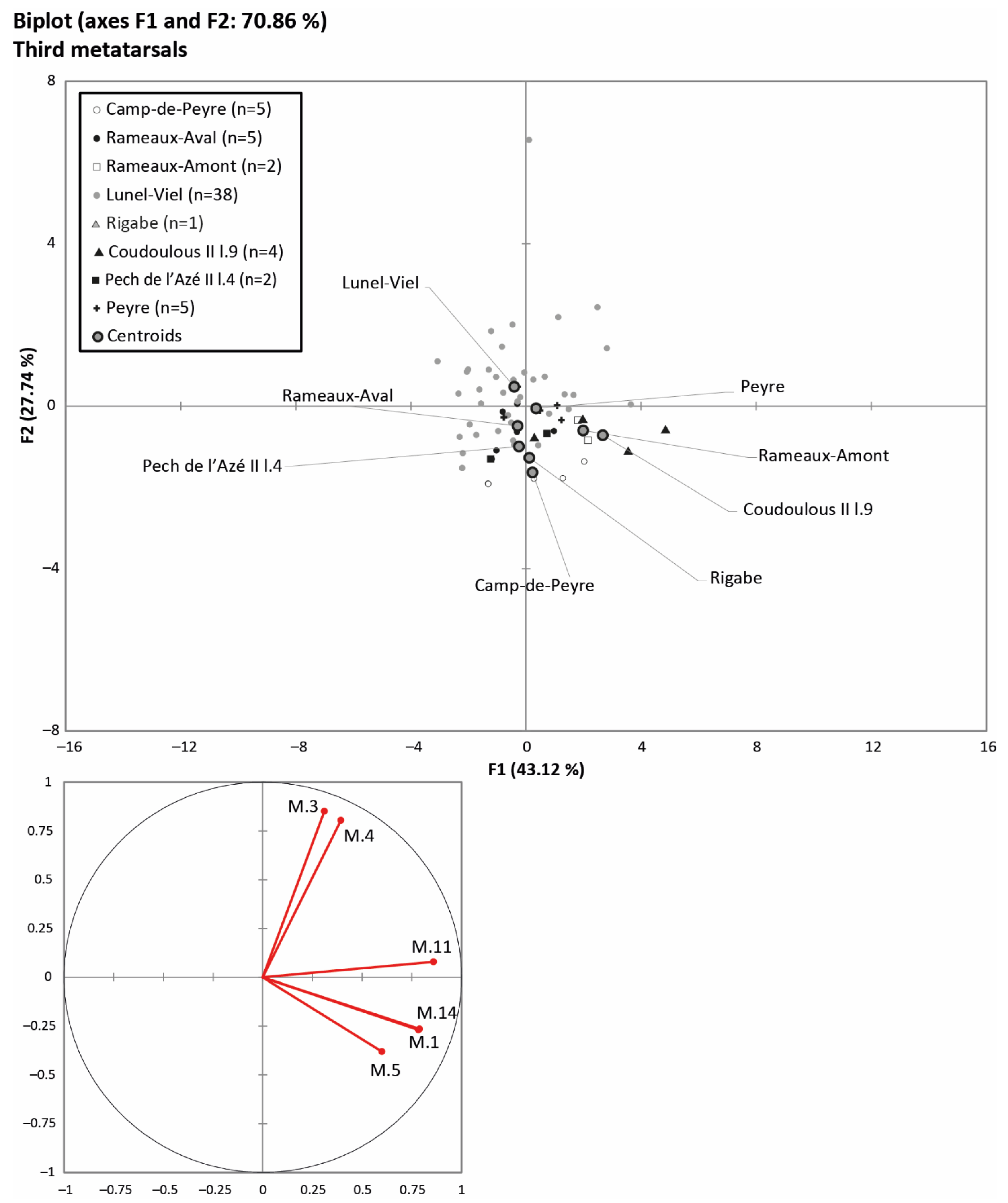{kind=link}
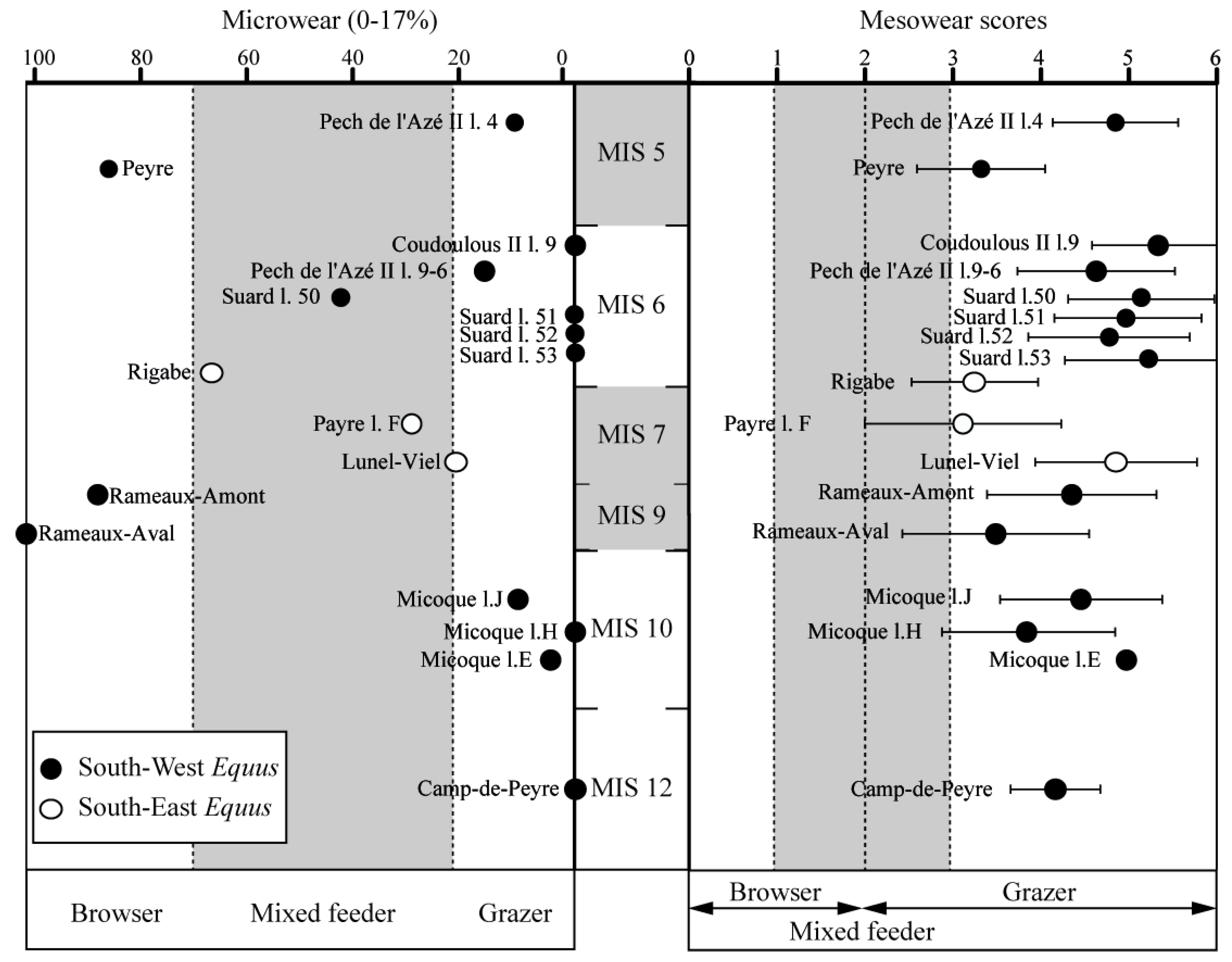{kind=link}
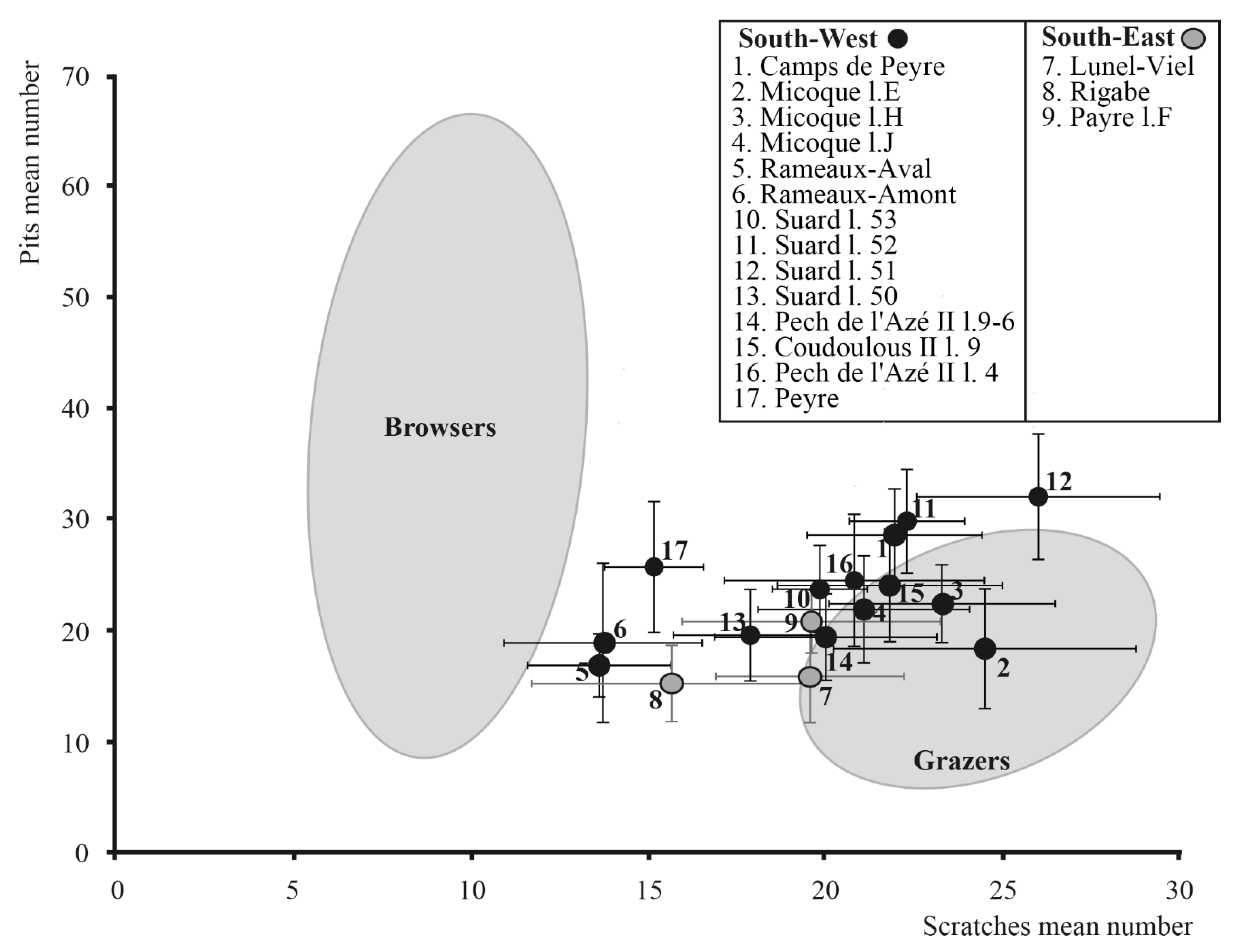{kind=link}
{kind=link}
Table 1.
Localities cited in this study with dating, location, location of the collection and enumeration of the bones and teeth studied. MMSH: Maison Méditérranéenne des Sciences de l’Homme; MNP: Musée National de Préhistoire. MIS 12: ca 478-424 ky; MIS 10: ca 374-337 ky; MIS9: ca 337-300 ky; MIS 7: ca 243-191 ky; MIS6: ca 191-130 ky; MIS 5: ca 130-71 ky (age interval for each isotopic stage in [22]).
Table 1.
Localities cited in this study with dating, location, location of the collection and enumeration of the bones and teeth studied. MMSH: Maison Méditérranéenne des Sciences de l’Homme; MNP: Musée National de Préhistoire. MIS 12: ca 478-424 ky; MIS 10: ca 374-337 ky; MIS9: ca 337-300 ky; MIS 7: ca 243-191 ky; MIS6: ca 191-130 ky; MIS 5: ca 130-71 ky (age interval for each isotopic stage in [22]).
| Locality | Region | Age | Reference for Dating | Location | Equus Specie | Reference for Palaeontological Determination | Number of Studied Bones | Number of Studied Teeth |
|---|---|---|---|---|---|---|---|---|
| Peyre | South-West | MIS 5 | [24] | MMSH | Equus caballus | [25] | 138 | 27 |
| Pech de l’Azé II l.4-2 | South-West | MIS 5 | [26] | Les Eyzies MNP | Equus caballus | [19,25] | 18 | 31 |
| Coudoulous II l.9 | South-West | MIS 6 | [27,28] | MMSH | Equus mosbachensis | [25,28] | 115 | 25 |
| Pech de l’Azé II l. 9-6 | South-West | MIS 6 | [26] | Les Eyzies MNP | Equus mosbachensis | [19,25] | 6 | 63 |
| Suard l.53-50 | South-West | MIS 6 | [29,30,31] | Angoulême museum | Equus caballus piveteaui | [25,32] | 190 | 116 |
| Rigabe | South-East | MIS 6 | [33] | Les Eyzies MNP | Equus mosbachensis palustris | [25] | 29 | 18 |
| Payre l. F | South-East | MIS 7 | [34] | Orgnac museum | Equus mosbachensis | [25] | 26 | 8 |
| Lunel-Viel | South-East | MIS 7 | [35] | Les Eyzies MNP | Equus mosbachensis palustris | [25,36] | 1109 | 65 |
| Rameaux-Amont | South-West | MIS 9 | [25,37,38] | MMSH | Equus mosbachensis | [25] | 247 | 151 |
| Rameaux-Aval | South West | MIS 9 | [25,37,38] | MMSH | Equus mosbachensis | [25] | 190 | 8 |
| La Micoque l. E, H, J | South-West | MIS 10 | [39] | Les Eyzies MNP | Equus mosbachensis | [25,40,41] | 38 | 34 |
| Camp-de-Peyre | South-West | MIS 12 | [42] | Les Eyzies MNP | Equus mosbachensis campdepeyri | [25,42] | 164 | 38 |
Table 2.
Measurements in millimetres of the greatest length of humerus, radius, femur, tibia, metapodials, first anterior phalanges (Phalanges I A), first posterior phalanges (Phalanges I P) and of the greatest width of the third phalange (Phalange III) of caballoid horses of the Middle and Late Pleistocene from France. n = number of specimen; m = mean. Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
Table 2.
Measurements in millimetres of the greatest length of humerus, radius, femur, tibia, metapodials, first anterior phalanges (Phalanges I A), first posterior phalanges (Phalanges I P) and of the greatest width of the third phalange (Phalange III) of caballoid horses of the Middle and Late Pleistocene from France. n = number of specimen; m = mean. Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
| Site/Bone | Humerus | Radius | Femur | Tibia | Metacarpal | Metatarsal | Phalange I A | Phalange I P | Phalange III | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | m | n | m | n | m | n | m | n | m | n | m | n | m | n | m | n | m | |
| Camp-de-Peyre | 1 | 300.15 | 3 | 355.43 | 3 | 373.27 | 4 | 242.69 | 5 | 287.78 | 7 | 86.9 | 1 | 82.65 | 2 | 85.03 | ||
| La Micoque | 1 | 90.73 | 1 | 95.18 | 1 | 77.65 | ||||||||||||
| Rameaux-Aval | 2 | 354.08 | 5 | 240.75 | 7 | 283.65 | 6 | 87.68 | 3 | 87.28 | 2 | 77.45 | ||||||
| Rameaux-Amont | 1 | 316.14 | 1 | 368 | 2 | 410.44 | 6 | 247.89 | 3 | 293.79 | 9 | 86.16 | 7 | 88.51 | 2 | 87.7 | ||
| Lunel-Viel | 2 | 305.51 | 15 | 338.48 | 1 | 409.74 | 33 | 233.81 | 45 | 277.75 | 29 | 86.7 | 14 | 85.4 | 6 | 78.78 | ||
| Payre l.F | 1 | 249 | 1 | 99.4 | 1 | 92 | ||||||||||||
| Suard | 3 | 95.14 | 13 | 87.7 | 4 | 85.27 | ||||||||||||
| Pech de l’Azé II l.9-6 | 1 | 72.81 | ||||||||||||||||
| Rigabe | 1 | 237.99 | 2 | 284.88 | ||||||||||||||
| Coudoulous II l.9 | 1 | 381.19 | 1 | 399.68 | 2 | 249 | 4 | 299.175 | 3 | 87.4 | 4 | 90.25 | 1 | 88.9 | ||||
| Pech de l’Azé II l.4 | 2 | 290.43 | ||||||||||||||||
| Peyre | 4 | 239.71 | 5 | 285.09 | 4 | 88.9 | 4 | 89.52 | 1 | 90.26 | ||||||||
Table 3.
Measurements in millimetres of the third metacarpals of horses from the caballoid lineage. 1: Maximal length; 3: Breadth at the middle of the diaphysis; 4: Depth of the diaphysis at the same level; 5: Proximal breadth; 6: Proximal depth; 10: Distal supra-articular breadth; 11: Distal articular breadth; 12: Depth of the sagittal crest; 14: Greatest depth of the medial condyle (measurements from V. Eisenmann [76]). n = number of specimens; m = mean; s = standard deviation; min-max: minimal and maximal values. Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Peyre: E. caballus.
Table 3.
Measurements in millimetres of the third metacarpals of horses from the caballoid lineage. 1: Maximal length; 3: Breadth at the middle of the diaphysis; 4: Depth of the diaphysis at the same level; 5: Proximal breadth; 6: Proximal depth; 10: Distal supra-articular breadth; 11: Distal articular breadth; 12: Depth of the sagittal crest; 14: Greatest depth of the medial condyle (measurements from V. Eisenmann [76]). n = number of specimens; m = mean; s = standard deviation; min-max: minimal and maximal values. Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Peyre: E. caballus.
| Site | 1 | 3 | 4 | 5 | 6 | 10 | 11 | 12 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Camp-de-Peyre | n | 4 | 7 | 7 | 4 | 5 | 4 | 4 | 4 | 4 |
| m | 242.70 | 37.92 | 29.42 | 54.29 | 36.95 | 53.33 | 52.97 | 38.82 | 32.99 | |
| s | 4.43 | 1.98 | 0.99 | 2.16 | 1.40 | 3.07 | 2.00 | 0.42 | 1.97 | |
| min | 238.25 | 35.98 | 28.18 | 51.34 | 34.55 | 51.05 | 50.60 | 38.21 | 30.47 | |
| max | 248.16 | 40.28 | 30.83 | 56.30 | 37.99 | 57.86 | 55.26 | 39.18 | 34.84 | |
| Micoque | n | 1 | 1 | 3 | 5 | 1 | 1 | 1 | 1 | |
| m | 43.65 | 26.49 | 55.19 | 35.88 | 52.03 | 53.03 | 40.60 | 34.18 | ||
| s | 3.04 | 1.21 | ||||||||
| min | 52.61 | 34.50 | ||||||||
| max | 58.54 | 37.67 | ||||||||
| Rameaux-Aval | n | 5 | 8 | 8 | 7 | 7 | 8 | 6 | 6 | 6 |
| m | 240.76 | 38.36 | 28.78 | 55.26 | 36.15 | 51.74 | 52.83 | 39.55 | 32.90 | |
| s | 4.08 | 4.69 | 2.69 | 1.97 | 2.11 | 2.66 | 0.97 | 1.18 | 0.96 | |
| min | 237.20 | 29.85 | 24.18 | 52.31 | 33.31 | 47.26 | 51.70 | 37.69 | 31.71 | |
| max | 247.40 | 42.71 | 31.18 | 57.66 | 38.86 | 54.10 | 54.19 | 40.72 | 33.80 | |
| Rameaux-Amont | n | 6 | 9 | 9 | 8 | 7 | 7 | 6 | 5 | 7 |
| m | 247.89 | 40.23 | 30.54 | 55.86 | 37.60 | 54.62 | 54.42 | 40.59 | 33.05 | |
| s | 12.18 | 2.03 | 0.88 | 2.43 | 2.05 | 3.06 | 2.55 | 1.98 | 1.88 | |
| min | 235.99 | 37.60 | 29.25 | 52.13 | 34.74 | 51.44 | 51.10 | 38.17 | 29.59 | |
| max | 267.10 | 42.77 | 31.75 | 60.13 | 40.83 | 59.53 | 56.97 | 42.57 | 35.01 | |
| Lunel-Viel | n | 33 | 43 | 44 | 37 | 38 | 37 | 37 | 34 | 37 |
| m | 233.81 | 39.54 | 30.26 | 54.13 | 36.35 | 51.77 | 52.07 | 40.47 | 32.90 | |
| s | 5.92 | 3.52 | 2.41 | 3.26 | 2.56 | 2.33 | 2.46 | 1.74 | 1.89 | |
| min | 221.20 | 26.16 | 24.26 | 46.82 | 31.34 | 47.17 | 45.51 | 37.53 | 27.79 | |
| max | 242.90 | 45.48 | 40.69 | 58.80 | 39.73 | 58.15 | 57.65 | 46.46 | 39.81 | |
| Payre l.F | n | 1 | 1 | 1 | 1 | 4 | 2 | 2 | 2 | 2 |
| m | 249.00 | 41.50 | 29.30 | 54.40 | 37.43 | 52.45 | 53.30 | 41.25 | 34.35 | |
| s | 2.06 | 0.35 | 0.14 | 1.20 | 0.07 | |||||
| min | 35.00 | 52.20 | 53.20 | 40.40 | 34.30 | |||||
| max | 40.00 | 52.70 | 53.40 | 42.10 | 34.40 | |||||
| Suard | n | 5 | 6 | 13 | 15 | 2 | 2 | 2 | 2 | |
| m | 42.87 | 31.02 | 56.30 | 36.81 | 56.28 | 55.42 | 41.23 | 34.26 | ||
| s | 1.40 | 0.61 | 2.01 | 1.15 | 0.34 | 0.21 | 0.57 | 0.30 | ||
| min | 41.39 | 30.20 | 53.67 | 33.88 | 56.04 | 55.27 | 40.83 | 34.05 | ||
| max | 45.15 | 32.02 | 60.05 | 38.61 | 56.52 | 55.56 | 41.63 | 34.47 | ||
| Pech de l’Azé II | n | 1 | 1 | 2 | ||||||
| l. 9-6 | m | 28.74 | 57.49 | 37.88 | ||||||
| s | 0.71 | |||||||||
| min | 37.38 | |||||||||
| max | 38.38 | |||||||||
| Rigabe | n | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | |
| m | 237.99 | 37.33 | 28.67 | 34.16 | 49.36 | 49.48 | 38.65 | 31.36 | ||
| Coudoulous II l.9 | n | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| m | 249.00 | 41.32 | 31.10 | 58.30 | 38.70 | 54.90 | 55.60 | 39.20 | 34.26 | |
| s | 1.41 | 2.91 | 0.42 | 2.83 | 2.12 | 1.56 | 2.83 | 1.70 | 0.21 | |
| min | 248.00 | 39.26 | 30.80 | 56.30 | 37.20 | 53.80 | 53.60 | 38.00 | 34.11 | |
| max | 250.00 | 43.37 | 31.40 | 60.30 | 40.20 | 56.00 | 57.60 | 40.40 | 34.40 | |
| Peyre | n | 4 | 4 | 4 | 2 | 2 | 4 | 4 | 3 | 4 |
| m | 239.72 | 39.75 | 30.45 | 57.54 | 37.12 | 50.24 | 50.90 | 38.92 | 32.40 | |
| s | 7.05 | 0.46 | 1.42 | 0.66 | 0.09 | 2.03 | 1.73 | 0.67 | 1.21 | |
| min | 233.59 | 39.22 | 29.08 | 57.07 | 37.05 | 47.22 | 48.36 | 38.19 | 30.64 | |
| max | 249.32 | 40.22 | 32.34 | 58.00 | 37.18 | 51.53 | 52.08 | 39.49 | 33.37 |
Table 4.
Measurements in millimetres of the third metatarsals of horses from the caballoid lineage. 1: Maximal length; 3: Breadth at the middle of the diaphysis; 4: Depth of the diaphysis at the same level; 5: Proximal breadth; 6: Proximal depth; 10: Distal supra-articular breadth; 11: Distal articular breadth; 12: Depth of the sagittal crest; 14: Greatest depth of the medial condyle (measurements from V. Eisenmann [76]). n= number of specimens; m=mean; s=standard deviation; min-max: minimal and maximal values. Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
Table 4.
Measurements in millimetres of the third metatarsals of horses from the caballoid lineage. 1: Maximal length; 3: Breadth at the middle of the diaphysis; 4: Depth of the diaphysis at the same level; 5: Proximal breadth; 6: Proximal depth; 10: Distal supra-articular breadth; 11: Distal articular breadth; 12: Depth of the sagittal crest; 14: Greatest depth of the medial condyle (measurements from V. Eisenmann [76]). n= number of specimens; m=mean; s=standard deviation; min-max: minimal and maximal values. Camp-de-Peyre: E. m. campdepeyri; La Micoque: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Suard: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
| Site | 1 | 3 | 4 | 5 | 6 | 10 | 11 | 12 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Camp-de-Peyre | n | 5 | 8 | 8 | 6 | 5 | 7 | 7 | 7 | 7 |
| m | 287.78 | 37.91 | 36.07 | 56.91 | 48.62 | 54.81 | 54.19 | 42.17 | 35.76 | |
| s | 7.59 | 1.52 | 1.17 | 2.96 | 2.27 | 2.14 | 1.98 | 1.69 | 1.29 | |
| min | 282.58 | 35.25 | 34.53 | 51.79 | 46.00 | 52.42 | 51.57 | 39.37 | 33.95 | |
| max | 301.08 | 40.35 | 37.85 | 59.46 | 51.09 | 58.65 | 57.01 | 43.95 | 37.15 | |
| Micoque | n | 2 | ||||||||
| m | 38.52 | |||||||||
| s | 3.71 | |||||||||
| min | 35.89 | |||||||||
| max | 41.14 | |||||||||
| Rameaux-Aval | n | 7 | 7 | 7 | 5 | 5 | 7 | 7 | 6 | 7 |
| m | 283.65 | 37.17 | 36.12 | 55.96 | 49.60 | 54.31 | 52.69 | 39.76 | 33.48 | |
| s | 5.96 | 3.04 | 2.74 | 1.08 | 2.49 | 1.64 | 2.00 | 1.18 | 1.07 | |
| min | 274.11 | 30.55 | 30.98 | 54.72 | 47.03 | 51.57 | 48.57 | 37.84 | 31.40 | |
| max | 290.80 | 39.80 | 38.56 | 57.36 | 53.71 | 55.82 | 54.99 | 41.32 | 34.90 | |
| Rameaux-Amont | n | 3 | 12 | 13 | 5 | 4 | 6 | 6 | 6 | 6 |
| m | 293.79 | 40.31 | 35.35 | 56.00 | 49.40 | 54.94 | 54.28 | 41.83 | 34.71 | |
| s | 13.38 | 1.83 | 4.71 | 2.14 | 2.70 | 2.88 | 1.23 | 2.07 | 1.29 | |
| min | 278.79 | 37.41 | 28.98 | 53.71 | 45.85 | 50.13 | 52.90 | 38.89 | 32.79 | |
| max | 304.50 | 43.13 | 44.71 | 58.41 | 52.38 | 58.38 | 55.86 | 43.87 | 36.07 | |
| Lunel-Viel | n | 44 | 51 | 50 | 42 | 41 | 47 | 48 | 45 | 48 |
| m | 277.63 | 41.00 | 37.73 | 55.59 | 49.45 | 54.37 | 53.79 | 42.12 | 33.75 | |
| s | 7.63 | 3.07 | 2.46 | 3.65 | 3.24 | 2.66 | 2.45 | 4.76 | 1.42 | |
| min | 262.61 | 36.24 | 33.61 | 40.47 | 38.32 | 50.30 | 48.43 | 38.84 | 30.89 | |
| max | 294.75 | 50.28 | 47.27 | 65.34 | 56.30 | 60.87 | 61.46 | 71.77 | 37.58 | |
| Suard | n | 3 | 2 | 6 | 6 | 2 | 2 | 2 | 2 | |
| m | 40.41 | 35.54 | 58.11 | 48.14 | 55.28 | 57.59 | 44.06 | 35.86 | ||
| s | 1.84 | 4.07 | 1.54 | 2.12 | 0.34 | 0.23 | 1.22 | 0.28 | ||
| min | 38.39 | 32.66 | 55.19 | 45.65 | 55.04 | 57.42 | 43.20 | 35.66 | ||
| max | 42.00 | 38.41 | 59.84 | 51.45 | 55.52 | 57.75 | 44.92 | 36.05 | ||
| Pech de l’Azé II | n | 1 | 1 | |||||||
| l. 9-6 | m | 56.84 | 52.37 | |||||||
| Rigabe | n | 2 | 3 | 3 | 1 | 2 | 3 | 3 | 2 | 2 |
| m | 284.88 | 39.05 | 35.53 | 56.22 | 47.58 | 53.14 | 50.82 | 43.89 | 35.61 | |
| s | 1.00 | 1.61 | 1.94 | 0.90 | 1.98 | 2.15 | 1.12 | 1.07 | ||
| min | 284.17 | 37.23 | 34.39 | 46.94 | 51.31 | 48.38 | 43.10 | 34.85 | ||
| max | 285.59 | 40.28 | 37.77 | 48.21 | 55.24 | 52.44 | 44.68 | 36.36 | ||
| Coudoulous II l.9 | n | 4 | 5 | 5 | 5 | 5 | 5 | 5 | 5 | 5 |
| m | 299.18 | 40.86 | 37.21 | 59.45 | 49.45 | 57.42 | 57.76 | 44.86 | 37.37 | |
| s | 9.06 | 0.63 | 1.33 | 3.90 | 1.03 | 2.56 | 3.11 | 1.84 | 1.83 | |
| min | 292.20 | 40.00 | 35.59 | 56.44 | 48.18 | 54.20 | 53.16 | 42.30 | 34.68 | |
| max | 311.80 | 41.62 | 39.30 | 65.44 | 50.90 | 60.31 | 60.42 | 46.70 | 39.34 | |
| Pech de l’Azé II | n | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| l.4 | m | 290.44 | 38.16 | 36.36 | 53.93 | 46.33 | 55.16 | 52.66 | 41.41 | 34.97 |
| s | 4.87 | 0.32 | 2.32 | 2.86 | 0.62 | 0.64 | 2.07 | 0.22 | 0.06 | |
| min | 286.99 | 37.93 | 34.72 | 51.90 | 45.89 | 54.70 | 51.19 | 41.25 | 34.93 | |
| max | 293.88 | 38.38 | 38.00 | 55.95 | 46.77 | 55.61 | 54.12 | 41.56 | 35.01 | |
| Peyre | n | 5 | 7 | 7 | 7 | 6 | 6 | 6 | 4 | 6 |
| m | 285.10 | 40.34 | 37.40 | 56.27 | 46.78 | 55.65 | 53.10 | 42.16 | 34.94 | |
| s | 9.90 | 0.72 | 1.51 | 1.30 | 1.77 | 2.05 | 0.92 | 1.01 | 1.63 | |
| min | 268.84 | 38.90 | 35.54 | 54.55 | 44.57 | 52.68 | 52.03 | 40.72 | 32.10 | |
| max | 292.30 | 40.93 | 38.91 | 58.75 | 49.12 | 58.74 | 54.59 | 42.93 | 36.48 |
Table 5.
Summary of the mesowear and microwear data for Pleistocene caballoid Equus from the South of France. Abbreviations: N = number of specimens; MWS = mesowear score; NP = average number of pits; NS = average number of scratches; LP% = percentage of specimens with large pits; G% = percentage of specimens with gouges; SWS = scratch width score; HC% = percentage of specimens with hypercoarse scratches; 0–17% = percentage of specimens with 17 or fewer scratches per counting area. M = average; SD = standard deviation. * Comm pers. F. Rivals. Camp-de-Peyre: E. m. campdepeyri; La Micoque l.E, H, J: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard l.53-50: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
Table 5.
Summary of the mesowear and microwear data for Pleistocene caballoid Equus from the South of France. Abbreviations: N = number of specimens; MWS = mesowear score; NP = average number of pits; NS = average number of scratches; LP% = percentage of specimens with large pits; G% = percentage of specimens with gouges; SWS = scratch width score; HC% = percentage of specimens with hypercoarse scratches; 0–17% = percentage of specimens with 17 or fewer scratches per counting area. M = average; SD = standard deviation. * Comm pers. F. Rivals. Camp-de-Peyre: E. m. campdepeyri; La Micoque l.E, H, J: E. mosbachensis; Rameaux-Aval and Amont: E. mosbachensis; Lunel-Viel: E. m. palustris; Payre l.F: E. mosbachensis; Suard l.53-50: E. caballus piveteaui; Pech de l’Azé II l.9-6: E. mosbachensis; Rigabe: E. m. palustris; Coudoulous II l.9: E. mosbachensis; Pech de l’Azé II l.4: E. caballus; Peyre: E. caballus.
| Site | Mesowear | Microwear | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | MWS | N | NP | NS | LP% | G% | SWS | HC% | 0–17% | ||
| Peyre | M | 27 | 3.33 | 14 | 25.71 | 15.21 | 50 | 28.57 | 0.92 | 7.14 | 85.71 |
| SD | 0.73 | 5.89 | 1.40 | ||||||||
| Pech de l’Azé II l.4-2 | M | 31 | 4.87 | 18 | 24.50 | 20.86 | 0 | 11.11 | 1 | 27.78 | 11.11 |
| SD | 0.72 | 5.94 | 3.67 | ||||||||
| Coudoulous II l.9 | M | 25 | 5.36 | 11 | 24.05 | 21.86 | 63.63 | 0 | 1 | 0 | 0 |
| SD | 0.76 | 5.07 | 3.17 | ||||||||
| Pech de l’Azé II l. 9-6 | M | 63 | 4.65 | 42 | 18.13 | 20.21 | 0 | 23.81 | 1 | 27.78 | 16.67 |
| SD | 0.9 | 4.25 | 2.63 | ||||||||
| Suard l. 50 | M | 12 | 5.17 | 7 | 19.57 | 17.93 | 57.14 | 14.28 | 1 | 0 | 42.86 |
| SD | 0.83 | 4.14 | 2.17 | ||||||||
| Suard l. 51 | M | 70 | 5.01 | 28 | 32.04 | 26.05 | 3.57 | 0 | 1 | 3.57 | 0 |
| SD | 0.84 | 5.66 | 3.42 | ||||||||
| Suard l. 52 | M | 30 | 4.80 | 26 | 29.83 | 22.35 | 3.84 | 3.84 | 0.92 | 0 | 0 |
| SD | 0.92 | 4.66 | 1.62 | ||||||||
| Suard l. 53 | M | 4 | 5.25 | 9 | 23.72 | 19.89 | 0 | 0 | 0.89 | 0 | 0 |
| SD | 0.96 | 3.91 | 1.34 | ||||||||
| Rigabe | M | 18 | 4.94 | 12 | 15.21 | 15.71 | 75 | 50 | 0.5 | 0 | 66.67 |
| SD | 0.73 | 3.44 | 3.95 | ||||||||
| Payre l. F * | M | 8 | 3.13 | 10 | 20.80 | 19.65 | 20 | 10 | 1.1 | 63.63 | 30 |
| SD | 1.13 | 2.84 | 3.64 | ||||||||
| Lunel-Viel | M | 65 | 4.88 | 36 | 15.85 | 19.61 | 31.43 | 8.33 | 1 | 2.78 | 22.22 |
| SD | 0.93 | 4.17 | 2.65 | ||||||||
| Rameaux-Amont | M | 151 | 4.37 | 24 | 18.88 | 13.77 | 37.5 | 12.5 | 1.04 | 4.17 | 87.5 |
| SD | 0.97 | 7.19 | 2.79 | ||||||||
| Rameaux-Aval | M | 8 | 3.50 | 3 | 16.83 | 13.67 | 66.66 | 33.33 | 1 | 0 | 100 |
| SD | 1.07 | 2.84 | 2.02 | ||||||||
| La Micoque l. J | M | 21 | 4.48 | 19 | 21.89 | 21.13 | 94.74 | 42.10 | 1.05 | 10.53 | 10.53 |
| SD | 0.93 | 4.82 | 2.99 | ||||||||
| La Micoque l. H | M | 8 | 3.88 | 15 | 22.40 | 23.33 | 46.66 | 0 | 1 | 0 | 0 |
| SD | 0.99 | 3.52 | 3.19 | ||||||||
| La Micoque l. E | M | 5 | 5 | 22 | 18.34 | 24.55 | 0 | 0 | 0.72 | 0 | 4.55 |
| SD | 0 | 5.41 | 4.27 | ||||||||
| Camp-de-Peyre | M | 38 | 4.18 | 7 | 28.57 | 22.00 | 0 | 0 | 0.71 | 14.28 | 0 |
| SD | 0.51 | 4.17 | 2.47 | ||||||||
Table 6.
Comparison of the mean (m) body mass with the standard deviation (s) in kg of the cohorts (n = number) of caballoid horses from the western and eastern part of France and from cold and temperate isotopic stages (IS) using the Mann-Withney statistical test.
Table 6.
Comparison of the mean (m) body mass with the standard deviation (s) in kg of the cohorts (n = number) of caballoid horses from the western and eastern part of France and from cold and temperate isotopic stages (IS) using the Mann-Withney statistical test.
| Group | n | m | s | p-Values |
|---|---|---|---|---|
| Western cohorts | 9 | 504.91 | 35.67 | 0.8636 |
| Eastern cohorts | 3 | 492.88 | 17.89 | |
| Cold IS | 6 | 502.22 | 37.91 | 0.9372 |
| Temperate IS | 6 | 501.59 | 28.13 |
Table 7.
Comparison of the mean (m) body mass with the standard deviation (s) in kg of the cohorts (n = number) of caballoid horses according to their diet using a Kruskal-Wallis statistical test. G-G: grazer according to meso- and microwear; G-MF: grazer according to mesowear analysis and mixed-feeder at the time of death (microwear analysis); G-B: grazer according to mesowear analysis and Browser at the time of death (microwear analysis).
Table 7.
Comparison of the mean (m) body mass with the standard deviation (s) in kg of the cohorts (n = number) of caballoid horses according to their diet using a Kruskal-Wallis statistical test. G-G: grazer according to meso- and microwear; G-MF: grazer according to mesowear analysis and mixed-feeder at the time of death (microwear analysis); G-B: grazer according to mesowear analysis and Browser at the time of death (microwear analysis).
| Group | n | m | s |
|---|---|---|---|
| G-G | 6 | 502.64 | 37.57 |
| G-MF | 3 | 492.88 | 17.89 |
| G-B | 3 | 509.45 | 38.91 |
| Kruskal-Wallis Test p-Values | |||
| G-G/G-MF | G-G/G-B | G-MF/G-B | |
| −0.3651 | 0.3651 | −0.3086 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Uzunidis, A. Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments. Quaternary 2021, 4, 23. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4030023
AMA Style
Uzunidis A. Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments. Quaternary. 2021; 4(3):23. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4030023
Chicago/Turabian StyleUzunidis, Antigone. 2021. "Middle Pleistocene Variations in the Diet of Equus in the South of France and Its Morphometric Adaptations to Local Environments" Quaternary 4, no. 3: 23. https://0-doi-org.brum.beds.ac.uk/10.3390/quat4030023