Characterization of a New Bifunctional and Cold-Adapted Polysaccharide Lyase (PL) Family 7 Alginate Lyase from Flavobacterium sp.

Abstract
:1. Introduction
2. Results and Discussions
2.1. Sequence Analysis of AlyS02
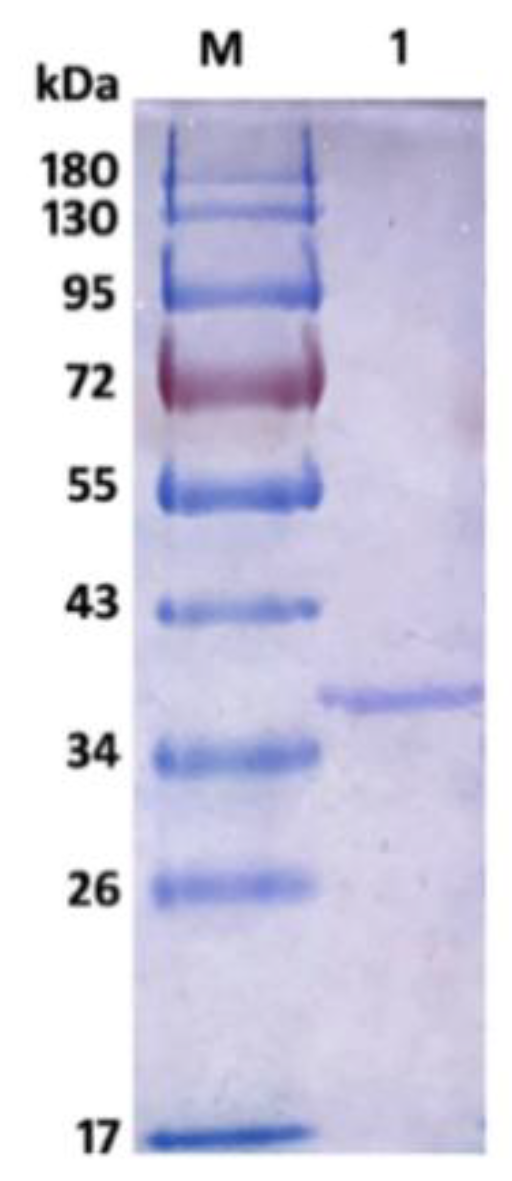
2.2. Expression and Purification of Recombinant AlyS02
2.3. Characterization of Recombinant AlyS02
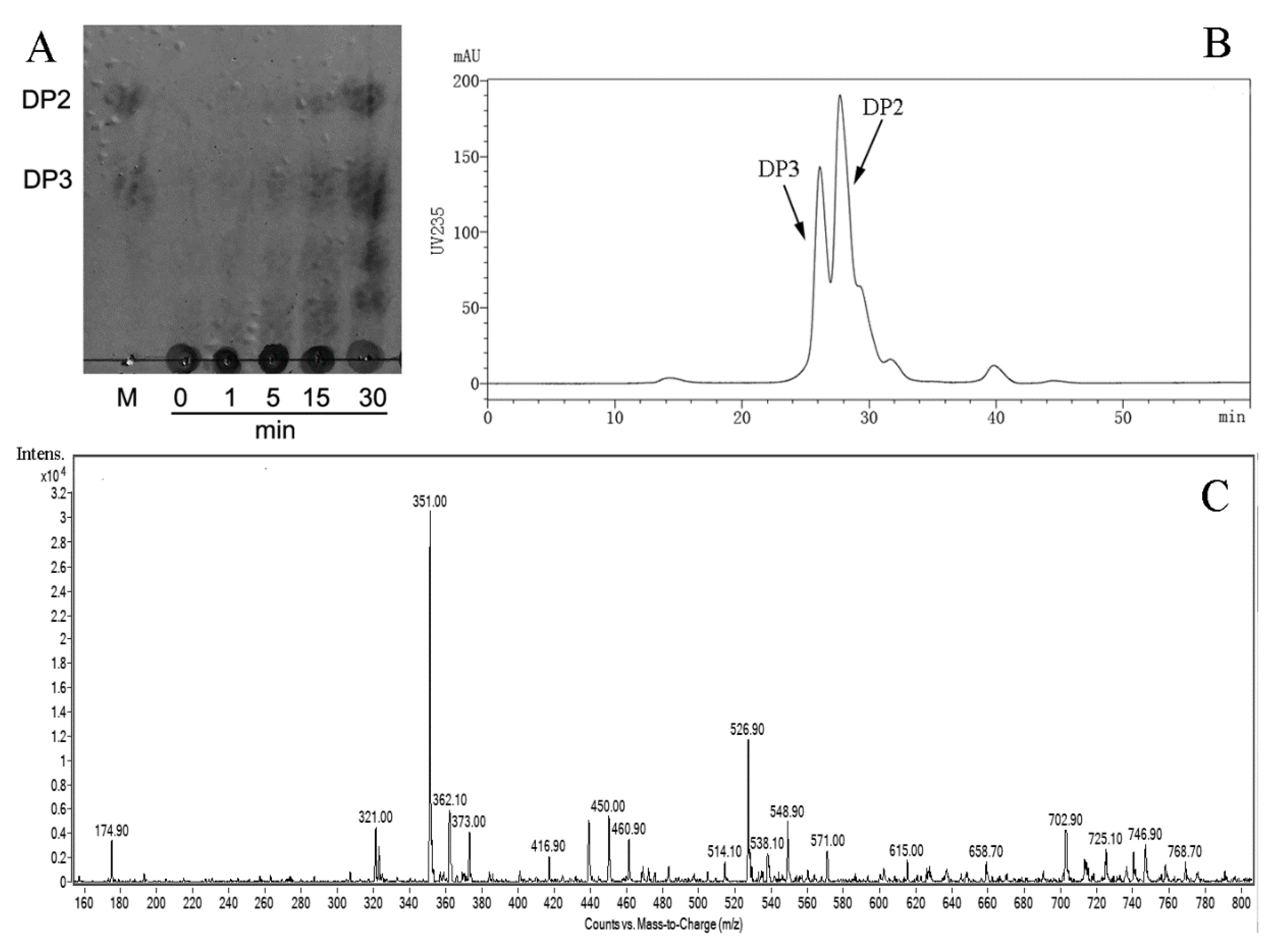
2.4. Substrate Specificity and Degradation Pattern of AlyS02
3. Materials and Methods
3.1. Materials, Strains and Culture Conditions
3.2. Sequence Analysis of AlyS02
3.3. Secretory Expression and Purification of Recombinant AlyS02
3.4. Enzymatic Activity Assay of the Alginate Lyase
3.5. Effects of Temperature and pH on Recombinant AlyS02 Activity and Stability
3.6. Effects of NaCl and Metal Ions on the Activity of Recombinant AlyS02
3.7. Substrate Specificity of Recombinant AlyS02
3.8. Degradation Products Analysis of Recombinant AlyS02
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wargacki, A.J.; Leonard, E.; Win, M.N.; Regitsky, D.D.; Santos, C.N.S.; Kim, P.B.; Cooper, S.R.; Raisner, R.M.; Herman, A.; Sivitz, A.B.; et al. An engineered Microbial platform for direct biofuel production from Brown Macroalgae. Science 2012, 335, 308–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ścieszka, S.; Klewicka, E. Algae in Food: A General Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.M.; Pereira, O.R.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, A.M.S. Seaweeds as preventive agents for cardiovascular diseases: From nutrients to functional foods. Mar. Drugs. 2015, 13, 6838–6865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanna, B.; Mishra, A. Nutraceutical potential of seaweed polysaccharides: Structure, bioactivity, safety, and toxicity. Compr. Rev. Food Sci. Food 2019, 18, 817–831. [Google Scholar] [CrossRef] [Green Version]
- Gacesa, P. Enzymic degradation of alginates. Int. J. Biochem. 1992, 24, 545–552. [Google Scholar] [CrossRef]
- Aliste, A.J.; Vieira, F.F.; Mastro, N.L.D. Radiation effects on agar, alginates and carrageenan to be used as food additives. Radiat. Phys. Chem. 2000, 57, 305–308. [Google Scholar] [CrossRef]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [Green Version]
- Mathieu, S.; Touvreyloiodice, M.; Poulet, L.; Drouillard, S.; Vincentelli, R.; Henrissat, B.; Braek, G.S.; Helbert, W. Ancient acquisition of “alginate utilization loci” by human gut microbiota. Sci. Rep. 2018, 8, 8075. [Google Scholar] [CrossRef] [Green Version]
- Laurienzo, P. Marine polysaccharides in pharmaceutical applications: An overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, Z.; Xu, X.; Jin, L. A Ca-alginate particle co-immobilized with Phanerochaete chrysosporium cells and the combined cross-linked enzyme aggregates from Trametes versicolor. Bioresour. Technol. 2015, 198, 464–469. [Google Scholar] [CrossRef]
- Zhu, B.; Chen, M.; Yin, H.; Du, Y.; Ning, L. Enzymatic hydrolysis of alginate to produce oligosaccharides by a new purified endo-type alginate lyase. Mar. Drugs 2016, 14, 108. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, L.; Liu, B.; He, N. Unsaturated alginate oligosaccharides attenuated obesity-related metabolic abnormalities by modulating gut microbiota in high-fat-diet mice. Food Funct. 2020, 11, 4773–4784. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Jiang, X.; Hwang, H.; Liu, S.; Guan, H. Antitumour activities of alginate-derived oligosaccharides and their sulphated substitution derivatives. Eur. J. Phycol. 2004, 39, 67–71. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kurachi, M.; Yamaguchi, K.; Oda. T. Induction of multiple cytokine secretion from RAW264.7 Cells by alginate oligosaccharides. Biosci. Biotechnol. Biochem. 2007, 71, 238–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Geng, M.; Xin, X.; Li, F.; Zhang, Z.; Li, J.; Ding, J. Multiple and multivalent interactions of novel anti-AIDS drug candidates, sulfated polymannuronate (SPMG)-derived oligosaccharides, with gp120 and their anti-HIV activities. Glycobiology 2004, 15. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, Y.; Xu, X.; Tamura, T.; Oda, T.; Muramatsu, T. Enzymatically depolymerized alginate oligomers that cause cytotoxic cytokine production in human mononuclear cells. Biosci. Biotechnol. Biochem. 2003, 67, 258–263. [Google Scholar] [CrossRef] [Green Version]
- Falkeborg, M.; Cheong, L.; Gianfico, C.; Sztukiel, K.M.; Kristensen, K.; Glasius, M.; Xu, X.; Guo, Z. Alginate oligosaccharides: Enzymatic preparation and antioxidant property evaluation. Food Chem. 2014, 164, 185–194. [Google Scholar] [CrossRef]
- Zhu, Y.; Wu, L.; Chen, Y.; Ni, H.; Xiao, A.; Cai, H. Characterization of an extracellular biofunctional alginate lyase from marine Microbulbifer sp. ALW1 and antioxidant activity of enzymatic hydrolysates. Microbiol. Res. 2016, 182, 49–58. [Google Scholar] [CrossRef]
- Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed polysaccharides and derived oligosaccharides stimulate defense responses and protection against pathogens in plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, H.; Zhao, X.; Wang, W.; Du, Y.; He, A.; Sun, K. The promoting effects of alginate oligosaccharides on root development in Oryza sativa L. mediated by auxin signaling. Carbohydr. Polym. 2014, 113, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Jiang, X.; Hwang, H.; Liu, S.; Guan, H. Promotive effects of alginate-derived oligosaccharide on maize seed germination. J. Appl. Phycol. 2004, 16, 73–76. [Google Scholar] [CrossRef]
- Wong, T.Y.; Preston, L.A.; Schiller, N.L. Alginate lyase: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 2000, 54, 289. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Lee, C.G.; Lee, E.Y. Alginate lyase: Structure, property, and application. Biotechnol. Bioprocess Eng. 2011, 16, 843–851. [Google Scholar] [CrossRef]
- Zhu, B.; Tan, H.; Qin, Y.; Xu, Q.; Du, Y.; Yin, H. Characterization of a new endo-type alginate lyase from Vibrio sp. W13. Int. J. Biol. Macromol. 2015, 75, 330–337. [Google Scholar] [CrossRef]
- Inoue, A.; Takadono, K.; Nishiyama, R.; Tajima, K.; Kobayashi, T.; Ojima, T. Characterization of an alginate lyase, FlAlyA, from Flavobacterium sp. strain UMI-01 and its expression in Escherichia coli. Mar. Drugs 2014, 12, 4693–4712. [Google Scholar] [CrossRef] [Green Version]
- Yagi, H.; Isobe, N.; Itabashi, N.; Fujise, A.; Ohshiro, T. Characterization of a long-lived alginate lyase derived from Shewanella species YH1. Mar. Drugs 2018, 16, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Tang, L.; Bao, M.; Liu, Z.; Yu, W.; Han, F. Functional characterization of carbohydrate-binding modules in a new alginate lyase, TsAly7B, from Thalassomonas sp. LD5. Mar. Drugs 2019, 18, 25. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, L.; Hao, J.; Xing, M.; Sun, J.; Sun, M. Purification and characterization of a new alginate lyase from marine bacterium Vibrio sp. SY08. Mar. Drugs 2017, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Park, H.H.; Kam, N.; Lee, E.Y.; Kim, H.S. Cloning and characterization of a novel oligoalginate Lyase from a newly isolated bacterium Sphingomonas sp. MJ-3. Mar. Biotechnol. 2012, 14, 189–202. [Google Scholar] [CrossRef]
- Jagtap, S.S.; Hehemann, J.H.; Polz, M.F.; Lee, J.K.; Zhao, H. Comparative biochemical characterization of three exolytic oligoalginate lyases from Vibrio splendidus reveals complementary substrate scope, temperature, and pH adaptations. Appl. Environ. Microbiol. 2014, 80, 4207–4214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Zhou, J.; Li, X.; Peng, Q.; Lu, H.; Du, Y. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. J. Ind. Microbiol. Biotechnol. 2013, 40, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Lee, E.Y.; Kim, H.S. Cloning and characterization of alginate lyase from a marine bacterium Streptomyces sp. ALG-5. Mar. Biotechnol. 2009, 11, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.-W.; Tan, H.-D.; Qin, Y.-Q.; Du, Y.-G.; Yin, H. Characterization of a new endo-type polyM-specific alginate lyase from Pseudomonas sp. Biotechnol. Lett. 2015, 37, 409–415. [Google Scholar] [CrossRef]
- Chen, P.; Zhu, Y.; Men, Y.; Zeng, Y.; Sun, Y. Purification and characterization of a novel alginate lyase from the marine bacterium Bacillus sp. Alg07. Mar. Drugs 2018, 16, 86. [Google Scholar] [CrossRef] [Green Version]
- Li, J.-W.; Dong, S.; Song, J.; Li, C.-B.; Chen, X.-L.; Xie, B.B.; Zhang, Y.Z. Purification and characterization of a bifunctional alginate lyase from Pseudoalteromonas sp. SM0524. Mar. Drugs 2011, 9, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Dou, W.; Wei, D.; Li, H.; Li, H.; Rahman, M.M.; Shi, J.; Xu, Z.; Ma, Y. Purification and characterisation of a bifunctional alginate lyase from novel Isoptericola halotolerans CGMCC 5336. Carbohydr. Polym. 2013, 98, 1476–1482. [Google Scholar] [CrossRef]
- Yan, J.; Chen, P.; Zeng, Y.; Men, Y.; Sun, Y. The Characterization and modification of a novel bifunctional and robust alginate lyase derived from Marinimicrobium sp. H1. Mar. Drugs 2019, 17, 545. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, M.; Moriwaki, S.; Miyake, O.; Hashimoto, W.; Murata, K.; Mikami, B. Structure and function of a hypothetical Pseudomonas aeruginosa protein PA1167 classified into family PL-7 a novel alginate lyase with a β-sandwich fold. Int. J. Biol. Chem. 2004, 279, 31863–31872. [Google Scholar] [CrossRef] [Green Version]
- Belik, A.A.; Silchenko, A.S.; Kusaykin, M.I.; Zvyagintseva, T.N.; Ermakova, S.P. Alginate lyases: Substrates, structure, properties, and prospects of application. Russ. J. Bioorg. Chem. 2018, 44, 386–396. [Google Scholar] [CrossRef]
- Huang, G.; Wang, Q.; Lu, M.; Chao, X.; Fei, L.; Zhang, R.; Liao, W.; Huang, S. AlgM4: A new salt-activated alginate lyase of the PL7 family with endolytic activity. Mar. Drugs 2018, 16, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Ni, F.; Sun, Y.; Ning, L.; Yao, Z. Elucidation of degrading pattern and substrate recognition of a novel bifunctional alginate lyase from Flammeovirga sp. NJ-04 and its use for preparation alginate oligosaccharides. Biotechnol. Biofuels 2019, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, Z.; Wang, L.; Peng, J.; Wang, Y.; Han, Y.; Zhao, S. Combined enzymatic hydrolysis and selective fermentation for green production of alginate oligosaccharides from Laminaria japonica. Bioresour. Technol. 2019, 281, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Aarstad, O.A.; Tondervik, A.; Sletta, H.; Skjakbraek, G. Alginate sequencing: An analysis of block distribution in alginates using specific alginate degrading enzymes. Biomacromolecules 2012, 13, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Inoue, A.; Mashino, C.; Kodama, T.; Ojima, T. Protoplast preparation from Laminaria japonica with recombinant alginate lyase and cellulase. Mar. Biotechnol. 2011, 13, 256–263. [Google Scholar] [CrossRef]
- Lamppa, J.W.; Griswold, K.E. Alginate lyase exhibits catalysis-independent biofilm dispersion and antibiotic synergy. Antimicrob. Agents Chemother. 2013, 57, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Islan, G.A.; Martinez, Y.N.; Illanes, A.; Castro, G.R. Development of novel alginate lyase cross-linked aggregates for the oral treatment of cystic fibrosis. RSC Adv. 2014, 4, 11758. [Google Scholar] [CrossRef]
- Tang, J.C.; Taniguchi, H.; Chu, H.; Zhou, Q.; Nagata, S. Isolation and characterization of alginate degrading bacteria for disposal of seaweed wastes. Lett. Appl. Microbiol. 2008, 48, 38–43. [Google Scholar] [CrossRef]
- Singh, R.P.; Gupta, V.; Kumari, P.; Kumar, M.; Reddy, C.R.K.; Prasad, K.; Jha, B. Purification and partial characterization of an extracellular alginate lyase from Aspergillus oryzae isolated from brown seaweed. J. Appl. Phycol. 2011, 23, 755–762. [Google Scholar] [CrossRef]
- Hashimoto, W.; Miyake, O.; Ochiai, A.; Murata, K. Molecular identification of Sphingomonas sp. A1 alginate lyase (A1-IV′) as a member of novel polysaccharide lyase family 15 and implications in alginate lyase evolution. J. Biosci. Bioeng. 2005, 99, 48–54. [Google Scholar] [CrossRef]
- Zhu, B.; Ni, F.; Ning, L.; Sun, Y.; Yao, Z. Cloning and characterization of a new pH-stable alginate lyase with high salt tolerance from marine Vibrio sp. NJ-04. Int. J. Biol. Macromol. 2018, 115, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Suzuki, K.I.; Inoue, A.; Ojima, T. A novel oligoalginate lyase from abalone, Haliotis discus hannai, that releases disaccharide from alginate polymer in an exolytic manner. Carbohydr. Res. 2006, 341, 1809–1819. [Google Scholar] [CrossRef]
- Shiraiwa, Y.; Abe, K.; Sasaki, S.F.; Ikawa, T.; Nisizawa, K. Alginate lyase activities in the extracts from several brown algae. Bot. Mar. 2009, 18, 97–104. [Google Scholar] [CrossRef]
- Ogura, K.; Yamasaki, M.; Yamada, T.; Mikami, B.; Hashimoto, W.; Murata, K. Crystal structure of Family 14 polysaccharide lyase with pH-dependent modes of action. Int. J. Biol. Chem. 2009, 284, 35572–35579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, S.J.; Baik, K.S.; Park, S.C.; Choe, H.N.; Seong, C.N.; Shin, T.S.; Woo, H.C.; Cho, J.Y.; Kim, D. Characterization of alginate lyase gene using a metagenomic library constructed from the gut microflora of abalone. J. Ind. Microbiol. Biotechnol. 2012, 39, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; An, D.; Jiao, C.; Xiao, Q.; Weng, H.; Yang, Q.; Xiao, A. Cloning, expression, and characterization of a new pH- and heat-stable alginate lyase from Pseudoalteromonas carrageenovora ASY5. J. Food Biochem. 2019, 43, e12886. [Google Scholar] [CrossRef]
- Inoue, A.; Anraku, M.; Nakagawa, S.; Ojima, T. Discovery of a novel alginate lyase from Nitratiruptor sp. SB155-2 Thriving at deep-sea hydrothermal vents and identification of the residues responsible for its heat stability. Int. J. Biol. Chem. 2016, 291, 15551–15563. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-L.; Dong, S.; Xu, F.; Dong, F.; Li, P.-Y.; Zhang, X.Y.; Zhou, B.C.; Zhang, Y.Z.; Xie, B.B. Characterization of a new cold-adapted and salt-activated polysaccharide lyase family 7 alginate lyase from Pseudoalteromonas sp. SM0524. Front. Microbiol. 2016, 7, 1120. [Google Scholar] [CrossRef]
- Gao, S.; Zhang, Z.; Li, S.; Su, H.; Tang, L.; Tan, Y.; Yu, W.; Han, F. Characterization of a new endo-type polysaccharide lyase (PL) family 6 alginate lyase with cold-adapted and metal ions-resisted property. Int. J. Biol. Macromol. 2018, 120, 729–735. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Bi, X.; Ren, Y.; Han, Q.; Zhou, Y.; Han, Y.; Li, S. Characterization of an alkaline alginate lyase with pH-stable and thermo-tolerance property. Mar. Drugs 2019, 17, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Uchimura, K.; Miyazaki, M.; Nogi, Y.; Horikoshi, K. A new high-alkaline alginate lyase from a deep-sea bacterium Agarivorans sp. Extremophiles 2009, 13, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Jiang, X.; Hwang, H.-M. Purification and characterization of an alginate lyase from marine bacterium Vibrio sp. mutant strain 510-64. Curr. Microbiol. 2006, 53, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shao, Y.; Jiao, C.; Yang, Q.; Weng, H.; Xiao, A.F. Characterization and application of an alginate lyase, Aly1281 from marine bacterium Pseudoalteromonas carrageenovora ASY5. Mar. Drugs 2020, 18, 95. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, M.; Ogura, K.; Hashimoto, W.; Mikami, B.; Murata, K. A structural basis for depolymerization of alginate by polysaccharide lyase family-7. J. Mol. Biol. 2005, 352, 11–21. [Google Scholar] [CrossRef]
- Thomas, F.; Lundqvist, L.C.E.; Jam, M.; Jeudy, A.; Barbeyron, T.; Sandstrom, C.; Michel, G.; Czjzek, M. Comparative characterization of two marine alginate lyases from Zobellia galactanivorans reveals distinct modes of action and exquisite adaptation to their natural substrate. Int. J. Biol. Chem. 2013, 288, 23021–23037. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wang, S.; Zhang, Y.; Chen, L. High-level expression of a thermally stable alginate lyase using pichia pastoris, characterization and application in producing brown alginate oligosaccharide. Mar. Drugs 2018, 16, 158. [Google Scholar] [CrossRef] [Green Version]
- Madzak, C. Yarrowia lipolytica: Recent achievements in heterologous protein expression and pathway engineering. Appl. Microbiol. Biotechnol. 2015, 99, 4559–4577. [Google Scholar] [CrossRef]
- Iwamoto, Y.; Araki, R.; Iriyama, K.-I.; Oda, T.; Fukuda, H.; Hayashida, S.; Muramastu, T. Purification and characterization of bifunctional alginate lyase from Alteromonas sp. strain no. 272 and its action on saturated oligomeric substrates. Biosci. Biotechnol. Biochem. 2001, 65, 133–142. [Google Scholar] [CrossRef]
- Han, M.; Wang, X.; Ding, H.; Jin, M.; Yu, L.; Wang, J.; Yu, X. The role of N-glycosylation sites in the activity, stability, and expression of the recombinant elastase expressed by Pichia pastoris. Enzyme. Microb. Technol. 2014, 54, 32–37. [Google Scholar] [CrossRef]
- Zou, S.; Huang, S.; Kaleema, I.; Li, C. N-glycosylation enhances functional and structural stability of recombinant β-glucuronidase expressed in Pichia pastoris. J. Biotechnol. 2013, 164, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Kunjukunju, S.; Roy, A.; . Shekhar, S.; Kumta, P.N. Cross-linked enzyme aggregates of alginate lyase: A systematic engineered approach to controlled degradation of alginate hydrogel. Int. J. Biol. Macromol. 2018, 115, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, Q.; Lu, D.; Han, W.; Li, F. A novel bifunctional endolytic alginate lyase with variable alginate-degrading modes and versatile monosaccharide-producing properties. Front. Microbiol. 2018, 9, 167. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.T.; Lin, H.; Kim, S.M. Purification and characterization of a Na+/K+ dependent alginate lyase from turban shell gut Vibrio sp. YKW-34. Enzyme Microb. Technol. 2007, 41, 828–834. [Google Scholar] [CrossRef]
- Ochi, Y.; Takeuchi, T.; Murata, K.; Kawabata, Y.; Kusakabe, I. A simple method for preparation of poly-mannuronate using poly-guluronate lyase. Biosci. Biotechnol. Biochem. 1995, 59, 1560–1561. [Google Scholar] [CrossRef]
- Miyake, O.; Ochiai, A.; Hashimoto, W.; Murata, K. Origin and diversity of alginate lyases of families PL-5 and -7 in Sphingomonas sp. strain A1. J. Bacteriol. 2004, 186, 2891–2896. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Hu, Y.; Zhang, L.; Wang, Y.; Wang, S.; Zhang, Y.; Guo, H.; Ji, D.; Wang, Y. Alginate oligosaccharide DP5 exhibits antitumor effects in osteosarcoma patients following surgery. Front. Pharmacol. 2017, 8, 623. [Google Scholar] [CrossRef]
- Kawada, A.; Hiura, N.; Tajima, S.; Takahara, H. Alginate oligosaccharides stimulate VEGF-mediated growth and migration of human endothelial cells. Arch. Dermatol. Res. 1999, 291, 542–547. [Google Scholar] [CrossRef]
- Iwasaki, K.; Matsubara, Y. Purification of alginate oligosaccharides with root growth-promoting activity toward lettuce. Biosci. Biotechnol. Biochem. 2000, 64, 1067–1070. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wang, Z.P.; Sheng, J.; Zheng, Y.; Ji, X.F.; Zhou, H.X.; Liu, X.Y.; Chi, Z.M. High and efficient isomaltulose production using an engineered Yarrowia lipolytica strain. Bioresour. Technol. 2018, 265, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Purification Fold | Yield (%) |
|---|---|---|---|---|---|
| Crude enzyme | 43.14 | 3680 | 85.3 | 1 | 100 |
| Ultrafiltration 1st | 38.32 | 3520.5 | 91.87 | 1.08 | 95.67 |
| Ni-NTA sepharose | 4.86 | 2217.9 | 456.36 | 5.35 | 60.27 |
| Ultrafiltration 2nd | 4.72 | 2090.2 | 442.84 | 5.19 | 56.8 |
| Enzyme Name | Organism/Source | PL Family | Mw(kDa) | Optimal pH/Temperature (°C) | Cation Activators | Cation Inhibitors | Substrate Specificity | Main Products (DP) | References |
|---|---|---|---|---|---|---|---|---|---|
| AlyS02 | Flavobacterium sp. S02/Y. lipolytica | 7 | 36.5 | 7.6/30 | Na+, K+, Ca2+, Mg2+ | Fe3+, Al3+, Mn2+, Cu2+, Zn2+, , Li+, SDS, EDTA | PolyG, polyM | 2, 3 | This study |
| Alg2A | Flavobacterium sp. S20/E. coli | 7 | 33 | 8.5/45 | Na+, K+ | Ca2+, Mg2+, Co2+, Cu2+, Zn2+, Mn2+, Fe2+ | PolyG | 5–7 | [32] |
| FlAlyA | Flavabacterium sp. UMI-01/native | 7 | 30 | 7.7/55 | Na+, K+, Ca2+, Mg2+ | Co2+, Ni2+ | PolyM, polyMG | 2–5 | [26] |
| PA1167 | P. aeruginosa PAO1/E. coli | 7 | 25 | 8.5/40 | N. D. | N. D. | PolyMG | 2–4 | [39] |
| Aly1281 | P. carrageenovora ASY5/E. coli | 7 | 40.65 | 8.0/50 | Na+, K+ | N. D. | PolyG, polyM | 2 | [64] |
| AlgNJ-04 | Vibrio sp. NJ-04/E. coli | 7 | 50.19 | 7.0/40 | Na+, K+, Ca2+ | Fe2+, Cu2+, Zn2+ | PolyG, polyM | 2–5 | [51] |
| A9mT | Vibrio sp. JAM-A9m/E. coli | 7 | 28 | 7.5/30 | Na+, K+, Li+, Rb+, Cs+, , Tween 20, Nonidet P40 | Cu2+, Zn2+, Ni2+, Co2+, Sr2+, SDS | PolyM | N. D. | [74] |
| Algb | Vibrio sp. W13/E. coli | 7 | 54.12 | 8.0/30 | Na+, Ca2+, Co2+, Fe2+ | Cu2+, Zn2+, Mn2+, Ba2+ | PolyMG, polyG, polyM | 2–5 | [25] |
| Aly08 | Vibrio sp. SY01/E. coli | 7 | 35 | 8.35/45 | Na+, Ca2+, Mn2+, Co2+, Zn2+ | SDS, EDTA | PolyG | 2, 3 | [61] |
| Aly510–64 | Vibrio sp. 510–64/native | N. D. | 34.6 | 7.5/35 | Na+, K+, Ca2+, Mg2+, Li+ | N. D. | PolyG, polyMG | 3 | [63] |
| AlySY08 | Vibrio sp. SY08/native | N. D. | 33 | 7.6/40 | Na+, K+, Ca2+, Mg2+ | Fe3+, Al3+, Mn2+, Cu2+, Zn2+, SDS, EDTA, 2-mercaptoethanol | PolyG, polyM | 2 | [29] |
| AlgA | Bacillus sp. Alg07/native | N. D. | 60 | 7.5/40 | Na+, Ca2+, Mn2+, Mg2+, Co2+ | Fe3+, Fe2+, Cu2+, Al3+, Hg2+, Ba2+, EDTA | PolyM | 2–4 | [35] |
| Cel32 | Cellulophaga sp. NJ-1/native | N. D. | 32 | 8.0/50 | Na+, K+, Ca2+, Mg2+ | Fe2+, Cu2+, Zn2+, Co2+, Ni2+ | PolyMG, polyG, polyM | 2, 3 | [11] |
| rSAGL | Flavobacterium sp. H63/P. pastoris | 7 | 32 | 7.5/45 | Na+, K+, Mg2+ | Co2+, Cu2+, Zn2+, Mn2+, Ca2+, Ni2+, Fe3+, Ag+ | PolyM | 2–4 | [67] |
| rAlgSV1-PL7 | Shewanella sp. YH1/E. coli | 7 | 33.216 | 8.0/45 | Na+, K+, Mg2+ | Cu2+, Fe3+, N-bromosuccinimide | PolyG, polyM, polyMG | 1–4 | [27] |
| Aly2 | Flammeovirga sp. MY04/E. coli | 7 | 60.58 | 6.0/40 | EDTA, glycerol, 2-mercaptoethanol | Ag+, Hg2+, Pb2+, Fe3+, Zn2+, Ni2+, Fe2+, Cu2+, Cr3+, K+, Mn2+, SDS | PolyG, polyM | 2, 3 | [73] |
| TsAly6A | Thalassomonas sp. LD5/E. coli | 6 | 83.9 | 8.0/35 | Ca2+, Mg2+, Na+ | EDTA, SDS | PolyG, polyM | 2, 3 | [60] |
| aly-SJ02 | Pseudoalteromonas sp. SM0524/native | 18 | 32 | 8.5/50 | Na+, K+, Ba2+, Ca2+, Mg2+, Mn2+, Co2+, Sr2+, Ni2+ | EDTA | PolyG, polyM | 2–4 | [36] |
| A1-IV’ | Sphingomonas sp. A1/E. coli | 15 | 90 | 8.5/50 | N. D. | Cu2+, Zn2+, Hg2+, Co2+ | PolyM, polyMG | 2, 3 | [50] |
| AlgA | Pseudomonas sp. E03/E. coli | 5 | 40.4 | 8.0/30 | Na+, K+, Ca2+, Mg2+, Zn2+, Ba2+, PMSF, DTT | Co2+, Cu2+, Mn2+, Fe2+, Triton X-100, Tween 20, EDTA, SDS, urea | PolyM | 2–5 | [34] |
| Ion Mode | m/z |
|---|---|
| [ΔDP2–H]– | 351 |
| [ΔDP2–2H]2– | 175 |
| [ΔDP2–2H+Na]– | 373 |
| [ΔDP3–H]– | 527 |
| [ΔDP3–2H+Na]– | 549 |
| [ΔDP3–3H+2Na]– | 571 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.-X.; Xu, S.-S.; Yin, X.-J.; Wang, F.-L.; Li, Y. Characterization of a New Bifunctional and Cold-Adapted Polysaccharide Lyase (PL) Family 7 Alginate Lyase from Flavobacterium sp. Mar. Drugs 2020, 18, 388. https://0-doi-org.brum.beds.ac.uk/10.3390/md18080388
Zhou H-X, Xu S-S, Yin X-J, Wang F-L, Li Y. Characterization of a New Bifunctional and Cold-Adapted Polysaccharide Lyase (PL) Family 7 Alginate Lyase from Flavobacterium sp. Marine Drugs. 2020; 18(8):388. https://0-doi-org.brum.beds.ac.uk/10.3390/md18080388
Chicago/Turabian StyleZhou, Hai-Xiang, Shan-Shan Xu, Xue-Jing Yin, Feng-Long Wang, and Yang Li. 2020. "Characterization of a New Bifunctional and Cold-Adapted Polysaccharide Lyase (PL) Family 7 Alginate Lyase from Flavobacterium sp." Marine Drugs 18, no. 8: 388. https://0-doi-org.brum.beds.ac.uk/10.3390/md18080388