
Suppressive Effects of Rosmarinic Acid Rich Fraction from Perilla on Oxidative Stress, Inflammation and Metastasis Ability in A549 Cells Exposed to PM via C-Jun, P-65-Nf-Κb and Akt Signaling Pathways

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. PMFF Sampling and GC-MS Analysis
2.2. Rosmarinic Acid Rich Fraction (RA-RF) Preparation
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. Cell Apoptosis Assay
2.6. Reactive Oxygen Species (ROS) Measurement
2.7. Total RNA Extraction and cDNA Synthesis
2.8. Quantitative Real-Time PCR (qPCR)
2.9. Cell Invasion and Migration Assay
2.10. Gelatin Zymography
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
3.1. The PAHs Concentrations in PMFF

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | PAHs Concentration (ng/m3) | Mol. wt. (g mol−1) [32] | Class [33] |
|---|---|---|---|
| Low molecular weight | |||
| Naphthalene | 0.082 ± 0.003 | 128 | - |
| Acenaphthylene | 0.083 ± 0.000 | 152 | - |
| Acenaphthene | 0.072 ± 0.004 | 154 | - |
| Phenanthrene | 0.208 ± 0.008 | 178 | - |
| Anthracene | 0.196 ± 0.011 | 178 | - |
| Fluorene | ND | 166 | - |
| High molecular weight | |||
| Benzo[ghi]perylene | 2.094 ± 0.016 | 276 | 3 |
| Indeno[1,2,3-cd]pyrene | 1.803 ± 0.036 | 276 | 2B |
| Benzo[b]fluoranthene | 1.531 ± 0.017 | 252 | 2B |
| Benzo[a]pyrene | 0.764 ± 0.017 | 252 | 1 |
| Benzo[k]fluoranthene | 0.609 ± 0.031 | 252 | 2B |
| Dibenz[a,h]anthracene | 0.509 ± 0.006 | 278 | 2A |
| Fluoranthene | 0.487 ± 0.016 | 202 | 3 |
| Pyrene | 0.396 ± 0.004 | 202 | 3 |
| Chrysene | 0.282 ± 0.001 | 228 | 2B |
| Benzo[a]anthracene | 0.184 ± 0.003 | 228 | 2B |
3.2. Quantification of Rosmarinic Acid Rich Fraction (RA-RF)
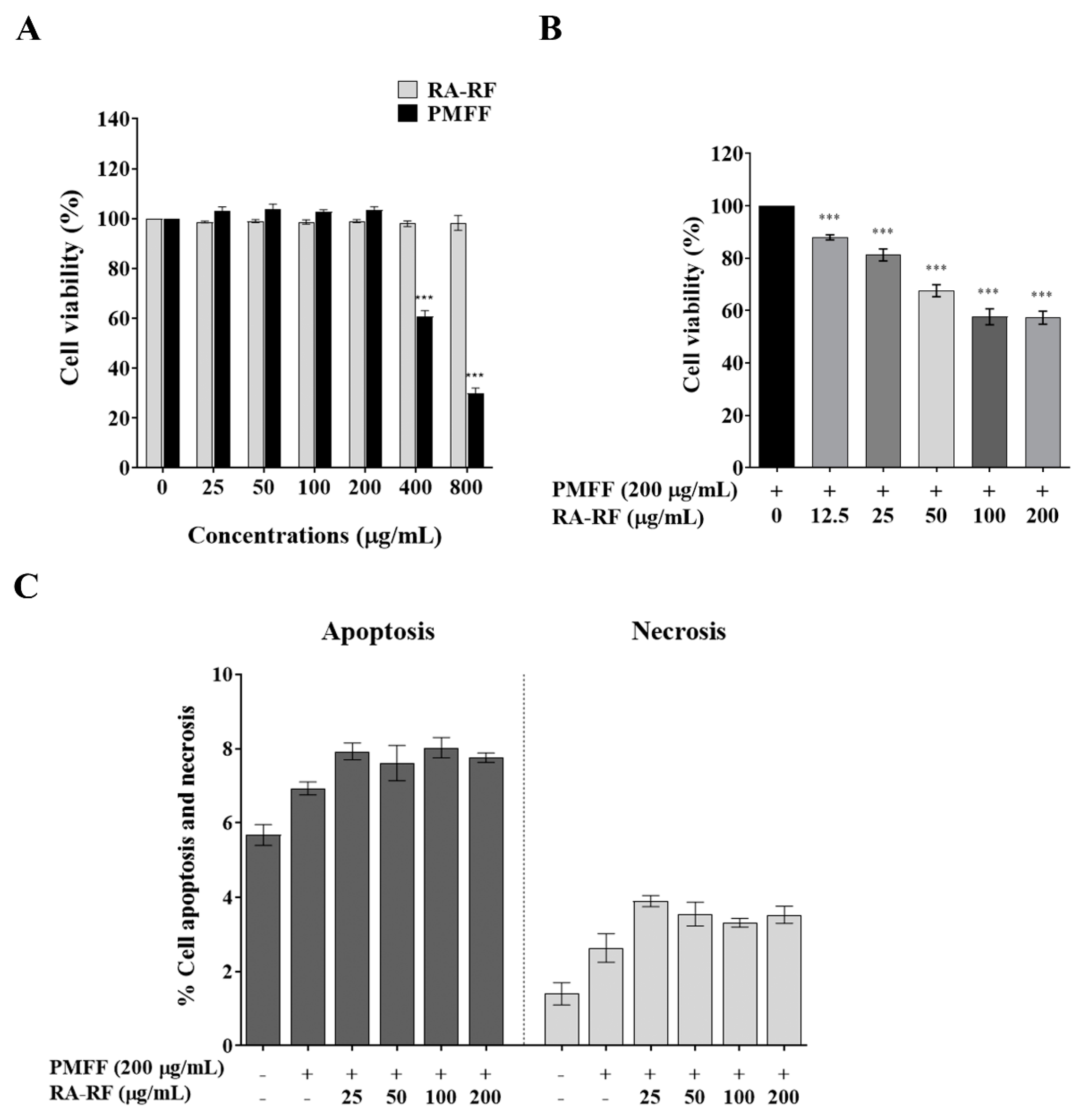
3.3. RA-RF Enhances Cell Viability and Apoptosis of PMFF-Induced A549 Cells
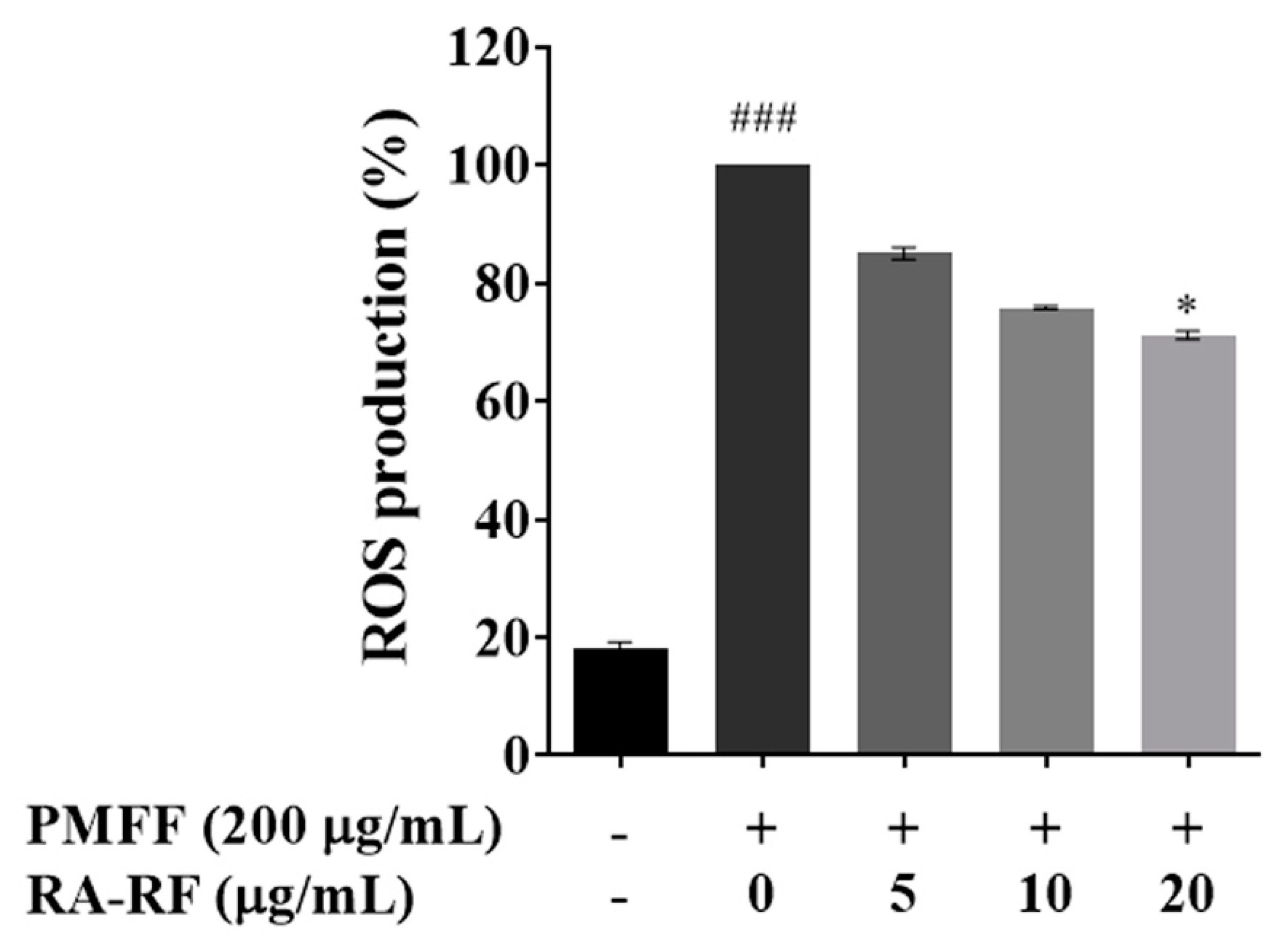
3.4. RA-RF Inhibits the Intracellular ROS Production
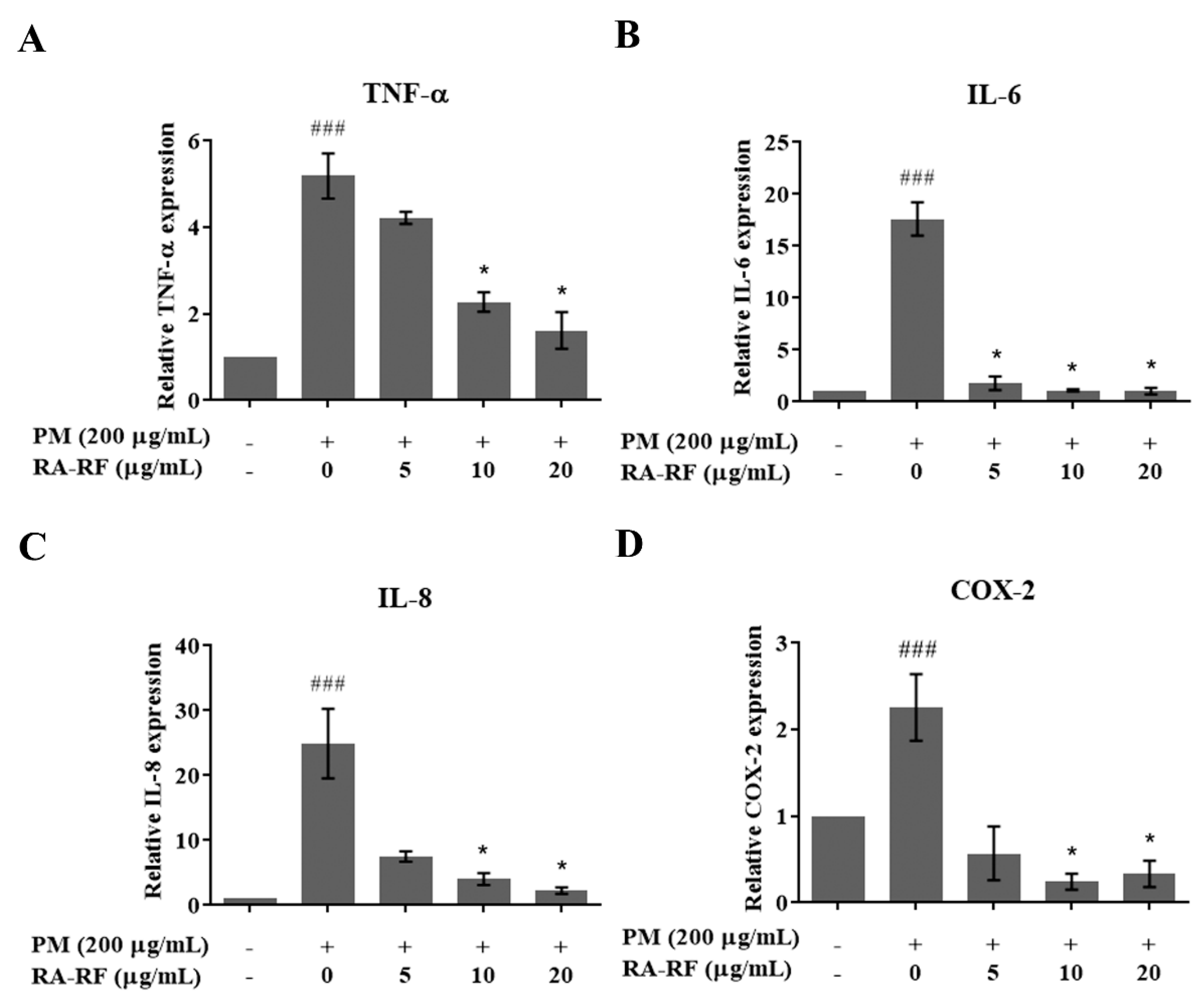
3.5. RA-RF Reducing the Inflammatory Cytokines and COX-2 Transcript
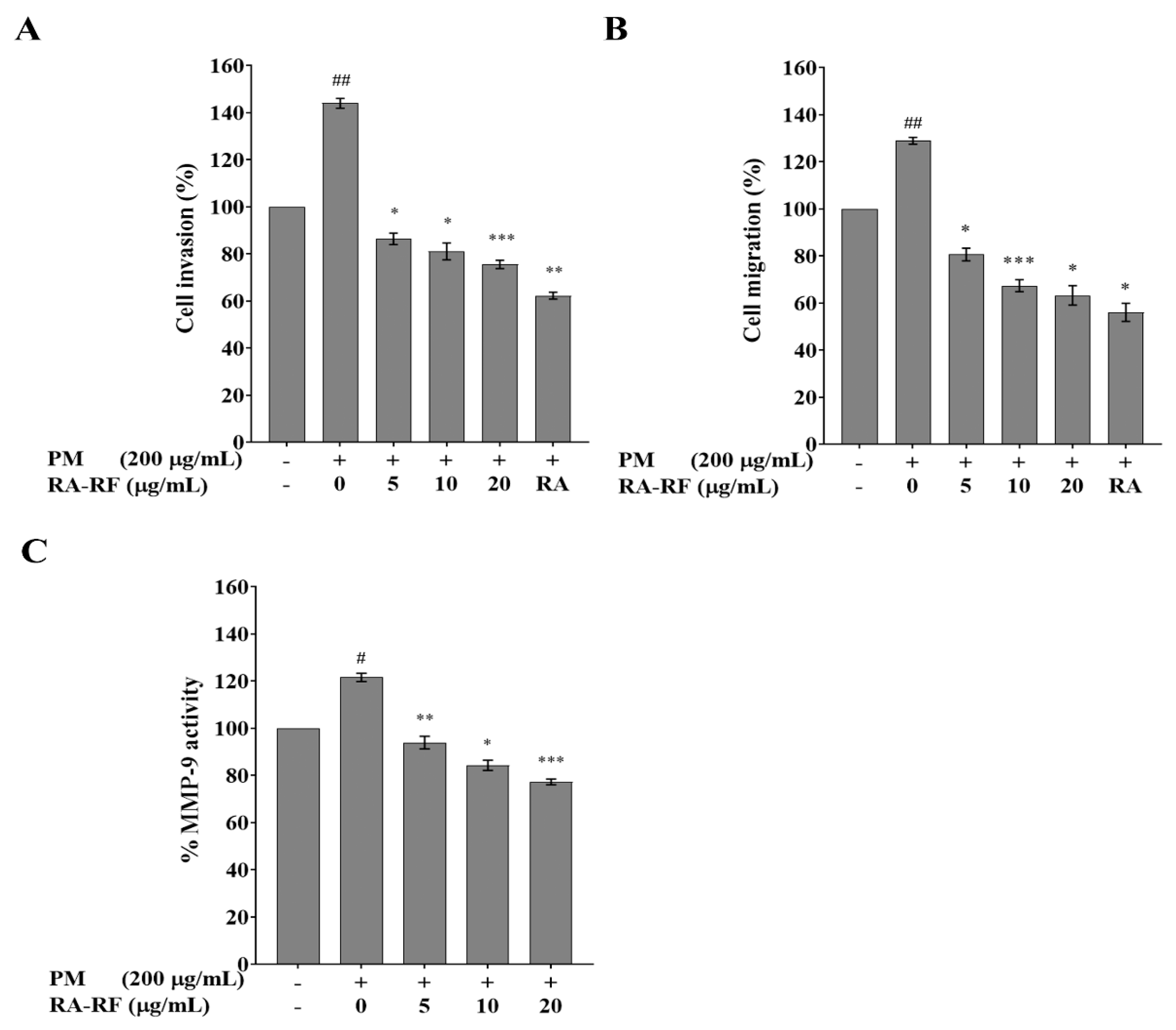
3.6. RA-RF Inhibits Invasion, Migration and MMP-9 Activity of A549 Cells Induced by PMFF
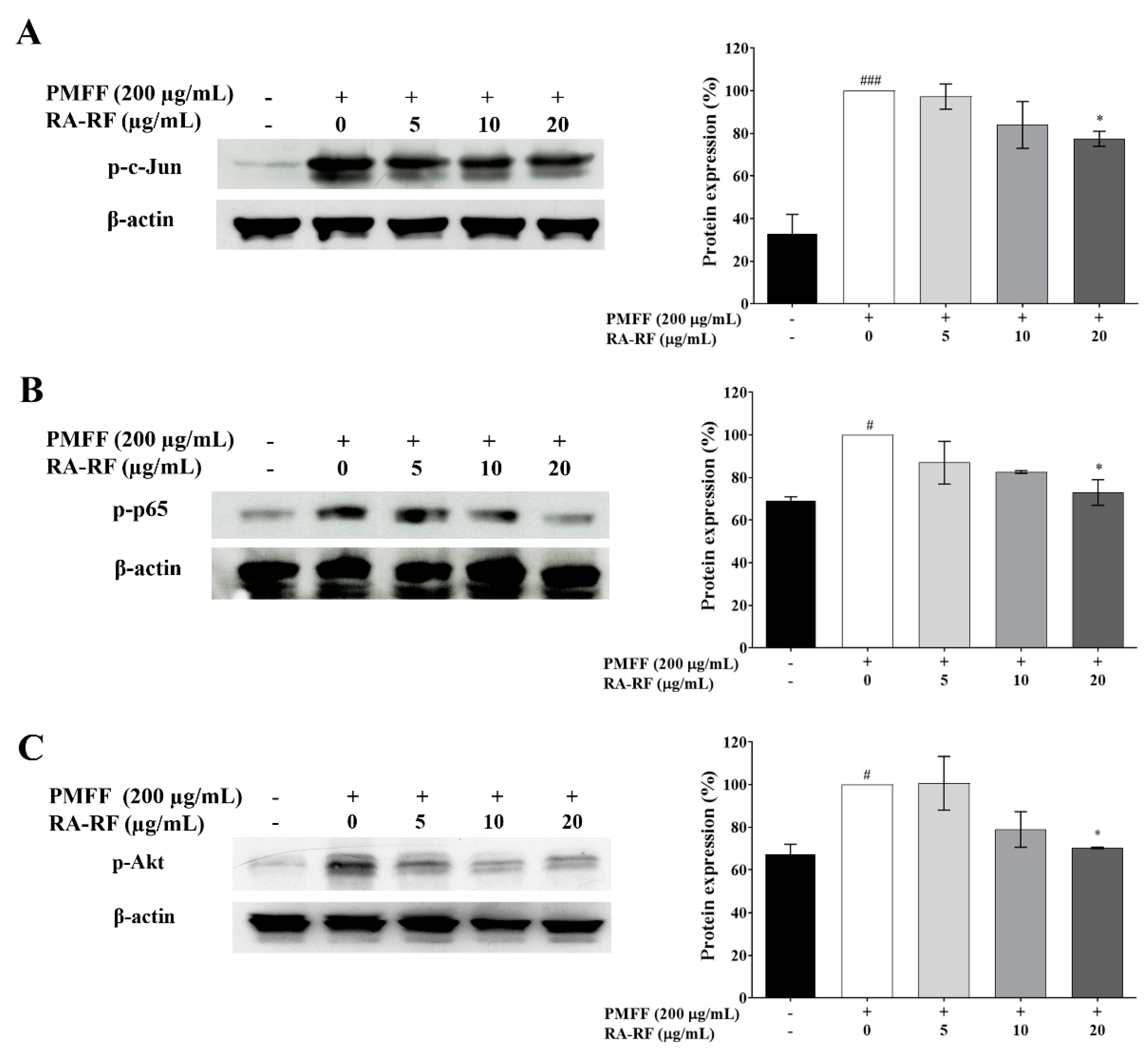
3.7. RA-RF Diminishes AP1, NF-κB and Akt Signaling Pathways
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mueller, W.; Loh, M.; Vardoulakis, S.; Johnston, H.J.; Steinle, S.; Precha, N.; Kliengchuay, W.; Tantrakarnapa, K.; Cherrie, J.W. Ambient particulate matter and biomass burning: An ecological time series study of respiratory and cardiovascular hospital visits in northern Thailand. Environ. Health 2020, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Phairuang, W.; Hata, M.; Furuuchi, M. Influence of agricultural activities, forest fires and agro-industries on air quality in Thailand. J. Environ. Sci. 2017, 52, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Farina, F.; Sancini, G.; Battaglia, C.; Tinaglia, V.; Mantecca, P.; Camatini, M.; Palestini, P. Milano summer particulate matter (PM10) triggers lung inflammation and extra pulmonary adverse events in mice. PLoS ONE 2013, 8, e56636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.; Kabir, E.; Kabir, S. A review on the human health impact of airborne particulate matter. Environ. Int. 2015, 74, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Jakovljević, I.; Pehnec, G.; Vađić, V.; Čačković, M.; Tomašić, V.; Jelinić, J.D. Polycyclic aromatic hydrocarbons in PM10, PM2.5 and PM1 particle fractions in an urban area. Air Qual. Atmos. Health 2018, 11, 843–854. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, K.; Loridas, S. Pulmonary oxidative stress, inflammation and cancer: Respirable particulate matter, fibrous dusts and ozone as major causes of lung carcinogenesis through reactive oxygen species mechanisms. J. Environ. Res. Public Health 2013, 10, 3886. [Google Scholar] [CrossRef] [PubMed]
- Freitas, M.; Alves, V.; Sarmento-Ribeiro, A.; Mota-Pinto, A. Polycyclic aromatic hydrocarbons may contibute for prostate cancer progression. J. Cancer Ther. 2013, 4, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Yang, T.; Gowrisankar, Y.V.; Liao, C.; Liao, J.; Huang, P.; Hseu, Y. Suppression of LPS-induced inflammation by chalcone flavokawain a through activation of Nrf2/ARE-mediated antioxidant genes and inhibition of ROS/NFκB signaling pathways in primary splenocytes. Oxid. Med. Cell. Longev. 2020, 3476212. [Google Scholar] [CrossRef]
- Kim, A.; Im, M.; Yim, N.; Jung, Y.P.; Ma, J.Y. Aqueous extract of bambusae caulis in taeniam inhibits PMA-induced tumor cell invasion and pulmonary metastasis: Suppression of NF-κB activation through ROS Signaling. PLoS ONE 2013, 8, e78061. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Jayawardena, T.U.; Kim, S.-Y.; Lee, H.G.; Je, J.-G.; Jee, Y.; Jeon, Y.-J. Sargassum horneri (Turner) inhibit urban particulate matter-induced inflammation in MH-S lung macrophages via blocking TLRs mediated NF-κB and MAPK activation. J. Ethnopharmacol. 2020, 249, 112363. [Google Scholar] [CrossRef]
- Wang, J.; Huang, J.; Wang, L.; Chen, C.; Yang, D.; Jin, M.; Bai, C.; Song, Y. Urban particulate matter triggers lung inflammation via the ROS-MAPK-NF-κB signaling pathway. J. Thorac. Dis. 2017, 9, 4398–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.; Lu, J.; Wang, B.; Sun, L.; Zhu, B.; Zhou, F.; Ding, Z. Inhibition of inflammation-induced injury and cell migration by coelonin and militarine in PM2.5-exposed human lung alveolar epithelial A549 cells. Eur. J. Pharmacol. 2021, 896, 173931. [Google Scholar] [CrossRef]
- Yue, H.; Yun, Y.; Gao, R.; Li, G.; Sang, N. Winter polycyclic aromatic hydrocarbon-bound particulate matter from peri-urban north china promotes lung cancer cell metastasis. Environ. Sci. Technol. 2015, 49, 14484–14493. [Google Scholar] [CrossRef]
- Vo, T.T.T.; Lee, C.; Wu, C.; Liu, J.; Lin, W.; Chen, Y.; Hsu, L.; Tsai11, M.; Lee, I. Surfactin from Bacillus subtilis attenuates ambient air particulate matter-promoted human oral cancer cells metastatic potential. J. Cancer 2020, 11, 6038–6049. [Google Scholar] [CrossRef]
- Seok, J.K.; Lee, J.; Kim, Y.M.; Boo, Y.C. Punicalagin and (–)-Epigallocatechin-3-Gallate rescue cell viability and attenuate inflammatory responses of human epidermal keratinocytes exposed to airborne particulate matter PM10. Skin Pharmacol. Physiol. 2018, 31, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C. Can plant phenolic compounds protect the skin from airborne particulate matter? Antioxidants 2019, 8, 379. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, H.M. Ethnomedicinal, phytochemical and pharmacological investigations of Perilla frutescens (L.) Britt. Molecules 2019, 24, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suttajit, M.; Khanaree, C.; Tantipaiboonwong, P.; Pintha, K. Omega-3, omega-6 fatty acids and nutrients of Nga-mon seeds in northern Thailand. Naresuan Phayao J. 2015, 8, 80–86. [Google Scholar]
- Pintha, K.; Tantipaiboonwong, P.; Yodkeeree, S.; Chaiwangyen, W.; Chumphukam, O.; Khantamat, O.; Khanaree, C.; Kangwan, N.; Thongchuai, B.; Suttajit, M.; et al. Thai perilla (Perilla frutescens) leaf extract inhibits human breast cancer invasion and migration. Maejo Int. J. Sci. 2018, 12, 112–123. [Google Scholar]
- Kangwan, N.; Pintha, K.; Lekawanvijit, S.; Suttajit, M. Rosmarinic acid enriched fraction from Perilla frutescens leaves strongly protects indomethacin-induced gastric ulcer in rats. BioMed Res. Int. 2019, 2019, 9514703. [Google Scholar] [CrossRef] [Green Version]
- Khanaree, C.; Pintha, K.; Tantipaiboonwong, P.; Suttajit, M.; Chewonarin, T. The effect of Perilla frutescens leaf on 1, 2-dimethylhydrazine-induced initiation of colon carcinogenesis in rats. J. Food Biochem. 2018, 42, e12493. [Google Scholar] [CrossRef]
- Chumphukam, O.; Pintha, K.; Khanaree, C.; Chewonarin, T.; Chaiwangyen, W.; Tantipaiboonwong, P.; Suttajit, M.; Khantamat, O. Potential anti-mutagenicity, antioxidant, and anti-inflammatory capacities of the extract from perilla seed meal. J. Food Biochem. 2018, 42, e12556. [Google Scholar] [CrossRef]
- Shekarchi, M.; Hajimehdipoor, H.; Saeidnia, S.; Gohari, A.R.; Hamedani, M.P. Comparative study of rosmarinic acid content in some plants of Labiatae family. Pharmacogn. Mag. 2012, 8, 37–41. [Google Scholar]
- Kaewnarin, K.; Shank, L.; Niamsup, H.; Rakariyatham, N. Inhibitory effects of Lamiaceae plants on the formation of advanced glycation endproducts (AGEs) in model proteins. J. Med. Biol. Eng. 2013, 2, 224–227. [Google Scholar] [CrossRef]
- Sik, B.; Hanczné, E.L.; Kapcsándi, V.; Ajtony, Z. Conventional and nonconventional extraction techniques for optimal extraction processes of rosmarinic acid from six Lamiaceae plants as determined by HPLC-DAD measurement. J. Pharm. Biomed. Anal. 2020, 184, 113173. [Google Scholar] [CrossRef]
- Yabueng, N.; Wiriya, W.; Chantara, S. Influence of zero-burning policy and climate phenomena on ambient PM2.5 patterns and PAHs inhalation cancer risk during episodes of smoke haze in Northern Thailand. Atmos. Environ. 2020, 232, 117485. [Google Scholar] [CrossRef]
- Kim, E.J.; Kim, G.T.; Kim, B.M.; Lim, E.G.; Kim, S.-Y.; Kim, Y.M. Apoptosis-induced effects of extract from Artemisia annua Linné by modulating PTEN/p53/PDK1/Akt/ signal pathways through PTEN/p53-independent manner in HCT116 colon cancer cells. BMC Complement. Altern. Med. 2017, 17, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chaiwangyen, W.; Ospina-Prieto, S.; Photini, S.M.; Schleussner, E.; Markert, U.R.; Morales-Prieto, D.M. Dissimilar microRNA-21 functions and targets in trophoblastic cell lines of different origin. Int. J. Biochem. Cell. Biol. 2015, 68, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Ooppachai, C.; Limtrakul, P.; Yodkeeree, S. Dicentrine potentiates TNF-α-induced apoptosis and suppresses invasion of A549 lung adenocarcinoma cells via modulation of NF-κB and AP-1 activation. Molecules 2019, 24, 4100. [Google Scholar] [CrossRef] [Green Version]
- Kasala, E.R.; Bodduluru, L.N.; Barua, C.C.; Sriram, C.S.; Gogoi, R. Benzo(a)pyrene induced lung cancer: Role of dietary phytochemicals in chemoprevention. Pharmacol. Rep. 2015, 67, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Jahan, S.A.; Kabir, E.; Brown, R.J. A review of airborne polycyclic aromatic hydrocarbons (PAHs) and their human health effects. Environ. Int. 2013, 60, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Luo, Y.; Zhong, R.; Law, P.T.Y.; Boon, S.S.; Chen, Z.; Wong, C.H.; Chan, P.K.S. Role of polycyclic aromatic hydrocarbons as a co-factor in human papillomavirus-mediated carcinogenesis. BMC Cancer 2019, 19, 138. [Google Scholar] [CrossRef]
- Alagawany, M.; Abd El-Hack, M.E.; Farag, M.R.; Gopi, M.; Karthik, K.; Malik, Y.S.; Dhama, K. Rosmarinic acid: Modes of action, medicinal values and health benefits. Anim. Health Res. Rev. 2017, 18, 167. [Google Scholar] [CrossRef]
- Yi, S.; Zhang, F.; Qu, F.; Ding, W. Water-insoluble fraction of airborne particulate matter (PM10) induces oxidative stress in human lung epithelial A549 cells. Environ. Toxicol. 2014, 29, 226–233. [Google Scholar] [CrossRef]
- Yun, Y.; Gao, R.; Yue, H.; Li, G.; Zhu, N.; Sang, N. Synergistic effects of particulate matter (PM10) and SO2 on human non-small cell lung cancer A549 via ROS-mediated NF-κB activation. J. Environ. Sci. 2015, 31, 146–153. [Google Scholar] [CrossRef]
- Kim, K.E.; Cho, D.; Park, H.J. Air pollution and skin diseases: Adverse effects of airborne particulate matter on various skin diseases. Life Sci. 2016, 152, 126–134. [Google Scholar] [CrossRef]
- Lee, G.H.; Jin, S.W.; Kim, S.J.; Pham, T.H.; Choi, J.H.; Jeong, H.G. Tetrabromobisphenol A induces MMP-9 expression via NADPH oxidase and the activation of ROS, MAPK, and Akt pathways in human breast cancer MCF-7 Cells. Toxicol. Res. 2019, 35, 93–101. [Google Scholar] [CrossRef]
- Yan, Z.; Jin, Y.; An, Z.; Liu, Y.; Samet, J.M.; Wu, W. Inflammatory cell signaling following exposures to particulate matter and ozone. Biochim. Biophys. Acta 2016, 1860, 2826–2834. [Google Scholar] [CrossRef] [PubMed]
- Radan, M.; Dianat, M.; Badavi, M.; Mard, S.A.; Bayati, V.; Goudarzi, G. In vivo and in vitro evidence for the involvement of Nrf2-antioxidant response element signaling pathway in the inflammation and oxidative stress induced by particulate matter (PM10): The effective role of gallic acid. Free Radic. Res. 2019, 53, 210–225. [Google Scholar] [CrossRef]
- Li, W.; Liu, T.; Xiong, Y.; Lv, J.; Cui, X.; He, R. Diesel exhaust particle promotes tumor lung metastasis via the induction of BLT1-mediated neutrophilic lung inflammation. Cytokine 2018, 111, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Jaafari, J.; Naddafi, K.; Yunesian, M.; Nabizadeh, R.; Hassanvand, M.S.; Ghozikali, M.G.; Shamsollahi, H.R.; Nazmara, S.; Yaghmaeian, K. Characterization, risk assessment and potential source identification of PM10 in Tehran. Microchem. J. 2020, 154, 104533. [Google Scholar] [CrossRef]
- Kord Mostafapour, F.; Jaafari, J.; Gharibi, H.; Sepand, M.R.; Hoseini, M.; Balarak, D.; Sillanpää, M.; Javid, A.B. Characterizing of fine particulate matter (PM1) on the platforms and outdoor areas of underground and surface subway stations. Hum. Ecol. Risk Assess. 2018, 24, 1016–1029. [Google Scholar] [CrossRef]
- Pongpiachan, S.; Hattayanone, M.; Cao, J. Effect of agricultural waste burning season on PM2.5-bound polycyclic aromatic hydrocarbon (PAH) levels in Northern Thailand. Atmos. Pollut. Res. 2017, 8, 1069–1080. [Google Scholar] [CrossRef]
- Pooltawee, J.; Pimpunchat, B.; Junyapoon, S. Size distribution, characterization and risk assessment of particle-bound polycyclic aromatic hydrocarbons during haze periods in Phayao Province, northern Thailand. Air Qual. Atmos. Health 2017, 10, 1097–1112. [Google Scholar] [CrossRef]
- Wiriya, W.; Prapamontol, T.; Chantara, S. PM10-bound polycyclic aromatic hydrocarbons in Chiang Mai (Thailand): Seasonal variations, source identification, health risk assessment and their relationship to air-mass movement. Atmos. Res. 2013, 124, 109–122. [Google Scholar] [CrossRef]
- Pengchai, P.; Chantara, S.; Sopajaree, K.; Wangkarn, S.; Tengcharoenkul, U.; Rayanakorn, M. Seasonal variation, risk assessment and source estimation of PM 10 and PM10-bound PAHs in the ambient air of Chiang Mai and Lamphun, Thailand. Environ. Monit. Assess. 2008, 154, 197. [Google Scholar] [CrossRef]
- US Environmental Protection Agency. Provisional Guidance for Quantitative Risk Assessment of Polycyclic Aromatic Hydrocarbons; EPA-600/R-93/089; US Environmental Protection Agency: Research Triangle Park, NC, USA, 1993; Volume 600.
- Cecinato, A. Polynuclear aromatic hydrocarbons (PAH), benz (a) pyrene (BaPY) and nitrated-PAH (N-PAH) in suspended particulate matter: Proposal for revision of the Italian reference method. Ann. Chim. 1997, 87, 483–496. [Google Scholar]
- US Environmental Protection Agency. Guidelines for Carcinogen Risk Assessment. Risk Assessment Forum. Available online: https://www.epa.gov/risk/guidelines-carcinogen-risk-assessment (accessed on 20 July 2021).
- World Health Organization. Air Quality Guidelines for Europe; WHO Regional Office for Europe: Copenhagen, Denmark, 2000. [Google Scholar]
- Liao, C.; Chiang, K. Probabilistic risk assessment for personal exposure to carcinogenic polycyclic aromatic hydrocarbons in Taiwanese temples. Chemosphere 2006, 63, 1610–1619. [Google Scholar] [CrossRef] [PubMed]
- AIRC. Some non-heterocyclic polycyclic aromatic hydrocarbons and some related exposures. IARC Monogr. Eval. Carcinog. Risks Hum. 2010, 92, 1–853. [Google Scholar]
- Adetona, O.; Reinhardt, T.E.; Domitrovich, J.; Broyles, G.; Adetona, A.M.; Kleinman, M.T.; Ottmar, R.D.; Naeher, L.P. Review of the health effects of wildland fire smoke on wildland firefighters and the public. Inhal. Toxicol. 2016, 28, 95–139. [Google Scholar] [CrossRef]
- Eom, S.; Yim, D.; Moon, S.I.; Youn, J.; Kwon, H.; Oh, H.C.; Yang, J.J.; Park, S.K.; Yoo, K.; Kim, H.S.; et al. Polycyclic aromatic hydrocarbon-induced oxidative stress, antioxidant capacity, and the risk of lung cancer: A pilot nested case-control study. Anticancer Res. 2013, 33, 3089. [Google Scholar]
- Wickramasinghe, A.P.; Karunaratne, D.G.G.P.; Sivakanesan, R. PM10-bound polycyclic aromatic hydrocarbons: Biological indicators, lung cancer risk of realistic receptors and ‘source-exposure-effect relationship’ under different source scenarios. Chemosphere 2012, 87, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Tantoh, D.M.; Hsu, S.; Nfor, O.N.; Frank, C.L.; Lung, C.; Ho, C.; Chen, C.; Liaw, Y. Association between coarse particulate matter (PM10-2.5) and nasopharyngeal carcinoma among Taiwanese men. J. Investig. Med. 2020, 68, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danielsen, P.H.; Møller, P.; Jensen, K.A.; Sharma, A.K.; Wallin, H.; Bossi, R.; Autrup, H.; Mølhave, L.; Ravanat, J.-L.; Briedé, J.J.; et al. Oxidative stress, DNA damage, and inflammation induced by ambient air and wood smoke particulate matter in human A549 and THP-1 cell lines. Chem. Chem. Res. Toxicol. 2011, 24, 168–184. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A. review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Shang, Y.; Zhou, Q.; Wang, T.; Jiang, Y.; Zhong, Y.; Qian, G.; Zhu, T.; Qiu, X.; An, J. Airborne nitro-PAHs induce Nrf2/ARE defense system against oxidative stress and promote inflammatory process by activating PI3K/Akt pathway in A549 cells. Toxicol. Vitr. 2017, 44, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, U.S.; McWhinney, R.D.; Rastogi, N.; Abbatt, J.P.D.; Evans, G.J.; Scott, J.A. Cytotoxic and proinflammatory effects of ambient and source-related particulate matter (PM) in relation to the production of reactive oxygen species (ROS) and cytokine adsorption by particles. Inhal. Toxicol. 2010, 22, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Habib, G.; Vivekanandan, P.; Kumar, A. Reactive oxygen species production and inflammatory effects of ambient PM2.5 -associated metals on human lung epithelial A549 cells “one year-long study”: The Delhi chapter. Chemosphere 2021, 262, 128305. [Google Scholar] [CrossRef]
- Long, Y.; Yang, X.; Yang, Q.; Clermont, A.C.; Yin, Y.; Liu, G.; Hu, L.; Liu, Q.; Zhou, Q.; Liu, Q.S.; et al. PM2.5 induces vascular permeability increase through activating MAPK/ERK signaling pathway and ROS generation. J. Hazard. Mater. 2020, 386, 121659. [Google Scholar] [CrossRef]
- Chua, M.L.; Setyawati, M.I.; Li, H.; Fang, C.H.Y.; Gurusamy, S.; Teoh, F.T.L.; Leong, D.T.; George, S. Particulate matter from indoor environments of classroom induced higher cytotoxicity and leakiness in human microvascular endothelial cells in comparison with those collected from corridor. Indoor Air 2017, 27, 551–563. [Google Scholar] [CrossRef]
- Lu, Y.; Su, S.; Jin, W.; Wang, B.; Li, N.; Shen, H.; Li, W.; Huang, Y.; Chen, H.; Zhang, Y.; et al. Characteristics and cellular effects of ambient particulate matter from Beijing. Environ. Pollut. 2014, 191, 63–69. [Google Scholar] [CrossRef]
- Danielsen, P.H.; Loft, S.; Kocbach, A.; Schwarze, P.E.; Møller, P. Oxidative damage to DNA and repair induced by Norwegian wood smoke particles in human A549 and THP-1 cell lines. Mutat. Res. 2009, 674, 116–122. [Google Scholar] [CrossRef]
- Libalova, H.; Milcova, A.; Cervena, T.; Vrbova, K.; Rossnerova, A.; Novakova, Z.; Topinka, J.; Rossner, P. Kinetics of ROS generation induced by polycyclic aromatic hydrocarbons and organic extracts from ambient air particulate matter in model human lung cell lines. Mutat Res. Genet. Toxicol. Environ. Mutagen. 2018, 827, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Tipsuwan, W.; Chaiwangyen, W. Preventive effects of polyphenol-rich perilla leaves on oxidative stress and haemolysis. Sci. Asia 2018, 44, 162–169. [Google Scholar] [CrossRef] [Green Version]
- Qiao, S.; Li, W.; Tsubouchi, R.; Haneda, M.; Murakami, K.; Takeuchi, F.; Nisimoto, Y.; Yoshino, M. Rosmarinic acid inhibits the formation of reactive oxygen and nitrogen species in RAW264.7 macrophages. Free Radic. Res. 2005, 39, 995–1003. [Google Scholar] [CrossRef]
- Liang, Z.; Wu, L.; Deng, X.; Liang, Q.; Xu, Y.; Deng, R.; Lv, L.; Ji, M.; Hao, Z.; He, J. The antioxidant rosmarinic acid ameliorates oxidative lung damage in experimental allergic asthma via modulation of NADPH oxidases and antioxidant enzymes. Inflammation 2020, 43, 1902–1912. [Google Scholar] [CrossRef]
- Erlandsson, L.; Lindgren, R.; Nääv, Å.; Krais, A.M.; Strandberg, B.; Lundh, T.; Boman, C.; Isaxon, C.; Hansson, S.R.; Malmqvist, E. Exposure to wood smoke particles leads to inflammation, disrupted proliferation and damage to cellular structures in a human first trimester trophoblast cell line. Environ. Pollut. 2020, 264, 114790. [Google Scholar] [CrossRef]
- Young, T.M.; Black, G.P.; Wong, L.; Bloszies, C.S.; Fiehn, O.; He, G.; Denison, M.S.; Vogel, C.F.A.; Durbin-Johnson, B. Identifying toxicologically significant compounds in urban wildfire ash using in vitro bioassays and high-resolution mass spectrometry. Environ. Sci. Technol. 2021, 55, 3657–3667. [Google Scholar] [CrossRef]
- Williams, K.M.; Franzi, L.M.; Last, J.A. Cell-specific oxidative stress and cytotoxicity after wildfire coarse particulate matter instillation into mouse lung. Toxicol. Appl. Pharmacol. 2013, 266, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Phromnoi, K.; Suttajit, M.; Saenjum, C.; Limtrakul, P.D. Inhibitory effect of a rosmarinic acid-enriched fraction prepared from Nga-Mon (Perilla frutescens) seed meal on osteoclastogenesis through the RANK signaling pathway. Antioxidants 2021, 10, 307. [Google Scholar] [CrossRef]
- Wang, G.; Chen, S.; Chen, Y.; Hong, C.; Hsu, Y.; Yen, G. Protective effect of rosmarinic acid-rich trichodesma khasianum clarke leaves against ethanol-induced gastric mucosal injury in vitro and in vivo. Phytomedicine 2021, 80, 153382. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Bastin, A.R.; Ghahremani, H.; Doustimotlagh, A.H. The effects of rosmarinic acid on oxidative stress parameters and inflammatory cytokines in lipopolysaccharide-induced peripheral blood mononuclear cells. Mol. Biol. Rep. 2020, 47, 3557–3566. [Google Scholar] [CrossRef] [PubMed]
- Lembo, S.; Balato, A.; Di Caprio, R.; Cirillo, T.; Giannini, V.; Gasparri, F.; Monfrecola, G. The modulatory effect of ellagic acid and rosmarinic acid on Ultraviolet-B-induced cytokine/chemokine gene expression in skin keratinocyte (HaCaT) cells. Biomed. Res. Int. 2014, 2014, 346793. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Chen, J.; Hu, Y.; Lu, W.; Zhang, X.; Wang, R.; Chu, K. Rosmarinic acid mitigates lipopolysaccharide-induced neuroinflammatory responses through the inhibition of TLR4 and CD14 expression and NF-κB and NLRP3 inflammasome activation. Inflammation 2018, 41, 732–740. [Google Scholar] [CrossRef]
- Rocha, J.; Eduardo-Figueira, M.; Barateiro, A.; Fernandes, A.; Brites, D.; Bronze, R.; Duarte, C.M.M.; Serra, A.T.; Pinto, R.; Freitas, M.; et al. Anti-inflammatory effect of rosmarinic acid and an extract of rosmarinus officinalis in rat models of local and systemic inflammation. Basic Clin. Pharmacol. Toxicol. 2015, 116, 398–413. [Google Scholar] [CrossRef]
- Domitrović, R.; Škoda, M.; Vasiljev Marchesi, V.; Cvijanović, O.; Pernjak Pugel, E.; Štefan, M.B. Rosmarinic acid ameliorates acute liver damage and fibrogenesis in carbon tetrachloride-intoxicated mice. Food Chem. Toxicol. 2013, 51, 370–378. [Google Scholar] [CrossRef]
- Morales-Bárcenas, R.; Chirino, Y.I.; Sánchez-Pérez, Y.; Osornio-Vargas, Á.R.; Melendez-Zajgla, J.; Rosas, I.; García-Cuellar, C.M. Particulate matter (PM10) induces metalloprotease activity and invasion in airway epithelial cells. Toxicol. Lett. 2015, 237, 167–173. [Google Scholar] [CrossRef]
- Han, Y.; Kee, J.; Hong, S. Rosmarinic acid activates ampk to inhibit metastasis of colorectal cancer. Front. Pharmacol. 2018, 9. [Google Scholar] [CrossRef]
- Han, Y.; Ma, L.; Zhao, L.; Feng, W.; Zheng, X. Rosmarinic inhibits cell proliferation, invasion and migration via up-regulating miR-506 and suppressing MMP2/16 expression in pancreatic cancer. Biomed. Pharmacother. 2019, 115, 108878. [Google Scholar] [CrossRef]
- Chen, X.; Su, H.Z.Z. Detailed studies on the anticancer action of rosmarinic acid in human Hep-G2 liver carcinoma cells: Evaluating its effects on cellular apoptosis, caspase activation and suppression of cell migration and invasion. J. BUON 2020, 25, 2011–2016. [Google Scholar] [PubMed]
- Liu, Y.; Xu, X.; Tang, H.; Pan, Y.; Hu, B.; Huang, G. Rosmarinic acid inhibits cell proliferation, migration, and invasion and induces apoptosis in human glioma cells. Int. J. Mol. Med. 2021, 47, 67. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.L.; Wang, B.; Chen, H.; Ho, K.F.; Cao, J.; Hai, G.; Jalaludin, B.; Herbert, C.; Thomas, P.S.; Saad, S.; et al. Pulmonary inflammation induced by low-dose particulate matter exposure in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 317, L424–L430. [Google Scholar] [CrossRef] [PubMed]
- Bejjani, F.; Evanno, E.; Zibara, K.; Piechaczyk, M.; Jariel-Encontre, I. The AP-1 transcriptional complex: Local switch or remote command? Biochim. Biophys. Acta Rev. Bioenergy Cancer 2019, 1872, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lee, W.; Jayawardena, T.U.; Cha, S.; Jeon, Y. Dieckol, an algae-derived phenolic compound, suppresses airborne particulate matter-induced skin aging by inhibiting the expressions of pro-inflammatory cytokines and matrix metalloproteinases through regulating NF-κB, AP-1, and MAPKs signaling pathways. Food Chem. Toxicol. 2020, 146, 111823. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, M.; Kim, J.M.; Lee, M.K.; Seo, S.J.; Park, K.Y. Afzelin suppresses proinflammatory responses in particulate matter-exposed human keratinocytes. Int. J. Mol. Med. 2019, 43, 2516–2522. [Google Scholar] [CrossRef]
- Zu, Y.-Y.; Liu, Q.-F.; Tian, S.-X.; Jin, L.-X.; Jiang, F.-S.; Li, M.-Y.; Zhu, B.-Q.; Ding, Z.-S. Effective fraction of Bletilla striata reduces the inflammatory cytokine production induced by water and organic extracts of airborne fine particulate matter (PM2.5) in vitro. BMC Complement. Altern. Med. 2019, 19, 369. [Google Scholar] [CrossRef]
- Jarvis, I.W.H.; Bergvall, C.; Morales, D.A.; Kummrow, F.; Umbuzeiro, G.A.; Westerholm, R.; Stenius, U.; Dreij, K. Nanomolar levels of PAHs in extracts from urban air induce MAPK signaling in HepG2 cells. Toxicol. Lett. 2014, 229, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Luo, M.; He, L.; Cao, Y.; Li, W.; Ying, S.; Chen, Z.; Shen, H. Necroptosis contributes to urban particulate matter-induced airway epithelial injury. Cell. Physiol. Biochem. 2018, 46, 699–712. [Google Scholar] [CrossRef]
- Bae, J.; Han, M.; Shin, H.S.; Kim, M.; Shin, C.; Lee, D.H.; Chung, J.H. Perilla frutescens leaves extract ameliorates ultraviolet radiation-induced extracellular matrix damage in human dermal fibroblasts and hairless mice skin. J. Ethnopharmacol. 2017, 195, 334–342. [Google Scholar] [CrossRef]
- Lin, S.; Wang, Y.; Chen, W.; Liao, S.; Chou, S.; Yang, C.; Chen, C. Hepatoprotective activities of rosmarinic acid against extrahepatic cholestasis in rats. Food Chem. Toxicol. 2017, 108, 214–223. [Google Scholar] [CrossRef]
- Yang, K.; Shen, Z.; Zou, Y.; Gao, K. Rosmarinic acid inhibits migration, invasion, and p38/AP-1 signaling via miR-1225-5p in colorectal cancer cells. J. Recept. Signal. Transduct. Res. 2021, 41, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-kB in development and progression of human cancer. Virchows Archiv 2005, 446, 475–482. [Google Scholar] [CrossRef] [PubMed]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-κB and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Liu, J.; Yin, J.; Cui, J.; Xiao, D.; Zhuo, W.; Luo, C.; Liu, R.; Li, X.; Zhang, W.; et al. Resistin facilitates metastasis of lung adenocarcinoma through the TLR4/Src/EGFR/PI3K/NF-κB pathway. Cancer Sci. 2018, 109, 2391–2400. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Cho, C.; Hsieh, W.; Tsai, C.; Chen, C.; Chang, H.; Lin, C. In vitro and in vivo experimental studies of PM2.5 on disease progression. Int. J. Environ. Res. Public Health 2018, 15, 1380. [Google Scholar] [CrossRef] [Green Version]
- Haghani, A.; Johnson, R.; Safi, N.; Zhang, H.; Thorwald, M.; Mousavi, A.; Woodward, N.C.; Shirmohammadi, F.; Coussa, V.; Wise, J.P.; et al. Toxicity of urban air pollution particulate matter in developing and adult mouse brain: Comparison of total and filter-eluted nanoparticles. Environ. Int. 2020, 136, 105510. [Google Scholar] [CrossRef]
- Taş, İ.; Zhou, R.; Park, S.-Y.; Yang, Y.; Gamage, C.D.B.; Son, Y.-J.; Paik, M.-J.; Kim, H. Inflammatory and tumorigenic effects of environmental pollutants found in particulate matter on lung epithelial cells. Toxicol. Vitr. 2019, 59, 300–311. [Google Scholar] [CrossRef]
- Joardar, S.; Dewanjee, S.; Bhowmick, S.; Dua, T.K.; Das, S.; Saha, A.; De Feo, V. Rosmarinic acid attenuates cadmium-induced nephrotoxicity via inhibition of oxidative stress, apoptosis, inflammation and fibrosis. Int. J. Mol. Sci. 2019, 20, 2027. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Hu, C.; Wu, L.; Xu, L.; Jiang, W. Rosmarinic acid inhibits inflammation and angiogenesis of hepatocellular carcinoma by suppression of NF-κB signaling in H22 tumor-bearing mice. J. Pharmacol. Sci. 2016, 132, 131–137. [Google Scholar] [CrossRef] [Green Version]
- An, Y.; Zhao, J.; Zhang, Y.; Wu, W.; Hu, J.; Hao, H.; Qiao, Y.; Tao, Y.; An, L. Rosmarinic acid induces proliferation suppression of hepatoma cells associated with NF-κB signaling pathway.Asian Pac. J. Cancer Prev. 2020, 22, 1623–1632. [Google Scholar]
- Duan, M.; Zhou, H.; Wu, Q.; Liu, C.; Xiao, Y.; Deng, W.; Tang, Q. Andrographolide protects against hg-induced inflammation, apoptosis, migration, and impairment of angiogenesis via PI3K/AKT-eNOS Signalling in HUVECs. Mediators Inflamm. 2019, 2019, 6168340. [Google Scholar] [CrossRef]
- Song, L.; Li, D.; Li, X.; Ma, L.; Bai, X.; Wen, Z.; Zhang, X.; Chen, D.; Peng, L. Exposure to PM2.5 induces aberrant activation of NF-κB in human airway epithelial cells by downregulating miR-331 expression. Environ. Toxicol. Pharmacol. 2017, 50, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Luo, Q.; Yuan, X.; Chai, L.; Li, D.; Liu, J.; Lv, Z. Atmospheric particulate matter2.5 promotes the migration and invasion of hepatocellular carcinoma cells. Oncol. Lett. 2017, 13, 3445–3450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Chen, J.; Cai, G.-E.; Lu, W.; Xu, W.; Wang, R.; Lin, Y.; Yang, C. Rosmarinic acid regulates microglial M1/M2 polarization via the PDPK1/Akt/HIF pathway under conditions of neuroinflammation. Inflammation 2021, 44, 129–147. [Google Scholar] [CrossRef]
- Wang, L.; Yang, H.; Wang, C.; Shi, X.; Li, K. Rosmarinic acid inhibits proliferation and invasion of hepatocellular carcinoma cells SMMC 7721 via PI3K/AKT/mTOR signal pathway. Biomed. Pharmacother. 2019, 120, 109443. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, J.; Yang, Y.; Wang, X.; Chen, G.; Shi, A.; Lu, Y.; Jia, S.; Kang, X.; Lu, L. Rosmarinic acid exerts an anticancer effect on osteosarcoma cells by inhibiting DJ-1 via regulation of the PTEN-PI3K-Akt signaling pathway. Phytomedicine 2020, 68, 153186. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Xu, G.; Liu, L.; Xu, D.; Liu, J. Anti-invasion effect of rosmarinic acid via the extracellular signal-regulated kinase and oxidation–reduction pathway in Ls174-T cells. J. Cell. Biochem. 2010, 111, 370–379. [Google Scholar] [CrossRef]





| Gene | Sequence |
|---|---|
| TNF-α | Forward: 5′-CCC AGG CAG TCA GAT CAT CTT C-3′ |
| Reverse: 5′-AGC TGC CCC TCA GCT TGA-3′ | |
| IL-6 | Forward: 5′-ATG AAC TCC TTC TCC ACA AGC-3′ |
| Reverse: 5′-GTT TTC TGC CAG TGC CTC TTT G-3′ | |
| IL-8 | Forward: 5′-AGA TAT TGC ACG GGA GAA-3′ |
| Reverse: 5′-GAA ATA AAG GAG AAA CCA-3′ | |
| COX-2 | Forward: 5′-CCC TTG GGT GTC AAA GGT AA-3′ |
| Reverse: 5′-GCC CTC GCT TAT GAT CTG TC-3′ | |
| GAPDH | Forward: 5′-GAA GGT GAA GGT CGA GTC A-3′ |
| Reverse: 5′-GCT CCT GGA AGA TGG TGA T-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintha, K.; Chaiwangyen, W.; Yodkeeree, S.; Suttajit, M.; Tantipaiboonwong, P. Suppressive Effects of Rosmarinic Acid Rich Fraction from Perilla on Oxidative Stress, Inflammation and Metastasis Ability in A549 Cells Exposed to PM via C-Jun, P-65-Nf-Κb and Akt Signaling Pathways. Biomolecules 2021, 11, 1090. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11081090
Pintha K, Chaiwangyen W, Yodkeeree S, Suttajit M, Tantipaiboonwong P. Suppressive Effects of Rosmarinic Acid Rich Fraction from Perilla on Oxidative Stress, Inflammation and Metastasis Ability in A549 Cells Exposed to PM via C-Jun, P-65-Nf-Κb and Akt Signaling Pathways. Biomolecules. 2021; 11(8):1090. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11081090
Chicago/Turabian StylePintha, Komsak, Wittaya Chaiwangyen, Supachai Yodkeeree, Maitree Suttajit, and Payungsak Tantipaiboonwong. 2021. "Suppressive Effects of Rosmarinic Acid Rich Fraction from Perilla on Oxidative Stress, Inflammation and Metastasis Ability in A549 Cells Exposed to PM via C-Jun, P-65-Nf-Κb and Akt Signaling Pathways" Biomolecules 11, no. 8: 1090. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11081090