
Comprehensive Identification of Reliable Reference Genes for qRT-PCR Normalization of Fusarium oxysporum-Resistant Genes’ Expressions in Lilium sargentiae Wilson

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Fungal Materials
2.2. Cultivation and Inoculation
2.3. Selection of Candidate Reference Genes
2.4. RNA Extraction and cDNA Synthesis
2.5. qRT-PCR Analysis
2.6. Expression Stability Analysis and Validation of Reference Genes
3. Results
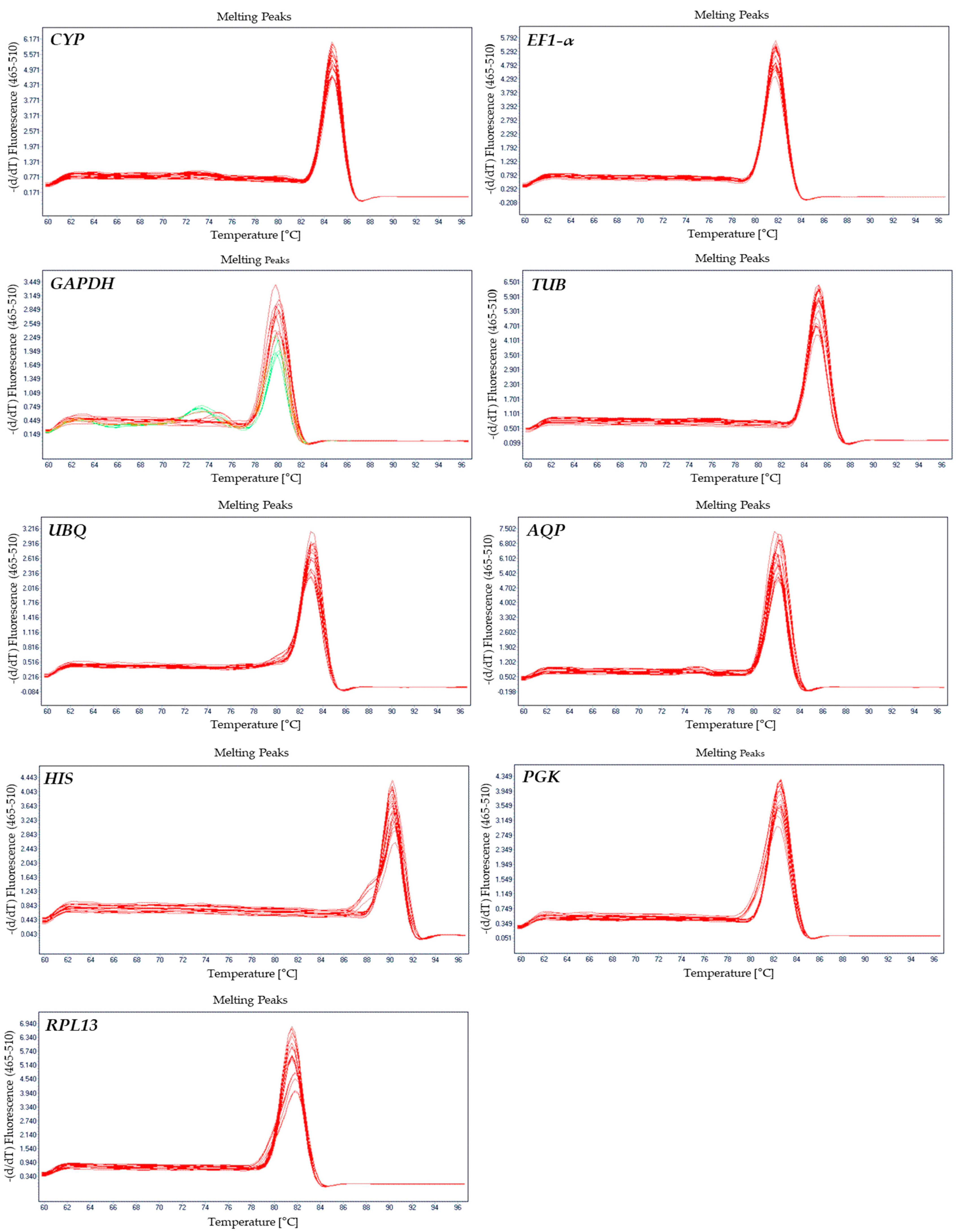
3.1. Verification of Primer Specificity and PCR Amplicons
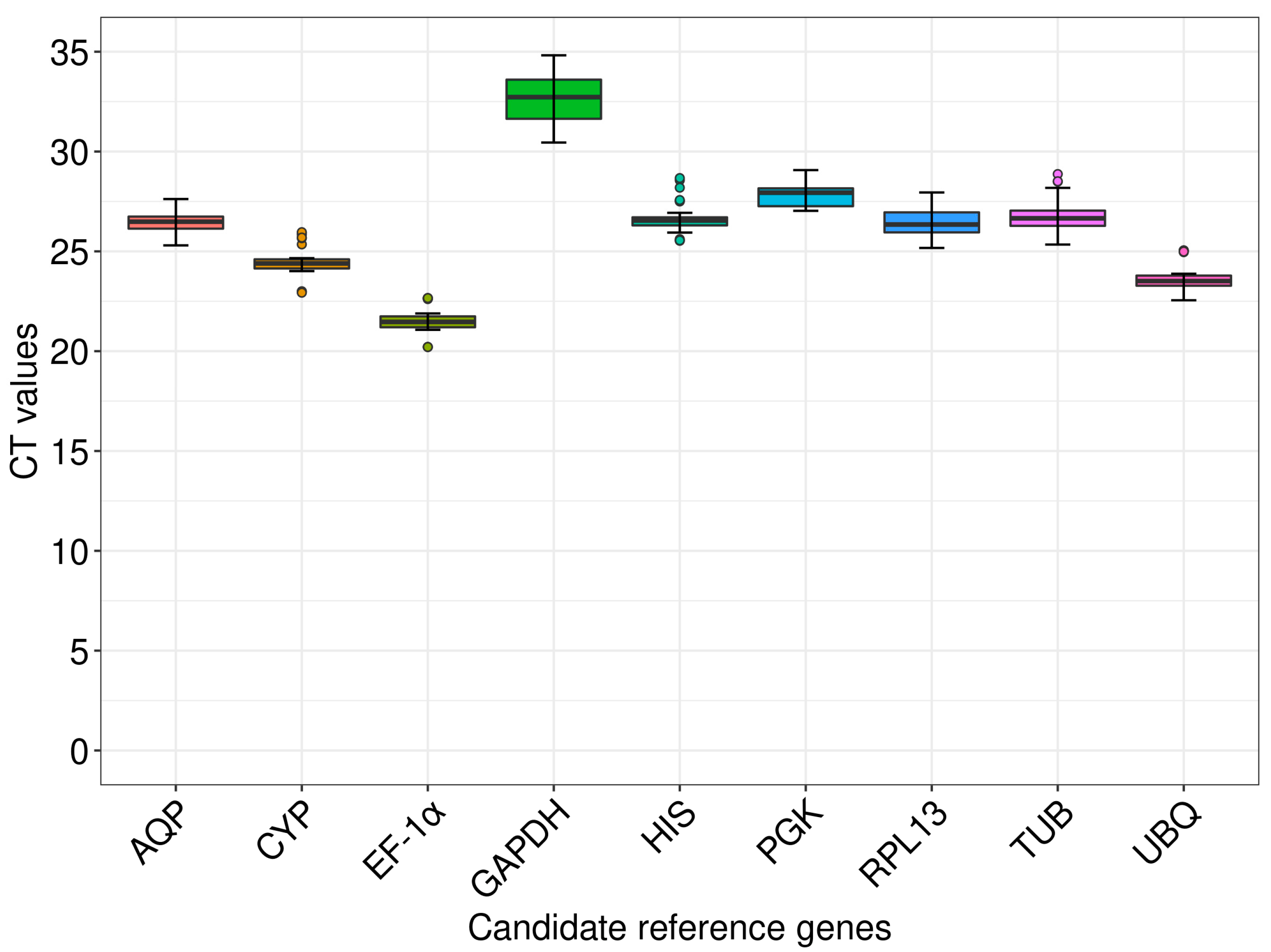
3.2. Expression Profile Analysis of the Candidate Reference Genes
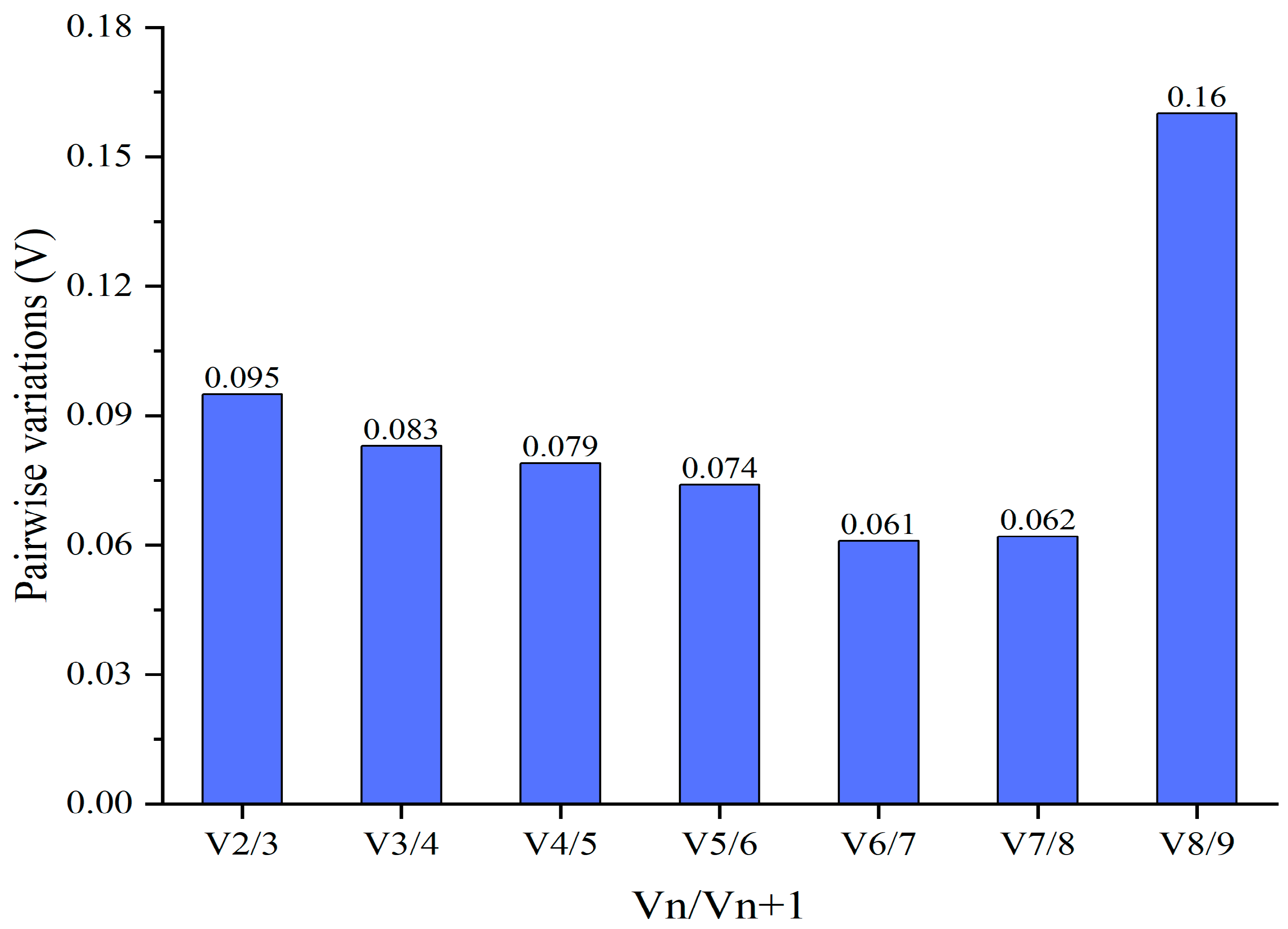
3.3. Expression Stability Analysis of the Candidate Reference Genes
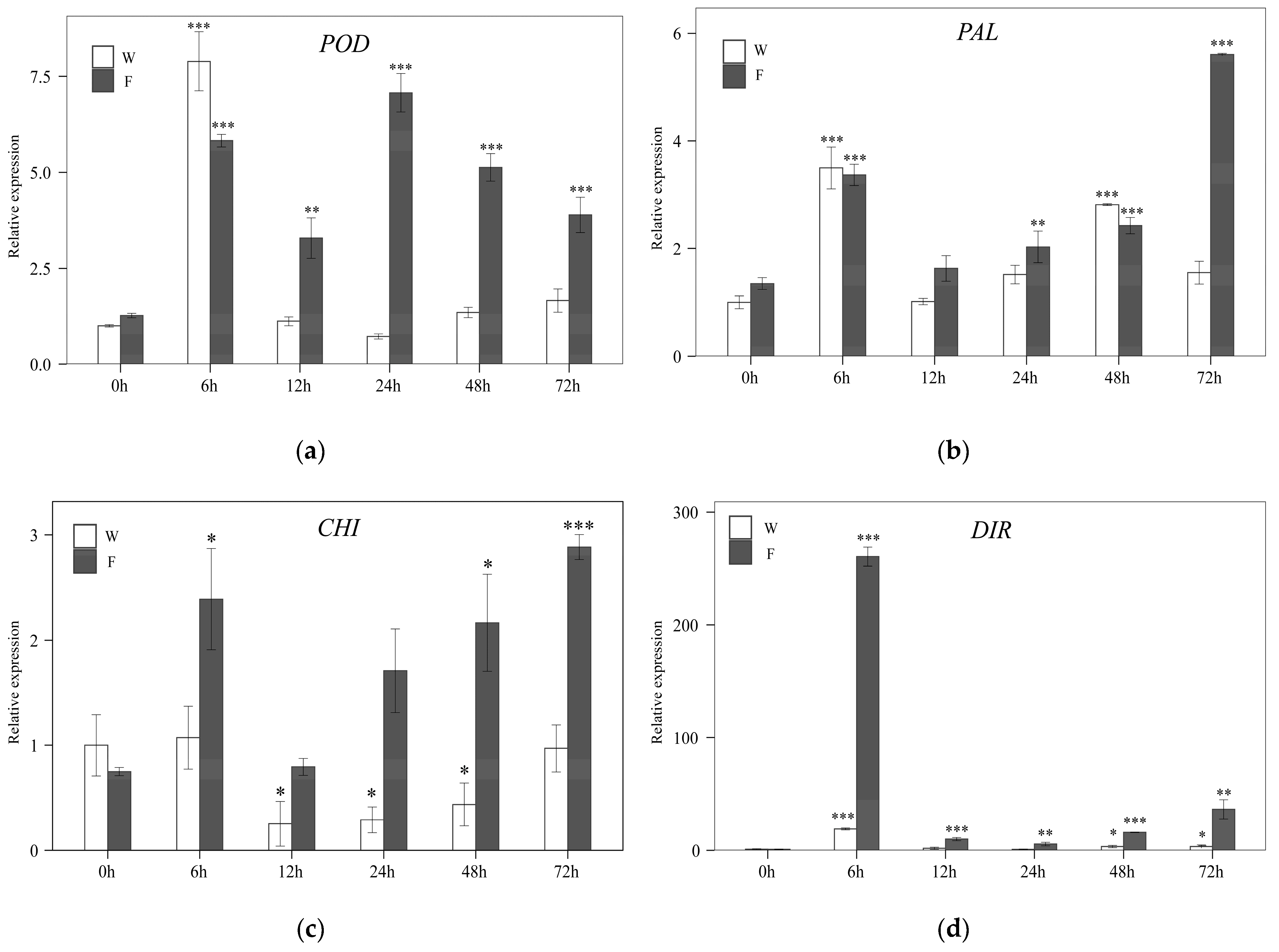
3.4. Expression Stability Validation of the Most Reliable Reference Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1–0034.11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Xu, H.; Cao, Y.; Yang, P.; Feng, Y.; Tang, Y.; Yuan, S.; Ming, J. Validation of Reference Genes for Quantitative Real-Time PCR during Bicolor Tepal Development in Asiatic Hybrid Lilies (Lilium spp.). Front. Plant Sci. 2017, 8, 669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Wu, J.; Hua, Q.; Tel-Zur, N.; Xie, F.; Zhang, Z.; Chen, J.; Zhang, R.; Hu, G.; Zhao, J.; et al. Identification of reliable reference genes for quantitative real-time PCR normalization in pitaya. Plant Methods 2019, 15, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, T.K.; Rim, Y.; Jang, H.-J.; Kim, C.H.; Park, J.; Kumar, R.; Lee, S.; Kim, B.C.; Bhak, J.; Nguyen-Quoc, B.; et al. De novo transcriptome sequencing of Momordica cochinchinensis to identify genes involved in the carotenoid biosynthesis. Plant Mol. Biol. 2012, 79, 413–427. [Google Scholar] [CrossRef]
- Chen, L.; Li, L.; Dai, Y.; Wang, X.; Duan, Y.; Yang, G. De novo transcriptome analysis of Osmanthus serrulatus Rehd. flowers and leaves by Illumina sequencing. Biochem. Syst. Ecol. 2015, 61, 531–540. [Google Scholar] [CrossRef]
- Cao, A.; Shao, D.; Cui, B.; Tong, X.; Zheng, Y.; Sun, J.; Li, H. Screening the Reference Genes for Quantitative Gene Expression by RT-qPCR During SE Initial Dedifferentiation in Four Gossypium hirsutum Cultivars that Have Different SE Capability. Genes 2019, 10, 497. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Zhu, Y.; Zhao, Y.; Wang, Y.; Li, J.; Wang, Q.; Liu, Y. De novo transcriptome sequencing and analysis of salt-, alkali-, and drought-responsive genes in Sophora alopecuroides. BMC Genom. 2020, 21, 423. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, J.; Liu, Y.; Gu, Y.; Zhang, H.; Ahmad, F.; Wang, G.; Ren, L. Selection and Validation of Reliable Reference Genes for qRT-PCR Normalization of Bursaphelenchus xylophilus from Different Temperature Conditions and Developmental Stages. Appl. Sci. 2022, 12, 2880. [Google Scholar] [CrossRef]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalisation; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Qi, X.; Yan, H.; Huang, L.; Nie, G.; Zhang, X. Reference Gene Selection for Quantitative Real-Time Reverse-Transcriptase PCR in Annual Ryegrass (Lolium multiflorum) Subjected to Various Abiotic Stresses. Molecules 2018, 23, 172. [Google Scholar] [CrossRef]
- Kou, X.; Zhang, L.; Yang, S.; Li, G.; Ye, J. Selection and validation of reference genes for quantitative RT-PCR analysis in peach fruit under different experimental conditions. Sci. Hortic. 2017, 225, 195–203. [Google Scholar] [CrossRef]
- Nolan, T.; Hands, R.E.; Bustin, S.A. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef]
- Quackenbush, J. Microarray data normalization and transformation. Nat. Genet. 2002, 32, 496–501. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, C.-S.; Huo, Y.-B.; Zhang, B.; Ma, Z.-Q.; Feng, J.-T.; Zhang, X. Validation of suitable reference genes for quantitative gene expression analysis in Tripterygium wilfordii. Mol. Biol. Rep. 2019, 46, 4161–4174. [Google Scholar] [CrossRef]
- Liang, L.; He, Z.; Yu, H.; Wang, E.; Zhang, X.; Zhang, B.; Zhang, C.; Liang, Z. Selection and Validation of Reference Genes for Gene Expression Studies in Codonopsis pilosula Based on Transcriptome Sequence Data. Sci. Rep. 2020, 10, 1362. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.; Zhao, Z.; Qian, C.; Sui, Y.; Malik, A.A.; Chen, J. Selection of appropriate reference genes for gene expression studies by quantitative real-time polymerase chain reaction in cucumber. Anal. Biochem. 2010, 399, 257–261. [Google Scholar] [CrossRef]
- Volkov, R.A.; Panchuk, I.I.; Schöffl, F. Heat-stress-dependency and developmental modulation of gene expression: The potential of house-keeping genes as internal standards in mRNA expression profiling using real-time RT-PCR. J. Exp. Bot. 2003, 54, 2343–2349. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.-Y.; Seo, P.J.; Yang, M.-S.; Xiang, F.; Park, C.-M. Exploring valid reference genes for gene expression studies in Brachypodium distachyon by real-time PCR. BMC Plant Biol. 2008, 8, 112. [Google Scholar] [CrossRef] [Green Version]
- Yi, S.; Qian, Y.; Han, L.; Sun, Z.; Fan, C.; Liu, J.; Ju, G. Selection of reliable reference genes for gene expression studies in Rhododendron micranthum Turcz. Sci. Hortic. 2012, 138, 128–133. [Google Scholar] [CrossRef]
- Li, J.; Huang, H.; Shan, T.; Pang, S. Selection of reference genes for real-time RT-PCR normalization in brown alga Undaria pinnatifida. J. Appl. Phycol. 2019, 31, 787–793. [Google Scholar] [CrossRef]
- Dheda, K.; Huggett, J.F.; Chang, J.S.; Kim, L.U.; Bustin, S.A.; Johnson, M.A.; Rook, G.A.W.; Zumla, A. The implications of using an inappropriate reference gene for real-time reverse transcription PCR data normalization. Anal. Biochem. 2005, 344, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yuan, F.; Long, G.; Qin, L.; Deng, Z. Selection of reference genes for quantitative real-time RT-PCR analysis in citrus. Mol. Biol. Rep. 2012, 39, 1831–1838. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Na, J.U.; Kim, J.S.; Lee, J.E.; Nie, H.; Lee, K.A.; Woo, S.Y.; Kim, S.H. Identification of valid reference genes for quantitative RT-PCR in Caragana microphylla under salt and drought stresses. Physiol. Mol. Biol. Plants 2020, 26, 2103–2108. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Rhee, H.K.; Kim, Y.J.; Lim, K.B.; van Tuyl, J.M. Resistance to Fusarium oxysporum f. sp. lilii in Lilium. Acta Hortic. 2003, 620, 311–318. [Google Scholar] [CrossRef]
- Rao, J.; Liu, D.; Zhang, N.; He, H.; Ge, F.; Chen, C. Identification of Genes Differentially Expressed in a Resistant Reaction to Fusarium Oxysporum in Lilium Regale by SSH. IERI Procedia 2013, 5, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.P.; Jiang, S.; Qu, S.P.; Yang, X.M.; Wang, X.N.; Ma, L.L.; Wu, L.L.; He, Y.Q.; Wang, J.H. In Vitro Selection for Fusarium Resistant Oriental Lily Mutants Using Culture Filtrate of the Fungal Agent. Acta Hortic. 2014, 1027, 205–212. [Google Scholar] [CrossRef]
- Yang, X.M.; Wang, J.H.; Wang, L.H.; Zhang, Y.P.; Zhang, L.F.; Qu, S.P. Resistance Identification and Genetic Diversity Among Lily Varieties Based on Resistance Gene Analogue. Acta Hortic. Sin. 2012, 39, 2404–2412. [Google Scholar]
- He, H.; Liu, D.; Zhang, N.; Zheng, W.; Han, Q.; Ji, B.; Ge, F.; Chen, C. The PR10 gene family is highly expressed in Lilium regale Wilson during Fusarium oxysporum f. sp. lilii infection. Genes Genom 2014, 36, 497–507. [Google Scholar] [CrossRef]
- Li, S.; Hai, J.; Wang, Z.; Deng, J.; Liang, T.; Su, L.; Liu, D. Lilium regale Wilson WRKY2 Regulates Chitinase Gene Expression During the Response to the Root Rot Pathogen Fusarium oxysporum. Front. Plant Sci. 2021, 12, 741463. [Google Scholar] [CrossRef]
- He, X.; Li, W.; Zhang, W.; Jin, X.; Shenkute, A.G.; Aynalem, T.; Xu, S.; Wang, W. Transcriptome Sequencing Analysis Provides Insights Into the Response to Fusarium oxysporum in Lilium pumilum. Evol. Bioinform. Online 2019, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-Q.; Sun, H. Pollination in Lilium sargentiae (Liliaceae) and the first confirmation of long-tongued hawkmoths as a pollinator niche in Asia. J. Syst. Evol. 2019, 57, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Van Huylenbroeck, J. Ornamental Crops; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-90697-3. [Google Scholar]
- Straathof, T.P.; Tuyl, J.M.V. Genetic variation in resistance to Fusarium oxysporum f.sp. lilii in the genus Lilium. Ann. Appl. Biol. 1994, 125, 61–72. [Google Scholar] [CrossRef]
- Yang, X.M.; Wang, J.H.; Wang, L.H.; Wu, X.W.; Peng, L.C.; Qu, S.P. Identification and ITS Sequence Analysis of Pathogen of Lily Wilt. Southwest China J. Agric. Sci. 2010, 23, 1914–1916. [Google Scholar] [CrossRef]
- Imai, T.; Ubi, B.E.; Saito, T.; Moriguchi, T. Evaluation of reference genes for accurate normalization of gene expression for real time-quantitative PCR in Pyrus pyrifolia using different tissue samples and seasonal conditions. PLoS ONE 2014, 9, e86492. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper--Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Li, F.; Cheng, Y.; Ma, L.; Li, S.; Wang, J. Identification of reference genes provides functional insights into meiotic recombination suppressors in Gerbera hybrida. Hortic. Plant J. 2022, 8, 123–132. [Google Scholar] [CrossRef]
- Tang, Q.-Y.; Zhang, C.-X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chung, W.-C.; Wu, R.-S.; Hsu, C.-P.; Huang, H.-C.; Huang, J.-W. Application of antagonistic rhizobacteria for control of Fusarium seedling blight and basal rot of lily. Australas. Plant Pathol. 2011, 40, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Liu, D.; Zheng, W.; He, H.; Ji, B.; Han, Q.; Ge, F.; Chen, C. A bZIP transcription factor, LrbZIP1, is involved in Lilium regale Wilson defense responses against Fusarium oxysporum f. sp. lilii. Genes Genom. 2014, 36, 789–798. [Google Scholar] [CrossRef]
- Zhang, N.; Guan, R.; Yang, Y.; Bai, Z.; Ge, F.; Liu, D. Isolation and characterization of a Fusarium oxysporum-resistant gene LrGLP1 from Lilium regale Wilson. Vitr. Cell. Dev. Biol.-Plant 2017, 53, 461–468. [Google Scholar] [CrossRef]
- Rao, J.; Liu, D.; Zhang, N.; He, H.; Ge, F.; Chen, C. Differential gene expression in incompatible interaction between Lilium regale Wilson and Fusarium oxysporum f. sp. lilii revealed by combined SSH and microarray analysis. Mol. Biol. 2014, 48, 802–812. [Google Scholar] [CrossRef]
- Luo, H.L.; Luo, L.P.; Guan, B.C.; Li, E.X.; Xiong, D.J.; Sun, B.T.; Peng, K.; Yang, B.Y. Evaluation of candidate reference genes for RT-qPCR in lily (Lilium brownii). J. Hortic. Sci. Biotechnol. 2014, 89, 345–351. [Google Scholar] [CrossRef]
- Li, X.; Cheng, J.; Zhang, J.; Da Teixeira Silva, J.A.; Wang, C.; Sun, H. Validation of Reference Genes for Accurate Normalization of Gene Expression in Lilium davidii var. unicolor for Real Time Quantitative PCR. PLoS ONE 2015, 10, e0141323. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.L.; Zhang, Y.P.; Ding, K.; Wu, L.F.; Wang, X.N.; Cui, G.F.; Jia, W.J.; Duan, Q.; Wang, J.H. Resistance Identification of lily Germplasms to Fusarium oxysporum and Screening of the Resistance Identification of lily Germplasms to Fusarium oxysporum and Screening of the Resistance Related Genes. Acta Hortic. Sin. 2012, 39, 1141–1150. [Google Scholar]
- Seneviratne, H.K.; Dalisay, D.S.; Kim, K.-W.; Moinuddin, S.G.A.; Yang, H.; Hartshorn, C.M.; Davin, L.B.; Lewis, N.G. Non-host disease resistance response in pea (Pisum sativum) pods: Biochemical function of DRR206 and phytoalexin pathway localization. Phytochemistry 2015, 113, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.F.; Ferreira, R.B.; Monteiro, S. Transcriptomic changes following the compatible interaction Vitis vinifera-Erysiphe necator. Paving the way towards an enantioselective role in plant defence modulation. Plant Physiol. Biochem. 2013, 68, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Reboledo, G.; Del Campo, R.; Alvarez, A.; Montesano, M.; Mara, H.; Ponce de León, I. Physcomitrella patens Activates Defense Responses against the Pathogen Colletotrichum gloeosporioides. Int. J. Mol. Sci. 2015, 16, 22280–22298. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Sun, W.; Zhou, P.; Wei, H.; Wang, P.; Li, H.; Rehman, S.; Li, D.; Zhuge, Q. Genome-Wide Characterization of Dirigent Proteins in Populus: Gene Expression Variation and Expression Pattern in Response to Marssonina brunnea and Phytohormones. Forests 2021, 12, 507. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) | Product Length (bp) |
|---|---|---|
| CYP | F:CGGTGGCGAGTCTATCTAT; R:TGGTATTAGGCCCAGCAT | 104 |
| EF1-α | F:TATTGACAAGCGTGTGATCG; R:TTATCAAGCACCCACGCA | 87 |
| GAPDH | F:ACATGGCTGCAACCAAGTA; R:CATGAAGCCGGCATTTATT | 80 |
| TUB | F:GCTGTTTCATCCCGAGCA; R:CAGGCACAGGTCAACAAT | 103 |
| UBQ | F:CACCCTTGCGGACTATAAC; R:TTCCGGTGAGCGTCTTGA | 98 |
| AQP | F:CGTTTCCTCTCTATCTCCAG;R:CCATGAGCAGATCCATGTATT | 117 |
| HIS | F:GATTCACAGGCAGTTGAAGT; R:TTCAGATCCTTGCTCGCAT | 138 |
| PGK | F:GGCTAAAGGCGTACGACTA; R:GCAGGTACAACCTTGCTC | 87 |
| RPL13 | F:GCAGGTACAACCTTGCTC; R:CCTTGCTCCTCTTGATCTTTA | 93 |
| POD | F:CCGAGCCTCTTGAGTACAATTA; R:CCATTGACAAAGCAGTCGT | 110 |
| PAL | F:CTGATCAAGGTCATAGACAGG; R:TAGTGCGTGCTCCACCAATA | 111 |
| CHI | F:GCCGACAATAAGATGGCG; R:AGGTTAGCTCCATAGCTGAC | 80 |
| DIR | F:GAGCGGCCTATTCAGGCTA; R:CGTACTCGACGAGAGATG | 89 |
| Stability Rank | geNorm | NormFinder | BestKeeper | |||
|---|---|---|---|---|---|---|
| Reference Genes | M | Reference Genes | SV | Reference Genes | SD | |
| 1 | HIS | 0.533 | HIS | 0.086 | CYP | 0.126 |
| 2 | EF-1α | 0.535 | PGK | 0.174 | AQP | 0.134 |
| 3 | CYP | 0.543 | CYP | 0.175 | EF-1α | 0.135 |
| 4 | UBQ | 0.598 | EF-1α | 0.182 | HIS | 0.146 |
| 5 | PGK | 0.611 | AQP | 0.211 | UBQ | 0.153 |
| 6 | TUB | 0.615 | TUB | 0.233 | TUB | 0.166 |
| 7 | AQP | 0.620 | UBQ | 0.280 | RPL13 | 0.180 |
| 8 | RPL13 | 0.717 | RPL13 | 0.420 | PGK | 0.204 |
| 9 | GAPDH | 1.466 | GAPDH | 0.994 | GAPDH | 0.217 |
| Comprehensive Ranking | Reference Genes | GM Values of Ranking |
|---|---|---|
| 1 | HIS | 1.587 |
| 2 | CYP | 2.080 |
| 3 | PGK | 2.884 |
| 4 | UBQ | 4.121 |
| 5 | AQP | 4.309 |
| 6 | EF-1α | 5.192 |
| 7 | TUB | 6.000 |
| 8 | RPL13 | 7.652 |
| 9 | GAPDH | 9.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Li, X.; Duan, Q.; Jia, W.; Du, W.; Wang, X.; Cui, G.; Zhang, Y.; Wang, J. Comprehensive Identification of Reliable Reference Genes for qRT-PCR Normalization of Fusarium oxysporum-Resistant Genes’ Expressions in Lilium sargentiae Wilson. Appl. Sci. 2023, 13, 3395. https://0-doi-org.brum.beds.ac.uk/10.3390/app13063395
Ma L, Li X, Duan Q, Jia W, Du W, Wang X, Cui G, Zhang Y, Wang J. Comprehensive Identification of Reliable Reference Genes for qRT-PCR Normalization of Fusarium oxysporum-Resistant Genes’ Expressions in Lilium sargentiae Wilson. Applied Sciences. 2023; 13(6):3395. https://0-doi-org.brum.beds.ac.uk/10.3390/app13063395
Chicago/Turabian StyleMa, Lulin, Xiang Li, Qing Duan, Wenjie Jia, Wenwen Du, Xiangning Wang, Guangfen Cui, Yiping Zhang, and Jihua Wang. 2023. "Comprehensive Identification of Reliable Reference Genes for qRT-PCR Normalization of Fusarium oxysporum-Resistant Genes’ Expressions in Lilium sargentiae Wilson" Applied Sciences 13, no. 6: 3395. https://0-doi-org.brum.beds.ac.uk/10.3390/app13063395